3.2. Phyllopodopsyllus saywakimi Kim & Lee, 2023 sp. nov.
Figure 2,
Figure 3,
Figure 4,
Figure 5,
Figure 6,
Figure 7,
Figure 8,
Figure 9,
Figure 10,
Figure 11,
Figure 12,
Figure 13,
Figure 14,
Figure 15,
Figure 16,
Figure 17,
Figure 18 and
Figure 19.
Zoobank registration
urn:lsid:zoobank.org:act:36472E42-EBEE-4ACE-9E96-7E09D9D37AE1
Type locality
Intertidal sand of the rocky shore, Udo, Republic of Korea, 23 July 2020. (L3: 33°31′14.53″ N 126°57′29.64″ E)
Specimens examined
Holotype: 1 female (MABIKCR00252865)
Paratype: 9 females and 10 males (NIBRIV0000901821–NIBRIV0000901839). The voucher specimen information is given in detail in
Table S1.
Etymology
The new species (saywakimi, Latin genitive, masculine) is named in honor of Prof. Saywa Kim, one of the first author’s scientific mentors who contributed to the ecology of marine zooplankton from Korean waters.
Description of adult female (based on the holotype and paratype 1–9)
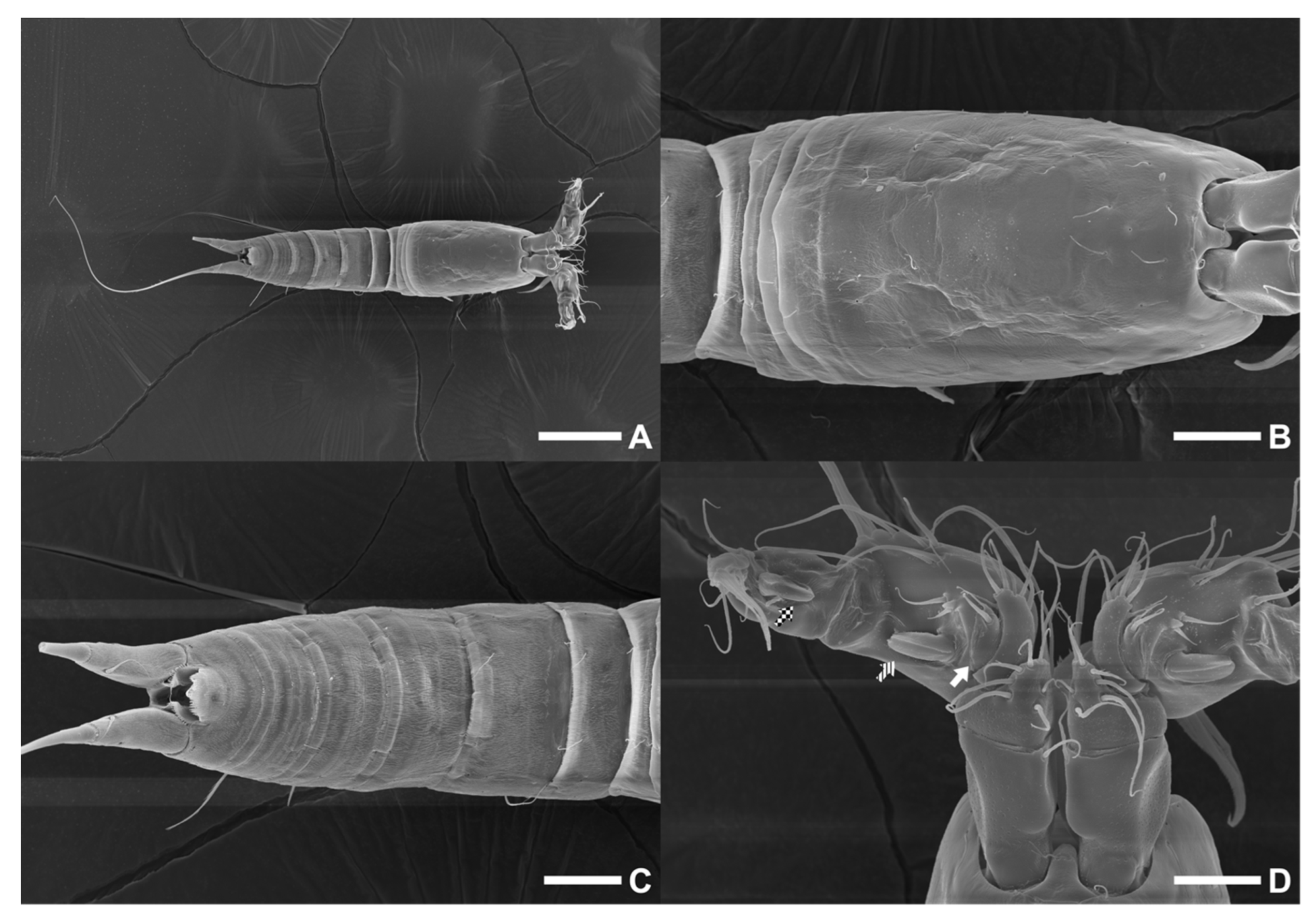
Body (
Figure 2A,
Figure 3A–C,
Figure 4A–E,
Figure 5,
Figure 6A,
Figure 7A,B,
Figure 8, and
Figure 9A) nine-segmented, cylindrical, slightly constricted in middle, furnished with sensilla, cuticular pores, (micro)spinules, and/or (micro)setules (details in figures), and without dimples on all body surfaces except for cephalothorax (resulting from complete fusion of cephalosome and first pedigerous somite).
Prosome (
Figure 2A,
Figure 3A,B,D,
Figure 4A–C,
Figure 6A,
Figure 7A,B, and
Figure 8A) four-segmented, slightly tapering distally and comprising cephalothorax and three free pedigerous somites. Cephalothorax somewhat longer than all succeeding prosomites combined and covered with vague punctures dorsally. Three free pedigerous somites nearly equal in length. Hyaline frills of prosomites smooth but frills of last prosomite finely serrated dorsally. Rostrum small, dimpled, weakly defined at base, semi-trapezoid in dorsal aspect, and with sensillum near each anterior corner. Eye not visible.
Urosome (
Figure 2A,
Figure 3A,C,
Figure 4A,D,E,
Figure 5,
Figure 7A,B,
Figure 8B–D, and
Figure 9A,B) five-segmented, comprising fifth pedigerous somite, genital double-somite (resulting from fusion of second and third urosomites), two abdominal somites and anal somite armed with caudal rami, and slightly longer than prosome (including caudal rami). Fifth pedigerous somite shortest and wrinkled on ventral surface; hyaline frills of fifth pedigerous somite finely serrated and smooth laterally. Genital double-somite longest among urosomites, armed with sixth pereiopod (P6), and with vestigial original segmentation marked by pattern of surface ornamentation, ventral suture, and dorsal and lateral ridge, of which dorsal one faint; hyaline frills of genital double-somite finely fringed dorsally and smooth laterally and ventrally; terminal part of lateral frill ornamented with spinules regularly; genital field with single ovoid copulatory pore near ventral suture, short copulatory duct in distal part of genital somite, and two seminal receptacles. Antepenultimate somite slightly longer than first urosomite and with finely serrated fringes dorsally; hyaline frill of antepenultimate somite smooth ventrally and laterally; lateral frill ornamented with spinules regularly; ventral frill extended as lappet, furnished with hair-like elements, and ornamented with long spinules on terminal margin (details in
Figure 5B and
Figure 9A). Penultimate somite about as long as preceding one; hyaline frill of penultimate somite finely serrated dorsally; lateral and ventral frills smooth and furnished with regular spinules terminally. Anal somite slightly narrower and longer than penultimate somite, cleft medially in posterior part; anal opening located in cleft; anal operculum rounded and densely ornamented with spinules terminally. Caudal rami cylindrical (outer margin longer than inner margin), about 3.3 times as long as greatest width, covered with microspinules throughout surface, ornamented with tube pore on outer distal margin, and armed with seven setae on each ramus, (three outer, one dorsal, and three terminal); setae I and II naked and set very close to each other dorsoventrally at outer margin medially; seta I tiny, thready, and about one fourth as long as seta II; seta III bipinnate, implanted near distal outer corner, and about as long as seta II; setae IV–VI bare and inserted inwardly; seta IV fused basally to seta V and about as long as caudal ramus; seta V modified into indefinable bulbous shape at base (see
Figure 3C,
Figure 4E,
Figure 5C and
Figure 8B–D) and about 3.5 times as long as caudal ramus; seta VI fine and approximately half in length to seta IV; seta VII located at inner distal third and triarticulate.
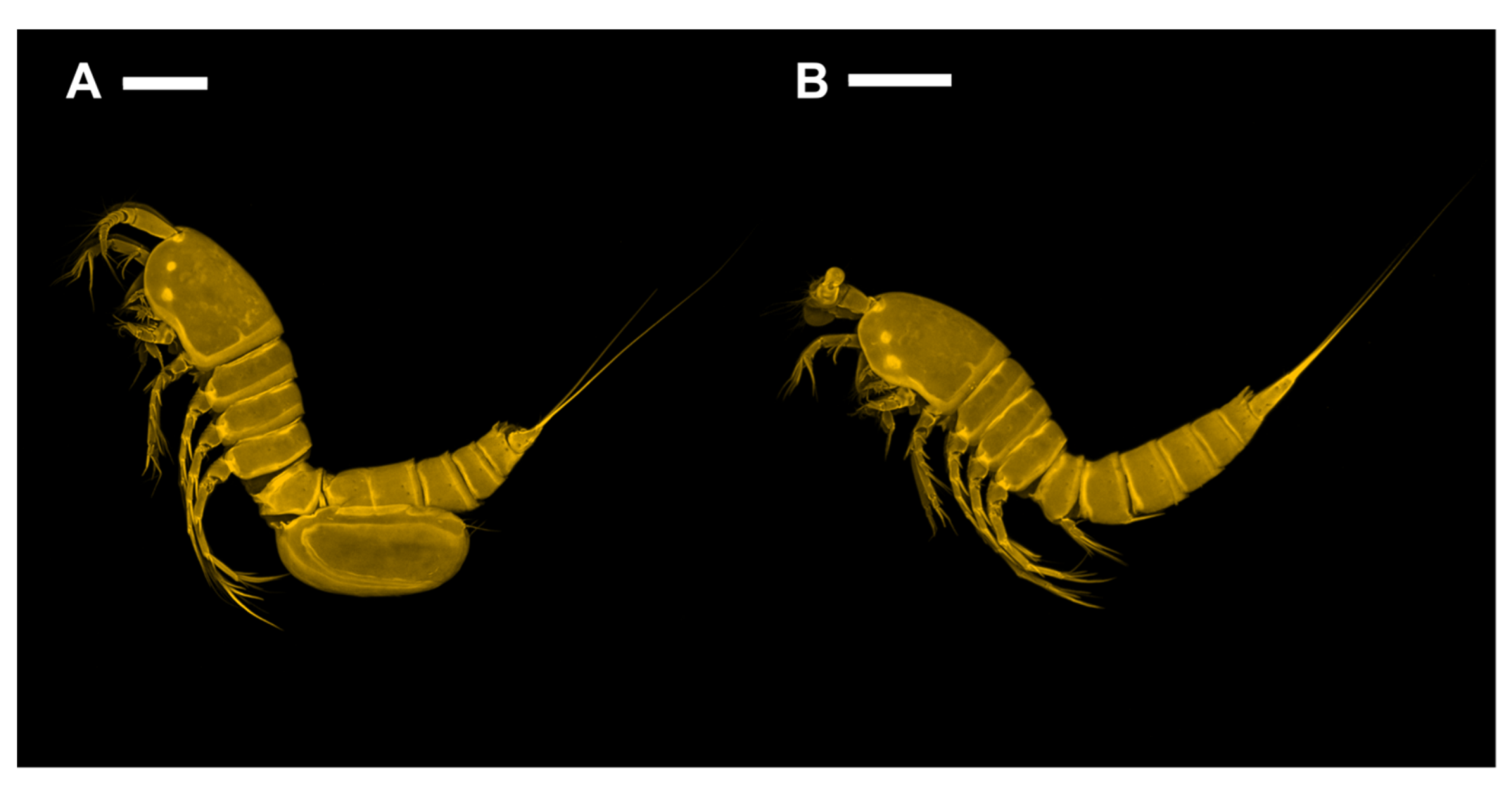
Figure 2.
Confocal laser scanning microscopy images of Phyllopodopsyllus saywakimi sp. nov. (A) female (paratype 1): habitus, lateral; (B) male (paratype 10): habitus, lateral. Scale bars: 100 µm.
Figure 2.
Confocal laser scanning microscopy images of Phyllopodopsyllus saywakimi sp. nov. (A) female (paratype 1): habitus, lateral; (B) male (paratype 10): habitus, lateral. Scale bars: 100 µm.
