
Genetic and Ecological Approaches to Introduced Populations of Pumpkinseed Sunfish (Lepomis gibbosus) in Southwestern Europe



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Sample Collection, DNA Extraction, Amplification, and Sequencing
2.2. Molecular Analysis
2.3. Phylogenetic Analysis
2.4. Analysis of Haplotypes, Genetic Distances, and Differentiation
2.5. Occurrence Data and Climatic Variables
2.6. Species Distribution Modeling (SDM)
2.7. Niche Overlap Analyses
3. Results
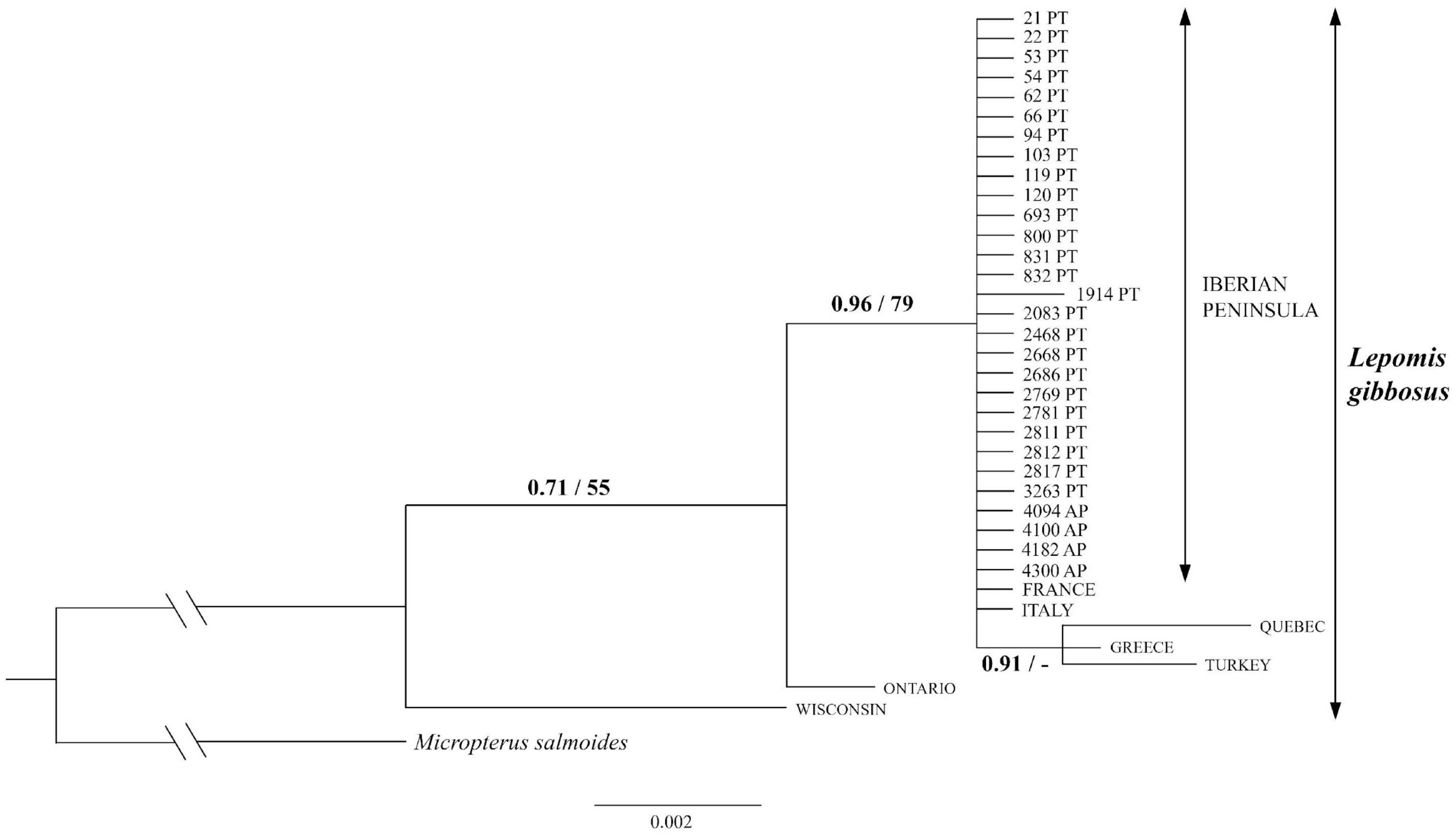
3.1. Molecular Characterization and Phylogenetic Analysis
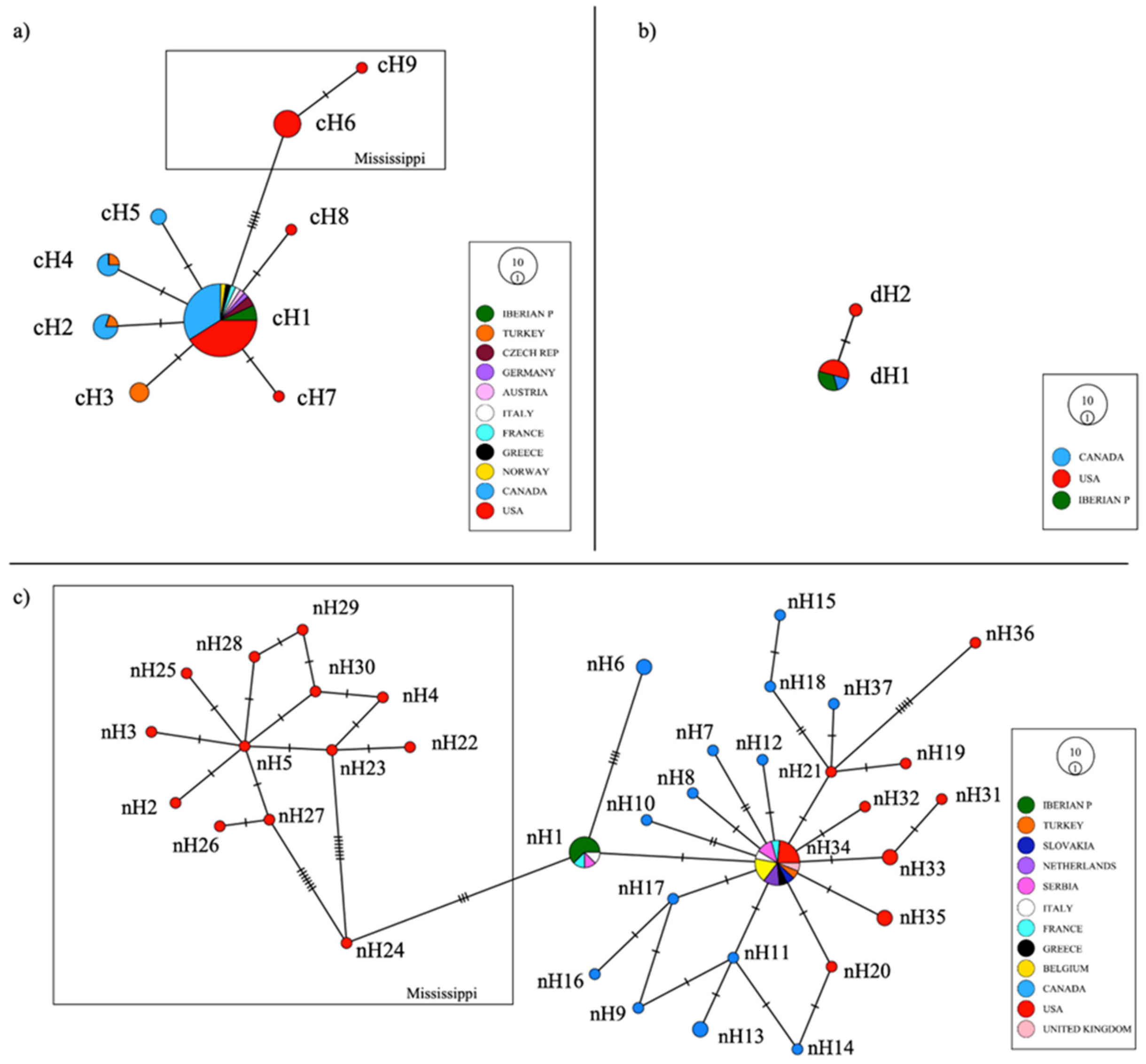
3.2. Population Analysis: Haplotype Network, Genetic Distances, and Differentiation
3.3. Species Distribution Modeling
3.4. Niche Identity and Overlap Analysis
4. Discussion
4.1. Origin and Dispersal of Lepomis gibbosus in Europe, Turkey, and the Iberian Peninsula
4.2. Niche Shift and Ecological Success
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bellard, C.; Jeschke, J.M.; Leroy, B.; Mace, G.M. Insights from modeling studies on how climate change affects invasive alien species geography. Ecol. Evol. 2018, 8, 5688–5700. [Google Scholar] [CrossRef] [PubMed]
- Drake, J.A. (Ed.) Handbook of Alien Species in Europe; Springer: Dordrecht, The Netherlands, 2009; pp. xxviii, 399. [Google Scholar]
- Broennimann, O.; Treier, U.A.; Müller-Schärer, H.; Thuiller, W.; Peterson, A.T.; Guisan, A. Evidence of climatic niche shift during biological invasion. Ecol. Lett. 2007, 10, 701–709. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.E. Evolutionary genetics of invasive species. Trends Ecol. Evol. 2002, 17, 386–391. [Google Scholar] [CrossRef]
- Rodríguez-Merino, A.; Fernández-Zamudio, R.; García-Murillo, P.; Muñoz, J. Climatic niche shift during Azolla filiculoides invasion and its potential distribution under future scenarios. Plants 2019, 8, 424. [Google Scholar] [CrossRef] [PubMed]
- Bernery, C.; Bellard, C.; Courchamp, F.; Brosse, S.; Gozlan, R.E.; Jarić, I.; Teletchea, F.; Leroy, B. Freshwater Fish Invasions: A Comprehensive Review, Annual Review of Ecology. Evol. Syst. 2022, 53, 427–456. [Google Scholar] [CrossRef]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W.; Zimmermann, N.E. Habitat Suitability and Distribution Models; R Cambridge University Press: Cambridge, UK, 2017; pp. xiii, 462. [Google Scholar]
