

Bioenergetic Model of the Highly Exploited Shark Mustelus schmitti under a Global Warming Context

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bioenergetics Model
2.1.1. Growth
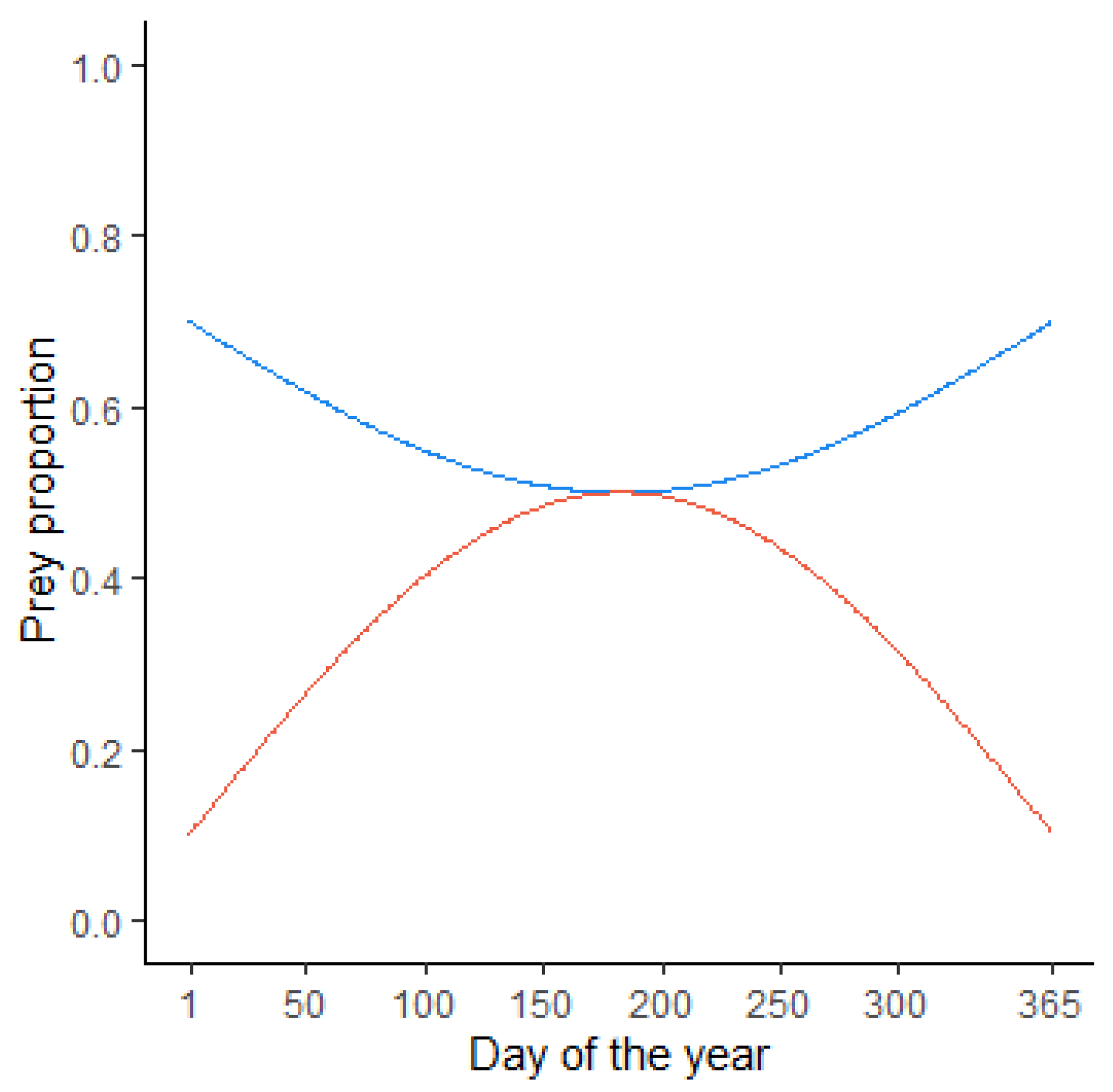
2.1.2. Consumption
2.1.3. Respiration
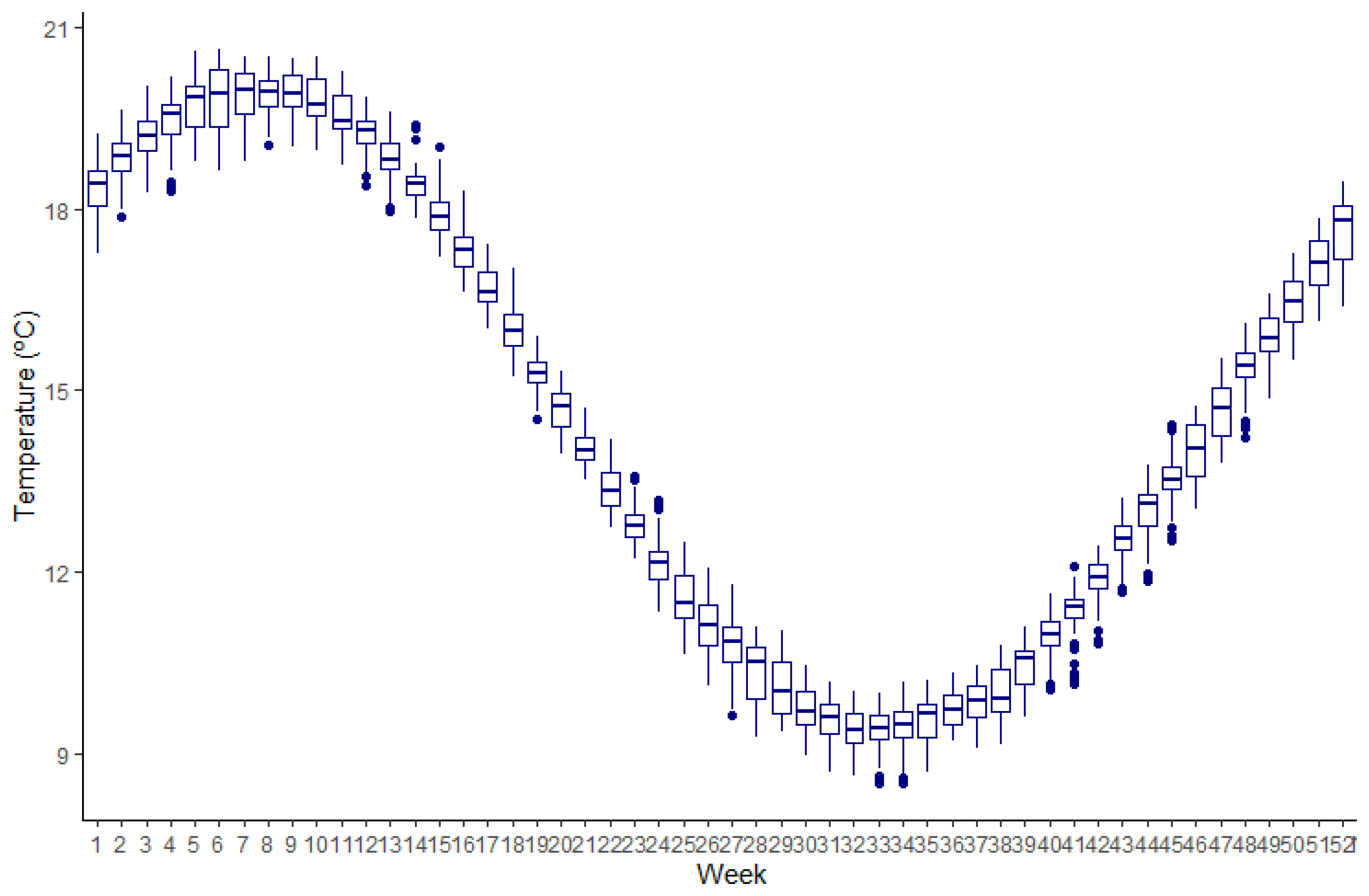
2.2. Temperature
2.3. Model Uncertainties, Simulated Variability, and Monte Carlo Methods
2.3.1. Individual-Level Variability
2.3.2. Environmental-Level Variability