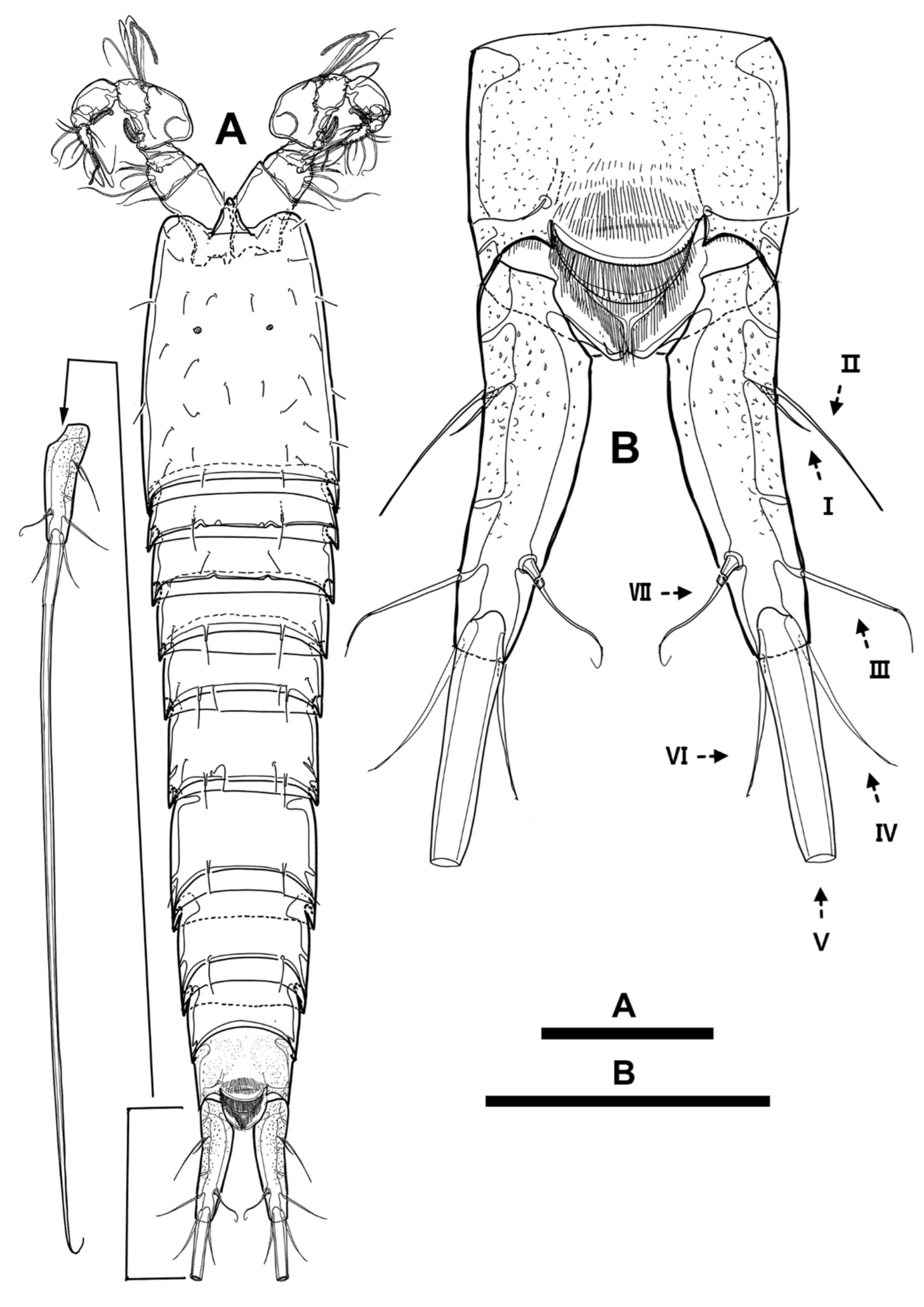
Antennule (
Figure 3D,
Figure 4F,
Figure 6B, and
Figure 10A) eight-segmented and covered with punctures; first segment elongated, longest, about four times longer than width, about 0.8 times as long as all succeeding segments combined, ornamented with row of small spinules on proximal inner margin and pore on dorsal surface proximally, and with pinnate seta on inner distal corner; second segment with robust pointy integumental projection on outer margin; fourth segment with pedestal armed with aesthetasc fused basally to long seta on inner distal corner; fifth segment shortest; eighth segment about as long as second one and with apical aesthetasc fused basally to two setae. Armature formula as follows: 1(1), 2(9), 3(8), 4(3+(ae+1)), 5(2), 6(4), 7(4), 8(5+(ae+2)); all setae naked and slender except for first two segments; all outer setae arising from penultimate and distal segments biarticulate.
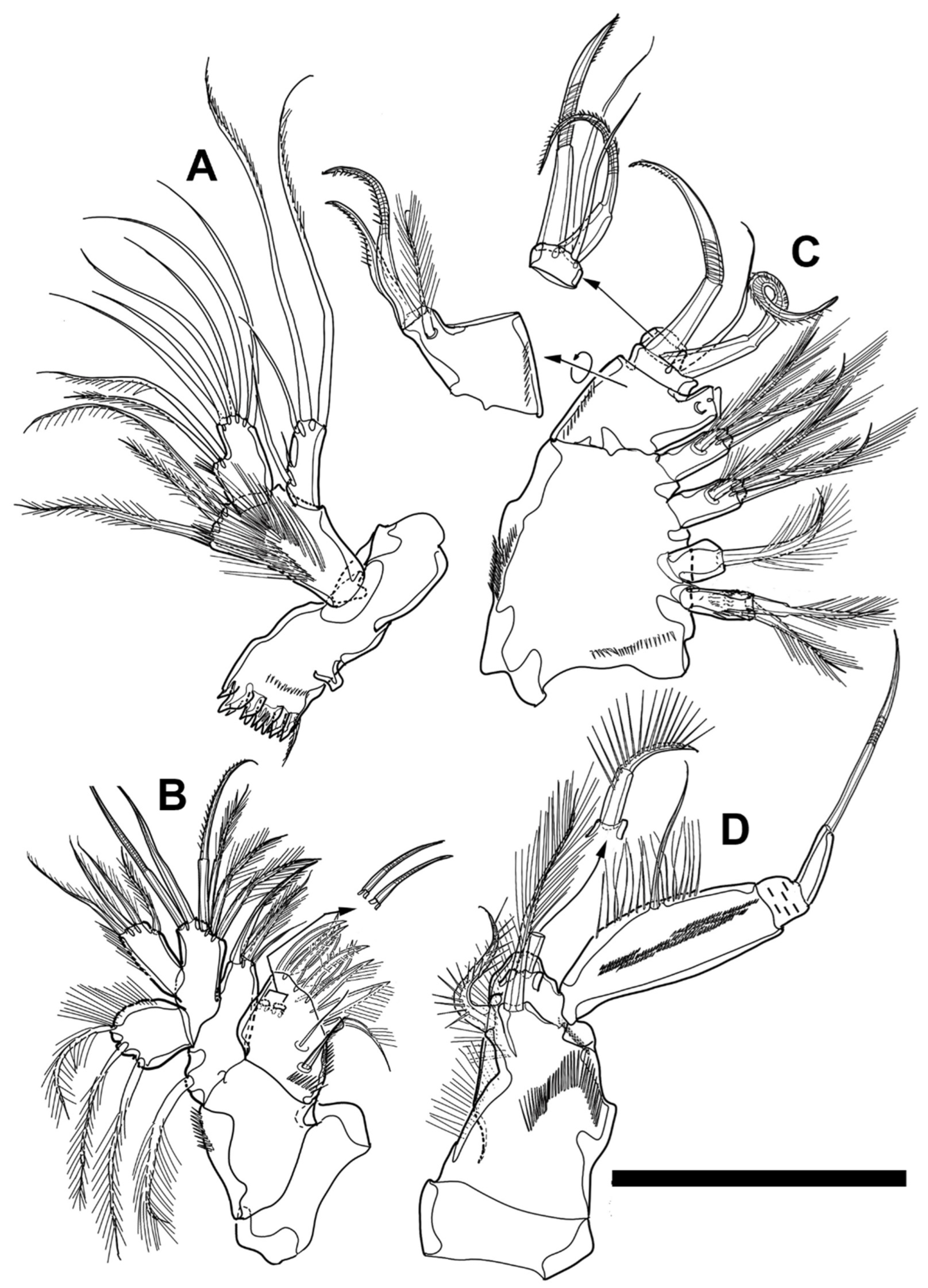
Antenna (
Figure 4F,
Figure 6C, and
Figure 10B) omposed of coxa, basis, endopod, and ex-opod. Coxa bare and about as long as greatest width. Basis unarmed, ornamented with longitudinal row of spinules along distal half of abexopodal margin, and about 2.2 times as long as width. Endopod two-segmented; first endopodal segment unarmed, about as long as basis, about 2.5 times as long as wide, and furnished with patch of spinules on proximal inner margin; second endopodal segment about 1.3 times longer than preceding segment, about 3.2 times as long as greatest width, decorated with patch of spinules on inner margin, row of strong spinules terminally, two rows of frills on middle and distal of outer margin, respectively, and armed with 11 armatures; four geniculate, two bare setae and one spine implanted terminally; two setae and two robust spines located in inner margin subapically; two lateral setae set very close to each other. Exopod one-segmented and with three pinnate setae (two apical and one lateral); outer apical seta fused basally to exopod.
Labrum (
Figure 6D and
Figure 7C) well-developed, gradually tapering ventrally, covered with dimples on each lateral side, decorated with patch of long hairly setules on distal margin posteriorly; cutting edge furnished with setules and strong spinules on cutting edge, and with cuticular pore on outside of each cutting edge.
Figure 3.
Scanning electron microscopy images of Phyllopodopsyllus saywakimi sp. nov. female (paratype 2) in dorsal view: (A) habitus; (B) prosome; (C) urosome; (D) antennule and rostrum. Scale bars: (A) 100 µm; (B–D) 50 µm.
Figure 3.
Scanning electron microscopy images of Phyllopodopsyllus saywakimi sp. nov. female (paratype 2) in dorsal view: (A) habitus; (B) prosome; (C) urosome; (D) antennule and rostrum. Scale bars: (A) 100 µm; (B–D) 50 µm.
Mandible (
Figure 6C and
Figure 11A) composed of coxa, basis, endopod, and exopod. Coxa enlarged and with gnathobase bearing two large ventral teeth, pinnate dorsal seta and row of multicuspidate teeth. Basis gradually widening distally, subpentagonal, about as long as coxa, furnished with two rows of long spinules on anterior surface and slender setules along distal half of inner margin, and armed with two pinnate setae inserted near inner distal corner and one plumose seta originating from distal margin medially. Endopod one-segmented, about 0.8 times as long as basis, and armed with two lateral pinnate setae and seven apical bare setae; lateral setae implanted very close to each other in proximal third. Exopod one-segmented, slender, about 0.36 times as long as endopod, and with five setae; two lateral setae implanted in proximal quarter and midlength, respectively, and other three setae located terminally.
Paragnaths (
Figure 6D) well-developed, composed of posterior and two lateral lobes; cutting edge of each lobe wrinkled; lateral robes ornamented with several rows of long and robust spinules on each inner side; posterior lobe decorated with some rows of spinules around wrinkles, medially.
Maxillule (
Figure 6D,E and
Figure 11B) composed of praecoxa, coxa, basis, endopod and exopod. Praecoxa large, trapezoidal, ornamented with row of spinules on proximal third of inner margin of ventral surface, and armed with well-developed arthrite; praecoxal arthrite with two pinnate spines arising from posterior margin, of which proximal spine shorter, two apical setae arising from distal corner, of which longer one plumose, two bare setae implanted dorsally, and seven robust unipinnate spines around distal margin. Coxa about 0.3 times shorter than praecoxa, armed with cylindrical endite incorporated into coxa and epipodite represented by one plumose seta; coxal endite with five setae (one naked and four pinnate) and ornamented with minute spinules on posterior sur-face. Basis unornamented and with eight setae apically. Endopod hexagonal, about twice as long as wide, unornamented, and with four plumose setae, of which three setae located around terminal margin and other seta inserted in lateral margin. Exopod about 1.3 times as long as greatest width, ornamented with setules along ventral margin, and with three plumose setae located in terminal margin.
Maxilla (
Figure 6E and
Figure 11C) composed of syncoxa, basis, and endopod. Syncoxa large and armed with four endites; endites with two, one, three, and three pinnate and/or plumose setae from proximal to distal, respectively; distal endite longest; proximal second endite shortest. Basis about 0.4 times as long as syncoxa, ornamented with row of spinules near terminal margin, and armed with cylindrical endite bearing one slender naked seta on distal margin, two robust apical spines and one bare seta arising from posterior surface; additional seta located in near boundary of basis and first endopodal segment (however not sure whether inserted in basis or following segment). Endopod two-segmented and much shorter than basis; first endopodal segment about 0.7 times as long as wide and with three setae of which two pinnate implanted around inner distal corner; other seta minute, thready, and located in anterior surface medially; second endopodal segment about equal to preceding segment in length and with four setae apically.
Figure 4.
Scanning electron microscopy images of Phyllopodopsyllus saywakimi sp. nov. female (paratype 3) in lateral view: (A) habitus; (B) cephalothorax; (C) second to fourth prosomites; (D) first to third urosomites and P5; (E) fourth and fifth urosomites, and caudal rami; (F) antennule and antenna. Scale bars: (A) 100 µm; (B–F) 50 µm.
Figure 4.
Scanning electron microscopy images of Phyllopodopsyllus saywakimi sp. nov. female (paratype 3) in lateral view: (A) habitus; (B) cephalothorax; (C) second to fourth prosomites; (D) first to third urosomites and P5; (E) fourth and fifth urosomites, and caudal rami; (F) antennule and antenna. Scale bars: (A) 100 µm; (B–F) 50 µm.