- Gherardi, F. Biological invasions in inland waters: An overview. In Biological Invaders in Inland Waters: Profiles, Distribution, and Threats; Gherardi, F., Ed.; Invading Nature—Springer Series in Invasion Ecology; Springer: Dordrecht, The Netherlands, 2007; Volume 2. [Google Scholar]
- Elvira, B.; Almodóvar, A. Freshwater fish introductions in Spain: Facts and figures at the beginning of the 21st century. J. Fish Biol. 2001, 59, 323–331. [Google Scholar] [CrossRef]
- García-Berthou, E.; Alcaraz, C.; Pou-Rovira, Q.; Zamora, L.; Coenders, G.; Feo, C. Introduction pathways and establishment rates of invasive aquatic species in Europe. Can. J. Fish. Aquat. Sci. 2005, 2, 453–463. [Google Scholar] [CrossRef]
- Ribeiro, F.; Collares-Pereira, M.J. Life-history variability of non-native centrarchids in regulated river systems of the lower River Guadiana drainage (south-west Iberian Peninsula). J. Fish Biol. 2010, 76, 522–537. [Google Scholar] [CrossRef]
- Hermoso, V.; Clavero, M. Threatening processes and conservation management of endemic freshwater fish in the Mediterranean basin: A review. Mar. Freshw. Res. 2011, 62, 244–254. [Google Scholar] [CrossRef]
- Anastácio, P.M.; Ribeiro, F.; Capinha, C.; Banha, F.; Gama, M.; Filipe, A.F.; Rebelo, R.; Sousa, R. Non-native freshwater fauna in Portugal: A review. Sci. Total Environ. 2019, 650, 1923–1934. [Google Scholar] [CrossRef]
- Doadrio, I. (Ed.) Atlas y Libro Rojo de los Peces Continentales de España; MNCN-CSIC: Madrid, Spain, 2001; p. 364. [Google Scholar]
- IUCN—The World Conservation Union, Unión Internacional para la Conservación de la Naturaleza, de los Recursos Naturales, Comisión de Supervivencia de Especies de la UICN & IUCN Species Survival Commission. In Categorías Y Criterios De La Lista Roja De La UICN Versión 3.1 Segunda Edición; IUCN: Gland, Switzerland, 2001.
- Pyšek, P.; Hulme, P.E.; Simberloff, D.; Bacher, S.; Blackburn, T.M.; Carlton, J.T.; Dawson, W.; Essl, F.; Foxcroft, L.C.; Genovesi, P.; et al. Scientists’ warning on invasive alien species. Biol. Rev. 2020, 95, 1511–1534. [Google Scholar] [CrossRef] [PubMed]
- Almeida, D.; Merino-Aguirre, R.; Vilizzi, L.; Copp, G.H. Interspecific aggressive behaviour of invasive pumpkinseed Lepomis gibbosus in Iberian fresh waters. PLoS ONE 2014, 9, e88038. [Google Scholar] [CrossRef] [PubMed]
