
Copepod Assemblages in A Large Arctic Coastal Area: A Baseline Summer Study



Abstract
:1. Introduction
2. Material and Methods
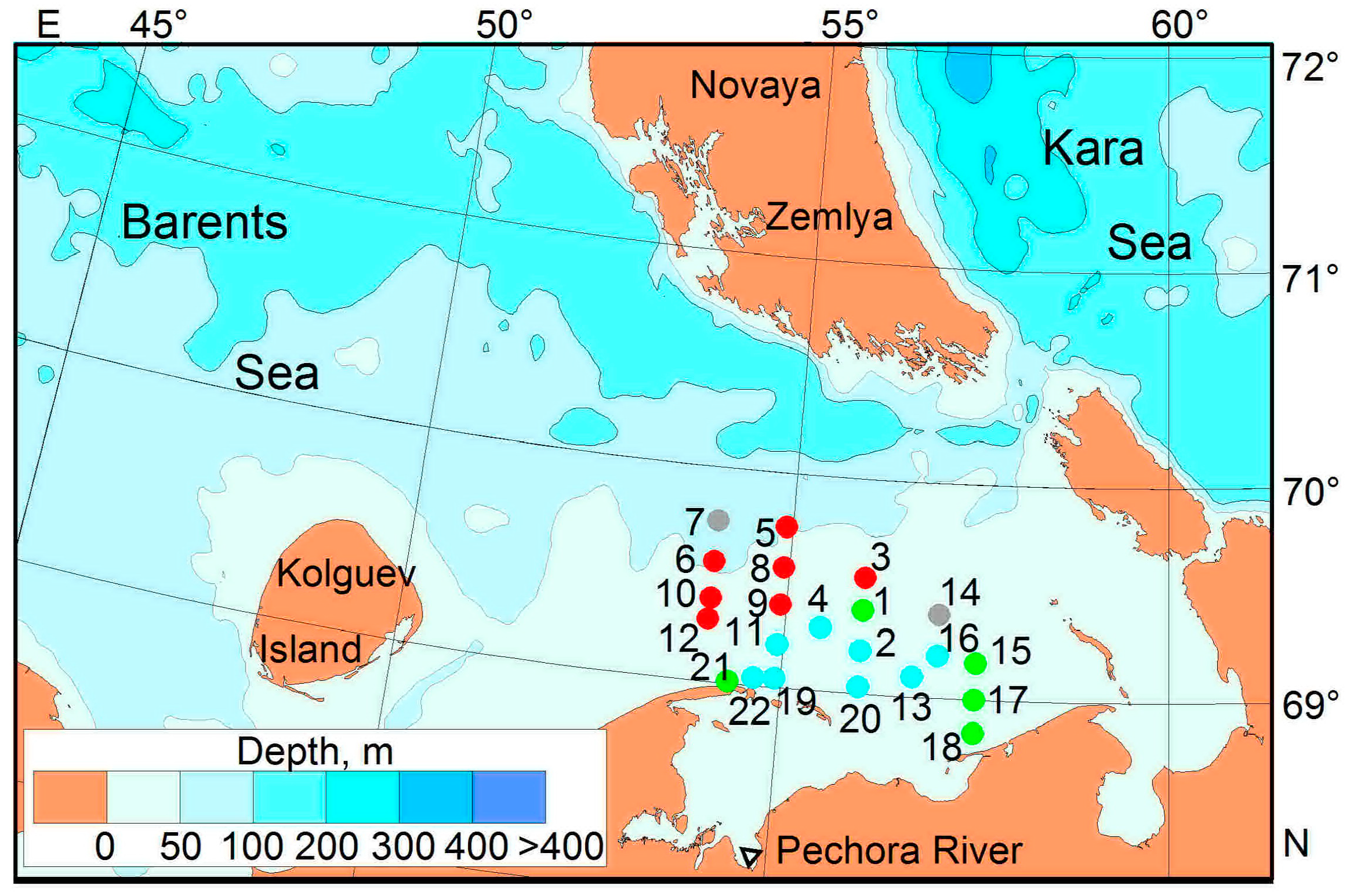
2.1. Sampling
2.2. Processing
2.3. Data Analysis
3. Results
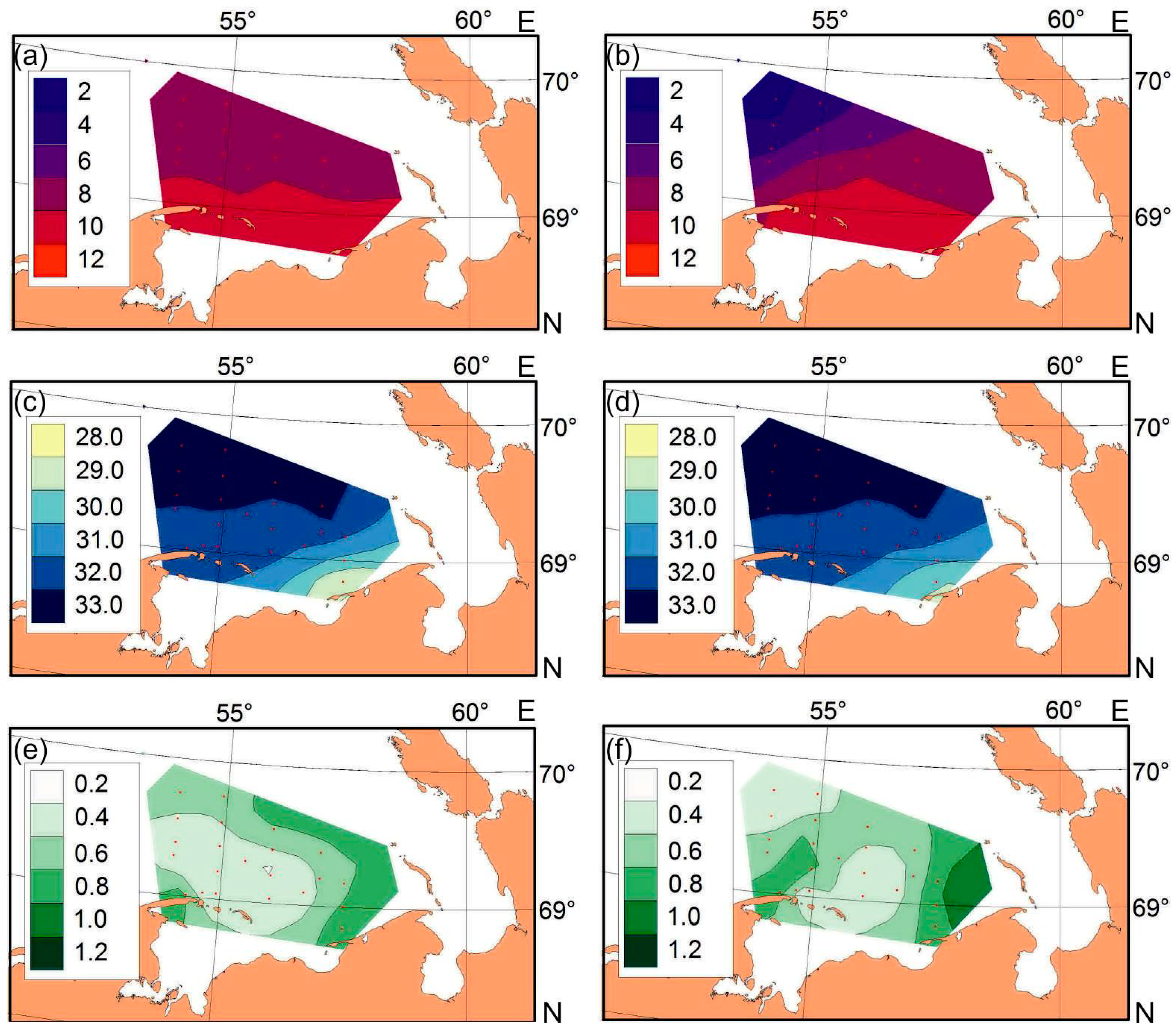
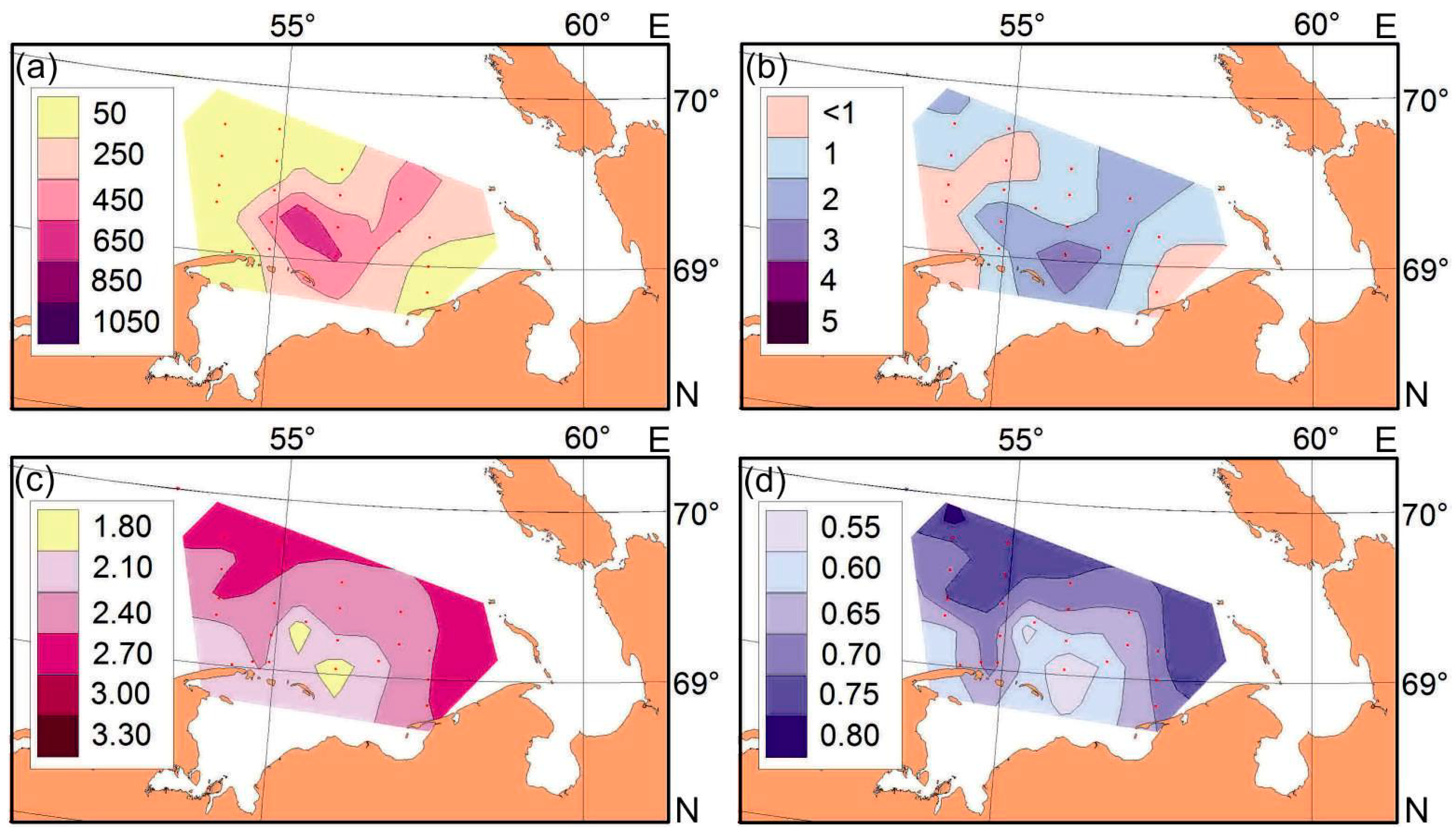
3.1. Environmental Conditions
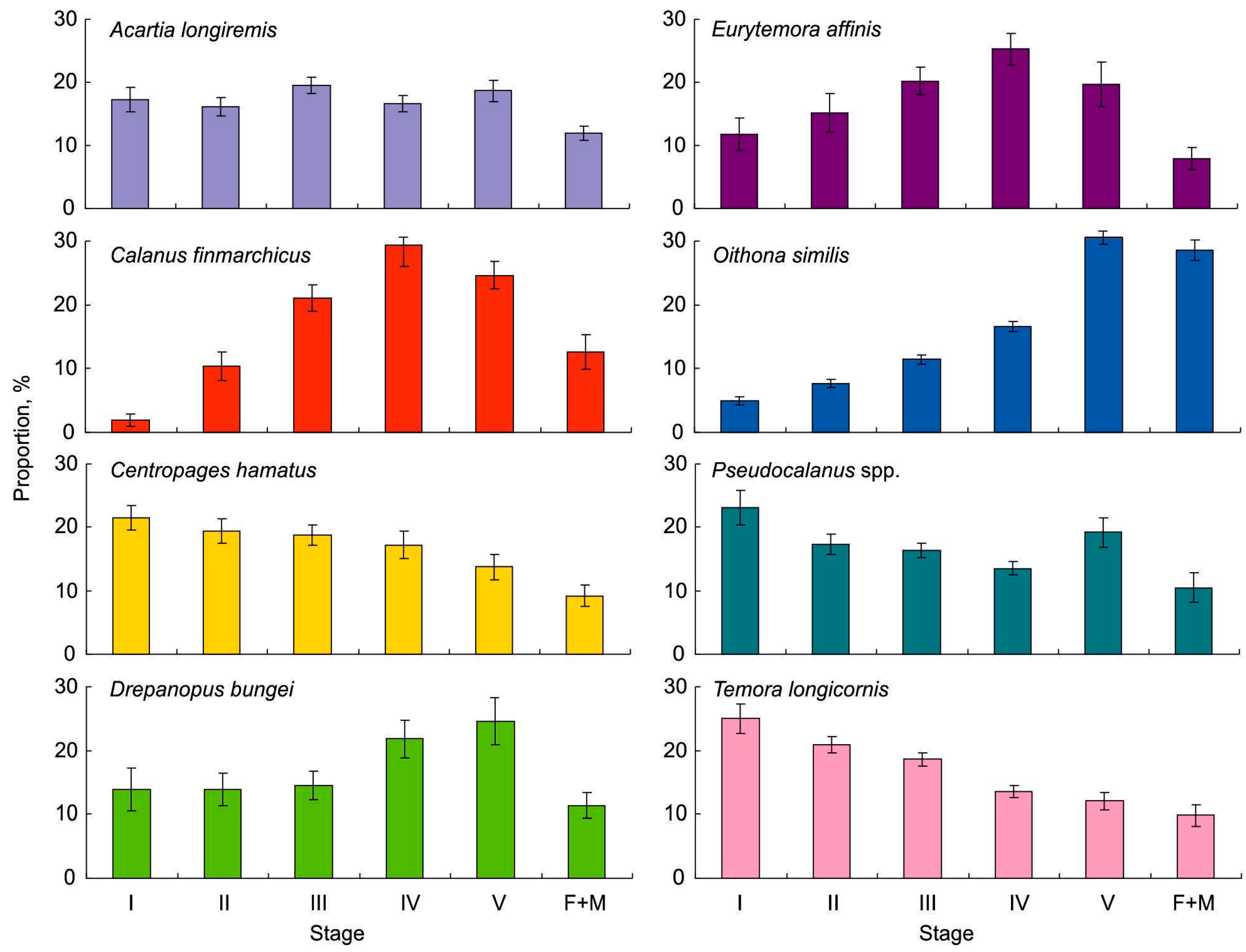
3.2. Copepod Composition, Abundance, Biomass, Diversity, and Population Structure of Common Taxa
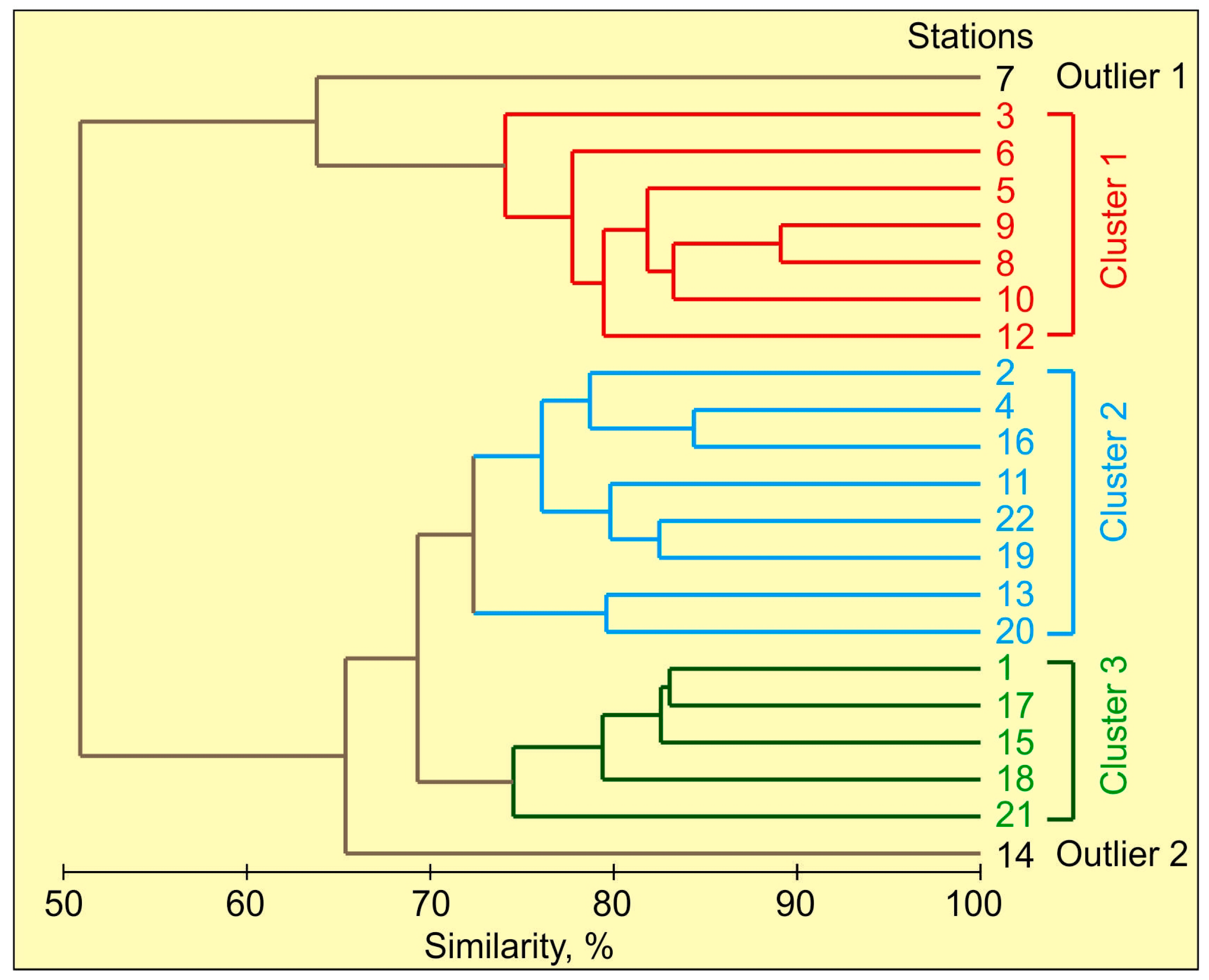
3.3. Copepod Assemblages
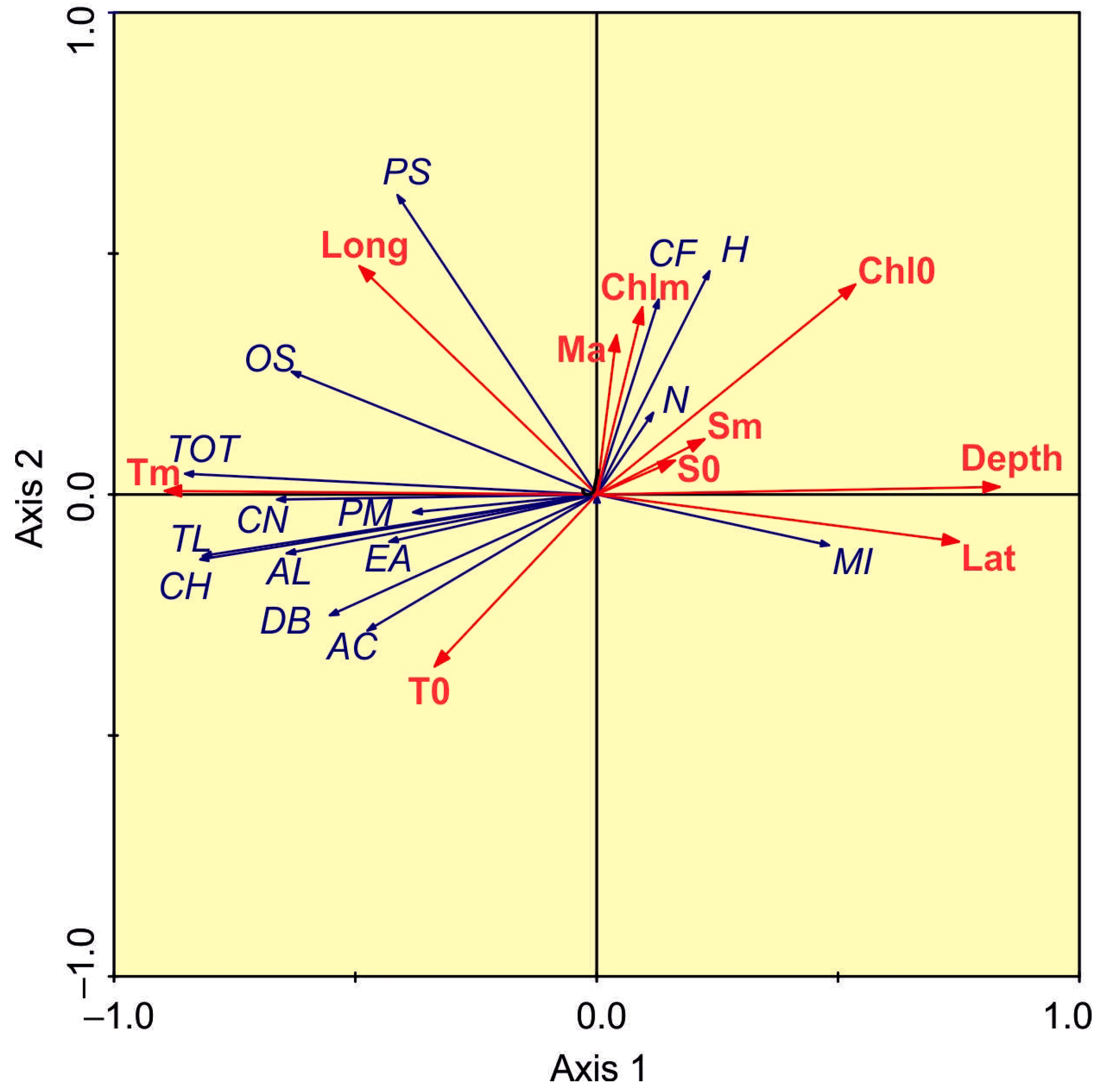
3.4. Environmental Control of Copepod Assemblages
4. Discussion
4.1. Environmental Conditions
4.2. Copepod Composition, Abundance, Biomass, Diversity, and Population Structure of Common Taxa
4.3. Copepod Assemblages
4.4. Environmental Control of Copepod Assemblages
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wassmann, P.; Reigstad, M.; Haug, T.; Rudels, B.; Carroll, M.L.; Hop, H.; Gabrielsen, G.W.; Falk-Petersen, S.; Denisenko, S.G.; Arashkevich, E.; et al. Food webs and carbon flux in the Barents Sea. Prog. Oceanogr. 2006, 71, 232–287. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Ecology and Distribution of Red King Crab Larvae in the Barents Sea: A Review. Water 2022, 14, 2328. [Google Scholar] [CrossRef]
- Ingvaldsen, R.; Loeng, H. Physical Oceanography. In Ecosystem Barents Sea; Sakshaug, E., Johnsen, G., Kovacs, K., Eds.; Tapir Academic Press: Trondheim, Norway, 2009; pp. 33–64. [Google Scholar]
- Loeng, H. Features of the physical oceanographic conditions in the central parts of the Barents Sea. Polar Res. 1991, 10, 5–18. [Google Scholar] [CrossRef] [Green Version]
- Ozhigin, V.K.; Ingvaldsen, R.B.; Loeng, H.; Boitsov, V.; Karsakov, A. Introduction to the Barents Sea. In The Barents Sea ecosystem: Russian-Norwegian cooperation in science and management; Jakobsen, T., Ozhigin, V., Eds.; Tapir Academic Press: Trondheim, Norway, 2011; pp. 315–328. [Google Scholar]
- Nikiforov, S.L.; Dunaev, N.N.; Politova, N.V. Modern environmental conditions of the Pechora Sea (climate, currents, waves, ice regime, tides, river runoff, and geological structure). Ber. Polarforsch. 2005, 501, 7–38. [Google Scholar]
- Sukhotin, A.; Denisenko, S.; Galaktionov, K. Pechora Sea ecosystems: Current state and future challenges. Polar Biol. 2019, 42, 1631–1645. [Google Scholar] [CrossRef]
- Sakshaug, E.; Johnsen, G.; Kristiansen, S.; von Quillfeldt, C.; Rey, F.; Slagstad, D.; Thingstad, F. Phytoplankton and primary production. In Ecosystem Barents Sea; Sakshaug, E., Johnsen, G., Kovacs, K., Eds.; Tapir Academic Press: Trondheim, Norway, 2009. [Google Scholar]
- Makarevich, P.; Druzhkova, E.; Larionov, V. Primary producers of the Barents Sea. In Diversity of Ecosystems; Mahamane, A., Ed.; In Tech: Rijeka, Croatia, 2012; pp. 367–392. [Google Scholar]
- Makarevich, P.R.; Vodopianova, V.V.; Bulavina, A.S.; Vashchenko, P.S.; Ishkulova, T.G. Features of the distribution of chlorophyll-a concentration along the western coast of the Novaya Zemlya archipelago in spring. Water 2021, 13, 3648. [Google Scholar] [CrossRef]