2.3.3. Monte Carlo Simulations
2.4. High Temperature Scenarios
2.5. Practical Application
2.6. Statistical Analysis
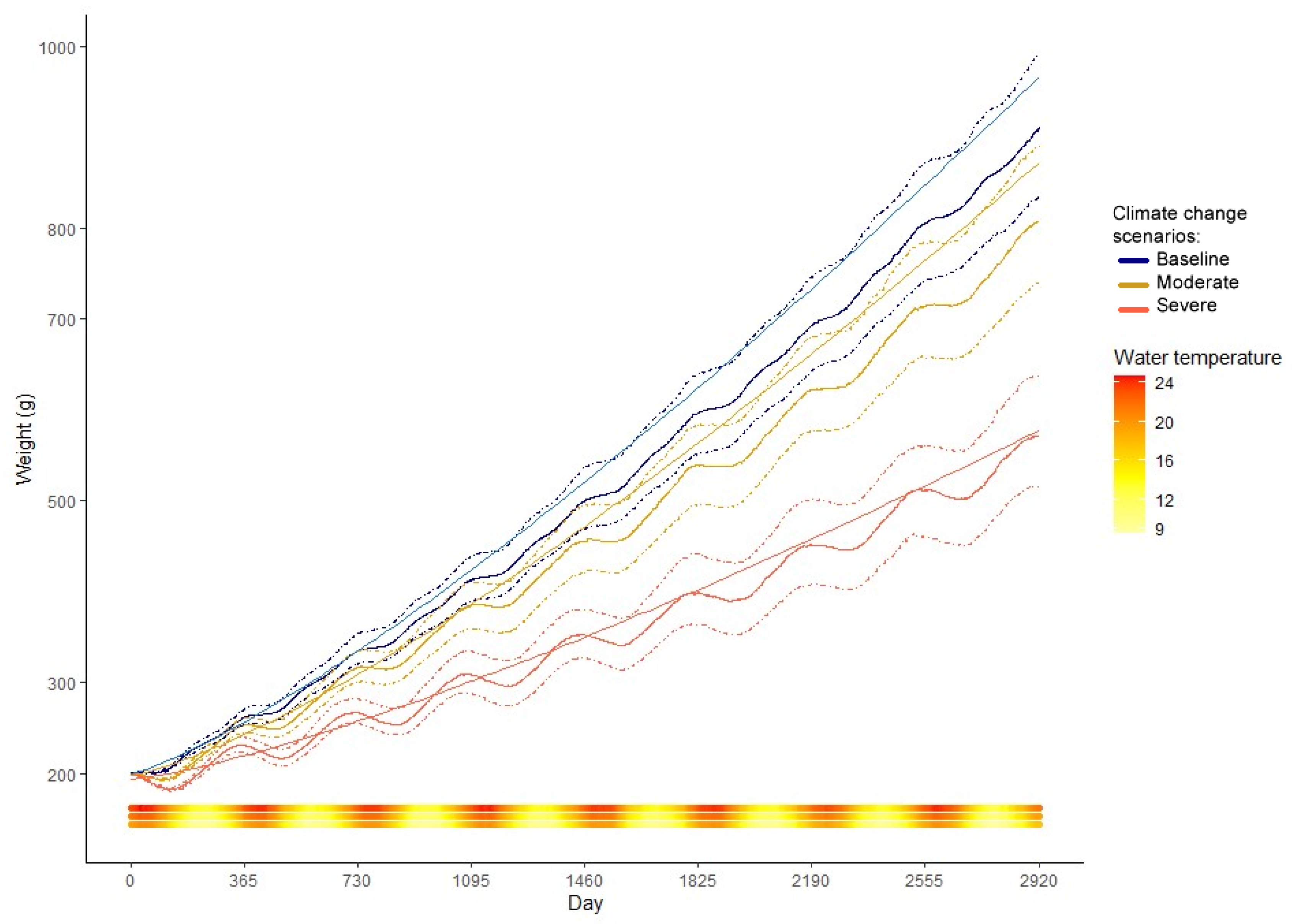
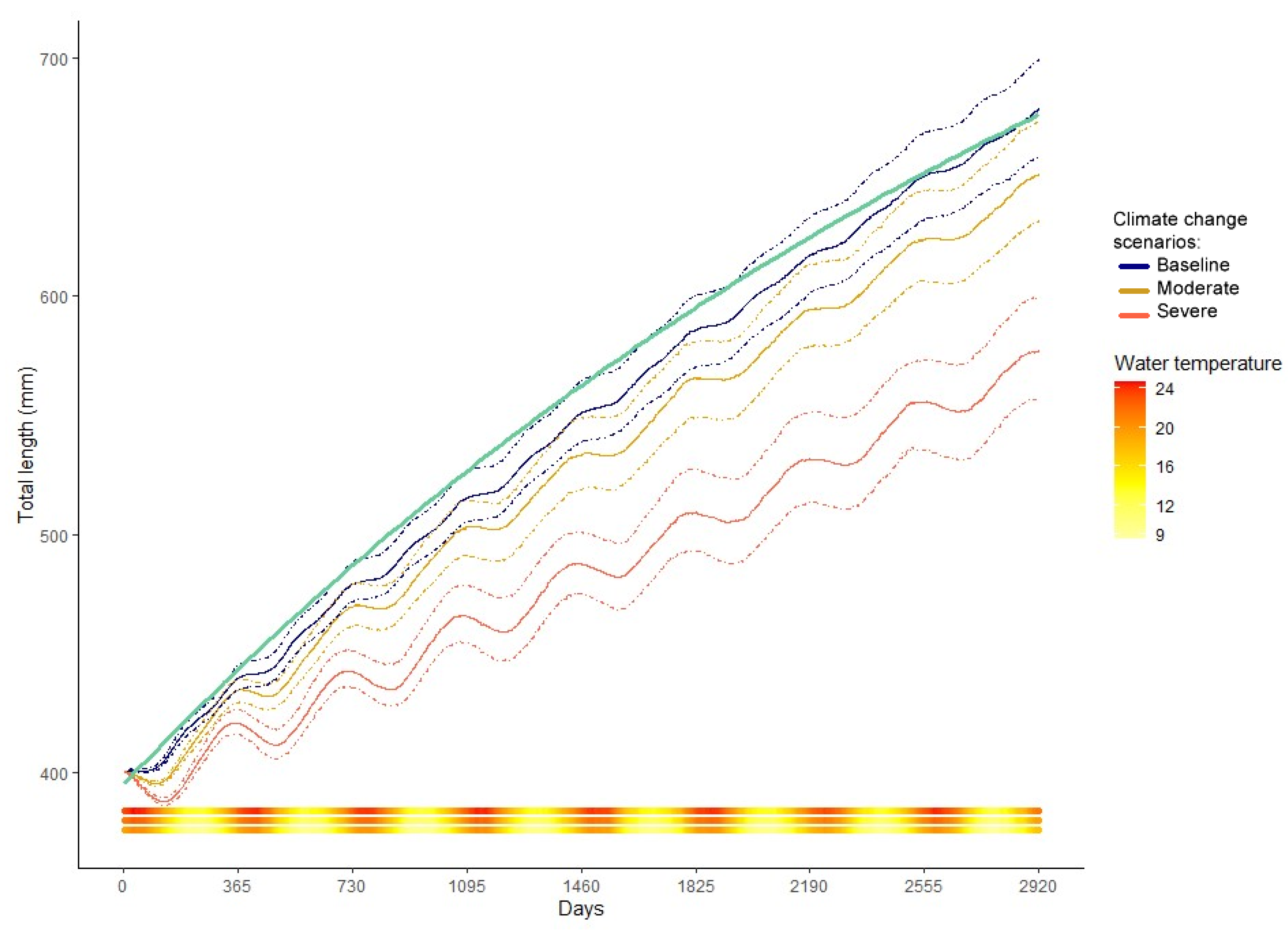
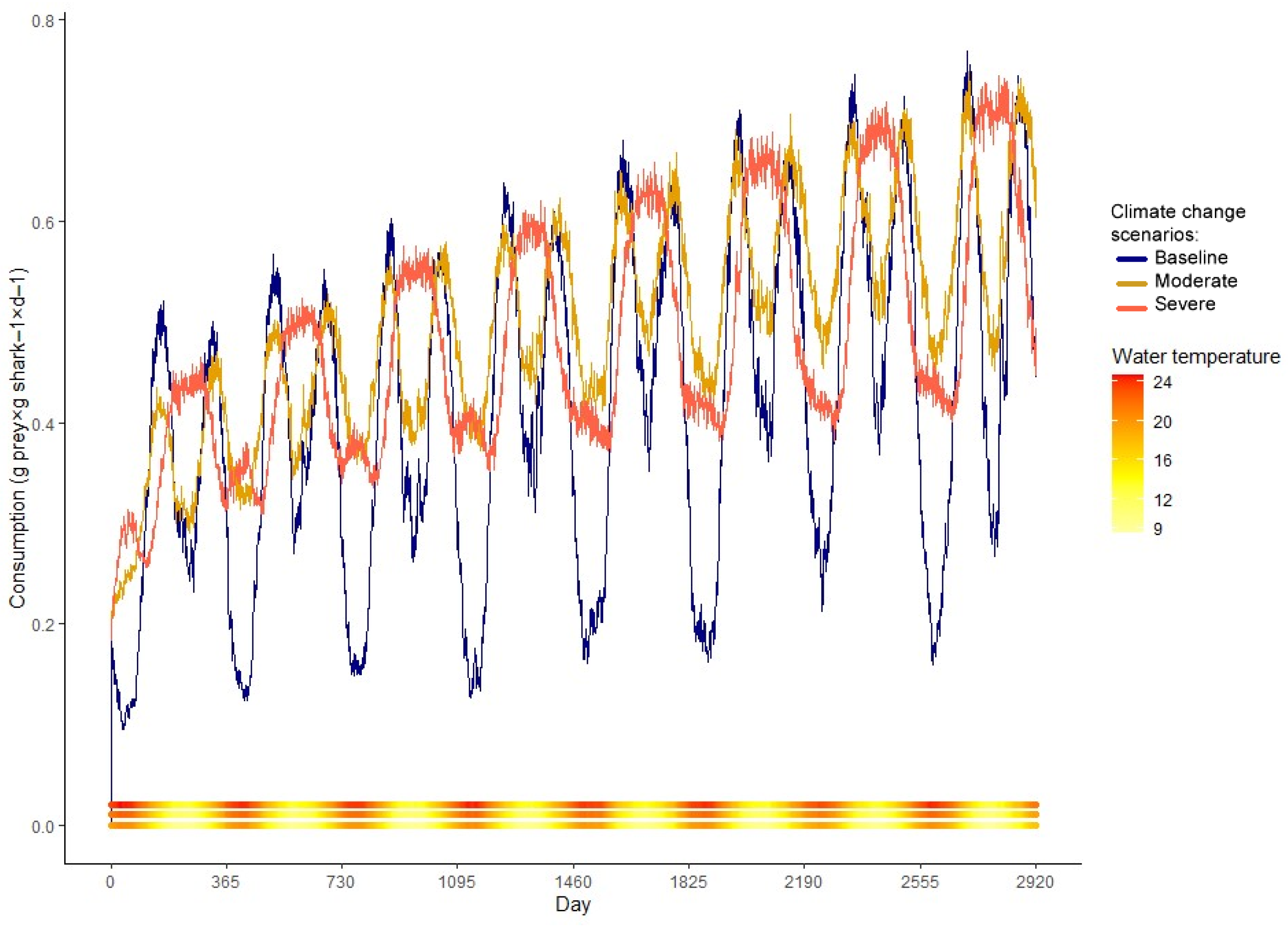
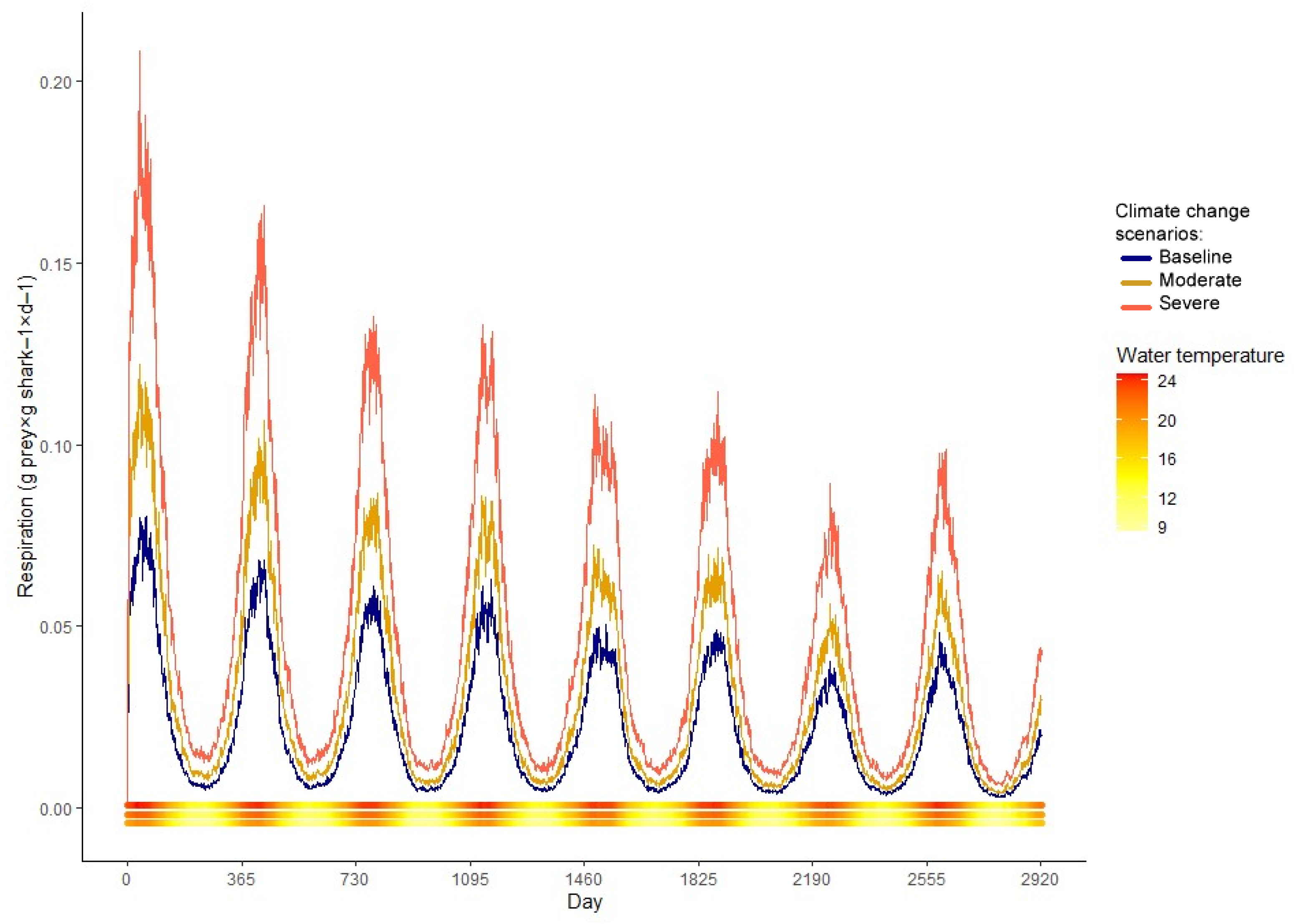
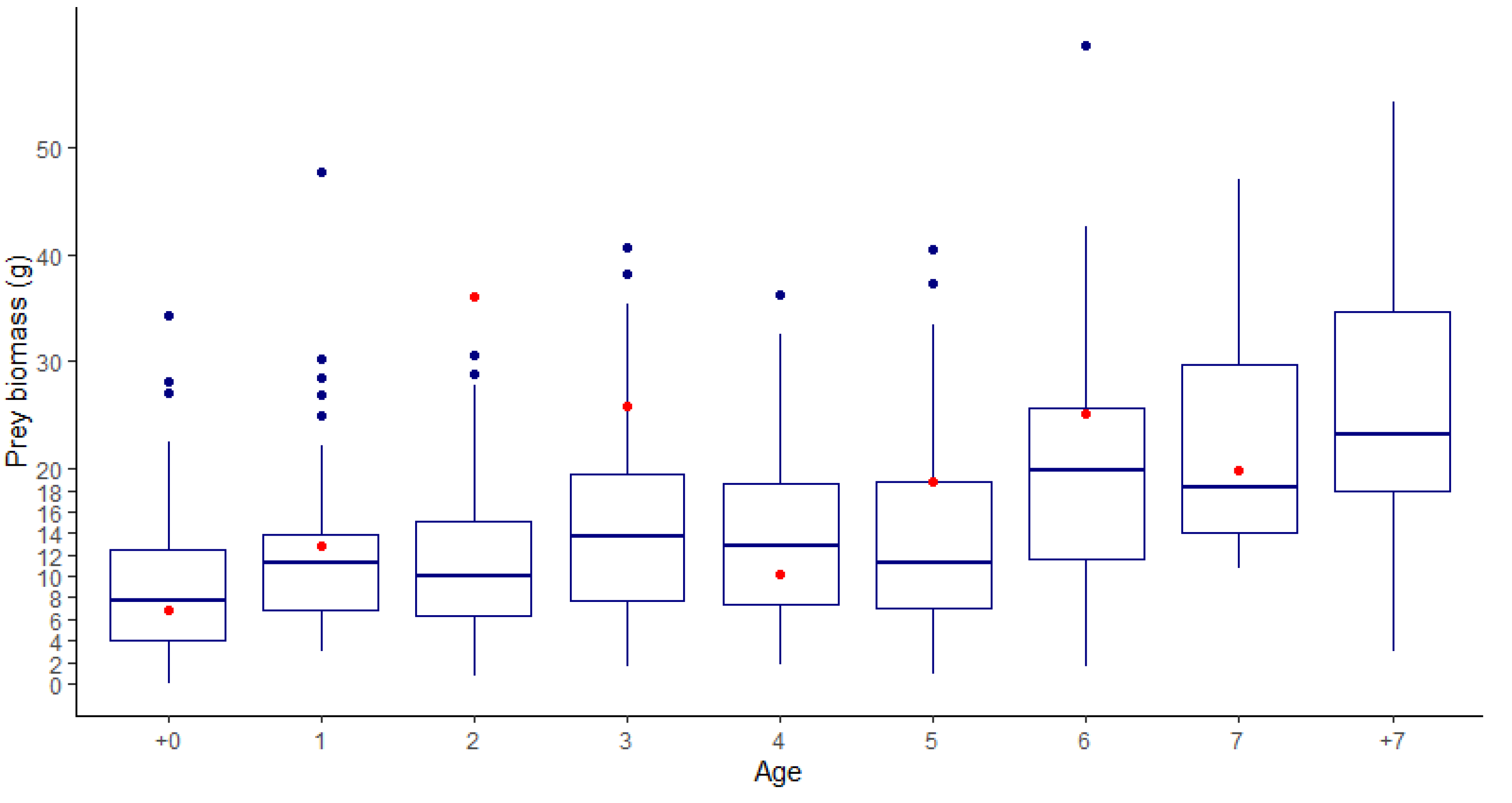
3. Results
4. Discussion
4.1. The Bioenergetic Model
4.2. Climate Change Scenarios
4.3. Model Limitations
- (1)
- Refinement and calibration: Further refinement of the bioenergetic model can be achieved by incorporating more precise and species-specific parameters [8,118]. Conducting controlled experimental studies to measure the energy consumption of M. schmitti, and determining the metabolizable energy of prey species would contribute to reducing uncertainties and improving model predictions [66,74,77]. Calibration of the model using independent field sampling data would also enhance its accuracy [115].