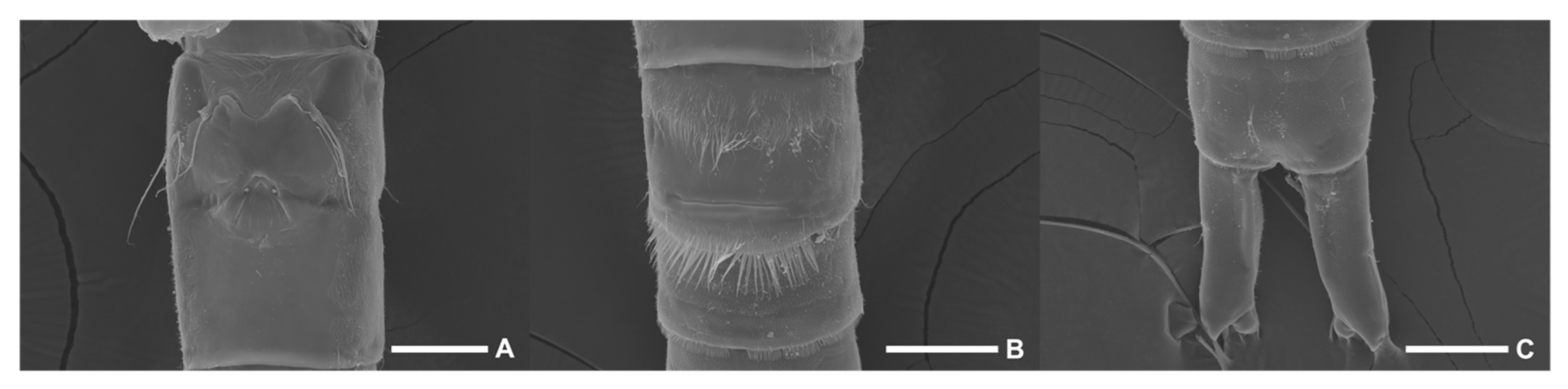
Figure 5.
Scanning electron microscopy images of Phyllopodopsyllus saywakimi sp. nov. female (paratype 4) in ventral view: (A) genital double-somite; (B) third and fourth urosomites; (C) fifth urosomite and caudal rami. Scale bars: 50 µm.
Figure 5.
Scanning electron microscopy images of Phyllopodopsyllus saywakimi sp. nov. female (paratype 4) in ventral view: (A) genital double-somite; (B) third and fourth urosomites; (C) fifth urosomite and caudal rami. Scale bars: 50 µm.
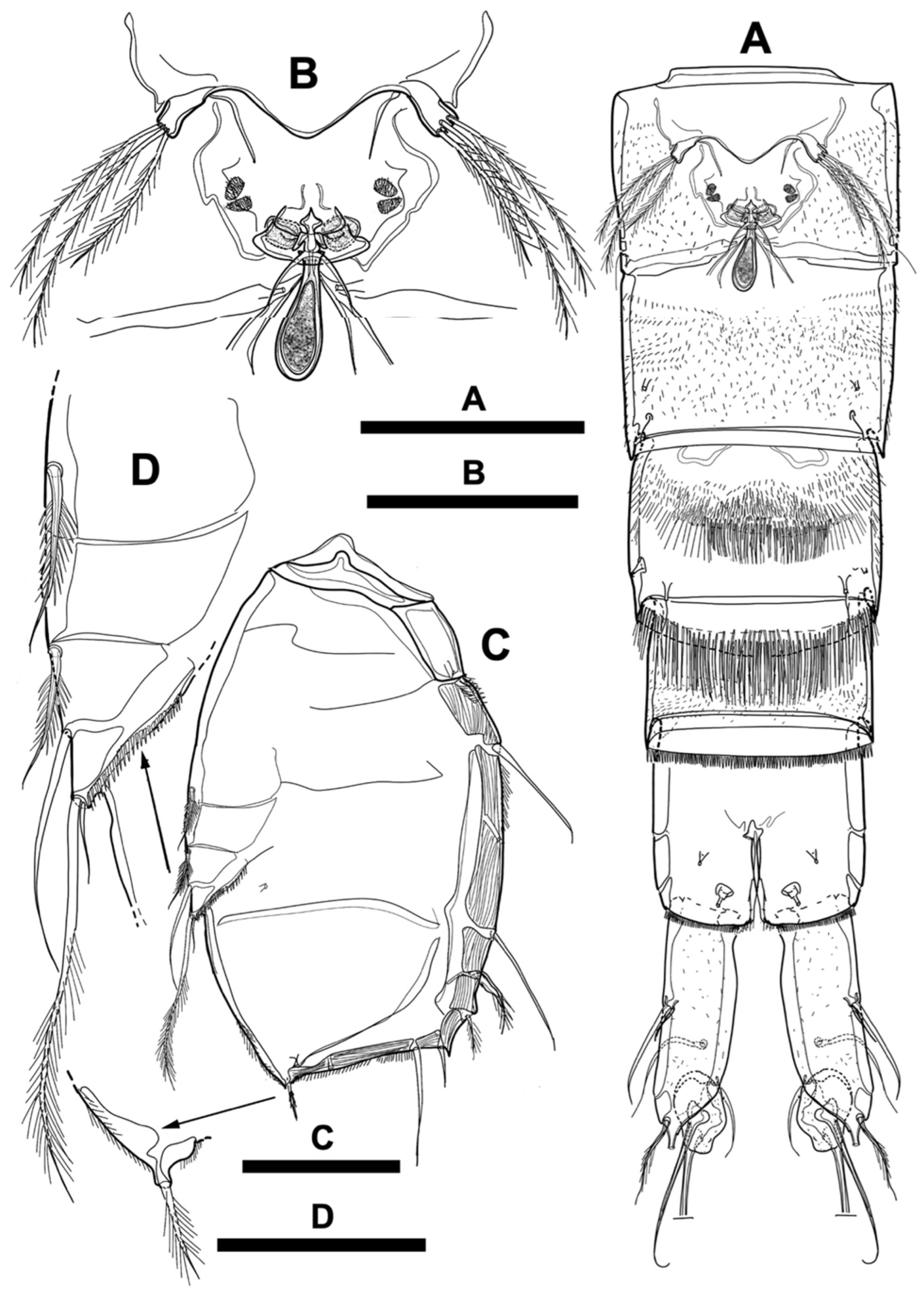
Figure 6.
Scanning electron microscopy images of Phyllopodopsyllus saywakimi sp. nov. female (paratype 4) in ventral view: (A) cephalothorax; (B) antennule; (C) antenna and mandible (excluding coxa); (D) mouth parts around oral opening; (E) maxillule and maxilla; (F) maxilliped. Scale bars: (A) 100 µm; (B–F) 20 µm.
Figure 6.
Scanning electron microscopy images of Phyllopodopsyllus saywakimi sp. nov. female (paratype 4) in ventral view: (A) cephalothorax; (B) antennule; (C) antenna and mandible (excluding coxa); (D) mouth parts around oral opening; (E) maxillule and maxilla; (F) maxilliped. Scale bars: (A) 100 µm; (B–F) 20 µm.
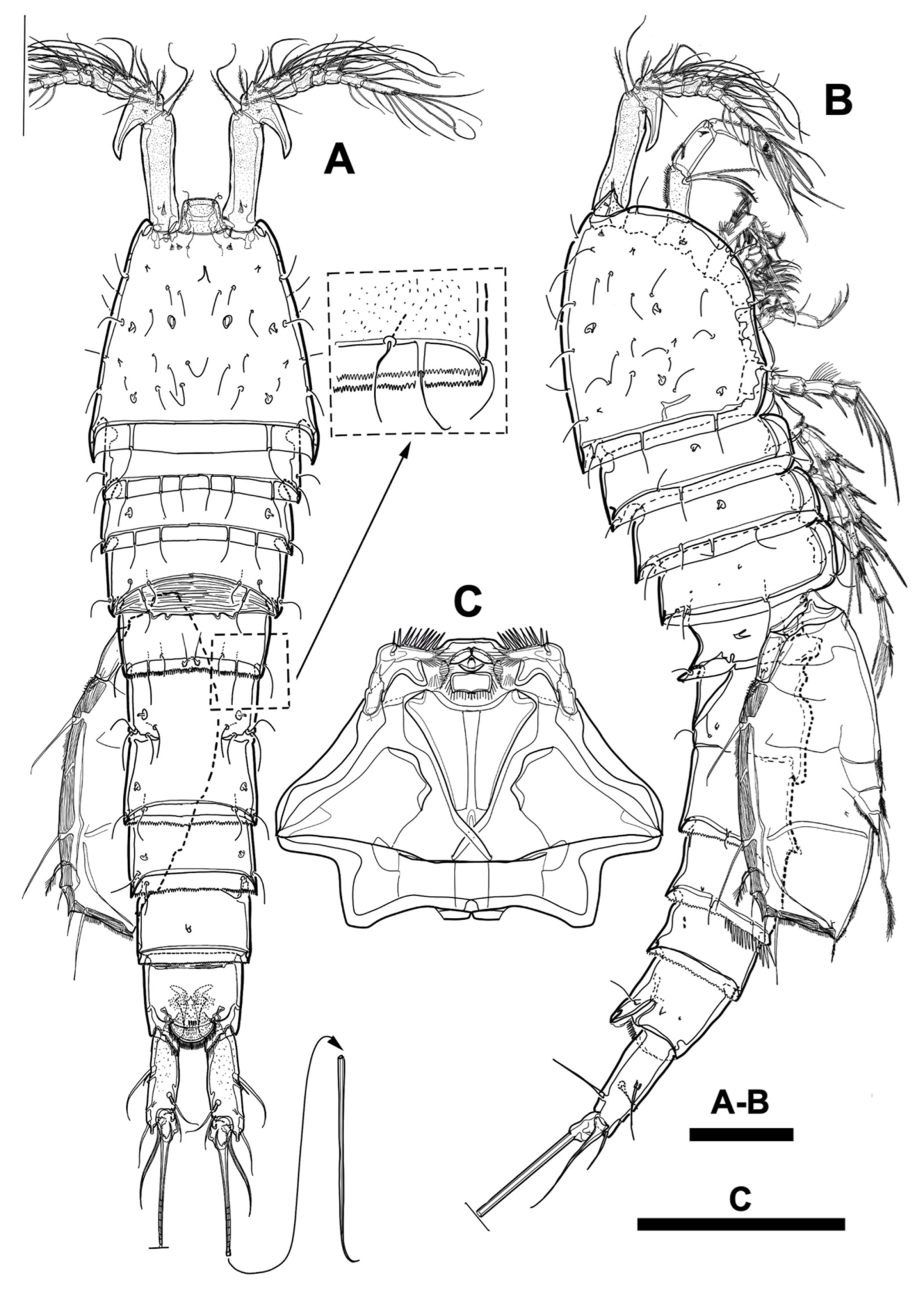
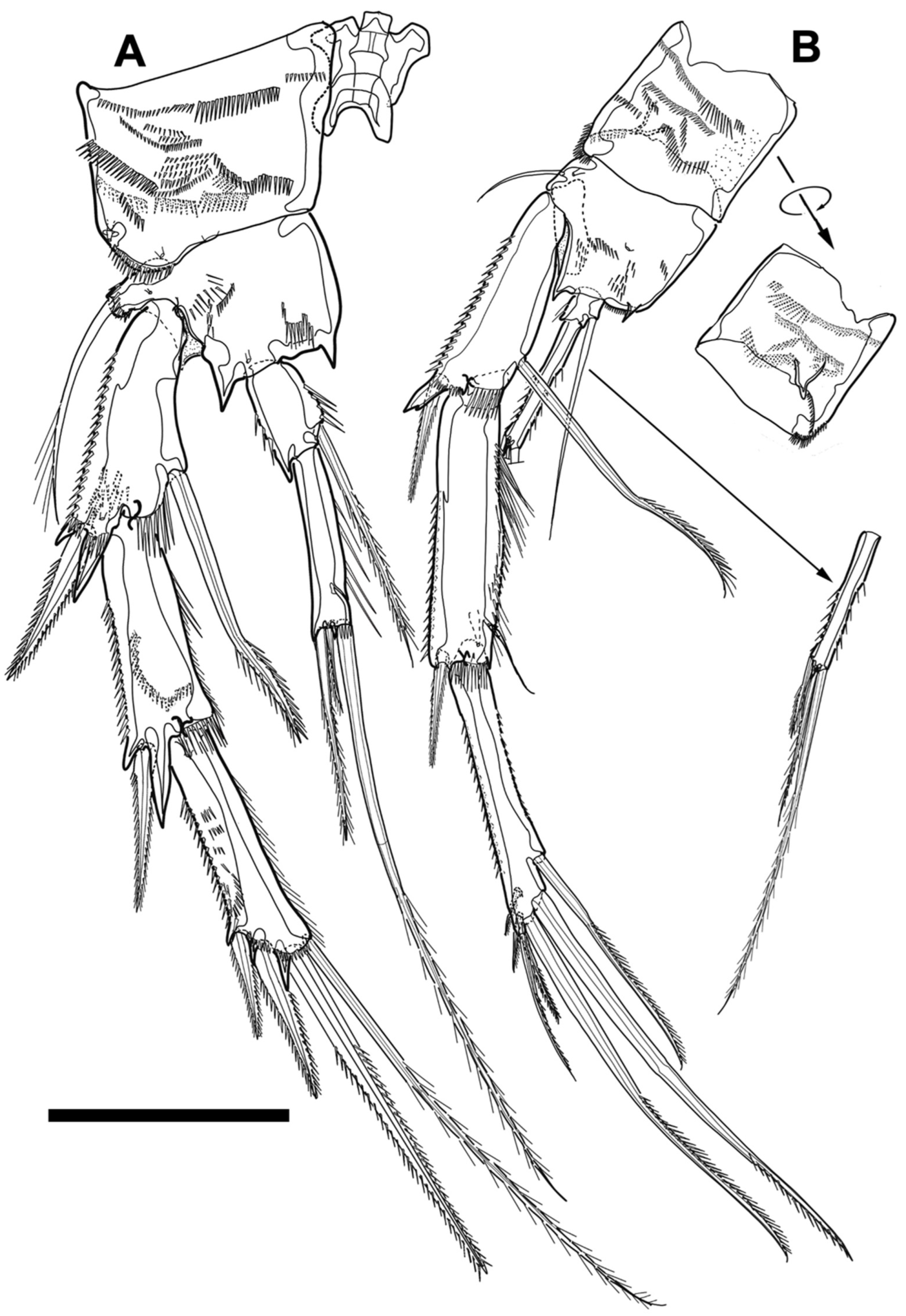
Figure 7.