- Raymont, J.E.G. Plankton and Productivity in the Ocean, 2. Zooplankton, 2nd ed.; Pergamon Press: Oxford, UK, 1983. [Google Scholar]
- Eiane, K.; Tande, K.S. Meso and macrozooplankton. In Ecosystem Barents Sea; Sakshaug, E., Johnsen, G., Kovacs, K., Eds.; Tapir Academic Press: Trondheim, Norway, 2009; pp. 209–234. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Commercial fish and shellfish in the Barents Sea: Have introduced crab species affected the population trajectories of commercial fish? Rev. Fish Biol. Fisheries 2015, 25, 297–322. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Red king crab (Paralithodes camtschaticus) fisheries in Russian waters: Historical review and present status. Rev. Fish Biol. Fish. 2017, 28, 331–353. [Google Scholar] [CrossRef]
- Olsen, E.; Aanes, S.; Mehl, S.; Holst, J.C.; Aglen, A.; Gjøsæter, H. Cod, haddock, saithe, herring, and capelin in the Barents Sea and adjacent waters: A review of the biological value of the area. ICES J. Mar. Sci. 2009, 67, 87–101. [Google Scholar] [CrossRef]
- Hop, H.; Gjøsæter, H. Polar cod (Boreogadus saida) and capelin (Mallotus villosus) as key species in marine food webs of the Arctic and the Barents Sea. Mar. Biol. Res. 2013, 9, 878–894. [Google Scholar] [CrossRef]
- Semushin, A.V.; Novoselov, A.P.; Sherstkov, V.S.; Levitsky, A.L.; Novikova, Y.V. Long-term changes in the ichthyofauna of the Pechora Sea in response to ocean warming. Polar Biol. 2019, 42, 1739–1751. [Google Scholar] [CrossRef]
- Shiskin, A.N. Ecological Atlas. Barents Sea; NIR Moscow: Moscow Russia, 2020. (In Russian) [Google Scholar]
- Margiotta, F.; Balestra, C.; Buondonno, A.; Casotti, R.; D’Ambra, I.; Di Capua, I.; Gallia, R.; Mazzocchi, M.G.; Merquiol, L.; Pepi, M.; et al. Do plankton reflect the environmental quality status? The case of a post-industrial Mediterranean Bay. Mar. Environ. Res. 2020, 160, 104980. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Effects of environmental factors on the abundance, biomass, and individual weight of juvenile red king crabs in the Barents Sea. Front. Mar. Sci. 2020, 7, 726. [Google Scholar] [CrossRef]
- McGinty, N.; Barton, A.D.; Record, N.R.; Finkel, Z.V.; Johns, D.G.; Stock, C.A.; Irwin, A.J. Anthropogenic climate change impacts on copepod trait biogeography. Glob. Chang. Biol. 2020, 27, 1431–1442. [Google Scholar] [CrossRef]
- Brodsky, K.A. Calanoida of the Far Eastern Seas and Polar Basin of the USSR; Israel Program Scientific Translation: Jerusalem, Israel, 1967. [Google Scholar]
- Castellani, C.; Edwards, M. Marine Plankton: A Practical Guide to Ecology, Methodology, and Taxonomy; Oxford University Press: Oxford, UK, 2017. [Google Scholar]
- Vadstein, O. Interactions in the planktonic food web. In Ecosystem Barents Sea; Sakshaug, E., Johnsen, G., Kovacs, K., Eds.; Tapir Academic Press: Trondheim, Norway, 2009; pp. 251–266. [Google Scholar]