- Copp, G.H.; Fox, M.G.; Przybylski, M.; Godinho, F.N.; Vila-Gispert, A. Life-time growth patterns of Pumpkinseed Lepomis gibbosus introduced to Europe, relative to North American populations. Folia Zool. Int. J. Vertebr. Zool. 2004, 53, 237–254. [Google Scholar]
- Dill, W.A. FAO Report Inland Fisheries of Europe; Food and Agriculture Organization of the United Nations: Rome, Italy, 1993; p. 281. [Google Scholar]
- Van Kleef, H.H.; Van der Velde, G.; Leuven, R.S.E.W.; Esselink, H. Pumpkinseed sunfish (Lepomis gibbosus) invasions facilitated by introductions and nature management strongly reduce macroinvertebrate abundance in isolated water bodies. Biol. Invasions 2008, 10, 1481–1490. [Google Scholar] [CrossRef]
- De Groot, S.J. Introductions of non-indigenous fish species for release and culture in the Netherlands. Aquaculture 1985, 46, 237–257. [Google Scholar] [CrossRef]
- Yavno, S.; Gobin, J.; Wilson, C.C.; Vila-Gispert, A.; Copp, G.H.; Fox, M.G. New and Old World phylogeography of pumpkinseed Lepomis gibbosus (Linnaeus, 1758): The North American origin of introduced populations in Europe. Hydrobiologia 2020, 847, 345–364. [Google Scholar] [CrossRef]
- Zięba, G.; Vilizzi, L.; Copp, G.H. How likely is Lepomis gibbosus to become invasive in Poland under conditions of climate warming? Acta Ichthyol. Piscat. 2020, 50, 35–51. [Google Scholar] [CrossRef]
- Elvira, B. Native and exotic freshwater fishes in Spanish river basins. Freshw. Biol. 1995, 33, 103–108. [Google Scholar] [CrossRef]
- Arranz, I.; Fournier, B.; Lester, N.P.; Shuter, B.J.; Peres-Neto, P.R. Species compositions mediate biomass conservation: The case of lake fish communities. Ecology 2022, 103, e3608. [Google Scholar] [CrossRef]
- Almeida, D.; Almodóvar, A.; Nicola, G.G.; Elvira, B. Feeding tactics and body condition of two introduced populations of pumpkinseed Lepomis gibbosus: Taking advantages of human disturbances? Ecol. Freshw. Fish 2009, 18, 15–23. [Google Scholar] [CrossRef]
- Bhagat, Y.; Wilson, C.C.; Fox, M.G.; Ferreira, M.T. Genetic relationships among pumpkinseed (Lepomis gibbosus) ecomorphs in freshwater reservoirs of Portugal. Ecol. Freshw. Fish 2011, 20, 287–298. [Google Scholar] [CrossRef]
- Naspleda, J.; Zamora, L.; Vila-Gispert, A. Pez sol—Lepomis gibbosus. In Enciclopedia Virtual de los Vertebrados Españoles; Salvador, A., Elvira, B., Eds.; Museo Nacional de Ciencias Naturales: Madrid, Spain, 2012; Available online: http://www.vertebradosibericos.org/peces/lepgib.html (accessed on 3 June 2022).