Line drawings of Phyllopodopsyllus saywakimi sp. nov. female (holotype): (A) habitus, dorsal; (B) habitus, lateral; (C) rostrum, ventral. Scale bars: (A,B) 100 µm; (C) 50 µm.
Figure 7.
Line drawings of Phyllopodopsyllus saywakimi sp. nov. female (holotype): (A) habitus, dorsal; (B) habitus, lateral; (C) rostrum, ventral. Scale bars: (A,B) 100 µm; (C) 50 µm.
Figure 8.
Line drawings of Phyllopodopsyllus saywakimi sp. nov. female (holotype): (A) rostrum, dorsal; (B) fifth and caudal rami, dorsal; (C) fifth and caudal rami, ventral; (D) distal part of the caudal ramus, lateral. Scale bars: (A,C,D) 50 µm; (B) 100 µm.
Figure 8.
Line drawings of Phyllopodopsyllus saywakimi sp. nov. female (holotype): (A) rostrum, dorsal; (B) fifth and caudal rami, dorsal; (C) fifth and caudal rami, ventral; (D) distal part of the caudal ramus, lateral. Scale bars: (A,C,D) 50 µm; (B) 100 µm.
Maxilliped (
Figure 6F and
Figure 11D) subchelate and composed of syncoxa, basis, and endopod. Syncoxa almost 3.1 times as long as greatest width, constricted in middle of inner margin, ornamented with several rows of spinules, and with three inner setae; proximal seta, arising from distal third, robust and pinnate; other two bipinnate setae inserted in distal corner and near distal sixth of inner margin, respectively. Basis cylindrical, about equal to syncoxa in length, almost four times as long as maximum width, with pinnate seta located on inner surface medially, and ornamented with two inner rows of spinules; posterior spinules much shorter and denser than anterior row. Endopod one-segmented, almost 0.3 times longer and much narrower than preceding segment, about 3.3 times as long as wide, unornamented, and with three armatures (lateral seta arising from inner distal quarter and two apical armatures); apical spine robust, claw-like, and furnished with row of short spinules on inner end; apical seta pinnate and shorter than spine; lateral seta shortest.
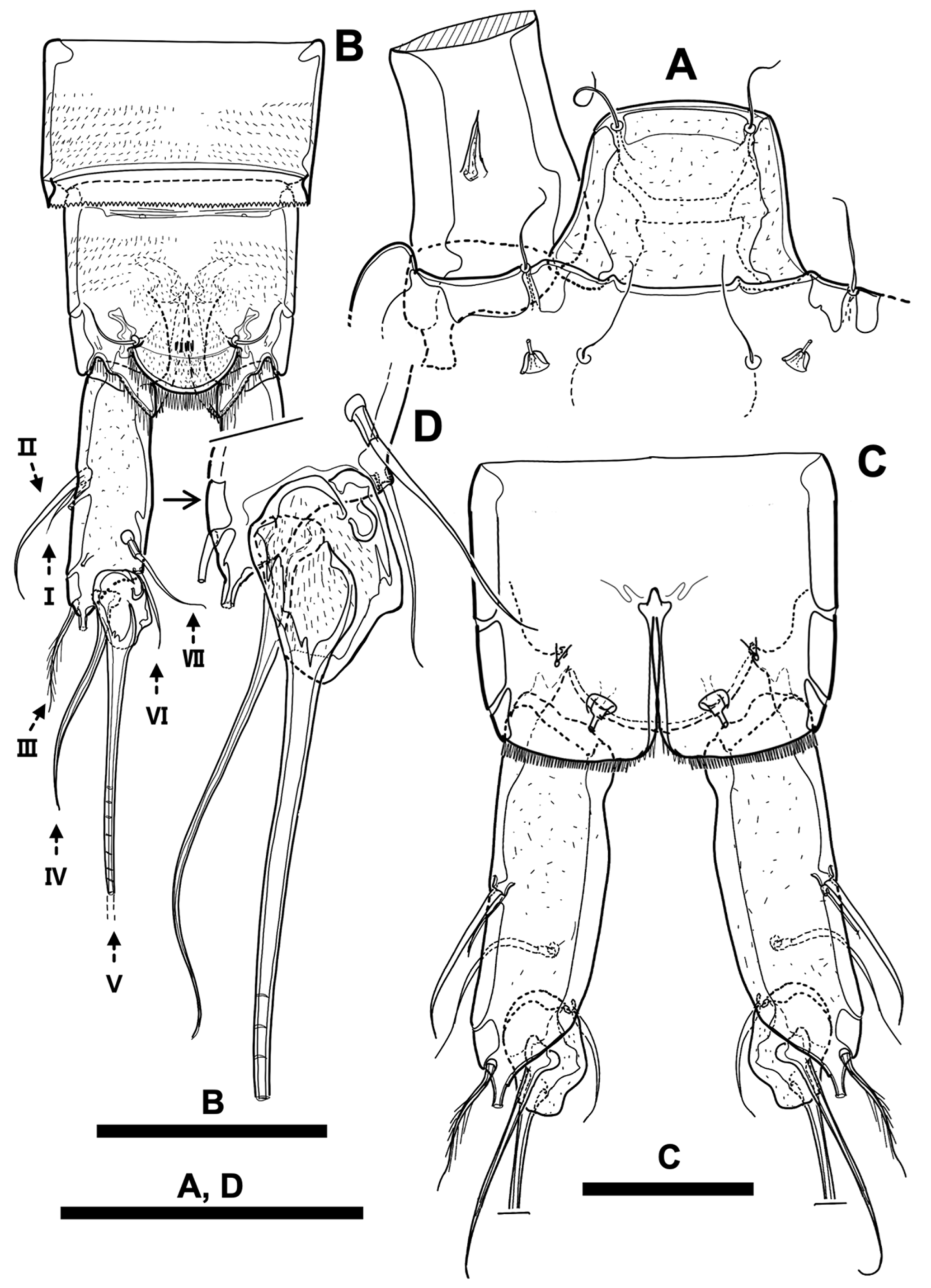
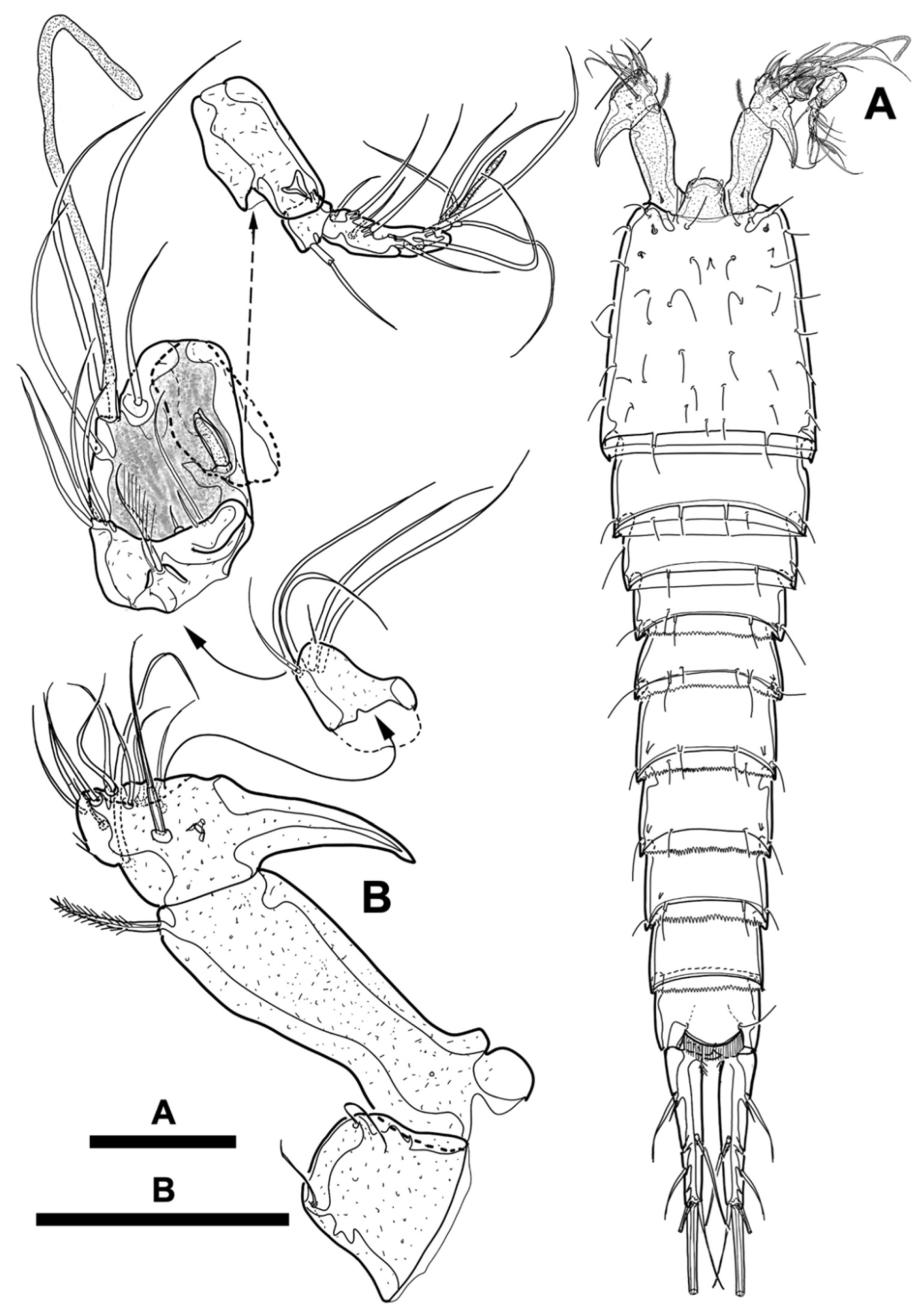
Figure 9.
Line drawings of Phyllopodopsyllus saywakimi sp. nov. female (holotype): (A) urosome excluding first urosomite, ventral; (B) genital field; (C) fifth pereiopod; (D) former baseoendopod of the fifth pereiopod. Scale bars: (A,C) 100 µm; (B,D) 50 µm.
Figure 9.
Line drawings of Phyllopodopsyllus saywakimi sp. nov. female (holotype): (A) urosome excluding first urosomite, ventral; (B) genital field; (C) fifth pereiopod; (D) former baseoendopod of the fifth pereiopod. Scale bars: (A,C) 100 µm; (B,D) 50 µm.
Figure 10.
Line drawings of Phyllopodopsyllus saywakimi sp. nov. female (holotype): (A) antennule, dorsal; (B) antenna, anterior. Scale bar: 50 µm.
Figure 10.
Line drawings of Phyllopodopsyllus saywakimi sp. nov. female (holotype): (A) antennule, dorsal; (B) antenna, anterior. Scale bar: 50 µm.
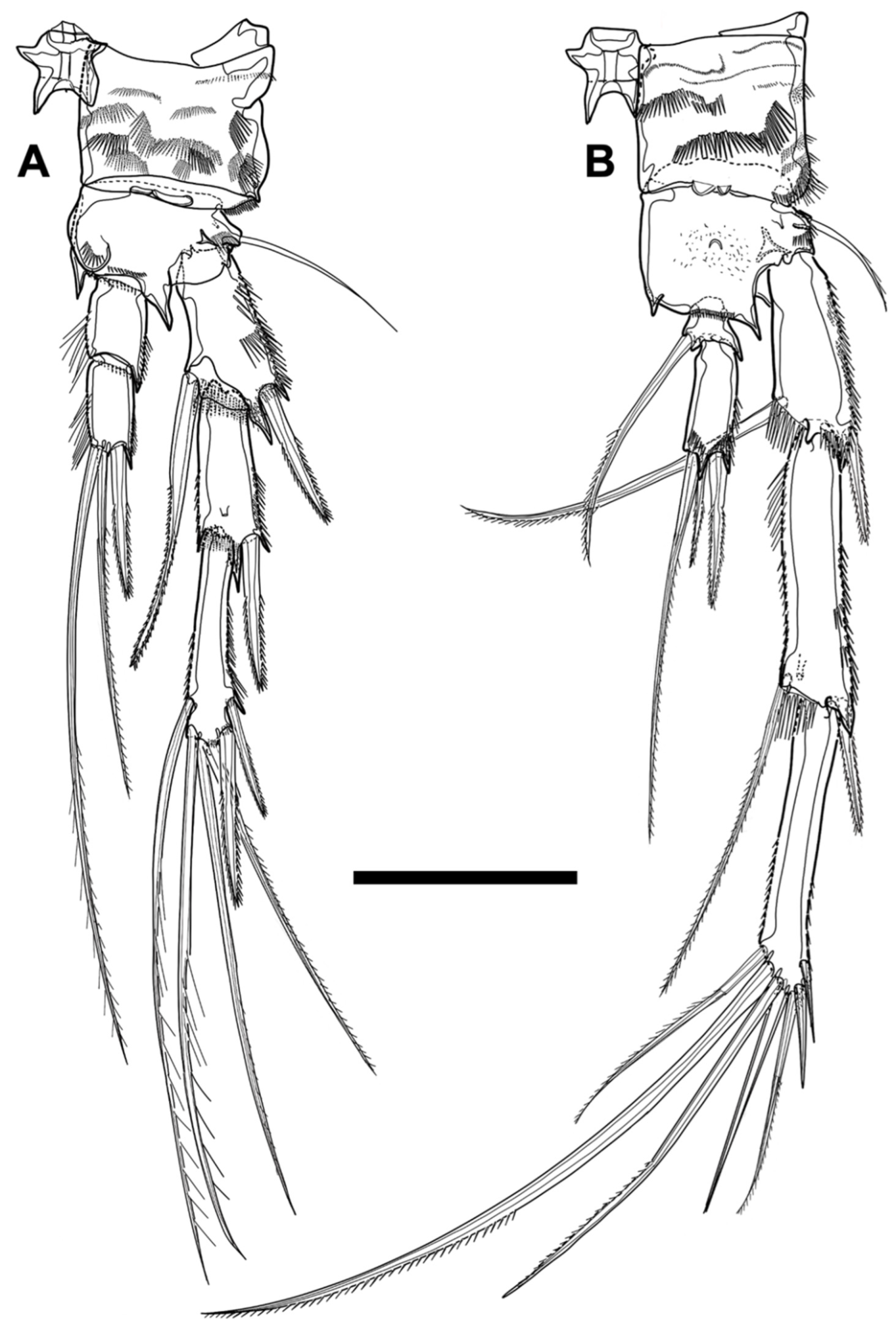
First four pereiopods (P1–P4;
Figure 12 and
Figure 13) biramous, composed of coxa, basis, two-segmented endopod, and three-segmented exopod. Protopod ornamented with numerous rows of various spinules on surface, but pattern of arrangement indefinable. Coxa rectangular and with small intercoxal sclerite connecting rami of each pair; cuticular pore located near outer distal corner, except for P4. Basis transformed into acute integumental structure on both anterior distal corners around endopod, apart from P1, and with cuticular pore on anterior surface near pedestal bearing outer basal seta. Exopod ornamented with rows of spinules or setules along inner and outer margins, but inner row absent on first segment; first two exopodal segments armed with bipinnate outer spine arising from distal corner; first two segments of P2–P4 decorated with frills on inner distal end and with cuticular extension transformed into thorn-like structure anteriorly, around base of outer spine, except for second segment of P4.