- Box, J.E.; Colgan, W.T.; Christensen, T.R.; Schmidt, N.M.; Lund, M.; Parmentier, F.-J.W.; Brown, R.; Bhatt, U.S.; Euskirchen, E.S.; Romanovsky, V.E.; et al. Key indicators of Arctic climate change: 1971–2017. Environ. Res. Lett. 2019, 14, 045010. [Google Scholar] [CrossRef]
- Long, Y.; Noman, A.; Chen, D.; Wang, S.; Yu, H.; Chen, H.; Wang, M.; Sun, J. Western Pacific Zooplankton Community along Latitudinal and Equatorial Transects in Autumn 2017 (Northern Hemisphere). Diversity 2021, 13, 58. [Google Scholar] [CrossRef]
- Richardson, A.J. In hot water: Zooplankton and climate change. ICES J. Mar. Sci. 2008, 65, 279–295. [Google Scholar] [CrossRef] [Green Version]
- Deschutter, Y.; Everaert, G.; De Schamphelaere, K.; De Troch, M. Relative contribution of multiple stressors on copepod density and diversity dynamics in the Belgian part of the North Sea. Mar. Pollut. Bull. 2017, 125, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Dvoretsky, V.G.; Dvoretsky, A.G. Summer mesozooplankton structure in the Pechora Sea (south-eastern Barents Sea). Estuarine Coast. Shelf Sci. 2009, 84, 11–20. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Summer mesozooplankton distribution near Novaya Zemlya (eastern Barents Sea). Polar Biol. 2009, 32, 719–731. [Google Scholar] [CrossRef]
- Dvoretsky, V.; Dvoretsky, A. Spatial variations in reproductive characteristics of the small copepod Oithona similis in the Barents Sea. Mar. Ecol. Prog. Ser. 2009, 386, 133–146. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Mesozooplankton structure in Dolgaya Bay (Barents Sea). Polar Biol. 2009, 33, 703–708. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Checklist of fauna found in zooplankton samples from the Barents Sea. Polar Biol. 2010, 33, 991–1005. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Copepod communities off Franz Josef Land (northern Barents Sea) in late summer of 2006 and 2007. Polar Biol. 2011, 34, 1231–1238. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Estimated copepod production rate and structure of mesozooplankton communities in the coastal Barents Sea during summer–autumn 2007. Polar Biol. 2012, 35, 1321–1342. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Epiplankton in the Barents sea: Summer variations of mesozooplankton biomass, community structure and diversity. Cont. Shelf Res. 2012, 52, 1–11. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Structure of mesozooplankton community in the Barents Sea and adjacent waters in August 2009. J. Nat. Hist. 2013, 47, 2095–2114. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Summer mesozooplankton community of Moller Bay (Novaya Zemlya Archipelago, Barents Sea). Oceanologia 2013, 55, 205–218. [Google Scholar] [CrossRef] [Green Version]