- European Union. Regulation (EU) Nº 1143/2014 of the European Parliament and of the Council of 22 October 2014 on the Prevention and Management of the Introduction and Spread of Invasive Alien Species [Consolidated Version 14/Dec/2019]; Publications Office of the European Union: Luxembourg, 2014; p. 21. [Google Scholar]
- Genovesi, P.; Carboneras, C.; Vilà, M.; Walton, P. EU adopts innovative legislation on invasive species: A step towards a global response to biological invasions? Biol. Invasions 2015, 17, 1307–1311. [Google Scholar] [CrossRef]
- Copp, G.H.; Fox, M.G. Growth and life history traits of introduced pumpkinseed (Lepomis gibbosus) in Europe, and the relevance to its potential invasiveness. In Biological Invaders in Inland Waters: Profiles, Distribution, and Threats; Gherardi, F., Ed.; Invading Nature—Springer SERIES in Invasion Ecology; Springer: Dordrecht, The Netherlands, 2007; Volume 2. [Google Scholar]
- Ağdamar, S.; Tarkan, A.S.; Keskin, E.; Top, N.; Doğaç, E.; Baysal, Ö.; Emiroğlu, Ö. The role of environmental factors and genetic diversity on colonization success of a non-native fish, Lepomis gibbosus from western part of Turkey. Biochem. Syst. Ecol. 2015, 58, 195–203. [Google Scholar] [CrossRef]
- Tarkan, A.S.; Karakuş, U.; Top-Karakuş, N.; Keskin, E.; Ünal, E.M.; Britton, J.R. Invasion of pumpkinseed Lepomis gibbosus is facilitated by phenotypic plasticity across its invasion gradient. Biol. Invasions 2021, 23, 3201–3214. [Google Scholar] [CrossRef]
- Detta, A. The genetic origin and dispersal of introduced pumpkinseed (Lepomis gibbosus) in the Iberian Peninsula. Master’s Thesis, Trent University, Peterborough, ON, Canada, 2011; p. 86. [Google Scholar]
- Keller, R.P.; Frang, K.; Lodge, D.M. Preventing the spread of invasive species: Economic benefits of intervention guided by ecological predictions. Conserv. Biol. 2008, 22, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Keskin, E.; Ağdamar, S.; Tarkan, A.S. DNA barcoding common non-native freshwater fish species in Turkey: Low genetic diversity but high population structuring. Mitochondrial DNA 2013, 24, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, K.; Yonekura, R.; Katano, O.; Taniguchi, Y.; Saitoh, K. Origin and dispersal of bluegill sunfish, Lepomis macrochirus, in Japan and Korea. Mol. Ecol. 2006, 15, 613–621. [Google Scholar] [CrossRef]
- Palumbi, S.R.; Martin, A.; Romano, S.; McMillan, W.O.; Stice, L.; Grabowski, G. The Simple Fool’s Guide to PCR, v. 2.0.; University of Hawaii: Honolulu, HI, USA, 1991. [Google Scholar]
- Kawamura, K.; Nagata, Y.; Ohtaka, H.; Kanoh, Y.; Kitamura, J. Genetic diversity in the Japanese rosy bitterling, Rhodeus ocellatus kurumeus (Cyprinidae). Ichthyol. Res. 2001, 48, 369–378. [Google Scholar] [CrossRef]
- Hubert, N.; Hanner, R.; Holm, E.; Mandrak, N.E.; Taylor, E.; Burridge, M.; Watkinson, D.; Dumont, P.; Curry, A.; Bentzen, P.; et al. Identifying Canadian freshwater fishes through DNA barcodes. PLoS ONE 2008, 3, e2490. [Google Scholar] [CrossRef]
- April, J.; Mayden, R.L.; Hanner, R.H.; Bernatchez, L. Genetic calibration of species diversity among North America’s freshwater fishes. Proc. Natl. Acad. Sci. USA 2011, 108, 10602–10607. [Google Scholar] [CrossRef] [PubMed]
- Knebelsberger, T.; Dunz, A.R.; Neumann, D.; Geiger, M.F. Molecular diversity of Germany’s freshwater fishes and lampreys assessed by DNA barcoding. Mol. Ecol. Resour. 2015, 15, 562–572. [Google Scholar] [CrossRef] [PubMed]
- Schroeter, J.C.; Maloy, A.P.; Rees, C.B.; Bartron, M.L. Fish mitochondrial genome sequencing: Expanding genetic resources to support species detection and biodiversity monitoring using environmental DNA. Conserv. Genet. Resour. 2020, 12, 433–446. [Google Scholar] [CrossRef]
- Kawamura, K.; Yonekura, R.; Katano, O.; Taniguchi, Y.; Saitoh, K. Phylogeography of the bluegill sunfish, Lepomis macrochirus, in the Mississippi river basin. Zool. Sci. 2009, 26, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A. FigTree v1.4.2. A Graphical Viewer of Phylogenetic Trees. Institute of Evolutionary Biology University of Edinburgh. 2014. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 20 July 2022).