- (2)
- Incorporation of ecological factors: The model can be expanded to consider additional ecological factors that influence the energy requirements and behavior of M. schmitti [74,114]. Factors such as predation risk, competition, and habitat preferences can significantly impact the energy dynamics of the species [116,117]. Incorporating these factors, when sufficient data become available, would improve the model’s ecological realism and predictive power.
- (3)
- Long-term monitoring and data collection: Long-term monitoring of M. schmitti populations, including growth rates, prey availability, and environmental conditions, would provide valuable data for validating and updating the model [115,119]. Continued data collection efforts, such as tagging studies and dietary analyses, can contribute to a better understanding of the species’ energy requirements and feeding ecology [120].
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Giovos, I.; Aga Spyridopoulou, R.N.; Doumpas, N.; Glaus, K.; Kleitou, P.; Kazlari, Z.; Katsada, D.; Loukovitis, D.; Mantzouni, I.; Papapetrou, M.; et al. Approaching the “real” state of elasmobranch fisheries and trade: A case study from the Mediterranean. Ocean. Coast. Manag. 2021, 211, 105743. [Google Scholar] [CrossRef]
- Bonfil, R.; Food and Agriculture Organization of the United Nations. Overview of World Elasmobranch Fisheries; Food & Agriculture Organization: Rome, Italy, 1994. [Google Scholar]
- Molina, J.M.; Cooke, S.J. Trends in shark bycatch research: Current status and research needs. Rev. Fish Biol. Fish. 2012, 22, 719–737. [Google Scholar] [CrossRef]
- Gallagher, A.J.; Kyne, P.M.; Hammerschlag, N. Ecological risk assessment and its application to elasmobranch conservation and management. J. Fish Biol. 2012, 80, 1727–1748. [Google Scholar] [CrossRef]
- Caswell, B.A.; Klein, E.S.; Alleway, H.K.; Ball, J.E.; Botero, J.; Cardinale, M.; Eero, M.; Engelhard, G.H.; Fortibuoni, T.; Giraldo, A.J.; et al. Something old, something new: Historical perspectives provide lessons for blue growth agendas. Fish Fish. 2020, 21, 774–796. [Google Scholar] [CrossRef]
- Parmesan, C.; Pörtner, H.-O.; Roberts, D.C. IPCC 2022: Summary for Policymakers. In Climate Change 2022: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2022. [Google Scholar]
- Koenigstein, S.; Mark, F.C.; Gößling-Reisemann, S.; Reuter, H.; Poertner, H.O. Modelling climate change impacts on marine fish populations: Process-based integration of ocean warming, acidification and other environmental drivers. Fish Fish. 2016, 17, 972–1004. [Google Scholar] [CrossRef]
- Neer, J.A.; Rose, K.A.; Cortés, E. Simulating the effects of temperature on individual and population growth of Rhinoptera bonasus: A coupled bioenergetics and matrix modeling approach. Mar. Ecol. Prog. Ser. 2007, 329, 211–223. [Google Scholar] [CrossRef]
- Bejarano, A.C.; Wells, R.S.; Costa, D.P. Development of a bioenergetic model for estimating energy requirements and prey biomass consumption of the bottlenose dolphin Tursiops truncatus. Ecol. Model. 2017, 356, 162–172. [Google Scholar] [CrossRef]
- Bouyoucos, I.A.; Simpfendorfer, C.A.; Rummer, J.L. Estimating oxygen uptake rates to understand stress in sharks and rays. Rev. Fish Biol. Fish. 2019, 29, 297–311. [Google Scholar] [CrossRef]
- Yoon, S.; Watanabe, E.; Ueno, H.; Kishi, M.J. Potential habitat for chum salmon (Oncorhynchus keta) in the Western Arctic based on a bioenergetics model coupled with a three-dimensional lower trophic ecosystem model. Prog. Oceanogr. 2015, 131, 146–158. [Google Scholar] [CrossRef]
- Kishi, M.J.; Kaeriyama, M.; Ueno, H.; Kamezawa, Y. The effect of climate change on the growth of Japanese chum salmon (Oncorhynchus keta) using a bioenergetics model coupled with a three-dimensional lower trophic ecosystem model (NEMURO). Deep. Sea Res. Part II Top. Stud. Oceanogr. 2010, 57, 1257–1265. [Google Scholar] [CrossRef]
- Megrey, B.A.; Rose, K.A.; Werner, F.E.; Klumb, R.A.; Hay, D. A generalized fish bioenergetics/biomass model with an application to Pacific herring. PICES Sci. Rep. 2002, 20, 4–12. [Google Scholar]
- Clark, T.D.; Sandblom, E.; Jutfelt, F. Aerobic scope measurements of fishes in an era of climate change: Respirometry, relevance and recommendations. J. Exp. Biol. 2013, 216, 2771–2782. [Google Scholar] [CrossRef]
- Molina, J.M.; Finotto, L.; Walker, T.I.; Reina, R.D. The effect of gillnet capture on the metabolic rate of two shark species with contrasting lifestyles. J. Exp. Mar. Biol. Ecol. 2020, 526, 151354. [Google Scholar] [CrossRef]
- Chabot, D.; Koenker, R.; Farrell, A.P. The measurement of specific dynamic action in fishes. J. Fish Biol. 2016, 88, 152–172. [Google Scholar] [CrossRef]
- Skomal, G.B.; Mandelman, J.W. The physiological response to anthropogenic stressors in marine elasmobranch fishes: A review with a focus on the secondary response. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2012, 162, 146–155. [Google Scholar] [CrossRef]
- Madliger, C.L.; Cooke, S.J.; Crespi, E.J.; Funk, J.L.; Hultine, K.R.; Hunt, K.E.; Rohr, J.R.; Sinclair, B.J.; Suski, C.D.; Willis, C.K.; et al. Success stories and emerging themes in conservation physiology. Conserv. Physiol. 2016, 4, cov057. [Google Scholar] [CrossRef]
- Chipps, S.R.; Wahl, D.H. Bioenergetics Modeling in the 21st Century: Reviewing New Insights and Revisiting Old Constraints. Trans. Am. Fish. Soc. 2008, 137, 298–313. [Google Scholar] [CrossRef]
- Ito, S.-I.; Kishi, M.J.; Kurita, Y.; Oozeki, Y.; Yamanaka, Y.; Megrey, B.A.; Werner, F.E. Initial design for a fish bioenergetics model of Pacific saury coupled to a lower trophic ecosystem model. Fish. Oceanogr. 2004, 13, 111–124. [Google Scholar] [CrossRef]
- Oyaizu, H.; Suyama, S.; Ambe, D.; Ito, S.; Itoh, S. Modeling the growth, transport, and feeding migration of age-0 Pacific saury Cololabis saira. Fish. Sci. 2022, 88, 131–147. [Google Scholar] [CrossRef]
- Compagno, L.J.V. Relationships of the megamouth shark, Megachasma pelagios (Lamniformes: Megachasmidae), with comments on its feeding habits. NOAA Tech. Rep. NMFS 1990, 90, 357–379. [Google Scholar]
- Cortés, E. Life history patterns and correlations in sharks. Rev. Fish. Sci. 2000, 8, 299–344. [Google Scholar] [CrossRef]
- Jaureguizar, A.J.; De Wysiecki, A.M.; Camiolo, M.D. Environmental influence on the inter-annual demographic variation of the narrownose smooth-hound shark (Mustelus schmitti, Springer 1939) in the Northern Argentine Coastal System (El Rincón, 38–42°S). Mar. Biol. Res. 2020, 16, 600–615. [Google Scholar] [CrossRef]
- Cortés, F.; Jaureguizar, A.J.; Menni, R.C.; Guerrero, R.A. Ontogenetic habitat preferences of the narrownose smooth-hound shark, Mustelus schmitti, in two Southwestern Atlantic coastal areas. Hydrobiologia 2011, 661, 445–456. [Google Scholar] [CrossRef]
- Colautti, D.; Baigun, C.; Cazorla, A.L.; Llompart, F.; Molina, J.M.; Suquele, P.; Calvo, S. Population biology and fishery characteristics of the smooth-hound Mustelus schmitti in Anegada Bay, Argentina. Fish. Res. 2010, 106, 351–357. [Google Scholar] [CrossRef]
- De Wysiecki, A.M.; Jaureguizar, A.J.; Cortés, F. The importance of environmental drivers on the narrownose smoothhound shark (Mustelus schmitti) yield in a small-scale gillnet fishery along the southern boundary of the Río de la Plata estuarine area. Fish. Res. 2017, 186, 345–355. [Google Scholar] [CrossRef]
- Pollom, R.; Barreto, R.; Charvet, P.; Chiaramonte, G.E.; Cuevas, J.M.; Herman, K.; Montealegre-Quijano, S.; Motta, F.; Paesch, L.; Rincon, G. Mustelus schmitti. The IUCN Red List of Threatened Species 2020, e.T60203A3092243. 2020. Available online: https://www.iucnredlist.org/species/60203/3092243 (accessed on 1 January 2023).