- Dvoretsky, V.G.; Dvoretsky, A.G. Mesozooplankton in the Kola Transect (Barents Sea): Autumn and winter structure. J. Sea Res. 2018, 142, 125–131. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Summer-fall macrozooplankton assemblages in a large Arctic estuarine zone (south-eastern Barents Sea): Environmental drivers of spatial distribution. Mar. Environ. Res. 2022, 173. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Coastal Mesozooplankton Assemblages during Spring Bloom in the Eastern Barents Sea. Biology 2022, 11, 204. [Google Scholar] [CrossRef]
- Dalpadado, P.; Arrigo, K.R.; van Dijken, G.L. Skjoldal, H.R.; Bagøien, E, Dolgov A.V.; Prokopchuk, I.P.; Sperfeld, E. Climate effects on temporal and spatial dynamics of phytoplankton and zooplankton in the Barents Sea. Progr. Oceanogr. 2020, 182, 102320. [Google Scholar]
- Usov, N.; Khaitov, V.; Smirnov, V.; Sukhotin, A. Spatial and temporal variation of hydrological characteristics and zooplankton community composition influenced by freshwater runoff in the shallow Pechora Sea. Polar Biol. 2018, 42, 1647–1665. [Google Scholar] [CrossRef]
- Lorenzen, C.J. Determination of chlorophyll and pheo-pigments: Spectrophotometric equations. Limnol. Oceanogr. 1967, 12, 343–346. [Google Scholar] [CrossRef]
- Parsons, T.R.; Maita, Y.; Lalli, C.M. A manual of chemical & biological methods for seawater analysis. Mar. Pollut. Bull. 1984, 1, 101–112. [Google Scholar]
- Jaschnov, V.A. Order Copepoda. In Guide to the Fauna and Flora of Northern Seas of the USSR; Gaevskaya, N.S., Ed.; Soviet Nauka Press: Moscow, USSR, 1948; pp. 183–215. (In Russian) [Google Scholar]
- Shuvalov, V.S. Copepod Cyclopoids of the Family Oithonidae of the World Ocean; Nauka Press: Leningrad, USSR, 1980. (In Russian) [Google Scholar]
- Madsen, S.D.; Nielsen, T.G.; Hansen, B.W. Annual population development and production by Calanus finmarchicus, C. glacialis and C. hyperboreus in Disko Bay, western Greenland. Mar. Biol. 2001, 139, 75–93. [Google Scholar]
- Dvoretsky, V.G. Distribution of Calanus species off Franz Josef Land (Arctic Barents Sea). Polar Sci. 2011, 5, 361–373. [Google Scholar] [CrossRef] [Green Version]
- Choquet, M.; Kosobokova, K.; Kwaśniewski, S.; Hatlebakk, M.; Dhanasiri, A.K.S.; Melle, W.; Daase, M.; Svensen, C.; Søreide, J.E.; Hoarau, G. Can morphology reliably distinguish between the copepods Calanus finmarchicus and C. glacialis, or is DNA the only way? Limnol. Oceanogr. Methods 2018, 16, 237–252. [Google Scholar] [CrossRef] [Green Version]
- Frost, B.W. A taxonomy of the marine calanoid copepod genus Pseudocalanus. Can. J. Zool. 1989, 67, 525–551. [Google Scholar] [CrossRef]
- Berggreen, U.; Hansen, B.; Kiørboe, T. Food size spectra, ingestion and growth of the copepod Acartia tonsa during development: Implications for determination of copepod production. Mar. Biol. 1988, 99, 341–352. [Google Scholar] [CrossRef]
- Ashjian, C.J.; Campbell, R.G.; Welch, H.E.; Butler, M.; Keuren, D.V. Annual cycle in abundance, distribution, and size in relation to hydrography of important copepod species in the western Arctic Ocean. Deep-Sea Res. I 2003, 50, 1235–1261. [Google Scholar] [CrossRef]