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. PopArt: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Swofford, D.L. Phylogenetic Analysis Using Parsimony (PAUP), Version 4.0 b10; Illinois Natural, History Survey: Champaign, IL, USA, 2002. [Google Scholar]
- Excoffier, L.; Lischer, H.E. Arlequin suite v3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Broennimann, O.; Di Cola, V.; Guisan, A. Ecospat: Spatial Ecology Miscellaneous Methods. R package Version 3.2. Available online: https://CRAN.R-project.org/package=ecospat (accessed on 3 June 2022).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 20 July 2022).
- De-Sostoa, A.; Lobón-Cerviá, J.; Fernández-Colomé, V.; De-Sostoa, F.J. La distribución del Pez-Sol (Lepomis gibbosus L.) en la Península Ibérica. Doñana Acta Vertebr. 1987, 14, 121–123. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1 km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Mamun, M.; Kim, S.; An, K.G. Distribution pattern prediction of an invasive alien species largemouth bass using a maximum entropy model (MaxEnt) in the Korean peninsula. J. Asia-Pac. Biodivers. 2018, 11, 516–524. [Google Scholar] [CrossRef]
- Manjarrés-Hernández, A.; Guisande, C.; García-Roselló, E.; Heine, J.; Pelayo-Villamil, P.; Pérez-Costas, E.; González-Vilas, L.; González-Dacosta, J.; Duque, S.R.; Granado-Lorencio, C.; et al. Predicting the effects of climate change on future freshwater fish diversity at global scale. Nat. Conserv. 2021, 43, 1–24. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.P. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Heibl, C.; Calenge, C. Phyloclim: Integrating Phylogenetics and Climatic Niche Modeling. R Package Version 0.9.5. 2018. Available online: https://CRAN.R-project.org/package=phyloclim (accessed on 20 July 2023).
- Warren, D.L.; Glor, R.E.; Turelli, M. Environmental niche equivalency versus conservatism: Quantitative approaches to niche evolution. Evolution 2008, 62, 2868–2883. [Google Scholar] [CrossRef] [PubMed]
- Warren, D.L.; Glor, R.E.; Turelli, M. ENMTools: A toolbox for comparative studies of environmental niche models. Ecography 2010, 33, 607–611. [Google Scholar] [CrossRef]
- Mandrak, N.E.; Crossman, E.J. Postglacial dispersal of freshwater fishes into Ontario. Can. J. Zool. 2002, 70, 2247–2259. [Google Scholar] [CrossRef]
- Yonekura, R.; Kawamura, K.; Uchii, K. A peculiar relationship between genetic diversity and adaptability in invasive exotic species: Bluegill sunfish as a model species. Ecol. Res. 2007, 22, 911–919. [Google Scholar] [CrossRef]
- Clavero, M.; García-Berthou, E. Homogenization dynamics and introduction routes of invasive freshwater fish in the Iberian Peninsula. Ecol. Appl. 2006, 16, 2313–2324. [Google Scholar] [CrossRef]
- Ribeiro, F.; Veríssimo, A. Full westward expansion of Rutilus rutilus (Linnaeus, 1758) in the Iberian Peninsula. J. Appl. Ichthyol. 2014, 30, 540–542. [Google Scholar] [CrossRef]
- Ashenden, T.; Rooke, A.C.; Fox, M.G. Boldness and dispersal tendency of native and invasive pumpkinseed (Lepomis gibbosus): Is spatial sorting creating superior invaders? Aquat. Invasions 2017, 12, 311–320. [Google Scholar] [CrossRef]
- Villeneuve, F.; Copp, G.H.; Fox, M.G.; Stakėnas, S. Interpopulation variation in growth and life-history traits of the introduced sunfish, pumpkinseed Lepomis gibbosus, in southern England. J. Appl. Ichthyol. 2005, 21, 275–281. [Google Scholar] [CrossRef]