- Pérez, M. Estimación de un índice de abundancia anual de gatuzo (Mustelus schmitti) a partir de datos de la flota comercial argentina. Período 1992–2008. INIDEP 2010, 79, 41. [Google Scholar]
- Molina, J.M.; Cazorla, A.L. Trophic ecology of Mustelus schmitti (Springer, 1939) in a nursery area of northern Patagonia. J. Sea Res. 2011, 65, 381–389. [Google Scholar] [CrossRef]
- Fiori, S.; Lopez Cazorla, A.; Martínez, A.; Carcedo, M.C.; Blasina, G.E.; Molina, J.M.; Garzón Cardona, J.; Moyano, J.; Menéndez, M.C. Life in the surf: Faunal assemblage diversity and distribution in temperate sandy beaches of the South American Atlantic. Cont. Shelf Res. 2022, 244, 104781. [Google Scholar] [CrossRef]
- Molina, J.M.; Blasina, G.E.; Lopez Cazorla, A.C. Age and growth of the highly exploited narrownose smooth-hound (Mustelus schmitti) (Pisces: Elasmobranchii). Fish. Bull. Natl. Ocean. Atmos. Adm. 2012, 115, 365–379. [Google Scholar] [CrossRef]
- Elisio, M.; Colonello, J.H.; Cortés, F.; Jaureguizar, A.J.; Somoza, G.M.; Macchi, G.J. Aggregations and reproductive events of the narrownose smooth-hound shark (Mustelus schmitti) in relation to temperature and depth in coastal waters of the south-western Atlantic Ocean (38–42° S). Mar. Freshwater Res. 2016, 68, 732–742. [Google Scholar] [CrossRef]
- Elisio, M.; Awruch, C.A.; Massa, A.M.; Macchi, G.J.; Somoza, G.M. Effects of temperature on the reproductive physiology of female elasmobranchs: The case of the narrownose smooth-hound shark (Mustelus schmitti). Gen. Comp. Endocrinol. 2019, 284, 113242. [Google Scholar] [CrossRef]
- Chiaramonte, G.E.; Pettovello, A.D. The biology of Mustelus schmitti in southern Patagonia, Argentina. J. Fish Biol. 2000, 57, 930–942. [Google Scholar] [CrossRef]
- Segura, A.M.; Milessi, A.C. Biological and reproductive characteristics of the Patagonian smoothhound Mustelus schmitti (Chondrichthyes, Triakidae) as documented from an artisanal fishery in Uruguay. J. Appl. Ichthyol. 2009, 25, 78–82. [Google Scholar] [CrossRef]
- Hozbor, N.M.; Saez, M.; Massa, A. Edad y crecimiento de Mustelus schmitti (gatuzo), en la región costera bonaerense y uruguaya. INIDEP 2010, 49, 1–14. [Google Scholar]
- Belleggia, M.; Figueroa, D.E.; Sánchez, F.; Bremec, C. The feeding ecology of Mustelus schmitti in the southwestern Atlantic: Geographic variations and dietary shifts. Environ. Biol. Fishes 2012, 95, 99–114. [Google Scholar] [CrossRef]
- Galíndez, E.J.; Díaz Andrade, M.; Estecondo, S. Morphological indicators of initial reproductive commitment in Mustelus schmitti (Springer 1939)(Chondrichthyes, Triakidae): Folliculogenesis and ovarian structure over the life cycle. Braz. J. Biol. 2014, 74, S154–S163. [Google Scholar] [CrossRef] [PubMed]
- Jaureguizar, A.J.; Wiff, R.; Clara, M.L. Role of the preferred habitat availability for small shark (Mustelus schmitti) on the interannual variation of abundance in a large Southwest Atlantic Coastal System (El Rincón, 39–41° S). Aquat. Living Resour. 2016, 29, 305. [Google Scholar] [CrossRef]
- Pasti, A.T.; Bovcon, N.D.; Ruibal-Nunez, J.; Navoa, X.; Jacobi, K.J.; Galvan, D.E. The diet of Mustelus schmitti in areas with and without commercial bottom trawling (Central Patagonia, Southwestern Atlantic): Is it evidence of trophic interaction with the Patagonian shrimp fishery? Food Webs 2021, 29, e00214. [Google Scholar] [CrossRef]
- Fuller, A.; Dawson, T.; Helmuth, B.; Hetem, R.; Mitchell, D.; Maloney, S. Physiological Mechanisms in Coping with Climate Change. Physiol. Biochem. Zool. 2010, 83, 713–720. [Google Scholar] [CrossRef]
- Hartman, K.; Cox, M. Refinement and Testing of a Brook Trout Bioenergetics Model. Trans. Am. Fish. Soc. 2008, 137, 357–363. [Google Scholar] [CrossRef]
- Kummu, M.; Sarkkula, J.; Koponen, J.; Nikula, J. Ecosystem Management of the Tonle Sap Lake: An Integrated Modelling Approach. Int. J. Water Resour. Dev. 2006, 22, 497–519. [Google Scholar] [CrossRef]
- Rudstam, L.G. Exploring the dynamics of herring consumption in the Baltic: Applications of an energetic model of fish growth. Kiel. Meeresforsch. Sonderh. 1988, 6, 312–322. [Google Scholar]
- Beauchamp, D.A.; Stewart, D.J.; Thomas, G.L. Corroboration of a bioenergetics model for sockeye salmon. Trans. Am. Fish. Soc. 1989, 118, 597–607. [Google Scholar] [CrossRef]
- Ware, D.M. Bioenergetics of pelagic fish: Theoretical change in swimming speed and ration with body size. J. Fish. Res. Board Can. 1978, 35, 220–228. [Google Scholar] [CrossRef]
- Trudel, M.; Geist, D.R.; Welch, D.W. Modeling the oxygen consumption rates in Pacific salmon and steelhead: An assessment of current models and practices. Trans. Am. Fish. Soc. 2004, 133, 326–348. [Google Scholar] [CrossRef]
- Kamezawa, M.; Azumaya, T.; Nagasawa, T.; Kishi, M.J. A fish bioenergetics model of Japanese chum salmon (Oncorhynchus keta) for studying the influence of environmental factor changes. Bull. Jpn. Soc. Fish. Oceanogr. 2007, 71, 87–95. [Google Scholar]
- Molina, J.M.; Cazorla, A.L. Biology of Myliobatis goodei (Springer, 1939), a widely distributed eagle ray, caught in northern Patagonia. J. Sea Res. 2015, 95, 106–114. [Google Scholar] [CrossRef]
- Elisio, M.; Maenza, R.A.; Luz Clara, M.; Baldoni, A.G. Modeling the bottom temperature variation patterns on a coastal marine ecosystem of the Southwestern Atlantic Ocean (El Rincón), with special emphasis on thermal changes affecting fish populations. J. Mar. Syst. 2020, 212, 103445. [Google Scholar] [CrossRef]
- Molina, J.M.; Blasina, G.; Cazorla, A.L. Ecology and Biology of Fish Assemblages. In The Bahía Blanca Estuary; Springer: Berlin/Heidelberg, Germany, 2021; pp. 275–306. [Google Scholar]
- He, E.; Wurtsbaugh, W.A. An Empirical Model of Gastric Evacuation Rates for Fish and an Analysis of Digestion in Piscivorous Brown Trout. Trans. Am. Fish. Soc. 1993, 122, 717–730. [Google Scholar] [CrossRef]
- Brown, S.C.; Bizzarro, J.J.; Cailliet, G.M.; Ebert, D.A. Breaking with tradition: Redefining measures for diet description with a case study of the Aleutian skate Bathyraja aleutica (Gilbert 1896). Environ. Biol. Fishes 2012, 95, 3–20. [Google Scholar] [CrossRef]
- Sims, D.W.; Davies, S.J.; Bone, Q. Gastric Emptying Rate and Return Of Appetite In Lesser Spotted Dogfish, Scyliorhinus canicula (Chondrichthyes: Elasmobranchii). J. Mar. Biol. Assoc. U. K. 1996, 76, 479–491. [Google Scholar] [CrossRef]
- Wetherbee, B.M.; Cortés, E.; Bizzarro, J.J. Food consumption and feeding habits. In Biology of Sharks and Their Relatives, 2nd ed.; Carrier, J.C., Musick, J.A., Heithaus, M.R., Eds.; CRC Press: Boca Raton, FL, USA, 2012; pp. 225–246. [Google Scholar] [CrossRef]
- Thornton, K.W.; Lessem, A.S. A temperature algorithm for modifying biological rates. Trans. Am. Fish. Soc. 1978, 107, 284–287. [Google Scholar] [CrossRef]
- Molina, J.M. La Comunidad Íctica de Bahía Anegada: Estructura, Composición, Dinámica Estacional y Aspectos Biológicos. Ph.D. Thesis, National University of the South, Bahia Blanca, Argentina, 2012; 234p. [Google Scholar]
- Orlando, L.; Pereyra, I.; Silveira, S.; Paesch, L.; Oddone, M.C.; Norbis, W. Determination of limited histotrophy as the reproductive mode in Mustelus schmitti Springer, 1939 (Chondrichthyes: Triakidae): Analysis of intrauterine growth of embryos. Neotrop. Ichthyol. 2015, 13, 699–706. [Google Scholar] [CrossRef]
- Quesada, G. Dinámica Energética Reproductiva en el Gatuzo (Mustelus schmitti). Implicancias Sobre la Variación Anual en el Rendimiento de Carne. Bachelor’s Thesis, Universidad Nacional de Mar del Plata, Mar del Plata, Argentina, 2018. [Google Scholar]
- Cousseau, M.B.; Carozza, C.R.; Macchi, G.J. Abundancia, reproducción y distribucion de tallas del gatuzo Mustelus schmitti en la Zona Comun de Pesca Argentino-Uruguaya y en el Rincón. Noviembre, 1994. INIDEP Inf. Técnico 1998, 21, 103–115. [Google Scholar]
- Casselberry, G.A.; Carlson, J.K. Endangered Species Act Review of the Narrownose Smoothhound (Mustelus schmitti); Report to the National Marine Fisheries Service, Office of Protected Resources; SFD Contribution PCB-15-07; NOAA: Washington, DC, USA, 2015.