- Klein Breteler, W.C.M.; Fransz, H.G.; Gonzales, S.R. Growth and development of four calanoid copepod species under experimental and natural conditions. Neth. J. Sea Res. 1982, 16, 195–207. [Google Scholar] [CrossRef]
- Sabatini, M.; Kiørboe, T. Egg production, growth and development of the cyclopoid copepod Oithona similis. J. Plankton Res. 1994, 16, 1329–1351. [Google Scholar] [CrossRef] [Green Version]
- Satapoomin, S. Carbon content of some common tropical Andaman Sea copepods. J. Plankton Res. 1999, 21, 2117–2123. [Google Scholar] [CrossRef]
- Middlebrook, K.; Roff, J.C. Comparison of Methods for Estimating Annual Productivity of the Copepods Acartia hudsonica and Eurytemora herdmani in Passamaquoddy Bay, New Brunswick. Can. J. Fish. Aquat. Sci. 1986, 43, 656–664. [Google Scholar] [CrossRef]
- Richter, C. Regional and seasonal variability in the vertical distribution of mesozooplankton in the Greenland Sea. Ber. Zur Polarforsch. 1994. [Google Scholar] [CrossRef]
- Kankaala, P.; Johansson, S. The influence of individual variation on length-biomass regressions in three crustacean zooplankton species. J. Plankton Res. 1986, 8, 1027–1038. [Google Scholar] [CrossRef]
- Liu, H.; Hopcroft, R.R. Growth and development of Metridia pacifica (Copepoda: Calanoida) in the northern Gulf of Alaska. J. Plankton Res. 2006, 28, 769–781. [Google Scholar] [CrossRef] [Green Version]
- Webber, M.K.; Roff, J.C. Annual biomass and production of the oceanic copepod community off Discovery Bay, Jamaica. Mar. Biol. 1995, 123, 481–495. [Google Scholar] [CrossRef]
- Postel, L.; Fock, H.; Hagen, W. Biomass and abundance. In ICES Zooplankton Methodology Manual; Harris, R., Wiebe, P., Lenz, J., Skjoldal, H.R., Huntley, M., Eds.; Academic Press: London, UK, 2000; pp. 83–192. [Google Scholar]
- Field, J.; Clarke, K.; Warwick, R. A Practical Strategy for Analysing Multispecies Distribution Patterns. Mar. Ecol. Prog. Ser. 1982, 8, 37–52. [Google Scholar] [CrossRef]
- Shannon, C.B.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, USA, 1963. [Google Scholar]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- ter Braak, C.J.F.; Verdonschot, P.F.M. Canonical correspondence analysis and related multivariate methods in aquatic ecology. Aquat. Sci. 1995, 57, 255–289. [Google Scholar] [CrossRef]
- ter Braak, C.J.F.; Smilauer, P. CANOCO Reference Manualand CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (version 4.5); Microcomputer Power: Ithaca, NY, USA, 2002. [Google Scholar]
- Pfirman, S.; Kögeler, J.W.; Anselme, B. Coastal environments of the Western Kara and Eastern Barents Seas. Deep Sea Res. 1995, 42, 1391–1412. [Google Scholar] [CrossRef]
- Matishov, G.G.; Zuyev, A.N.; A Golubev, V.; Adrov, N.M.; Timofeev, S.F.; Karamusko, O.; Pavlova, L.; Fadyakin, O.; Buzan, A.; Braunstein, A.; et al. Original source of the “Climatic Atlas of the Arctic Seas 2004: Part I. Database of the Barents, Kara, Laptev, and White Seas-Oceanography and Marine Biology”. Available online: https://elibrary.ru/item.asp?id=23200679 (accessed on 29 December 2022).