- Rooke, A.C.; Fox, M.G. Living a century in warm thermal conditions: Introduced populations of Pumpkinseed (Lepomis gibbosus) inhabiting the Iberian Peninsula (Spain) show reduced adaptation to North American winter conditions. Aquat. Sci. 2014, 76, 497–509. [Google Scholar] [CrossRef]
- Artaev, O. Prediction of Current and Future Suitable Habitats for Three Invasive Freshwater Fish Species in Europe. Water 2023, 15, 2091. [Google Scholar] [CrossRef]
- Osenberg, C.W.; Werner, E.E.; Mittelbach, G.G.; Hall, D.J. Growth patterns in bluegill (Lepomis macrochirus) and pumpkinseed (L. gibbosus) sunfish: Environmental variation and the importance of ontogenetic niche shifts. Can. J. Fish. Aquat. Sci. 1988, 45, 17–26. [Google Scholar] [CrossRef]
- Fobert, E.; Fox, M.G.; Ridgway, M.; Copp, G.H. Heated competition: How climate change will affect non-native pumpkinseed Lepomis gibbosus and native perch Perca fluviatilis interactions in the UK. J. Fish Biol. 2011, 79, 1592–1607. [Google Scholar] [CrossRef] [PubMed]
- Yavno, S.; Fox, M.G. Morphological change and phenotypic plasticity in native and non-native pumpkinseed sunfish in response to sustained water velocities. J. Evol. Biol. 2013, 26, 2383–2395. [Google Scholar] [CrossRef] [PubMed]
- Almeida, D.; Ribeiro, F.; Leunda, P.M.; Vilizzi, L.; Copp, G.H. Effectiveness of FISK, an invasiveness screening tool for non-native freshwater fishes, to perform risk identification assessments in the Iberian Peninsula. Risk Anal. 2013, 33, 1404–1413. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence | PCR Conditions | Reference | |||
|---|---|---|---|---|---|---|
| Cytochrome c oxidase subunit I (COI) | Fish-1F | 5′-TCAACCAACCACAAAGACATTGGCAC-3′ | 95 °C | 2′ | [37] | |
| 35× | 95 °C | 30″ | ||||
| 54 °C | 30″ | |||||
| Fish-1R | 5′-TAGACTTCTGGGTGGCCAAAGAATCA-3′ | 72 °C | 1′ | [37] | ||
| 72 °C | 10′ | |||||
| 10 °C | ∞ | |||||
| Control region or D loop | CB3R-L | 5′-CAYATYMARCCMGAATGRTATTT-3′ | 94 °C | 2′ | [38] | |
| 40× | 94 °C | 30″ | ||||
| 55 °C | 30″ | |||||
| 12SAR-H | 5′-ATARTRGGGTATCTAATCCYAGTT-3′ | 72 °C | 2′ | [39] | ||
| 72 °C | 5′ | |||||
| 10 °C | ∞ | |||||
| NADH dehydrogenase 1 (ND1) | L2949-LMA | 5′-AGTTACCCTAGGGATAACAGCGCAATC-3′ | 95 °C | 5′ | [23] | |
| 10× (−0.5 °C) | 95 °C | 45″ | ||||
| 55 °C | 1′ | |||||
| 72 °C | 1′30″ | |||||
| 30× | 95 °C | 45″ | ||||
| S2-LMA | 5′-GGTATGGGCCCAAAAGCTTA-3′ | 52 °C | 1′30″ | [40] | ||
| 72 °C | 1′30″ | |||||
| 72 °C | 6′ | |||||
| 10 °C | ∞ | |||||
| GenBank ID. | Locality | Country | COI | Reference | GenBank | Locality | Country | D-loop | Reference | GenBank | Locality | Country | ND1 | Reference |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EU524723 | Quebec | CA | cH1 | [41] | MF621725 | Lake Erie | CA | dH1 | [42] | MN516524 | Quebec | CA | nH13 | [23] |
| EU524724 | Quebec | CA | cH1 | [41] | MF621724 | Pennsylvania | US | dH2 | [42] | MN516520 | Quebec | CA | nH14 | [23] |
| BCFB584-06 * | Quebec | CA | cH1 | [41] | MT667250 | Pennsylvania | US | dH1 | Unpublished | MN516505 | Quebec | CA | nH15 | [23] |
| BCFB585-06 * | Quebec | CA | cH1 | [41] | MF621726 | New York | US | dH1 | [42] | MN516501 | Quebec | CA | nH16 | [23] |
| BCFB586-06 * | Quebec | CA | cH1 | [41] | Iberian Peninsula | ES/PT | dH1 | MN516500 | Quebec | CA | nH17 | [23] | ||
| BCFB587-06 * | Quebec | CA | cH1 | [41] | MN516496 | Quebec | CA | nH18 | [23] | |||||
| BCFB588-06 * | Quebec | CA | cH1 | [41] | MN516527 | Ontario | CA | nH6 | [23] | |||||
| NHFEC079 * | Quebec | CA | cH1 | [41] | MN516526 | Ontario | CA | nH7 | [23] | |||||
| BCFB583-06 * | Quebec | CA | cH4 | [41] | MN516525 | Ontario | CA | nH8 | [23] | |||||