- Paesch, L. Índices de abundancia para Mustelus schmitti, Squatina guggenheim y Zearaja chilensis en la Zona Común de Pesca Argentino-Uruguaya. INIDEP 2018, 56, 1–32. [Google Scholar]
- Bethea, D.M.; Hale, L.; Carlson, J.K.; Cortés, E.; Manire, C.A.; Gelsleichter, J. Geographic and ontogenetic variation in the diet and daily ration of the bonnethead shark, Sphyrna tiburo, from the eastern Gulf of Mexico. Mar. Biol. 2007, 152, 1009–1020. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 1 January 2023).
- Cortés, E. Incorporating uncertainty into demographic modeling: Application to shark populations and their conservation. Conserv. Biol. 2002, 16, 1048–1062. [Google Scholar] [CrossRef]
- Simpfendorfer, C.A.; Hueter, R.E.; Bergman, U.; Connett, S.M. Results of a fishery-independent survey for pelagic sharks in the western North Atlantic, 1977–1994. Fish. Res. 2002, 55, 175–192. [Google Scholar] [CrossRef]
- Simpfendorfer, C.A.; Heupel, M.R.; Hueter, R.E. Estimation of short-term centers of activity from an array of omnidirectional hydrophones and its use in studying animal movements. Can. J. Fish. Aquat. Sci. 2002, 59, 23–32. [Google Scholar] [CrossRef]
- Dulvy, N.K.; Baum, J.K.; Clarke, S.; Compagno, L.J.; Cortés, E.; Domingo, A.; Fordham, S.; Fowler, S.; Francis, M.P.; Gibson, C.; et al. You can swim but you can’t hide: The global status and conservation of oceanic pelagic sharks and rays. Aquat. Conserv. Mar. Freshw. Ecosyst. 2004, 14, 459–482. [Google Scholar] [CrossRef]
- Frisk, M.; Miller, T.; Dulvy, N. Life histories and vulnerability to exploitation of elasmobranchs: Inferences from elasticity, perturbation and phylogenetic analyses. J. Northwest Atl. Fish. Sci. 2014, 46, 27–45. [Google Scholar] [CrossRef]
- Hussey, N.E.; Kessel, S.T.; Aarestrup, K.; Cooke, S.J.; Cowley, P.D.; Fisk, A.T.; Harcourt, R.G.; Holland, K.N.; Iverson, S.J.; Kocik, J.F.; et al. Aquatic animal telemetry: A panoramic window into the underwater world. Science 2015, 348, 1255642. [Google Scholar] [CrossRef]
- Roff, G.; Doropoulos, C.; Rogers, A.; Bozec, Y.M.; Krueck, N.C.; Aurellado, E.; Priest, M.; Birrell, C.; Mumby, P.J. The ecological role of sharks on coral reefs. Trends Ecol. Evol. 2016, 31, 395–407. [Google Scholar] [CrossRef]
- Queiroz, N.; Humphries, N.E.; Couto, A.; Vedor, M.; Da Costa, I.; Sequeira, A.M.; Mucientes, G.; Santos, A.M.; Abascal, F.J.; Abercrombie, D.L.; et al. Global spatial risk assessment of sharks under the footprint of fisheries. Nature 2019, 572, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Pirotta, E. A review of bioenergetic modelling for marine mammal populations. Conserv. Physiol. 2022, 10, coac036. [Google Scholar] [CrossRef] [PubMed]
- Beukers-Stewart, B.; Jones, G. The influence of prey abundance on the feeding ecology of two piscivorous species of coral reef fish. J. Exp. Mar. Biol. Ecol. 2004, 299, 155–184. [Google Scholar] [CrossRef]
- Queiroz, N.; Humphries, N.E.; Mucientes, G.; Hammerschlag, N.; Lima, F.P.; Scales, K.L.; Miller, P.I.; Sousa, L.L.; Seabra, R.; Sims, D.W. Ocean-wide tracking of pelagic sharks reveals extent of overlap with longline fishing hotspots. Proc. Natl. Acad. Sci. USA 2016, 113, 1582–1587. [Google Scholar] [CrossRef]
- McClintock, J. On estimating energetic values of prey: Implications in optimal diet models. Oecologia 1986, 70, 161–162. [Google Scholar] [CrossRef]
- Finstad, A.; Forseth, T.; Ugedal, O.; Næsje, T. Metabolic rate, behaviour and winter performance in juvenile Atlantic salmon. Funct. Ecol. 2007, 21, 905–912. [Google Scholar] [CrossRef]
- Hussey, N.E.; MacNeil, M.A.; McMeans, B.C.; Olin, J.A.; Dudley, S.F.; Cliff, G.; Wintner, S.P.; Fennessy, S.T.; Fisk, A.T. Rescaling the trophic structure of marine food webs. Ecol. Lett. 2014, 17, 239–250. [Google Scholar] [CrossRef]
- Dulvy, N.K.; Reynolds, J.D. Evolutionary transitions among egg-laying, live-bearing and maternal inputs in sharks and rays. Proc. R. Soc. London. Ser. B Biol. Sci. 1997, 264, 1309–1315. [Google Scholar] [CrossRef]
- Cuadrado, D.G.; Gómez, E.A. Geomorfología y dinámica del canal San Blas, Provincia de Buenos Aires (Argentina). Lat. Am. J. Sedimentol. Basin Anal. 2010, 17, 3–16. [Google Scholar]
- Croce, M.E.; Parodi, E.R. The Japanese alga Polysiphonia morrowii (Rhodomelaceae, Rhodophyta) on the South Atlantic Ocean: First report of an invasive macroalga inhabiting oyster reefs. Helgol. Mar. Res. 2014, 68, 241–252. [Google Scholar] [CrossRef]
- Heithaus, M.R. Nursery areas as essential shark habitats: A theoretical perspective. In American Fisheries Society Symposium; American Fisheries Society: Bethesda, MD, USA, 2007; Volume 50, p. 3. [Google Scholar]
- Cuevas, J.M.; y Michelson, A.M. Estado Actual del Conocimiento Sobre Condrictios en la Reserva Natural de Usos Múltiples Bahía San Blas, Provincia de Buenos Aires. 2023. Available online: https://argentina.wcs.org/ (accessed on 20 August 2023).