Figure 11.
Line drawings of Phyllopodopsyllus saywakimi sp. nov. female (holotype): (A) mandible, anterior; (B) maxillule, posterior; (C) maxilla, anterior; (D) maxilliped, anterior. Scale bar: 50 µm.
Figure 11.
Line drawings of Phyllopodopsyllus saywakimi sp. nov. female (holotype): (A) mandible, anterior; (B) maxillule, posterior; (C) maxilla, anterior; (D) maxilliped, anterior. Scale bar: 50 µm.
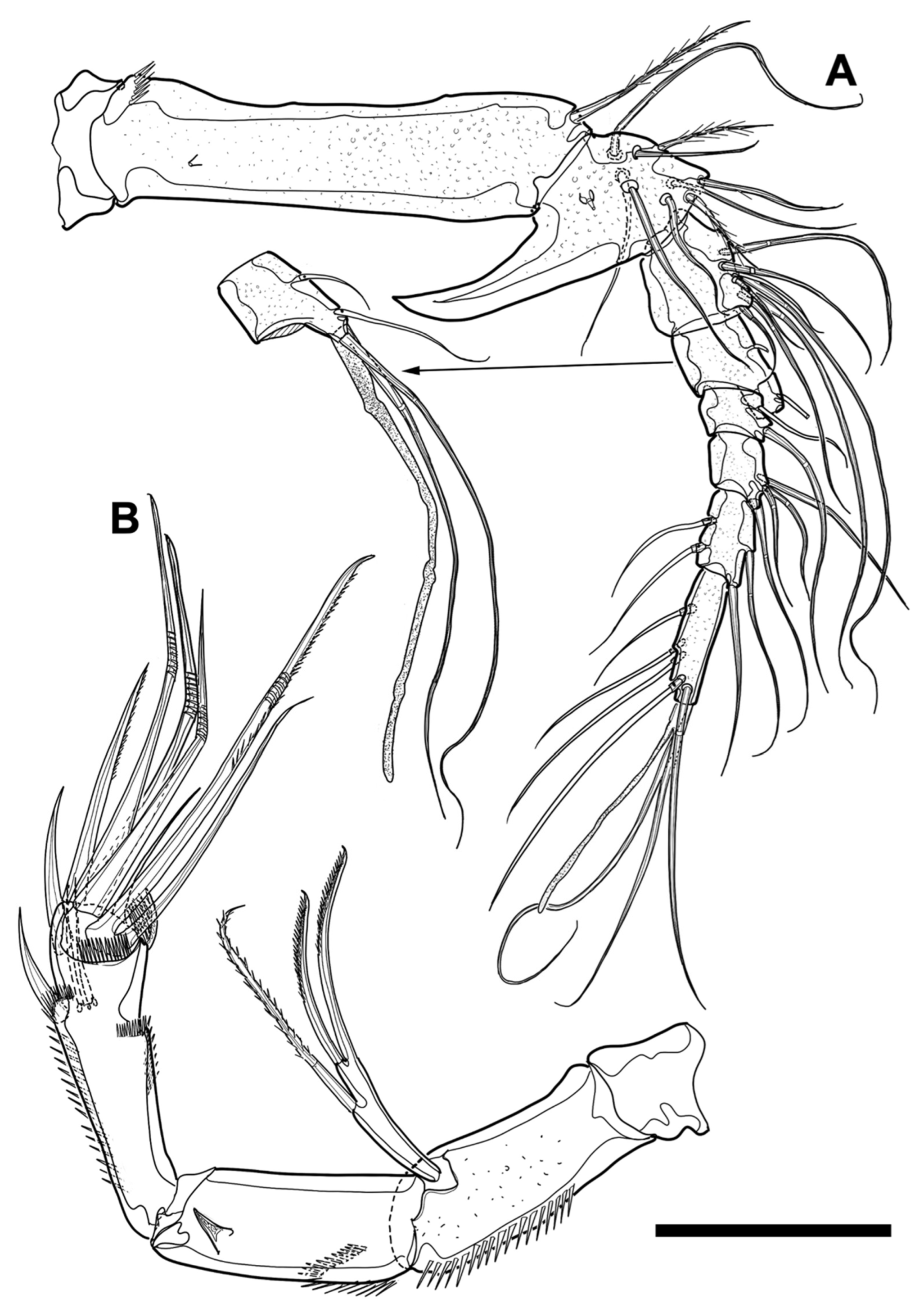
P1 (
Figure 12A). Coxa about 0.9 times as long as greatest width and tapering distally. Basis shorter (about 0.6 times) and narrower (about 0.6 times) than coxa, and with two pedestals bearing inner and outer spine, respectively; inner pedestal ornamented with patch of long and short spinules; inner spine robust, bipinnate, and about as long as basis; outer spine shorter and finer than inner spine; distal margin bearing endopod furnished transversely with row of minute spinules. Endopod prehensile and longer than protopod (about 1.6 times) and exopod (about 1.5 times); first segment elongated, about seven times as long as wide, furnished with spinules along outer margin and setules from inner proximal quarter to inner distal quarter, armed with pinnate inner seta arising from distal quarter; inner seta about two-thirds of first segment in length; second segment much shorter (about 0.3 times) and slightly narrower than first segment, ornamented with row of spinules on outer margin, and armed with two geniculate setae arising from terminal margin; two apical setae much longer than distal endopodal segment and decorated terminally with row of minute spinules. Exopod not reaching distal end of first endopodal segment, without inner seta, and slightly getting shorter gradually (length ratio of three segments from proximal one to distal, 1:0.85:0.7); distal segment with two apical setae and two outer spines; apical setae geniculate and decorated with row of minute spinules terminally, inner of which longer than outer.
P2 (
Figure 12B). Coxa about 1.4 times as broad as length. Basis ornamented with row of long setules on inner margin, shorter (about three quarters), and narrower (about 0.9 times) than coxa; basal spine bipinnate and almost as long as basis. Endopod reaching approximately midlength of distal exopodal segment, about as long as protopod, and shorter than exopod (about 0.6 times); first endopodal segment about twice as long as width, with pinnate seta inserted in inner distal third, and decorated with spinules along inner and outer margins; inner spinules longer and sparser than outer row; second seg-ment much slenderer (about half) and longer (about 1.4 times) than preceding segment, ornamented sparsely with long spinules, and with three terminal setae of which middle seta bare and minute; other two setae pinnate, innermost of which longest and much longer than entire endopod. Exopod tapering distally; length ratio of three exopodal segments from first to third, 1:0.9:0.9; first segment widening distally and with inner robust seta; distal part of inner seta slightly curved and bipinnate terminally; second segment without inner seta; third segment with two outer spines, one apical spine, and one apical seta; apical spine longer than two outer spines; innermost seta slender and about as long as exopod.
P3 (
Figure 13A). Coxa about 1.2 times as broad as length. Basis about 1.5 times as broad as length, shorter (about two-thirds), and narrower (about 0.9 times) than preceding segment; outer basal seta slender and much longer than basis. Endopod not reaching end of second exopodal segment, about 0.9 times as long as protopod, and shorter (about 0.4 times) than exopod; first endopodal segment about 1.5 times as long as greatest width, ornamented with spinules on inner and outer margins and with pinnate seta located in inner distal third; second segment slenderer (about 0.6 times) and longer (about twice) than preceding segment, furnished with spinules along inner and outer margins, and armed with three apical bipinnate armatures; innermost seta longest and much longer than entire endopod; middle seta shortest and thready; outmost armature spine-like. Exopod tapering distally; length ratio of three exopodal segments from proximal to distal, 1:1:1.1; first segment broadening distally and armed with robust seta arising from inner distal quarter; inner seta somewhat bent terminally and bipinnate; second segment without inner seta; third segment with four bipinnate armatures (two outer spines, one apical spine, and one innermost apical seta); apical spine much longer than outer spines; innermost seta about as long as entire exopod.
P4 (
Figure 13B). Coxa about 0.9 times as long as width. Basis shorter (about 0.9 times) than coxa but almost equal in width; pedestal bearing outer seta smaller than two preceding pereiopods; basal seta slender and naked. Endopod about 0.6 times as long as protopod, much shorter (about 0.2 times) than entire exopod, and not reaching midlength of second exopodal segment; first endopodal segment minute, almost as long as wide, and armed with seta arising from inner margin; inner seta slightly longer than entire endopod; second endopodal segment more than five times as long as greatest width, decorated with minute spinules along inner, outer, and terminal margin, and armed with three bipinnate terminal armatures; outermost armature shortest and spine-like; innermost seta longest, longer than entire endopod, and exceeding end of second exopodal segment. Exopod elongate and tapering distally; length ratio of three exopodal segments from first to third, 1:1.3:1.4; first segment cylindrical and with strong pectinate seta originating from about inner distal third; second segment cylindrical and armed with thready inner seta implanted near distal end; third segment with seven pinnate armatures (three inner setae, two terminal setae, and two outer spines); inner setae robust and long, middle of which longest; apical setae slender; outer spines minute.
Figure 12.
Line drawings of Phyllopodopsyllus saywakimi sp. nov. female (holotype): (A) P1; (B) P2. Scale bar: 50 µm.
Figure 12.
Line drawings of Phyllopodopsyllus saywakimi sp. nov. female (holotype): (A) P1; (B) P2. Scale bar: 50 µm.
Figure 13.
Line drawings of Phyllopodopsyllus saywakimi sp. nov. female (holotype): (A) P3; (B) P4. Scale bar: 50 µm.
Figure 13.
Line drawings of Phyllopodopsyllus saywakimi sp. nov. female (holotype): (A) P3; (B) P4. Scale bar: 50 µm.