| EU524725 | Quebec | CA | cH4 | [41] | MN516521 | Ontario | CA | nH9 | [23] | |||||
| BCFB589-06 * | Ontario | CA | cH1 | [41] | MN516523 | Ontario | CA | nH10 | [23] | |||||
| BCFB590-06 * | Ontario | CA | cH1 | [41] | MN516519 | Ontario | CA | nH11 | [23] | |||||
| BCFB606-06 * | Ontario | CA | cH1 | [41] | MN516499 | Ontario | CA | nH12 | [23] | |||||
| BCFB592-06 * | Ontario | CA | cH2 | [41] | MF621725 | Lake Erie | CA | nH37 | Unpublished | |||||
| BCFB593-06 * | Ontario | CA | cH2 | [41] | MN516514 | Wisconsin | US | nH28 | [23] | |||||
| BCFB594-06 * | Ontario | CA | cH2 | [41] | MN516509 | Wisconsin | US | nH29 | [23] | |||||
| EU524717 | Ontario | CA | cH2 | [41] | MN516508 | Wisconsin | US | nH30 | [23] | |||||
| CFF194-16 * | Lake Erie | CA | cH1 | Unpublished | MN516490 | North Carolina | US | nH34 | [23] | |||||
| CFF048-16 * | Lake Erie | CA | cH5 | Unpublished | MN516528 | Minnesota | US | nH2 | [23] | |||||
| CFF104-16 * | Durand Lake | CA | cH1 | Unpublished | MN516515 | Minnesota | US | nH3 | [23] | |||||
| CFF107-16 * | Riviere Saint-Jean | CA | cH1 | Unpublished | MN516507 | Minnesota | US | nH4 | [23] | |||||
| CFF117-16 * | Opinicon Lake | CA | cH1 | Unpublished | MN516513 | Minnesota | US | nH5 | [23] | |||||
| CFF171-16 | Saint Louis Lake | CA | cH4 | Unpublished | MN516502 | Pennsylvania | US | nH19 | [23] | |||||
| RMAYB187-07 * | Ottawa | US | cH7 | [43] | MN516498 | Pennsylvania | US | nH20 | [23] | |||||
| SERCA041-12 * | Maryland | US | cH1 | [44] | MN516493 | Pennsylvania | US | nH21 | [23] | |||||
| UKFBJ845-08 * | New Hampshire | US | cH1 | Unpublished | MF621726 | New York | US | nH34 | Unpublished | |||||
| BNAFB548 * | Wisconsin | US | cH6 | Unpublished | MN516497 | New Jersey | US | nH35 | [23] | |||||
| HQ557271 | Wisconsin | US | cH6 | Unpublished | MN516518 | Iowa | US | nH22 | [23] | |||||
| RMAYB188-07 * | Wisconsin | US | cH8 | [43] | MN516511 | Iowa | US | nH23 | [23] | |||||
| BNAFB547-09 * | Wisconsin | US | cH6 | [43] | MN516510 | Indiana | US | nH24 | [23] | |||||
| JN026988 | South Carolina | US | cH1 | [43] | MN516517 | Indiana | US | nH25 | [23] | |||||
| RMAYB189-07 * | South Carolina | US | cH1 | [43] | MN516516 | Michigan | US | nH26 | [23] | |||||
| EPAMC397-20 * | Cincinnati | US | cH1 | Unpublished | MN516512 | Michigan | US | nH27 | [23] | |||||
| EPAMC398-20 * | Cincinnati | US | cH1 | Unpublished | MN516522 | Virginia | US | nH31 | [23] | |||||
| EPAMC399-20 * | Cincinnati | US | cH1 | Unpublished | MN516503 | Virginia | US | nH32 | [23] | |||||
| EPAMC400-20 * | Cincinnati | US | cH1 | Unpublished | MN516504 | Virginia | US | nH32 | [23] | |||||
| EPAMC401-20 * | Cincinnati | US | cH1 | Unpublished | MN516494 | Virginia | US | nH33 | [23] | |||||
| EPAMC402-20 * | Cincinnati | US | cH1 | Unpublished | MN516495 | Virginia | US | nH33 | [23] | |||||
| EPAMC403-20 * | Cincinnati | US | cH1 | Unpublished | MN516492 | Virginia | US | nH34 | [23] | |||||
| EPAMC404-20 * | Cincinnati | US | cH1 | Unpublished | MN516489 | Virginia | US | nH34 | [23] | |||||
| EPAMC405-20 * | Cincinnati | US | cH1 | Unpublished | AB271766 | Hamilton | US | nH36 | [45] | |||||
| EPAMC406-20 * | Cincinnati | US | cH1 | Unpublished | Iberian Peninsula | ES/PT | nH1 | |||||||
| EPAMC407-20 * | Cincinnati | US | cH1 | Unpublished | MN516506 | Monte Novo | PT | nH1 | [23] | |||||
| EPAMC408-20 * | Cincinnati | US | cH1 | Unpublished | MN516506 | River Ebro and | ES/PT | nH1 | [23] | |||||
| EPAMC409-20 * | Cincinnati | US | cH1 | Unpublished | MN516506 | Lake Bolsena | IT | nH1 | [23] | |||||