- Drinkwater, K.; Loeng, H.; Titov, O.V.; Boitsov, V.D. Global warming and climate change. In The Barents Sea. Ecosystem, Resources, Management; Jakobsen, T., Ozhigin, V.K., Eds.; Tapir Academic Publishers: Trondheim, Norway, 2011; pp. 777–807. [Google Scholar]
- Møller, E.F.; Nielsen, T.G. Borealization of Arctic zooplankton – smaller and less fat zooplankton species in Disko Bay. Western Greenland. Limnol. Oceanogr. 2020, 65, 1175–1188. [Google Scholar] [CrossRef]
- Evseeva, O.Y.; Ishkulova, T.G.; Dvoretsky, A.G. Environmental Drivers of an Intertidal Bryozoan Community in the Barents Sea: A Case Study. Animals 2022, 12, 552. [Google Scholar] [CrossRef] [PubMed]
- Dvoretsky, A.G.; Dvoretsky, V.G. Inter-annual dynamics of the Barents Sea red king crab (Paralithodes camtschaticus) stock indices in relation to environmental factors. Polar Sci. 2016, 10, 541–552. [Google Scholar] [CrossRef]
- Makarevich, P.R.; Vodopianova, V.V.; Bulavina, A.S. Dynamics of the Spatial Chlorophyll-A Distribution at the Polar Front in the Marginal Ice Zone of the Barents Sea during Spring. Water 2022, 14, 101. [Google Scholar] [CrossRef]
- Lischka, S.; Knickmeier, K.; Hagen, W. Mesozooplankton assemblages in the shallow Arctic Laptev Sea in summer 1993 and autumn 1995. Polar Biol. 2001, 24, 186–199. [Google Scholar] [CrossRef]
- Fetzer, I.; Hirche, H.; Kolosova, E. The influence of freshwater discharge on the distribution of zooplankton in the southern Kara Sea. Polar Biol. 2002, 25, 404–415. [Google Scholar] [CrossRef] [Green Version]
- Schmid, M.K.; Piepenburg, D.; Golikov, A.A.; von Juterzenka, K.; Petryashov, V.V.; Spindler, M. Trophic pathways and carbon flux patterns in the Laptev Sea. Prog. Oceanogr. 2006, 71, 314–330. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Regional differences of mesozooplankton communities in the Kara Sea. Cont. Shelf Res. 2015, 105, 26–41. [Google Scholar] [CrossRef]
- Smoot, C.A.; Hopcroft, R.R. Cross-shelf gradients of epipelagic zooplankton communities of the Beaufort Sea and the influence of localized hydrographic features. J. Plankton Res. 2016, 39, 65–78. [Google Scholar] [CrossRef] [Green Version]
- Turner, J.T. The importance of small planktonic copepods and their roles in pelagic marine food webs. Zool. Stud. 2004, 43, 255–266. [Google Scholar]
- Dvoretsky, V.G.; Venger, M.P.; Vashchenko, A.V.; Maksimovskaya, T.M.; Ishkulova, T.G.; Vodopianova, V.V. Pelagic Bacteria and Viruses in a High Arctic Region: Environmental Control in the Autumn Period. Biology 2022, 11, 845. [Google Scholar] [CrossRef]
- Falk-Petersen, S.; Timofeev, S.; Pavlov, V.; Sargent, J.R. Climate variability and possible effects on arctic food chains: The role of Calanus. In Arctic Alpine Ecosystems and People in a Changing Environment; Orbok, J.B., Tombre, T., Kallenborn, R., Hegseth, E., Falk-Petersen, S., Hoel, A.H., Eds.; Springer Verlag: Berlin, Germany, 2007; pp. 147–166. [Google Scholar]
- Lischka, S.; Hagen, W. Life histories of the copepod Psedocalanus minutus, P. acuspes (Calanoida) and Oithona similis (Cyclopoida) in the Arctic Kongsfjorden (Svalbard). Polar Biol. 2005, 28, 910–921. [Google Scholar] [CrossRef]
- A Ershova, E.; Nyeggen, M.U.; A Yurikova, D.; E Søreide, J. Seasonal dynamics and life histories of three sympatric species of Pseudocalanus in two Svalbard fjords. J. Plankton Res. 2021, 43, 209–223. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wen, Y.; Zhang, G.; Zhang, G.; Sun, J.; Xu, W. Effects of Terrestrial Inputs on Mesozooplankton Community Structure in Bohai Bay, China. Diversity 2022, 14, 410. [Google Scholar] [CrossRef]
- Seo, M.-H.; Kim, H.-J.; Lee, S.-J.; Kim, S.-Y.; Yoon, Y.-H.; Han, K.-H.; Choi, S.-D.; Kwak, M.-T.; Jeong, M.-K.; Soh, H.-Y. Environmental factors affecting the spatiotemporal distribution of copepods in a small mesotidal inlet and estuary. Diversity 2021, 13, 389. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Macrozooplankton of the Arctic – The Kara Sea in relation to environmental conditions. Estuarine, Coast. Shelf Sci. 2017, 188, 38–55. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Summer macrozooplankton assemblages of Arctic shelf: A latitudinal study. Cont. Shelf Res. 2019, 188, 103967. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Mesozooplankton structure in the northern White Sea in July 2008. Polar Biol. 2010, 34, 469–474. [Google Scholar] [CrossRef]
- Gislason, A.; Petursdottir, H.; Astthorsson, O.S.; Gudmundsson, K.; Valdimarsson, H. Inter-annual variability in abundance and community structure of zooplankton south and north of Iceland in relation to environmental conditions in spring 1990–2007. J. Plankton Res. 2009, 31, 541–551. [Google Scholar] [CrossRef] [Green Version]
- Norrbin, F.; Eilertsen, H. Chr.; Degerlund, M. Vertical distribution of primary producers and zooplankton grazers during different phases of the Arctic spring bloom. Deep-Sea Res. II 2009, 56, 1945–1958. [Google Scholar] [CrossRef]
- Blachowiak-Samolyk, K.; Kwasniewski, S.; Hop, H.; Falk-Petersen, S. Magnitude of mesozooplankton variability: A case study from the Marginal Ice Zone of the Barents Sea in spring. J. Plankton Res. 2007, 30, 311–323. [Google Scholar] [CrossRef] [Green Version]
- Trudnowska, E.; Sagan, S.; Kwasniewski, S.; Darecki, M.; Blachowiak-Samolyk, K. Fine-scale zooplankton vertical distribution in relation to hydrographic and optical characteristics of the surface waters on the Arctic shelf. J. Plankton Res. 2014, 37, 120–133. [Google Scholar] [CrossRef]