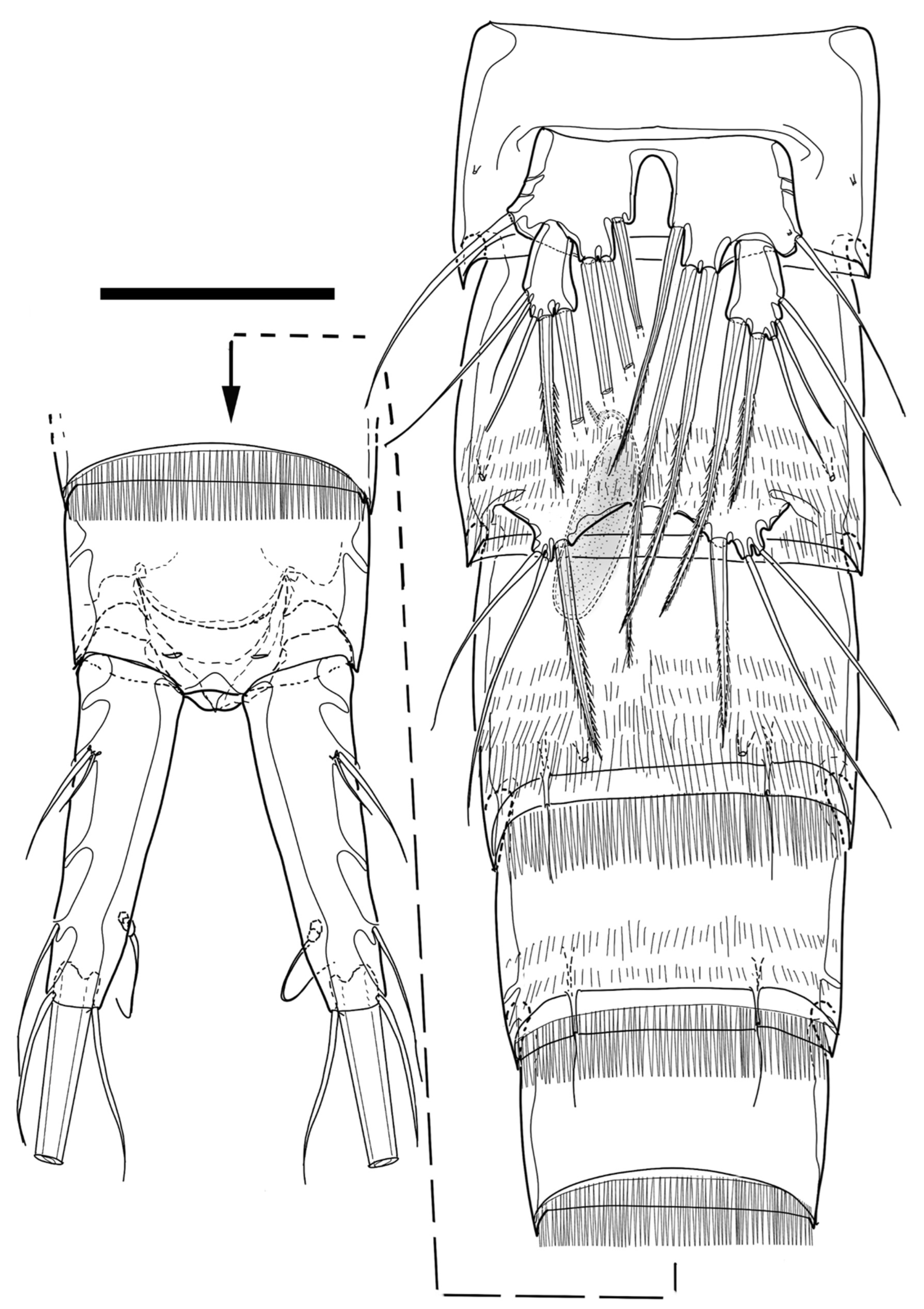
The fifth pereiopod (P5,
Figure 4D and
Figure 9C,D) enlarged, foliaceous, with confluence of baseoendopod and exopod, covered with microspinules and/or microsetules throughout surface, reaching end of third urosomite, formed into brood pouch, and armed with 12 setae (one basal, five baseoendopodal, and six exopodal); baseoendopodal setae located along inner edge; two proximalmost baseoendopodal setae short, bipinnate, and inserted on ventral surface; other three baseoendopodal setae located on small lobe; small baseoendopodal lobe ornamented with row of tiny spinules; two most distal baseoendopodal setae implanted very close to each other in end of lobe, of which proximal seta bipinnate, longest among baseoendopodal setae, and reaching distal end of P5; other distal seta hair-like and minute; basal seta bare and located in near proximal third of outer margin; three of exopodal setae located in distal margin; one of distal setae, arising from ventral corner, minute and bipinnate; central of distal setae much longer than other distal setae; other three exopodal setae located around distal third of dorsal edge; distalmost dorsal seta bipinnate and minute; central of dorsal setae small, bipinnate, and somewhat swollen at base; other dorsal seta about as long as central distal seta.
P6 (
Figure 5A and
Figure 9A,B) P6 (
Figure 5A and
Figure 9A,B) reduced and with three apical setae; outermost seta longest, reaching ventral suture of genital double-somite, and plumose.
Description of adult male (based on paratype 10–19)
Sexual dimorphism appeared in body ornamentation, segmentation of urosome, caudal rami, antennule, and P2–P6. The body ornamentations, such as hyaline frills, cuticular pores, sensilla, (micro)setules, (micro)spinules, and their arrangements are presented in
Figure 2B,
Figure 14A–E,
Figure 15A–E,
Figure 16A–C,
Figure 17A and
Figure 18A.
Figure 14.
Scanning electron microscopy images of Phyllopodopsyllus saywakimi sp. nov. male (paratype 11) in dorsal view: (A) habitus; (B) cephalothorax; (C) second prosomite to the second urosomite; (D) third to fifth urosomites; (E) ana somite and caudal rami; (F) antennule. Scale bars: 50 µm.
Figure 14.
Scanning electron microscopy images of Phyllopodopsyllus saywakimi sp. nov. male (paratype 11) in dorsal view: (A) habitus; (B) cephalothorax; (C) second prosomite to the second urosomite; (D) third to fifth urosomites; (E) ana somite and caudal rami; (F) antennule. Scale bars: 50 µm.
Urosome (
Figure 2B,
Figure 14C–E,
Figure 15D,E,
Figure 16A–C,
Figure 17A and
Figure 18A,B) six-segmented and tapering gradually. First urosomite cylindrical and with smooth hyaline frill ventrally. Genital and first abdominal somite separated; genital somite armed with P6 and longest among urosomites; three free abdominal somites getting shorter posteriorly (length ratio of third to fifth urosomite in lateral aspect, 1:0.9:0.8) and with regularly spinulose hyaline frills ventrally. Caudal rami elongated, about 5.4 times as long as greatest width, with dimples on dorsolateral surface, covered with microspinules throughout surface, tapering from midlength to distal, and armed with seven setae (three lateral, three apical, and one dorsal); setae I and II, arising from proximal third, naked and inserted very close to each other dorsoventrally; seta I minute, hair-like, and less than one-tenth of seta II in length; seta III implanted in distal quarter and about as long as seta II; setae IV and VI slender and almost equal in length; seta V longest and robust; seta VII inserted in about distal third of dorsal surface and triarticulate.
Figure 15.
Scanning electron microscopy images of Phyllopodopsyllus saywakimi sp. nov. male (paratype 12) in lateral view: (A) habitus; (B) cephalothorax; (C) second to fourth prosomites; (D) first to third urosomites, P5, and P6; (E) fourth urosomite to anal somite and caudal rami; (F) antennule. Scale bars: (A) 100 µm; (B–D) 30 µm; (E,F) 20 µm.
Figure 15.
Scanning electron microscopy images of Phyllopodopsyllus saywakimi sp. nov. male (paratype 12) in lateral view: (A) habitus; (B) cephalothorax; (C) second to fourth prosomites; (D) first to third urosomites, P5, and P6; (E) fourth urosomite to anal somite and caudal rami; (F) antennule. Scale bars: (A) 100 µm; (B–D) 30 µm; (E,F) 20 µm.
Antennule (
Figure 14F,
Figure 15F,
Figure 16D and
Figure 17A) seven-segmented, subchirocer, covered with dimples dorsally, and with geniculation between fourth and fifth segments; first segment longest, with cuticular pore on dorsal surface proximally, ornamented with row of spinules near dorsal pore, and armed with pinnate seta inserted in inner distal corner; second segment with integumental process as female; third segment small; fourth segment enlarged, shorter than all followings combined, and armed with aesthetasc fused basally to seta; sixth segment shortest; fourth to sixth segments modified into pincer-like structure with several cuticular ridges; seventh segment with short aesthetasc fused with two setae at base; most of setae arising from distal segment biarticulate. Armature formula as follow: 1(1), 2(11), 3(6), 4(13+(1+ae)), 5(1), 6(1), 7(9+(2+ae)).
It was difficult to determine whether: (a) the small lobe bearing two setae (marked by an empty arrow in
Figure 14F) was a part of the third or the fourth segment; (b) the flame-like structures (marked by a hatched arrow in
Figure 14F) on the fourth segment and the cuticular process (marked by a checkered arrow in
Figure 14F) on the fifth segment were the transformed armatures or the integumental ridges. The above descriptions and formula were written, assuming they were a part of the fourth segment and the transformed armatures, respectively.
Figure 16.
Scanning electron microscopy images of Phyllopodopsyllus saywakimi sp. nov. male (paratype 13) in ventral view: (A) second to third urosomites, P5, and P6; (B) fourth urosomite to anal somite; (C) caudal rami; (D) antennule. Scale bars: 30 µm.
Figure 16.
Scanning electron microscopy images of Phyllopodopsyllus saywakimi sp. nov. male (paratype 13) in ventral view: (A) second to third urosomites, P5, and P6; (B) fourth urosomite to anal somite; (C) caudal rami; (D) antennule. Scale bars: 30 µm.
P2 (
Figure 19A) as in female excluding the following: Endopod reaching, at least, middle part of third exopodal segment; innermost seta of second endopodal segment shortest and hairy; middle seta of second endopodal segment longest.
P3 (
Figure 19B) as in female excluding the following: Second endopodal segment longer (about 1.5 times) than preceding segment; outermost armature of second endopodal segment spine-like and shortest; other two setae inserted very close to each other.
P4 (
Figure 19C) as in female excluding the following: Second endopodal segment about 4.4 times as long as greatest width, about 3.5 times longer than preceding segment, and armed with two pinnate spine-like armatures, of which outer armature much shorter. Distal segment of exopod with two inner setae, one apical seta, one apical spine, and two outer spines; two inner setae robust, pinnate, and much longer than other armatures.
Figure 17.
Line drawings of Phyllopodopsyllus saywakimi sp. nov. male (paratype 14): (A) habitus, dorsal; (B) rostrum and antennule, dorsal. Scale bar: (A) 100 µm; (B) 50 µm.
Figure 17.
Line drawings of Phyllopodopsyllus saywakimi sp. nov. male (paratype 14): (A) habitus, dorsal; (B) rostrum and antennule, dorsal. Scale bar: (A) 100 µm; (B) 50 µm.
Figure 18.
Line drawings of Phyllopodopsyllus saywakimi sp. nov. male (paratype 14): (A) urosome (excluding first urosomite); (B) caudal ramus; (C) P5; (D) P6. Scale bars: (A) 100 µm; (B–D) 50 µm.
Figure 18.
Line drawings of Phyllopodopsyllus saywakimi sp. nov. male (paratype 14): (A) urosome (excluding first urosomite); (B) caudal ramus; (C) P5; (D) P6. Scale bars: (A) 100 µm; (B–D) 50 µm.
P5 (
Figure 18C) not foliaceous and composed of baseoendopod and exopod; both rami fused at central part completely. Baseoendopod large, with cuticular pore near base of basal seta, and armed with three pinnate setae on endopodal lobe; basal seta as long as P5 and slender; median seta longest among endopodal armatures; innermost seta bipinnate terminally. Exopod about twice as long as greatest width, shorter than baseoendopod, and armed with five setae; two innermost setae bipinnate; other setae slender and bare.
Figure 19.
Line drawings of Phyllopodopsyllus saywakimi sp. nov. male (paratype 14): (A) P2; (B) endopod of P3; (C) P4. Scale bar: 50 µm.
Figure 19.
Line drawings of Phyllopodopsyllus saywakimi sp. nov. male (paratype 14): (A) P2; (B) endopod of P3; (C) P4. Scale bar: 50 µm.
P6 (
Figure 18A,D) reduced into single plate, fused at base with second urosomite completely, and armed with three setae; outermost seta slender, shortest, and bare; meddle seta slender and naked; innermost seta longest, bipinnate, and robust.
Variability
The body length ranged from about 807.6–856.1 µm (mean = 829.3 µm,
n = 3) in females (
Figure S1) and about 554.6–640.8 µm (mean = 589.5 µm,
n = 5) in males (
Figure S2).
3.3. Phyllopodopsyllus similis Kim & Lee, 2023 sp. nov.
Figure 20,
Figure 21,
Figure 22,
Figure 23,
Figure 24,
Figure 25,
Figure 26,
Figure 27,
Figure 28,
Figure 29,
Figure 30,
Figure 31,
Figure 32,
Figure 33,
Figure 34,
Figure 35,
Figure 36 and
Figure 37 and
Figures S5–S12.
Zoobank registration
urn:lsid:zoobank.org:act:C0F18E60-A5B5-4672-BA24-0E85FF001B61
Type locality
Intertidal sand of the rocky shore, Udo, Republic of Korea, 23 July 2020. (L3: 33°31′14.53″ N 126°57′29.64″ E)
Other localities
The sandy habitat around the tidal pool near Daepyeong Pier in Jeju, Korea, 26 July 2020 (L1: 33°14′11.54″ N 126°21′35.56″ E); the intertidal sand near Jeju Ocean and Fisheries Research Institute in Jeju, Republic of Korea, 24 July 2020 (L2: 33°18′34.782″ N 126°49′53.328″ E); the intertidal sand of the rocky shore, Busan, Republic of Korea, 25 August 2022 (L4: 35°10′17.61″ N 129°11′52.68″ E).
Specimens examined
Holotype: 1 female (MABIKCR00252866)
Paratype: 20 females and 14 males (NIBRIV0000901840–NIBRIV0000901873). The voucher specimen information is given in detail in
Table S1.
Etymology
The specific name similis is derived from the Latin adjective similis, meaning “similar”. It alludes to the similarity between the new species and the widely distributed species, Phyllopodopsyllus aegypticus Nicholls, 1944.
Description of adult female (based on Holotype and Paratype 1–20)
Body (
Figure 20A,
Figure 21A–D,
Figure 22A–E,
Figure 23,
Figure 24A,
Figure 25,
Figure 26A,B and
Figures S5–S7) nine-segmented, cylindrical, slightly constricted in middle, furnished with sensilla, cuticular pores, (micro)spinules, and/or (micro)setules (details in figures), and without dimples on all body surface except for anal somite.
Prosome (
Figure 20A,
Figure 21A,B,
Figure 22A–C,
Figure 24A,
Figure 25,
Figures S5A,B,D, S6A–C, S8A, S9A, and S10A) four-segmented, slightly tapering distally, and composed of cephalothorax and three free pedigerous somites. Cephalothorax somewhat longer than all succeeding prosomites combined. Three free pedigerous somites nearly equal in length. Hyalin frills of prosomites smooth except for fourth somite; dorsal frills of last prosomite serrated but inconspicuous. Rostrum small, weakly defined at base, triangular in dorsal aspect, and with two sensilla near apex. Eye not visible.