- Myers, R.A.; Worm, B. Rapid worldwide depletion of predatory fish communities. Nature 2003, 423, 280–283. [Google Scholar] [CrossRef]
- Block, B.; Jonsen, I.; Jorgensen, S.; Winship, A.; Shaffer, S.; Bograd, S.; Hazen, E.; Foley, D.; Breed, G.; Harrison, A.; et al. Tracking apex marine predator movements in a dynamic ocean. Nature 2011, 475, 86–90. [Google Scholar] [CrossRef]
- Gallagher, A.J.; Skubel, R.A.; Pethybridge, H.R.; Hammerschlag, N. Energy metabolism in mobile, wild-sampled sharks inferred by plasma lipids. Conserv. Physiol. 2017, 5, cox002. [Google Scholar] [CrossRef]
- Pörtner, H.O. Climate variations and the physiological basis of temperature dependent biogeography: Systemic to molecular hierarchy of thermal tolerance in animals. Comparative Biochem. Physiol. Part A Mol. Integr. Physiol. 2002, 132, 739–761. [Google Scholar] [CrossRef]
- Cheung, W.W.; Lam, V.W.; Sarmiento, J.L.; Kearney, K.; Watson, R.; Pauly, D. Projecting global marine biodiversity impacts under climate change scenarios. Fish Fish. 2009, 10, 235–251. [Google Scholar] [CrossRef]
- Perry, A.L.; Low, P.J.; Ellis, J.R.; Reynolds, J.D. Climate change and distribution shifts in marine fishes. Science 2005, 308, 1912–1915. [Google Scholar] [CrossRef]
- Staudinger, M.D.; Grimm, N.B.; Staudt, A.; Carter, S.L.; Chapin, F.S. Impacts of climate change on biodiversity, ecosystems, and ecosystem services. United States Glob. Chang. Res. Program Wash. DC 2012, 139, 205. [Google Scholar]
- Sunday, J.M.; Bates, A.E.; Dulvy, N.K. Global analysis of thermal tolerance and latitude in ectotherms. Proc. R. Soc. B Biol. Sci. 2011, 278, 1823–1830. [Google Scholar] [CrossRef]
- Rennie, M.; Purchase, C.; Shuter, B.; Collins, N.; Abrams, P.; Morgan, G. Prey life-history and bioenergetic responses across a predation gradient. J. Fish Biol. 2010, 77, 1230–1251. [Google Scholar] [CrossRef]
- Gasalla, M.A.; Abdallah, P.R.; Lemos, D. Potential impacts of climate change in Brazilian marine fisheries and aquaculture. Clim. Chang. Impacts Fish. Aquac. A Glob. Anal. 2017, 1, 455–477. [Google Scholar]
- Jennings, S.; Brander, K. Predicting the effects of climate change on marine communities and the consequences for fisheries. J. Mar. Syst. 2010, 79, 418–426. [Google Scholar] [CrossRef]
- Beck, M.W.; Heck, K.L.; Able, K.W.; Childers, D.L.; Eggleston, D.B.; Gillanders, B.M.; Halpern, B.; Hays, C.G.; Hoshino, K.; Minello, T.J.; et al. The identification, conservation, and management of estuarine and marine nurseries for fish and invertebrates: A better understanding of the habitats that serve as nurseries for marine species and the factors that create site-specific variability in nursery quality will improve conservation and management of these areas. Bioscience 2001, 51, 633–641. [Google Scholar]
- Heupel, M.; Simpfendorfer, C.; Hueter, R. Estimation of Shark Home Ranges using Passive Monitoring Techniques. Environ. Biol. Fishes 2004, 71, 135–142. [Google Scholar] [CrossRef]
- Lucifora, L.O.; Menni, R.C.; Escalante, A.H. Reproduction, abundance and feeding habits of the broadnose sevengill shark Notorynchus cepedianus in north Patagonia, Argentina. Mar. Ecol. Prog. Ser. 2005, 289, 237–244. [Google Scholar] [CrossRef]
- Bland, S.; Valdovinos, F.S.; Hutchings, J.A.; Kuparinen, A. The role of fish life histories in allometrically scaled food-web dynamics. Ecol. Evol. 2019, 9, 3651–3660. [Google Scholar] [CrossRef]
- Funes, M.; De Wysiecki, A.M.; Bovcon, N.D.; Jaureguizar, A.J.; Irigoyen, A.J. One marine protected area is not enough: The trophic ecology of the broadnose sevengill shark (Notorynchus cepedianus) in the Southwest Atlantic. bioRxiv 2023. [Google Scholar] [CrossRef]
- Cheung, W.W.; Lam, V.W.; Sarmiento, J.L.; Kearney, K.; Watson, R.E.; Zeller, D.; Pauly, D. Large-scale redistribution of maximum fisheries catch potential in the global ocean under climate change. Glob. Chang. Biol. 2012, 18, 1439–1459. [Google Scholar] [CrossRef]
- Pinsky, M.L.; Reygondeau, G.; Caddell, R.; Palacios-Abrantes, J.; Spijkers, J.; Cheung, W.W. Preparing ocean governance for species on the move. Science 2013, 360, 1189–1191. [Google Scholar] [CrossRef]
- Weatherdon, L.V.; Magnan, A.K.; Rogers, A.D.; Sumaila, U.R.; Cheung, W.W. Observed and projected impacts of climate change on marine fisheries, aquaculture, coastal tourism, and human health: An update. Front. Mar. Sci. 2016, 3, 48. [Google Scholar] [CrossRef]
- Barneche, D.R.; Robertson, D.R.; White, C.R.; Marshall, D.J. Fish reproductive-energy output increases disproportionately with body size. Science 2018, 360, 642–645. [Google Scholar] [CrossRef]
- Gervais, C.R.; Nay, T.J.; Renshaw, G.; Johansen, J.L.; Steffensen, J.F.; Rummer, J.L. Too hot to handle? Using movement to alleviate effects of elevated temperatures in a benthic elasmobranch, Hemiscyllium ocellatum. Mar. Biol. 2018, 165, 162. [Google Scholar] [CrossRef]
- Cheung, W.W.; Sarmiento, J.L.; Dunne, J.; Frölicher, T.L.; Lam, V.W.; Deng Palomares, M.L.; Watson, R.; Pauly, D. Shrinking of fishes exacerbates impacts of global ocean changes on marine ecosystems. Nat. Clim. Chang. 2013, 3, 254–258. [Google Scholar] [CrossRef]
- García Molinos, J.; Halpern, B.S.; Schoeman, D.S.; Brown, C.J.; Kiessling, W.; Moore, P.J.; Pandolfi, J.M.; Poloczanska, E.S.; Richardson, A.J.; Burrows, M.T. Climate velocity and the future global redistribution of marine biodiversity. Nat. Clim. Chang. 2015, 6, 83–88. [Google Scholar] [CrossRef]
- Olden, J.D.; Neff, B.D. Cross-correlation bias in lag analysis of aquatic time series. Mar. Biol. 2001, 138, 1063–1070. [Google Scholar] [CrossRef]
- Trujillo, A.P.; Thurman, H.V. Essentials of Oceanography, 12th ed.; Pearson Education, Inc.: Boston, MA, USA, 2016; pp. 403–444. [Google Scholar]
- Wang, L.; Kerr, L.A.; Record, N.R.; Bridger, E.; Tupper, B.; Mills, K.E.; Armstrong, E.M.; Pershing, A.J. Modeling marine pelagic fish species spatiotemporal distributions utilizing a maximum entropy approach. Fish. Oceanogr. 2018, 27, 571–586. [Google Scholar] [CrossRef]
- Hutchings, J. Conservation biology of marine fishes: Perceptions and caveats regarding assignment of extinction risk. Can. J. Fish. Aquat. Sci. 2001, 58, 108–121. [Google Scholar] [CrossRef]
- Brown, C.J.; Fulton, E.A.; Hobday, A.J.; Matear, R.J.; Possingham, H.P.; Bulman, C.; Christensen, V.; Forrest, R.E.; Gehrke, P.C.; Gribble, N.A. Effects of climate-driven primary production change on marine food webs: Implications for fisheries and conservation. Glob. Chang. Biol. 2010, 16, 1194–1212. [Google Scholar] [CrossRef]
- Morris, J.A.; Akins, J.L. Feeding ecology of invasive lionfish (Pterois volitans) in the Bahamian archipelago. Environ. Biol. Fishes 2009, 86, 389–398. [Google Scholar] [CrossRef]
- Zhao, Q.; Arnold, T.; Devries, J.; Howerter, D.; Clark, R.; Weegman, M. Using integrated population models to prioritize region-specific conservation strategies under global change. Biol. Conserv. 2020, 252, 108832. [Google Scholar] [CrossRef]
- Koen-Alonso, M.; Lindstrøm, U.; Cuff, A. Comparative modeling of cod-capelin dynamics in the Newfoundland-Labrador shelves and Barents Sea ecosystems. Front. Mar. Sci. 2021, 8, 579946. [Google Scholar] [CrossRef]
- Martin, R.A.; Rossmo, D.K.; Hammerschlag, N. Hunting patterns and geographic profiling of white shark predation. J. Zool. 2009, 279, 111–118. [Google Scholar] [CrossRef]