- Trudnowska, E.; Gluchowska, M.; Beszczynska-Möller, A.; Blachowiak-Samolyk, K.; Kwasniewski, S. Plankton patchiness in the Polar Front region of the West Spitsbergen Shelf. Mar. Ecol. Prog. Ser. 2016, 560, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Jacobsen, S.; Gaard, E.; Húsgarð Larsen, K.M.; Eliasen, S.K.; Hátún, H. Temporal and spatial variability of zooplankton on the Faroe shelf in spring 1997–2016. J. Mar. Syst. 2018, 177, 28–38. [Google Scholar] [CrossRef]
- Turner, J.T.; Tester, P.A. Zooplankton feeding ecology: Bacterivory by metazoan microzooplankton. J. Exp. Mar. Biol. Ecol. 1992, 160, 149–167. [Google Scholar] [CrossRef]
- Froneman, P.W. Predator Diversity Does Not Contribute to Increased Prey Risk: Evidence from a Mesocosm Study. Diversity 2022, 14, 584. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| St. | Date (dd.mo.ye) | Local Time | Coordinates | Depth, m | Sampling Layer, m | |
|---|---|---|---|---|---|---|
| 1 | 29 July 2012 | 4:15 | 69°23.2′ N | 55°59.9′ E | 22 | 20–0 |
| 2 | 29-July-2012 | 0:20 | 69°12.1′ N | 56°00.2′ E | 20 | 18–0 |
| 3 | 29-July-2012 | 16:50 | 69°32.6′ N | 56°00.9′ E | 36 | 34–0 |
| 4 | 29-July-2012 | 4:10 | 69°15.1′ N | 55°27.8′ E | 23 | 21–0 |
| 5 | 31-July-2012 | 20:20 | 69°45.1′ N | 54°52.4′ E | 54 | 50–0 |
| 6 | 31-July-2012 | 15:35 | 69°33.2′ N | 54°00.4′ E | 54 | 50–0 |
| 7 | 01-August-2012 | 0:50 | 69°45.3′ N | 54°00.4′ E | 74 | 70–0 |
| 8 | 01-August-2012 | 6:50 | 69°33.3′ N | 54°55.6′ E | 38 | 35–0 |
| 9 | 02-August-2012 | 17:05 | 69°23.3′ N | 54°54.6′ E | 29 | 27–0 |
| 10 | 02-August-2012 | 23:10 | 69°23.1′ N | 53°59.7′ E | 42 | 40–0 |
| 11 | 03-August-2012 | 15:33 | 69°12.1′ N | 54°55.8′ E | 24 | 20–0 |
| 12 | 03-August-2012 | 7:57 | 69°17.0′ N | 53°59.0′ E | 34 | 32–0 |
| 13 | 03-August-2012 | 21:32 | 69°05.7′ N | 56°40.5′ E | 13 | 11–0 |
| 14 | 04-August-2012 | 21:37 | 69°23.4′ N | 56°59.6′ E | 19 | 17–0 |
| 15 | 05-August-2012 | 4:15 | 69°10.6′ N | 57°29.7′ E | 18 | 16–0 |
| 16 | 05-August-2012 | 0:25 | 69°11.5′ N | 57°00.5′ E | 15 | 13–0 |
| 17 | 05-August-2012 | 7:00 | 69°00.8′ N | 57°30.7′ E | 17 | 14–0 |
| 18 | 05-August-2012 | 13:20 | 68°56.6′ N | 57°34.2′ E | 18 | 16–0 |
| 19 | 06-August-2012 | 15:55 | 69°03.8′ N | 54°57.5′ E | 15 | 13–0 |
| 20 | 06-August-2012 | 8:50 | 69°02.3′ N | 55°59.9′ E | 18 | 15–0 |
| 21 | 06-August-2012 | 20:50 | 69°02.3′ N | 54°38.9′ E | 10 | 8–0 |
| 22 | 07-August-2012 | 13:10 | 69°00.7′ N | 54°19.0′ E | 10 | 7.5–0 |
| Taxon | Equation | Reference |
|---|---|---|
| Acartia spp. | Ln DW (μg) = 2.92 Ln PL (μm)–18.316 | [52] |
| Calanus finmarchicus | Ln C (mg) = 3.5687 Ln PL (mm)–1.004 | [48] |
| Calanus glacialis | Ln DW (mg) = 3.414 Ln PL (mm)–4.605 | [53] |
| Calanus hyperboreus | Ln DW (mg) = 3.718 Ln PL (mm)–5.809 | [53] |
| Centropages hamatus | Lg DW (μg) = 2.4492 Lg PL (μm)–6.0984 | [54] |
| Copepoda nauplii | Ln DW (μg) = 3.31 Ln PL (μm)–19.566 | [52] |
| Cyclopina gracilisa | Ln C (μg) = 2.16 Ln PL (μm)–13.870 | [55] |
| Drepanopus bungeib | Lg DW (μg) = 2.7302 Lg PL (μm)–6.9121 | [54] |
| Ectinosoma spp. c | Ln C (μg) = 1.15 Ln TL (μm)–7.79 | [56] |
| Eurytemora affinis | Lg DW (μg) = 2.96 Lg PL (μm)–7.6 | [57] |
| Jaschnovia tollid | Lg DW (mg) = 3.412 Lg PL (mm)–2 | [58] |
| Limnocalanus macrurus | Ln C (μg) = 1.47 Ln PL (mm) + 0.239 | [59] |
| Metridia longae | Lg DW (μg) = 3.29 Lg PL (μm)–8.75 | [60] |
| Microcalanus pusillusa | Ln C (μg) = 2.16 Ln PL (μm)–13.870 | [55] |
| Microsetella norvegica | Ln C (μg) = 1.15 Ln TL (μm)–7.79 | [56] |
| Oithona atlanticaa | Ln C (μg) = 2.16 Ln PL (μm)–13.870 | [55] |
| Oithona similis | Ln C (μg) = 2.16 Ln PL (μm)–13.870 | [55] |
| Pseudocalanus spp. | Lg DW (μg) = 2.7302 Lg PL (μm)–6.9121 | [54] |
| Temora longicornis | Lg DW (μg) = 3.064 Lg PL (μm)–7.6958 | [54] |
| Triconia borealisf | Ln DW (μg) = 2.10 Ln PL (μm)–11.62 | [61] |
| Taxon | Abundance | Biomass | |||
|---|---|---|---|---|---|
| Occurrence | Range | Mean ± SE | Range | Mean ± SE | |
| Acartia longiremis | 95 | 0–120/0–6020 | 16 ± 5/1007 ± 322 | 0–843/0–42.2 | 108 ± 38/6.5 ± 2.1 |
| Acartia clausi | 36 | 0–9/0–626 | 1 ± 0/43 ± 28 | 0–106/0–7.1 | 7 ± 5/0.5 ± 0.3 |
| Calanus finmarchicus | 95 | 0–16/0–916 | 2 ± 1/93 ± 41 | 0–1766/0–103.9 | 250 ± 85/10.2 ± 4.7 |
| Calanus glacialis | 18 | 0–3/0–46 | <1/4 ± 2 | 0–1426/0–20.4 | 88 ± 65/1.6 ± 1 |
| Calanus hyperboreus | 5 | <1/0–6 | <1/<1 | 0–197/0–2.8 | 9 ± 9/0.1 ± 0.1 |
| Centropages hamatus | 100 | 0–77/3–5688 | 24 ± 5/1591 ± 371 | 3–937/0–51.3 | 220 ± 54/14.8 ± 3.7 |
| Copepoda nauplii | 100 | 8–502/142–23,916 | 72 ± 22/4152 ± 1192 | 3–139/0.1–6.6 | 25 ± 6/1.3 ± 0.3 |
| Cyclopina gracilis | 5 | <1/0–2 | < 1/< 1 | <1/<1 | <1/<1 |
| Drepanopus bungei | 82 | 0–32/0–2938 | 4 ± 2/313 ± 1 47 | 0–59/0–4.5 | 8 ± 3/0.6 ± 0.3 |
| Ectinosoma spp. | 14 | <1/0–2 | < 1/< 1 | 0–1/<1.1 | <1/<1 |
| Eurytemora affinis | 91 | 0–39/0–3568 | 3 ± 2/22 ± 160 | 0–189/0–17.2 | 15 ± 8/1.2 ± 0.8 |
| Jaschnovia tolli | 9 | 0–1/0–43 | <1/3 ± 2 | 0–10/<1.6 | 1 ± 0/<1 |
| Limnocalanus macrurus | 18 | 0–1/0–98 | <1/7 ± 5 | 0–19/0–1.3 | 2 ± 1/0.1 ± 0.1 |
| Metridia longa | 5 | 0–1/0–10 | <1/<1 | 0–4/<1.1 | <1/<1 |
| Microcalanus pusillus | 36 | 0–6/0–106 | 1 ± 0/15 ± 6 | 0–18/<1.3 | 2 ± 1/<1 |
| Microsetella norvegica | 23 | <1/0–20 | <1/2 ± 1 | <1/<1 | <1/<1 |
| Oithona atlantica | 9 | 0–2/0–26 | <1/1 ± 1 | 0–2/<1 | <1/<1 |
| Oithona similis | 100 | 9–214/416–12,589 | 54 ± 10/2864 ± 599 | 9–225/0.4–13.2 | 52 ± 10/2.8 ± 0.6 |
| Triconia borealis | 27 | <1/0–22 | <1/2 ± 1 | 0–1/<1.1 | <1/<1 |
| Pseudocalanus spp. I-IV | 100 | 1–201/121–11,844 | 35 ± 9/1903 ± 550 | 2–251/0.2–14.7 | 55 ± 11/2.9 ± 0.7 |