Urosome (
Figure 20A,
Figure 21A,C,D,
Figure 22A,C–E,
Figure 23,
Figure 25,
Figure 26A–C,
Figures S5A,C, S6A,D, S7, and S8E,F) five-segmented, about as long as prosome (including caudal rami), and composed of fifth pedigerous somite, genital double-somite, two abdominal somites, and anal somite armed with caudal rami. Fifth pedigerous somite shortest and wrinkled on ventral surface; hyaline frill of fifth pedigerous somite finely serrated dorsally and smooth laterally. Genital double-somite longest among urosomites, armed with P6, and with vestigial original segmentation marked by suture and pattern of surface ornamentation; hyaline frill of genital double-somite finely fringed dorsally but smooth laterally and ventrally; lateral frill ornamented with spinules regularly; genital field with single copulatory pore near ventral suture, short copulatory duct in distal part of genital somite, and two seminal receptacles. Antepenultimate somite slightly longer than first urosomite; hyaline frill of antepenultimate somite finely fringed dorsally; ventral part of frill smooth and ornamented with long spinules regularly on terminal margin. Penultimate somite about as long as first urosomite and finely fringed dorsally; ventral part of frill smooth and ornamented with regular spinules terminally. Anal somite about as long as antepenultimate somite, dimpled, cleft medially in posterior part; anal opening located in cleft; anal operculum dimpled, rounded, and ornamented densely with spinules on terminal margin. Caudal rami subconical, about 1.8 times as long as greatest width, dimpled laterally, swelled in proximal part of inner margin dorsally, furnished with one bunch of long setules near inner bulge, and armed with seven naked setae on each ramus, (three outer, one dorsal, and three terminal); setae I and II set very close to each other dorsoventrally at proximal third of outer margin; seta I tiny, thready, and about one-fourth as long as seta II; seta III inserted near outer distal quarter; seta V about as long as urosomite; setae IV and VI slender and equal in length; seta VII triarticulate and implanted in inner distal corner.
Figure 20.
Confocal laser scanning microscopy images of Phyllopodopsyllus similis sp. nov. (A) female (paratype 1): habitus, lateral; (B) male (paratype 21): habitus, lateral. Scale bars: 100 µm.
Figure 20.
Confocal laser scanning microscopy images of Phyllopodopsyllus similis sp. nov. (A) female (paratype 1): habitus, lateral; (B) male (paratype 21): habitus, lateral. Scale bars: 100 µm.
Antennule (
Figure 21E,
Figure 22F,
Figure 24B,
Figure 27A,
Figures S5D, S6B, S8B, S9B, S10B, and S11A) nine-segmented and covered with dimples; first segment elongated, longest, about three times longer than maximum width, about 0.6 times as long as all succeeding segments combined, ornamented with row of tiny spinules on proximal inner margin, and with pinnate seta on inner distal corner; second segment without protuberance and with cuticular pore on dorsal surface medially; fourth segment with pedestal armed with aesthetasc fused basally to long seta on inner distal corner; eighth segment shortest; ninth segment about 3.5 times as long as wide and with apical aesthetasc fused basally to two setae. Armature formula as follows: 1(1), 2(9), 3(8), 4(3+(ae+1)), 5(2), 6(4), 7(2), 8(2), 9(5+(ae+2)); all of setae naked and slender, except for first two segments; all outer setae arising from antepenultimate to distal segments biarticulate.
Antenna (
Figure 22F,
Figure 24B,
Figure 27B,
Figures S8B, S9B, S10B, and S11A) composed of coxa, basis, endopod, and exopod. Coxa bare and about as long as greatest width. Basis unarmed, ornamented with longitudinal row of spinules along abexopodal margin, and about 2.5 times as long as wide. Endopod two-segmented; first endopodal segment bare, about as long as basis, about 2.7 times long as greatest width; second endopodal segment about as long as preceding segment, about 2.7 times as long as greatest width, decorated with patch of spinules on inner margin, row of strong spinules terminally, and two rows of frills on outer margin and armed with nine armatures (six apical and three lateral); one bare, one pinnate, and four geniculate setae located apically; three lateral armatures composed of two robust inner spines and one slender seta; one of inner spines located near distal corner and other spine inserted in distal third; lateral seta implanted between outer spines. Exopod one-segmented and with three pinnate setae (two apical and one lateral); outer apical seta completely fused basally to exopod.
Figure 21.
Scanning electron microscopy images of Phyllopodopsyllus similis sp. nov. female (paratype 2) in dorsal view: (A) habitus; (B) prosome; (C) urosome; (D) anal somite and caudal rami; (E) antennule; (F) atypical antennule. Scale bars: (A) 100 µm; (B–F) 30 µm.
Figure 21.
Scanning electron microscopy images of Phyllopodopsyllus similis sp. nov. female (paratype 2) in dorsal view: (A) habitus; (B) prosome; (C) urosome; (D) anal somite and caudal rami; (E) antennule; (F) atypical antennule. Scale bars: (A) 100 µm; (B–F) 30 µm.
Labrum (
Figure 24C,
Figures S8C, S9C, S10C, and S11B) well-developed, gradually tapering ventrally, decorated with patch of long setules along posterior margin distally, and covered with long and slender setules throughout cutting edge; anterior and lateral surfaces dimpled
Mandible (
Figure 24C,
Figure 28A,
Figures S10C, and S11B) composed of coxa, basis, one-segmented endopod, and one-segmented exopod. Coxa enlarged and with gnathobase bearing pinnate dorsal seta and row of multicuspidate teeth. Basis gradually widening distally, pentagonal, shorter than coxa, furnished with longitudinal rows of long spinules on anterior surface, and armed with three pinnate setae originating from terminal margin. Endopod much shorter than basis and armed with two lateral setae and six apical bare setae. Exopod slightly narrower than endopod, about as long as endopod, and with five setae (three lateral and two apical); proximal-most of lateral setae plumose and long; middle seta naked, short, and thready; distal-most lateral seta naked; two apical setae plumose and long.
Figure 22.
Scanning electron microscopy images of Phyllopodopsyllus similis sp. nov. female (paratype 3) in lateral view: (A) habitus; (B) cephalothorax; (C) second prosomite to first urosomite; (D) P5 and genital double-somite; (E) third urosomite to caudal rami; (F) antennule and antenna. Scale bars: (A) 100 µm; (B–F) 30 µm.
Figure 22.
Scanning electron microscopy images of Phyllopodopsyllus similis sp. nov. female (paratype 3) in lateral view: (A) habitus; (B) cephalothorax; (C) second prosomite to first urosomite; (D) P5 and genital double-somite; (E) third urosomite to caudal rami; (F) antennule and antenna. Scale bars: (A) 100 µm; (B–F) 30 µm.
Paragnaths (
Figure 24C,
Figures S8C, S9C, and S10C) well-developed and composed of posterior and two lateral lobes; cutting edge of each lobe wrinkled; lateral lobes ornamented with several rows of long spinules on each inner side; posterior lobe decorated with fine setules around median wrinkles.
Figure 23.
Scanning electron microscopy images of Phyllopodopsyllus similis sp. nov. female (paratype 4) in ventral view: (A) genital double-somite; (B) third and fourth urosomites; (C) anal somite and caudal rami. Scale bars: 30 µm.
Figure 23.
Scanning electron microscopy images of Phyllopodopsyllus similis sp. nov. female (paratype 4) in ventral view: (A) genital double-somite; (B) third and fourth urosomites; (C) anal somite and caudal rami. Scale bars: 30 µm.
Figure 24.
Scanning electron microscopy images of Phyllopodopsyllus similis sp. nov. female (paratype 4) in ventral view: (A) cephalothorax; (B) antennule and antenna; (C) mouth parts around oral opening; (D) maxilliped. Scale bars: (A) 50 µm; (B) 30 µm; (C,D) 20 µm.
Figure 24.
Scanning electron microscopy images of Phyllopodopsyllus similis sp. nov. female (paratype 4) in ventral view: (A) cephalothorax; (B) antennule and antenna; (C) mouth parts around oral opening; (D) maxilliped. Scale bars: (A) 50 µm; (B) 30 µm; (C,D) 20 µm.
Maxillule (
Figure 24C,
Figure 28B,
Figures S8C, S9C, S10C and S11B) composed of praecoxa, coxa, basis, endopod, and exopod. Praecoxa large, trapezoidal, ornamented with row of spinules on distal half of outer margin, and armed with well-developed arthrite; praecoxal arthrite armed with 13 elements and ornamented with row of spinules; two setae located in near posterior margin; one apical pinnate seta inserted in distal corner; two bare setae implanted in anterior surface; eight robust armatures arranged around distal margin. Coxa about half as long as praecoxa, with cylindrical endite incorporated into coxa, and armed with epipodite represented by one plumose seta; coxal endite with four setae. Basis unornamented and armed with six setae apically. Endopod about twice as long as greatest width, and with four plumose setae; three endopodal setae originating around terminal margin, and one seta inserted in inner margin subapically. Exopod broader than endopod, about as long as greatest width, ornamented with setules along inner margin, and with three plumose setae located apically.
Maxilla (
Figure 24C,
Figure 28C,
Figures S8C, S9C and S11B) composed of syncoxa, basis, and endopod. Syncoxa large and armed with four endites; endites with two, one, three, and three pinnate and/or plumose setae from proximal to distal, respectively; proximal second endite shortest; other endites almost equal in length; Basis approximately half as long as syncoxa and armed with cylindrical endite bearing one slender plumose seta arising from posterior surface distally and two robust setae apically; Endopod two-segmented and much shorter than basis; first endopodal segment about 0.2 times as long as greatest width and armed with three setae (two robust pinnate setae and one slender bare seta); second endopodal segment about as long as preceding segment and armed with four setae apically (two slender naked setae and two pinnate robust setae).
Maxiliped (
Figure 24D,
Figure 28D,
Figures S8D, S9D, S10D and S11B) subchelate and composed of syncoxa, basis, and endopod. Syncoxa about twice as long as greatest width, constricted in middle of inner margin, ornamented with several rows of spinules and setules, and with three inner setae; proximal-most seta, arising from distal third, robust and pinnate; central inner seta plumose; distal-most seta pinnate and located near distal corner. Basis cylindrical, almost as long as preceding segment, about 2.8 times as long as greatest width, armed with plumose seta inserted in inner margin medially, and ornamented with two rows of spinules along inner margin; posterior spinules much shorter and denser than anterior row. Endopod one-segmented, much narrower and shorter (about 0.3 times) than preceding segment, about 2.7 times as long as maximum width, unornamented, with claw-like spine originating from terminal margin; terminal spine robust and pinnate on inner end.
P1–P4 (Figure 29 and
Figure 30) biramous, composed of coxa, basis, two-segmented endopod, and three-segmented exopod. Protopod ornamented with several rows of spinules on surface but arrangement pattern indefinable. Coxa rectangular and with small intercoxal sclerite connecting rami of each pair; cuticular pore located near outer distal corner except for P4. Basis transformed into acute integumental structure on both anterior distal corners around endopod apart from P1 and with cuticular pore on anterior surface near pedestal bearing outer basal armature. Exopod ornamented with row of spinules or setules along outer margin of each segment; first two exopodal segments armed with bipinnate outer spine originating from distal corner; first two segments of P2–P4 decorated with frills on inner distal end and with cuticular extension transformed into thorn-like structure, anteriorly, near base of outer spine.
P1 (
Figure 29A). Coxa about 0.6 times as long as greatest width and somewhat tapering distally. Basis shorter (about two-thirds) and narrower (about 0.6 times) than coxa, and with two bipinnate spines originating from inner and outer margins, respectively; base of inner spine ornamented with patch of long and short spinules; inner spine robust and longer than basis; distal margin bearing endopod furnished with row of spinules, transversely. Endopod longer than protopod (about 1.7 times); first segment elongated, about six times as long as greatest width, furnished with spinules and setules along outer and inner margins, respectively, and armed with pinnate inner seta arising from distal third; inner seta about half of first segment in length; second segment much shorter (about 0.2 times) and slightly narrower than preceding segment, ornamented with row of spinules on anterior surface, and with two geniculate setae apically; two apical setae much longer than distal endopodal segment, and pinnate terminally. Exopod reaching about midlength of first endopodal segment (about 0.7 times) and without inner seta; length ratio of three exopodal segments from proximal to distal, 1:0.8:0.8; distal segment with two apical setae and two outer spines; apical setae geniculate and pinnate terminally; inner apical seta longer than outer.
Figure 25.
Line drawings of Phyllopodopsyllus similis sp. nov. female (holotype): (A) habitus; dorsal; (B) habitus, lateral. Scale bar: 100 µm.
Figure 25.
Line drawings of Phyllopodopsyllus similis sp. nov. female (holotype): (A) habitus; dorsal; (B) habitus, lateral. Scale bar: 100 µm.
P2 (
Figure 29B). Coxa almost as long as wide. Basis ornamented with row of long setules along inner margin, shorter (about half), and narrower (about 0.9 times) than coxa; basal spine bipinnate and longer than basis. Endopod not reaching end of second exopodal segment, about 0.7 times as long as protopod, and shorter than exopod (about 0.4 times); first endopodal segment unarmed, about 1.3 times as long as greatest width, and decorated with spinules along inner and outer margins; inner spinules longer and sparser than outer spinules; second segment narrower (about 0.7 times) than preceding segment but almost equal in length, ornamented with long spinules, and with three terminal armatures; innermost seta longest; outermost armature spine-like and inserted deeper than other two setae. Exopod tapering distally; length ratio of three exopodal segments from first to third, 1:1:1.2; first segment widening distally and armed with inner bipinnate seta; second segment unarmed; third segment with five pinnate armatures (two outer spines, one apical spine, one apical seta, and one inner seta); apical spine much longer than outer spines.