| EPAMC410-20 * | Cincinnati | US | cH1 | Unpublished | MN516489 | River Picocca | IT | nH34 | [23] | |||||
| EFA105-16 * | Lake Superior | US | cH6 | Unpublished | MN516506 | Argancy Pond | FR | nH1 | [23] | |||||
| EFA207-17 * | Lake Superior | US | cH9 | Unpublished | MN516489 | Birazel | FR | nH34 | [23] | |||||
| Iberian Peninsula | ES/PT | cH1 | MN516489 | Lake Prespa | GR | nH34 | [23] | |||||||
| HQ960772 | Hradec Králové | CZ | cH1 | Unpublished | MN516506 | Mrzenica | RS | nH1 | [23] | |||||
| ANGBF56447-19 * | Czech Republic | CZ | cH1 | MN516489 | Mrzenica | RS | nH34 | [23] | ||||||
| FBPIS137-10 * | Baviera | DE | cH1 | [44] | MN516491 | Mrzenica | RS | nH34 | [23] | |||||
| FBPIS140-10 * | Styria | AT | cH1 | [44] | MN516489 | De Maten Ponds | BE | nH34 | [23] | |||||
| FFMBH104-14 * | Turin | IT | cH1 | Unpublished | MN516491 | Mastbos Pond | NL | nH34 | [23] | |||||
| MN516489 | Mastbos Pond | NL | nH34 | [23] | ||||||||||
| FFMBH1592-14 * | Canton du Pilat | FR | cH1 | Unpublished | MN516489 | Ogosta Reservoir | BG | nH34 | [23] | |||||
| MN516489 | Slovakia | SK | nH34 | [23] | ||||||||||
| FFMBH2708-14 * | Sitagri | GR | cH1 | Unpublished | MN516489 | Tanyard Fisheries | GB | nH34 | [23] | |||||
| NOFIS136-18 * | Asker | NO | cH1 | Unpublished | MN516489 | Sariçay Stream | TR | nH34 | [23] | |||||
| JQ979159 | Ipsala | TR | cH2 | [37] | ||||||||||
| JQ979160 | Cayirkoy | TR | cH3 | [37] | ||||||||||
| JQ979161 | Bayraktar | TR | cH3 | [37] | ||||||||||
| JQ979162 | Davuldere | TR | cH3 | [37] | ||||||||||
| JQ979163 | Mugla | TR | cH4 | [37] |
| Gene | Group | America | Europe | Iberian Peninsula | Gene Mean | ||
|---|---|---|---|---|---|---|---|
| COI | America | 0.32 ± 0.42 {0.0–1.36} | COI | 0.16 ± 0.33 {0.00–1.36} | Within | ||
| Europe | 0.25 ± 000.35 {0.00–1.35} | 0.13 ± 0.12 {0.00–0.39} | 0.17 ± 0.32 {0.00–1.35} | Between | |||
| Iberian P | 0.18 ± 0.34 {0.00–1.16} | 0.07 ± 0.09 {0.00–0.19} | 0.00 ± 0.00 {0.00–0.00} | ||||
| ND-1 | America | 0.76 ± 0.53 {0.00–1.95} | ND-1 | 0.36 ± 0.53 {0.00–1.95} | Within | ||
| Europe | 0.49 ± 0.47 {0.00–1.33} | 0.03 ± 0.05 {0.00–0.10} | 0.42 ± 0.42 {0.00–1.33} | Between | |||
| Iberian P | 0.52 ± 0.42 {0.10–1.33} | 0.08 ± 0.04 {0.00–0.10} | 0.00 ± 0.00 {0.00–0.00} | ||||
| D-loop | America | 0.05 ± 0.06 {0.00–0.12} | D-loop | 0.00 ± 0.01 {0.00–0.12} | Within | ||
| Iberian P | 0.02 ± 0.05 {0.00–0.12} | 0.00 ± 0.00 {0.00–0.00} | 0.02 ± 0.05 {0.00–0.12} | Between |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lambea-Camblor, A.; Morcillo, F.; Muñoz, J.; Perdices, A. Genetic and Ecological Approaches to Introduced Populations of Pumpkinseed Sunfish (Lepomis gibbosus) in Southwestern Europe. Diversity 2023, 15, 1059. https://doi.org/10.3390/d15101059
Lambea-Camblor A, Morcillo F, Muñoz J, Perdices A. Genetic and Ecological Approaches to Introduced Populations of Pumpkinseed Sunfish (Lepomis gibbosus) in Southwestern Europe. Diversity. 2023; 15(10):1059. https://doi.org/10.3390/d15101059
Chicago/Turabian StyleLambea-Camblor, Angela, Felipe Morcillo, Jesús Muñoz, and Anabel Perdices. 2023. "Genetic and Ecological Approaches to Introduced Populations of Pumpkinseed Sunfish (Lepomis gibbosus) in Southwestern Europe" Diversity 15, no. 10: 1059. https://doi.org/10.3390/d15101059
APA StyleLambea-Camblor, A., Morcillo, F., Muñoz, J., & Perdices, A. (2023). Genetic and Ecological Approaches to Introduced Populations of Pumpkinseed Sunfish (Lepomis gibbosus) in Southwestern Europe. Diversity, 15(10), 1059. https://doi.org/10.3390/d15101059