- Brena, P.F.; Mourier, J.; Planes, S.; Clua, E.E. Concede or clash? Solitary sharks competing for food assess rivals to decide. Proc. R. Soc. B Biol. Sci. 2018, 285. [Google Scholar] [CrossRef] [PubMed]
- Molina, J.M.; Kunzmann, A.; Reis, J.P.; Guerreiro, P.M. Metabolic Responses and Resilience to Environmental Challenges in the Sedentary Batrachoid Halobatrachus didactylus (Bloch & Schneider, 1801). Animals 2023, 13, 632. [Google Scholar]
- Fordham, D.A.; Wigley, T.M.; Brook, B.W. Multi-model climate projections for biodiversity risk assessments. Ecol. Appl. 2011, 21, 3317–3331. [Google Scholar] [CrossRef]
- Horodysky, A.Z.; Cooke, S.J.; Graves, J.E.; Brill, R.W. Fisheries conservation on the high seas: Linking conservation physiology and fisheries ecology for the management of large pelagic fishes. Conserv. Physiol. 2016, 4, cov059. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter Name | Parameter Description and Symbol | Value | Sampling Distribution | Units | Sources |
|---|---|---|---|---|---|
| Initial weight | W | 200 | Normal (µ = 200, σ = 35) | g | [26,32,58] |
| Consumption | Cmax intercept (ac) | 0.28 | Log-normal (log-µ = 0.28, log-σ = 0.2) | gprey/gshark/day | [30,38,58] |
| Cmax/weight exponent (bc) | 0.3 | - | unitless | [30,38,58] | |
| R intercept | 0.016 | - | unitless | [53] | |
| R constant | 0.06 | - | unitless | [53] | |
| Respiration | MR/weight intercept (ar) | 0.256 | Log-normal (log-µ = 0.256, log-σ = 0.1) | gO2/gshark/day | [15] |
| MR/weight exponent (br) | −0.38 | - | unitless | [15] | |
| Routine metabolic rate | RMR | 0.208 | Log-normal (log-µ = 0.208, log-σ = 0.17) | gO2/kg/day | [15] |
| Standard metabolic rate | SMR | 0.168 | Log-normal (log-µ = 0.168, log-σ = 0.13) | gO2/kg/day | [15] |
| Maximum temperature for respiration | RTM | 22 | - | °C | [15,27] |
| Temperature effect on respiration | RQ10 | 2.3 | - | unitless | [15] |
| Specific dynamic action | sda | 0.3 | Normal (µ = 0.3, σ = 0.3) | unitless | [8] |
| Egestion | Proportion of consumption (peg) | 0.2 | Normal (µ = 0.2, σ = 0.03) | proportion | [8] |
| Excretion | Proportion of consumption (pex) | 0.08 | Normal (µ = 0.08, σ = 0.04) | proportion | [8] |
| Effect of temperature on consumption | Proportion C 1 (xk1) | 0.1 | - | proportion | [57] |
| Proportion C 2 (xk2) | 0.85 | - | proportion | [57] | |
| Proportion C 3 (xk3) | 0.98 | - | proportion | [57] | |
| Proportion C 4 (xk4) | 0.2 | - | proportion | [57] | |
| T° 1 (te1) | 7 | - | °C | [40] | |
| T° 2 (te2) | 10 | - | °C | [40] | |
| T° 3 (te3) | 14.9 | - | °C | [24] | |
| T° 4 (te4) | 22 | - | °C | [40] | |
| Fish caloric density | CALS | 1294 | - | cal/gshark | [15] |
| Prey (Polychaetes) caloric density | CALp1 | 991 | - | cal/gcrab | [8] |
| Prey (Crabs) caloric density | CALp2 | 1300 | - | cal/gcrab | [8] |
| Prey availability | Cden1 and Cdem2 | Uniform (+/−20% of model output) | [30,58] |
| Growth | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Baseline | Moderate | Severe | |||||||||
| Age | Mean | CI | Rate | Mean | CI | Rate | %D | Mean | CI | Rate | %D |
| 0+ | 220.38 | 38.58 | 0.60 | 205.84 | 35.81 | 0.56 | 6.598 | 196.59 | 28.92 | 0.54 | 10.79 |
| 1 | 288.94 | 49.84 | 0.79 | 268.17 | 47.85 | 0.73 | 7.189 | 233.84 | 38.40 | 0.64 | 19.07 |
| 2 | 365.84 | 59.85 | 1.00 | 336.99 | 60.16 | 0.92 | 7.886 | 274.00 | 48.10 | 0.75 | 25.10 |
| 3 | 446.94 | 75.89 | 1.22 | 406.55 | 77.78 | 1.11 | 9.037 | 318.43 | 61.91 | 0.87 | 28.75 |
| 4 | 538.43 | 94.33 | 1.48 | 483.40 | 94.86 | 1.32 | 10.22 | 362.98 | 74.07 | 0.99 | 32.59 |
| 5 | 634.50 | 111.29 | 1.74 | 566.68 | 108.95 | 1.55 | 10.69 | 409.42 | 96.83 | 1.12 | 35.47 |
| 6 | 740.46 | 131.78 | 2.03 | 658.24 | 131.86 | 1.80 | 11.1 | 469.49 | 108.93 | 1.29 | 36.59 |
| 7 | 846.12 | 152.58 | 2.32 | 747.54 | 147.97 | 2.05 | 11.65 | 527.51 | 118.58 | 1.45 | 37.66 |
| Consumption | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Baseline | Moderate | Severe | |||||||||
| Age | Mean | CI | KPY | Mean | CI | KPY | %D | Mean | CI | KPY | %D |
| 0+ | 0.35 | 0.32 | 28.08 | 0.38 | 0.19 | 30.35 | 8.065 | 0.37 | 0.19 | 29.99 | 6.79 |
| 1 | 0.40 | 0.35 | 41.75 | 0.40 | 0.21 | 42.30 | 1.315 | 0.45 | 0.22 | 47.14 | 12.91 |
| 2 | 0.41 | 0.36 | 54.66 | 0.50 | 0.23 | 66.20 | 21.12 | 0.57 | 0.25 | 75.98 | 39.02 |
| 3 | 0.45 | 0.41 | 73.63 | 0.54 | 0.26 | 87.67 | 19.06 | 0.57 | 0.26 | 92.67 | 25.85 |
| 4 | 0.48 | 0.42 | 94.89 | 0.54 | 0.27 | 105.52 | 11.21 | 0.53 | 0.29 | 104.52 | 10.15 |
| 5 | 0.50 | 0.45 | 115.88 | 0.57 | 0.29 | 135.57 | 13.83 | 0.59 | 0.31 | 137.62 | 18.76 |
| 6 | 0.55 | 0.45 | 149.47 | 0.57 | 0.30 | 186.93 | 3.693 | 0.69 | 0.32 | 186.93 | 25.06 |
| 7 | 0.52 | 0.46 | 160.62 | 0.59 | 0.31 | 177.10 | 12.56 | 0.62 | 0.34 | 192.55 | 19.88 |
| Energy Requirements | ||||||||
|---|---|---|---|---|---|---|---|---|
| Baseline | Moderate | Severe | ||||||
| Age | Daily (cal) | Anual (Kcal) | Daily (cal) | Anual (Kcal) | %D | Daily (cal) | Anual (Kcal) | %D |
| 0+ | 30.14 | 2424.33 | 47.16 | 3543.39 | 46.16 | 71.16 | 5106.36 | 110.63 |
| 1 | 26.52 | 2796.98 | 40.71 | 3985.02 | 42.48 | 61.11 | 5215.64 | 86.47 |
| 2 | 23.97 | 3200.79 | 35.78 | 4400.65 | 37.49 | 53.63 | 5363.97 | 67.58 |
| 3 | 23.05 | 3759.42 | 33.80 | 5015.48 | 33.41 | 50.94 | 5920.70 | 57.49 |
| 4 | 21.10 | 4147.55 | 30.97 | 5463.91 | 31.74 | 46.79 | 6199.20 | 49.47 |
| 5 | 19.74 | 4572.15 | 28.55 | 5905.91 | 29.17 | 43.40 | 6486.16 | 41.86 |
| 6 | 16.75 | 4528.16 | 23.86 | 5731.63 | 26.58 | 35.98 | 6165.39 | 36.16 |
| 7 | 17.41 | 5377.22 | 24.47 | 6678.03 | 24.19 | 37.29 | 7179.42 | 33.52 |
| Stock Abundances | Prey Consumed (t) | Daily Ration (%W) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Age | Proportion | Abundance (t) | N estimated | Baseline | Moderate | Severe | Baseline | Moderate | Severe |
| 0+ | 0.16 | 13,513.5 | 61,319,230.0 | 172,200.4 | 186,088.6 | 183,895.8 | 3.5 | 3.8 | 3.7 |
| 1 | 0.31 | 26,107.0 | 90,354,691.3 | 377,199.5 | 382,160.0 | 425,913.1 | 4.0 | 4.0 | 4.5 |
| 2 | 0.20 | 16,906.3 | 46,212,393.1 | 252,585.9 | 305,935.6 | 351,136.1 | 4.1 | 5.0 | 5.7 |
| 3 | 0.16 | 13,456.0 | 30,107,273.2 | 221,690.6 | 263,953.9 | 279,003.7 | 4.5 | 5.4 | 5.7 |
| 4 | 0.08 | 7015.5 | 13,029,665.2 | 123,636.9 | 137,492.5 | 136,180.8 | 4.8 | 5.4 | 5.3 |
| 5 | 0.03 | 2242.7 | 3,534,540.2 | 40,957.5 | 47,918.6 | 48,641.6 | 5.0 | 5.9 | 5.9 |
| 6 | 0.03 | 2300.2 | 3,106,390.1 | 46,431.1 | 58,066.3 | 58,066.3 | 5.5 | 6.9 | 6.9 |
| 7 | 0.02 | 1380.1 | 1,631,103.4 | 26,198.2 | 28,887.3 | 31,407.5 | 5.2 | 5.7 | 6.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molina, J.M.; Yoon, S.; Elisio, M.; Kasai, A. Bioenergetic Model of the Highly Exploited Shark Mustelus schmitti under a Global Warming Context. Diversity 2023, 15, 1118. https://doi.org/10.3390/d15111118
Molina JM, Yoon S, Elisio M, Kasai A. Bioenergetic Model of the Highly Exploited Shark Mustelus schmitti under a Global Warming Context. Diversity. 2023; 15(11):1118. https://doi.org/10.3390/d15111118
Chicago/Turabian StyleMolina, Juan Manuel, Seokjin Yoon, Mariano Elisio, and Akihide Kasai. 2023. "Bioenergetic Model of the Highly Exploited Shark Mustelus schmitti under a Global Warming Context" Diversity 15, no. 11: 1118. https://doi.org/10.3390/d15111118
APA StyleMolina, J. M., Yoon, S., Elisio, M., & Kasai, A. (2023). Bioenergetic Model of the Highly Exploited Shark Mustelus schmitti under a Global Warming Context. Diversity, 15(11), 1118. https://doi.org/10.3390/d15111118