Figure 26.
Line drawings of Phyllopodopsyllus similis sp. nov. female (holotype): (A) genital double-somite, ventral; (B) anal somite and caudal rami, dorsal; (C) caudal ramus, ventral; (D) P5. Scale bars: 50 µm.
Figure 26.
Line drawings of Phyllopodopsyllus similis sp. nov. female (holotype): (A) genital double-somite, ventral; (B) anal somite and caudal rami, dorsal; (C) caudal ramus, ventral; (D) P5. Scale bars: 50 µm.
Figure 27.
Line drawings of Phyllopodopsyllus similis sp. nov. female (holotype): (A) rostrum and antennule, dorsal; (B) antenna, anterior. Scale bars: 50 µm.
Figure 27.
Line drawings of Phyllopodopsyllus similis sp. nov. female (holotype): (A) rostrum and antennule, dorsal; (B) antenna, anterior. Scale bars: 50 µm.
P3 (
Figure 30A). Coxa almost as long as greatest width. Basis about half as long as wide, shorter (about 0.5 times) and narrower (about 0.9 times) than coxa; outer basal seta slender, naked, and much longer than basis. Endopod reaching midlength of second exopodal segment, about 0.7 times as long as protopod, and shorter than exopod (about 0.4 times); first endopodal segment unarmed, about 1.4 times as long as greatest width, ornamented with spinules on inner and outer margins; second segment slenderer (about 0.7 times) than first segment but almost equal in length and armed with three bipinnate armatures terminally; innermost seta longest and much longer than entire endopod; outmost armature spine-like and inserted deeper than other apical setae. Exopod tapering distally; length ratio of three exopodal segments from proximal to distal, 1:1:1.3; first segment broadening distally and armed with robust seta arising from inner distal corner; inner seta somewhat bent terminally and bipinnate; second segment unarmed; third segment with six pinnate armatures (two outer spines, two apical setae, and two inner setae).
Figure 28.
Line drawings of Phyllopodopsyllus similis sp. nov. female (holotype): (A) mandible; (B) maxillule; (C) maxilla; (D) maxilliped. Scale bar: 50 µm.
Figure 28.
Line drawings of Phyllopodopsyllus similis sp. nov. female (holotype): (A) mandible; (B) maxillule; (C) maxilla; (D) maxilliped. Scale bar: 50 µm.
Figure 29.
Line drawings of Phyllopodopsyllus similis sp. nov. female (holotype): (A) P1; (B) P2. Scale bars: 50 µm.
Figure 29.
Line drawings of Phyllopodopsyllus similis sp. nov. female (holotype): (A) P1; (B) P2. Scale bars: 50 µm.
P4 (
Figure 30B). Coxa about 1.1 times as broad as length. Basis shorter (about 0.8 times) than coxa but almost equal in maximum width; pedestal bearing outer basal seta smaller than ped-estals of two preceding pereiopods; basal seta slender and naked. Endopod about half as long as protopod, much shorter than exopod (about 0.2 times), and not reaching midlength of second exopodal segment; first endopodal segment minute, about 0.9 times as long as wide, and armed with inner pinnate seta; inner seta of first endopodal segment much longer than entire endopod; second endopodal segment about 2.4 times as long as greatest width and armed with three bipinnate terminal armatures; outermost spine-like and inserted deeper than other terminal setae; innermost seta longest, much longer than entire endopod, and exceeding end of second exopodal segment. Exopod elongated and tapering distally; length ratio of three exopodal segments from first to third segment, 1:1.4:1.4; first segment cylindrical and with pinnate seta originating from inner distal corner; second segment cylindrical and armed with long inner seta implanted near distal end; third segment with seven armatures (three inner setae, two terminal setae, and two outer spines); inner setae robust and pinnate terminally; middle one of inner setae longest; outer spines minute.
Figure 30.
Line drawings of Phyllopodopsyllus similis sp. nov. female (holotype): (A) P3; (B) P4. Scale bars: 50 µm.
Figure 30.
Line drawings of Phyllopodopsyllus similis sp. nov. female (holotype): (A) P3; (B) P4. Scale bars: 50 µm.
P5 (
Figure 22D and
Figure 26D) enlarged, foliaceous, formed into brood pouch, with confluence of baseoendopod and exopod, not reaching end of third urosomite, ornamented with microspinules and/or microsetules on posterior surface and dorsal margin, and armed with 12 setae (one basal, five baseoendopodal, six exopodal); baseoendopodal setae located along inner edge; two proximal-most baseoendopodal setae bipinnate; median seta plumose; two distal-most setae implanted very close to each other; distal-most baseoendopodal seta hair-like and minute; basal seta bare and inserted in proximal part of dorsal edge; exopodal setae located around from distal third of dorsal edge to distal corner; distal-most exopodal seta pinnate; second proximal seta pinnate, short, and some what swollen at base; other exopodal setae bare; second distal seta longest among exopodal setae and exceeding end of antepenultimate urosomite.
P6 (
Figure 26A,
Figures S7A, and S8E) reduced, longer than maximum width, and armed with two apical setae and one inner seta; all setae plumose and almost equal in length.
Description of adult male (based on paratype 21–34)
Sexual dimorphism appeared in the body ornamentation, segmentation of urosome, caudal rami, antennule, and P2–P6. The body ornamentations, such as hyaline frills, cuticular pores, sensilla, (micro)setules, and (micro)spinules, and their arrangements, were presented in
Figure 20B,
Figure 31A–C,
Figure 32A–E,
Figure 33A–C,
Figure 34,
Figure 35 and
Figure S12A.
Figure 31.
Scanning electron microscopy images of Phyllopodopsyllus similis sp. nov. male (paratype 22) in dorsal view: (A) habitus; (B) prosome; (C) urosome; (D) antennule. Scale bars: (A) 100 µm; (B,C) 30 µm; (D) 20 µm.
Figure 31.
Scanning electron microscopy images of Phyllopodopsyllus similis sp. nov. male (paratype 22) in dorsal view: (A) habitus; (B) prosome; (C) urosome; (D) antennule. Scale bars: (A) 100 µm; (B,C) 30 µm; (D) 20 µm.
Figure 32.
Scanning electron microscopy images of Phyllopodopsyllus similis sp. nov. male (paratype 23) in lateral view: (A) habitus; (B) cephalothorax; (C) second prosomite to the second urosomite; (D) third to fifth urosomites; (E) anal somite and caudal rami; (F) antennule. Scale bars: (A) 100 µm; (B–D) 30 µm; (E,F) 20 µm.
Figure 32.
Scanning electron microscopy images of Phyllopodopsyllus similis sp. nov. male (paratype 23) in lateral view: (A) habitus; (B) cephalothorax; (C) second prosomite to the second urosomite; (D) third to fifth urosomites; (E) anal somite and caudal rami; (F) antennule. Scale bars: (A) 100 µm; (B–D) 30 µm; (E,F) 20 µm.
Urosome (
Figure 31C,
Figure 32C–E,
Figure 33A–C,
Figure 34,
Figure 35 and
Figure S12A) six-segmented and tapering gradually. First urosomite cylindrical and with smooth hyaline frill ventrally. Genital and first abdominal somite separated; genital somite armed with P6; three free abdominal somites getting shorter posteriorly (length ratio from third to fifth urosomite in lateral aspect, 1:0.9:0.7) and regularly ornamented with spinules along terminal margin. Caudal rami elongated, about 3.3 times as long as greatest width, with dimples on dorsal surface, covered with fine microsetules throughout surface, tapering distally, and armed with seven setae (one dorsal, three lateral, and three apical setae); setae I and II naked and inserted very close to each other dorsoventrally; seta I minute, hair-like, and about one-fourth of seta II in length; seta III about as long as seta VII and implanted in distal fifth; seta V longest and robust; setae IV and VI slender and almost equal in length; seta VII triarticulate and inserted in about distal third of inner margin.
Figure 33.
Scanning electron microscopy images of Phyllopodopsyllus similis sp. nov. male (paratype 24) in ventral view: (A) P5 and second to third urosomites; (B) fourth urosomite to anal somite; (C) caudal rami; (D) antennule and antenna. Scale bars: 30 µm.
Figure 33.
Scanning electron microscopy images of Phyllopodopsyllus similis sp. nov. male (paratype 24) in ventral view: (A) P5 and second to third urosomites; (B) fourth urosomite to anal somite; (C) caudal rami; (D) antennule and antenna. Scale bars: 30 µm.
Antennule (
Figure 31D,
Figure 32F,
Figure 33D,
Figure 36A,B and
Figure S12B) seven-segmented, subchirocer, covered with dimples on first three segments, and with geniculation between fourth and fifth segments; first segment longest, dented in inner margin proximally, beveled in outer margin, ornamented with row of spinules on proximal concave surface, and armed with seta implanted in inner distal corner; second segment without cuticular projection and with cuticular pore near boundary with following segment dorsally; third segment shortest; fourth segment enlarged, shorter than all following segments combined, and armed with aesthetasc fused basally to seta; fifth segment unarmed; sixth segment about as long as maximum width; fourth to sixth segments modified into pincer-like structure with several cuticular ridges; seventh segment with short aesthetasc fused with two setae at base; most of setae on seventh segment biarticulate. Armature formula as follow: 1(1), 2(11), 3(7), 4(13+(1+ae)), 5(1), 6(1), 7(9+(2+ae)).
It was difficult to determine whether: (a) the small lobe bearing two setae (marked by an empty arrow in
Figure 31D) was a part of the third or the fourth segment; (b) the coffee-bean-like projections on the fourth segment (marked by a hatched arrow in
Figure 31D) and on the fifth segment (marked by a checkered arrow in
Figure 31D) were the transformed armatures or the integumental ridges. The above descriptions and the formula were written assuming they were a part of the fourth segment and the transformed armatures, respectively.
P2 (
Figure 36C) as in female excluding following: Endopod almost reaching end of second exopodal segfment; first endopodal segment about 1.5 times as long as greatest width; second endopodal segment shorter (about 0.8 times) than preceding segment; middle armature narrowed sharply from middle and longest; outermost spine fused with endopodal segment at base; other two setae inserted very close to each other; innermost seta shortest and hairly.
Figure 34.
Line drawings of Phyllopodopsyllus similis sp. nov. male (paratype 25): (A) habitus, dorsal; (B) anal somite, dorsal; Scale bars: (A) 100 µm; (B) 50 µm.
Figure 34.
Line drawings of Phyllopodopsyllus similis sp. nov. male (paratype 25): (A) habitus, dorsal; (B) anal somite, dorsal; Scale bars: (A) 100 µm; (B) 50 µm.
Figure 35.
Line drawings of Phyllopodopsyllus similis sp. nov. male (paratype 25): urosomite, ventral. Scale bar: 50 µm.
Figure 35.
Line drawings of Phyllopodopsyllus similis sp. nov. male (paratype 25): urosomite, ventral. Scale bar: 50 µm.
Figure 36.
Line drawings of Phyllopodopsyllus similis sp. nov. male (paratype 25): (A) antennule, lateral; (B) antennule, dorsal; (C) P2. Scale bar: 50 µm.
Figure 36.
Line drawings of Phyllopodopsyllus similis sp. nov. male (paratype 25): (A) antennule, lateral; (B) antennule, dorsal; (C) P2. Scale bar: 50 µm.
Figure 37.
Line drawings of Phyllopodopsyllus similis sp. nov. male (paratype 25): (A) P3; (B) P4. Scale bar: 50 µm.
Figure 37.
Line drawings of Phyllopodopsyllus similis sp. nov. male (paratype 25): (A) P3; (B) P4. Scale bar: 50 µm.
P3 (
Figure 37A) as in female excluding following: Outer armature of second endopodal segment somewhat bent terminally, and much deeper embedded than other two setae; other two setae inserted very close to each other
P4 (
Figure 37B). as in female excluding following: Second endopodal segment about twice as long as wide and armed with two stout spines apically; inner spine longer than outer spine; third exopodal segment with two inner setae, two terminal setae, and two outer spines.
P5 (
Figure 33A,
Figure 35 and
Figure S12A) not foliaceous and composed of baseoendopod and exopod; both rami completely fused at central part. Baseoendopod large, with cuticular pore near base of basal seta, and armed with three pinnate setae on endopodal lobe; basal seta bare, slender, and much longer than P5; two apical setae equal in length; distal half of innermost seta bipinnate. Exopod about 1.6 times as long as greatest width, shorter than baseoendopod, and armed with five setae; two innermost setae robust and bipinnate, of which inner seta longest; other three setae slender.
P6 (
Figure 33A,
Figure 35 and
Figure S12A) reduced into single plate, completely fused at base with second urosomite, and armed with three armatures; innermost spine bipinnate and robust; middle seta longest, bare, and slender; outermost seta naked and about as long as innermost spine.
Variability
The body length ranged from about 561.5–685.9 µm (mean = 608.4 µm,
n = 8) in females (
Figure S3) and about 461.6–520.6 µm (mean = 485.7 µm,
n = 5) in males (
Figure S4). Another morphological variation appears in a single female individual (paratype 2). The left antennule of this individual shows the abnormalities, such as segmentation, chaetotaxy, and the shape of each segment (
Figure 21F).











































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}