| Pseudocalanus minutus V-VI | 100 | 0–25/27–825 | 6 ± 1/204 ± 44 | 2–228/0.2–6.7 | 47 ± 12/1.7 ± 0.4 |
| Pseudocalanus acuspes V-VI | 100 | 0–31/55–1816 | 7 ± 2/305 ± 88 | 3–244/0.4–14.4 | 51 ± 13/2.3 ± 0.7 |
| Pseudocalanus major | 36 | 0–3/0–183 | 1 ± 0/33 ± 13 | 0–11/0–0.7 | 2 ± 1/0.1 ± 0 |
| Temora longicornis | 95 | 0–420/0–28,032 | 89 ± 22/5949 ± 1544 | 0–2901/0–193.4 | 624 ± 158/42.6 ± 11.8 |
| Total | 69–935/1581–47,381 | 314 ± 52/18,720 ± 3376 | 312–4239/11–283 | 1565 ± 250/89 ± 18 | |
| Abundance | Biomass | ||||
| Taxon | Occurence | Range | Mean ± SE | Range | Mean ± SE |
| Acartia longiremis | 95 | 0–120/0–6020 | 16 ± 5/1007 ± 322 | 0–843/0–42.2 | 108 ± 38/6.5 ± 2.1 |
| Acartia clausi | 36 | 0–9/0–626 | 1 ± 0/43 ± 28 | 0–106/0–7.1 | 7 ± 5/0.5 ± 0.3 |
| Calanus finmarchicus | 95 | 0–16/0–916 | 2 ± 1/93 ± 41 | 0–1766/0–103.9 | 250 ± 85/10.2 ± 4.7 |
| Calanus glacialis | 18 | 0–3/0–46 | <1/4 ± 2 | 0–1426/0–20.4 | 88 ± 65/1.6 ± 1 |
| Calanus hyperboreus | 5 | <1/0–6 | <1/<1 | 0–197/0–2.8 | 9 ± 9/0.1 ± 0.1 |
| Centropages hamatus | 100 | 0–77/3–5688 | 24 ± 5/1591 ± 371 | 3–937/0–51.3 | 220 ± 54/14.8 ± 3.7 |
| Copepoda nauplii | 100 | 8–502/142–23,916 | 72 ± 22/4152 ± 1192 | 3–139/0.1–6.6 | 25 ± 6/1.3 ± 0.3 |
| Cyclopina gracilis | 5 | <1/0–2 | <1/<1 | <1/<1 | <1/<1 |
| Drepanopus bungei | 82 | 0–32/0–2938 | 4 ± 2/313 ± 1 47 | 0–59/0–4.5 | 8 ± 3/0.6 ± 0.3 |
| Ectinosoma spp. | 14 | <1/0–2 | < 1/< 1 | 0–1/<1.1 | <1/<1 |
| Eurytemora affinis | 91 | 0–39/0–3568 | 3 ± 2/22 ± 160 | 0–189/0–17.2 | 15 ± 8/1.2 ± 0.8 |
| Jaschnovia tolli | 9 | 0–1/0–43 | <1/3 ± 2 | 0–10/<1.6 | 1 ± 0/<1 |
| Limnocalanus macrurus | 18 | 0–1/0–98 | <1/7 ± 5 | 0–19/0–1.3 | 2 ± 1/0.1 ± 0.1 |
| Metridia longa | 5 | 0–1/0–10 | <1/<1 | 0–4/<1.1 | <1/<1 |
| Microcalanus pusillus | 36 | 0–6/0–106 | 1 ± 0/15 ± 6 | 0–18/<1.3 | 2 ± 1/<1 |
| Microsetella norvegica | 23 | <1/0–20 | <1/2 ± 1 | <1/<1 | <1/<1 |
| Oithona atlantica | 9 | 0–2/0–26 | <1/1 ± 1 | 0–2/<1 | <1/<1 |
| Oithona similis | 100 | 9–214/416–12,589 | 54 ± 10/2864 ± 599 | 9–225/0.4–13.2 | 52 ± 10/2.8 ± 0.6 |
| Triconia borealis | 27 | <1/0–22 | <1/2 ± 1 | 0–1/<1.1 | <1/<1 |
| Pseudocalanus spp. I-IV | 100 | 1–201/121–11,844 | 35 ± 9/1903 ± 550 | 2–251/0.2–14.7 | 55 ± 11/2.9 ± 0.7 |
| Pseudocalanus minutus V-VI | 100 | 0–25/27–825 | 6 ± 1/204 ± 44 | 2–228/0.2–6.7 | 47 ± 12/1.7 ± 0.4 |
| Pseudocalanus acuspes V-VI | 100 | 0–31/55–1816 | 7 ± 2/305 ± 88 | 3–244/0.4–14.4 | 51 ± 13/2.3 ± 0.7 |
| Pseudocalanus major | 36 | 0–3/0–183 | 1 ± 0/33 ± 13 | 0–11/0–0.7 | 2 ± 1/0.1 ± 0 |
| Temora longicornis | 95 | 0–420/0–28,032 | 89 ± 22/5949 ± 1544 | 0–2901/0–193.4 | 624 ± 158/42.6 ± 11.8 |
| Total | 69–935/1581–47,381 | 314 ± 52/18,720 ± 3376 | 312–4239/11–283 | 1565 ± 250/89 ± 18 | |
| Average Similarity, % | 78.68 | 75.59 | 78.45 |
|---|---|---|---|
| Taxon | Cluster 1 | Cluster 2 | Cluster 3 |
| Oithona similis | 21.00 | 14.63 | 16.61 |
| Copepoda nauplii | 18.71 | 16.60 | 15.43 |
| Pseudocalanus minutus/acuspes V-VI | 15.26 | 5.16 | 7.90 |
| Pseudocalanus minutus/acuspes I-IV | 12.94 | 10.23 | 10.37 |
| Temora longicornis | 8.37 | 26.46 | 20.47 |
| Acartia longiremis | 8.32 | 7.65 | 6.90 |
| Calanus finmarchicus | 3.69 | 0.87 | 1.23 |
| Centropages hamatus | 2.35 | 12.72 | 11.72 |
| Eurytemora affinis | 1.38 | 2.30 | 3.00 |
| Microcalanus pusillus | 0.74 | – | 0.10 |
| Drepanopus bungei | 0.71 | 2.31 | 4.55 |
| Acartia clausi | – | 0.84 | 0.13 |
| Parameter | Lat | Long | Tm | Sm | T0 | S0 | Chl0 | Chlm | Depth | MAb |
|---|---|---|---|---|---|---|---|---|---|---|
| N | 0.00 | 0.35 | −0.09 | −0.37 | −0.11 | −0.42 | 0.19 | −0.02 | 0.17 | 0.51 |
| AL | −0.43 | −0.01 | 0.55 | −0.03 | 0.32 | 0.04 | −0.40 | 0.01 | −0.53 | −0.20 |
| AC | −0.40 | 0.07 | 0.42 | −0.06 | 0.35 | 0.00 | −0.21 | −0.22 | −0.39 | −0.22 |
| CF | 0.43 | 0.07 | −0.23 | 0.33 | −0.46 | 0.31 | 0.21 | 0.14 | 0.20 | 0.09 |
| CH | −0.70 | 0.34 | 0.77 | −0.35 | 0.55 | −0.26 | −0.39 | −0.11 | −0.73 | 0.08 |
| CN | −0.35 | 0.17 | 0.53 | −0.03 | 0.24 | 0.04 | −0.39 | −0.33 | −0.53 | −0.06 |
| DB | −0.49 | 0.43 | 0.54 | −0.41 | 0.46 | −0.34 | −0.19 | −0.28 | −0.50 | 0.03 |
| EA | −0.39 | 0.26 | 0.40 | −0.24 | 0.31 | −0.18 | −0.29 | −0.26 | −0.43 | 0.13 |
| MI | 0.49 | −0.24 | −0.55 | 0.26 | −0.41 | 0.18 | 0.09 | −0.21 | 0.65 | 0.12 |
| OS | −0.31 | 0.41 | 0.57 | −0.14 | 0.20 | −0.08 | −0.15 | 0.05 | −0.52 | 0.13 |
| PS | −0.20 | 0.58 | 0.37 | −0.16 | 0.00 | −0.13 | 0.10 | 0.12 | −0.36 | 0.17 |
| PM | −0.49 | 0.64 | 0.49 | −0.67 | 0.46 | −0.65 | 0.13 | 0.07 | −0.37 | 0.27 |
| TL | −0.63 | 0.38 | 0.74 | −0.27 | 0.52 | −0.18 | −0.33 | −0.17 | −0.69 | −0.11 |
| TOT | −0.56 | 0.41 | 0.75 | −0.21 | 0.41 | −0.12 | −0.35 | −0.19 | −0.71 | −0.02 |
| H | 0.37 | −0.01 | −0.42 | 0.01 | −0.32 | −0.06 | 0.29 | 0.25 | 0.45 | 0.16 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dvoretsky, V.G.; Dvoretsky, A.G. Copepod Assemblages in A Large Arctic Coastal Area: A Baseline Summer Study. Diversity 2023, 15, 81. https://doi.org/10.3390/d15010081
Dvoretsky VG, Dvoretsky AG. Copepod Assemblages in A Large Arctic Coastal Area: A Baseline Summer Study. Diversity. 2023; 15(1):81. https://doi.org/10.3390/d15010081
Chicago/Turabian StyleDvoretsky, Vladimir G., and Alexander G. Dvoretsky. 2023. "Copepod Assemblages in A Large Arctic Coastal Area: A Baseline Summer Study" Diversity 15, no. 1: 81. https://doi.org/10.3390/d15010081
APA StyleDvoretsky, V. G., & Dvoretsky, A. G. (2023). Copepod Assemblages in A Large Arctic Coastal Area: A Baseline Summer Study. Diversity, 15(1), 81. https://doi.org/10.3390/d15010081