
Patterns of Diversity, Structure and Local Ecology of Arthropod-Pathogenic Fungi in the Amazonian Forest of Cusco and Madre de Dios Regions, Southern Peru

Abstract
:1. Introduction
2. Materials and Methods
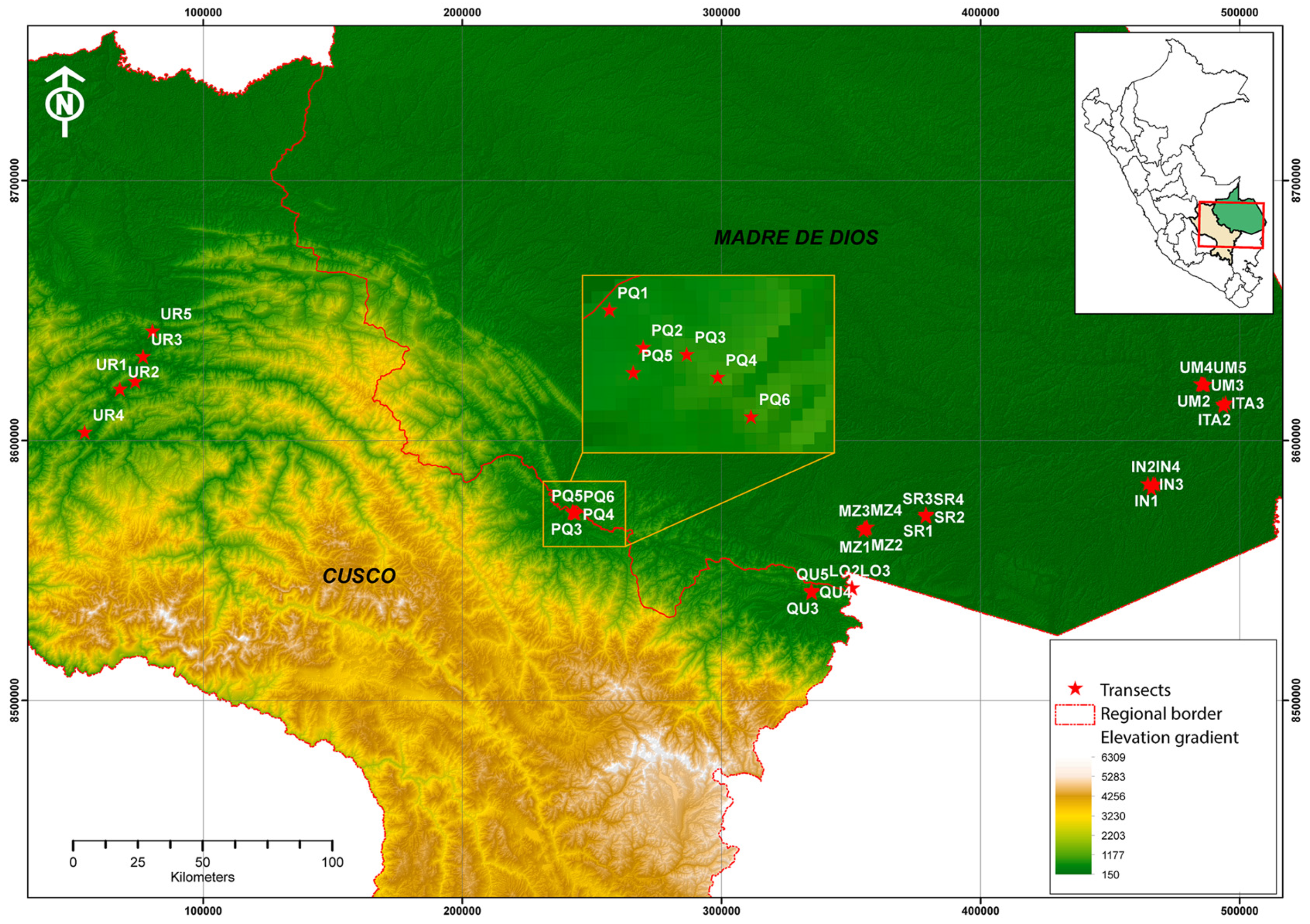
2.1. Study Area
2.2. Methodology
2.3. Data Analysis
3. Results
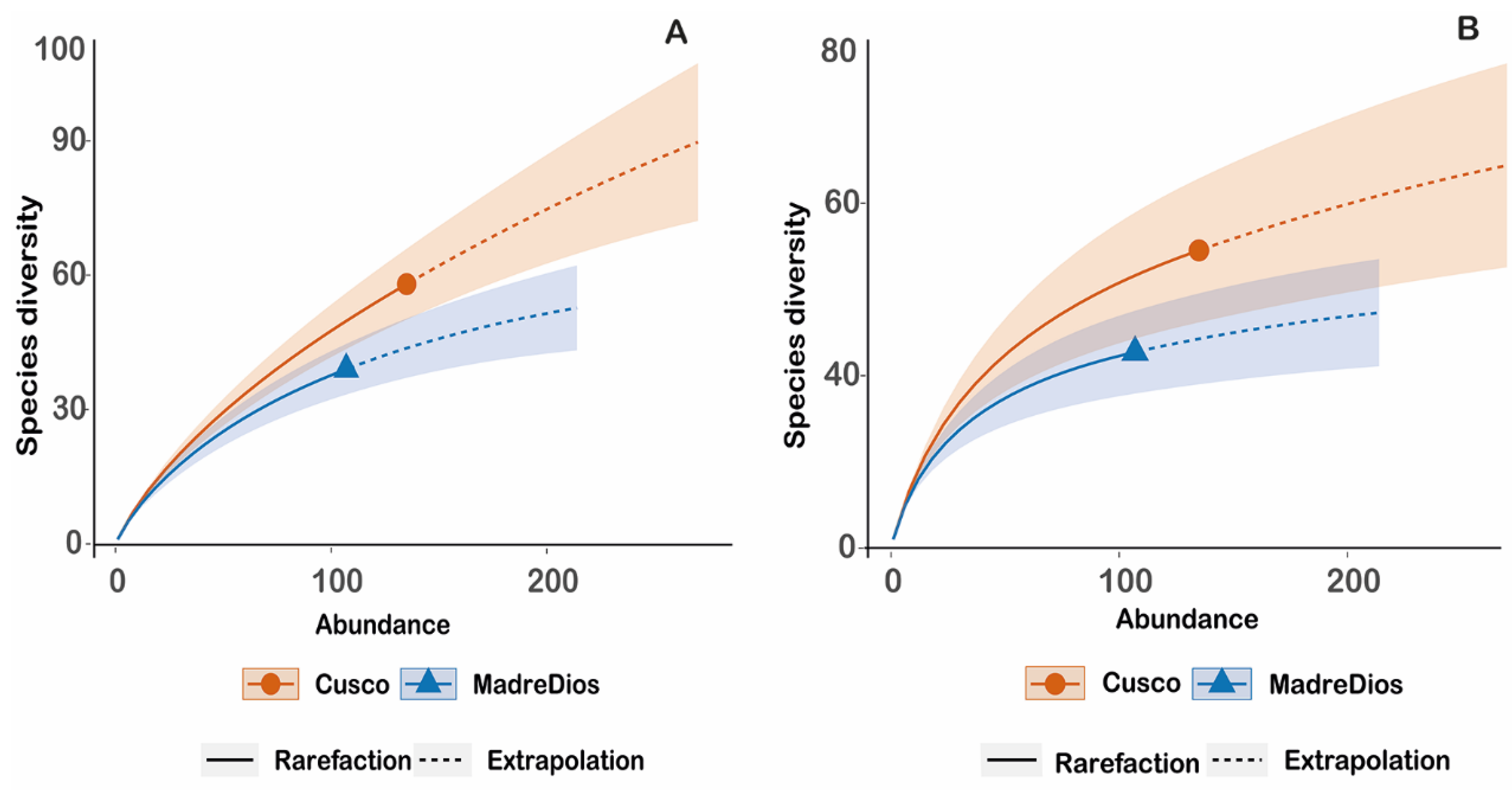
3.1. Diversity and Composition
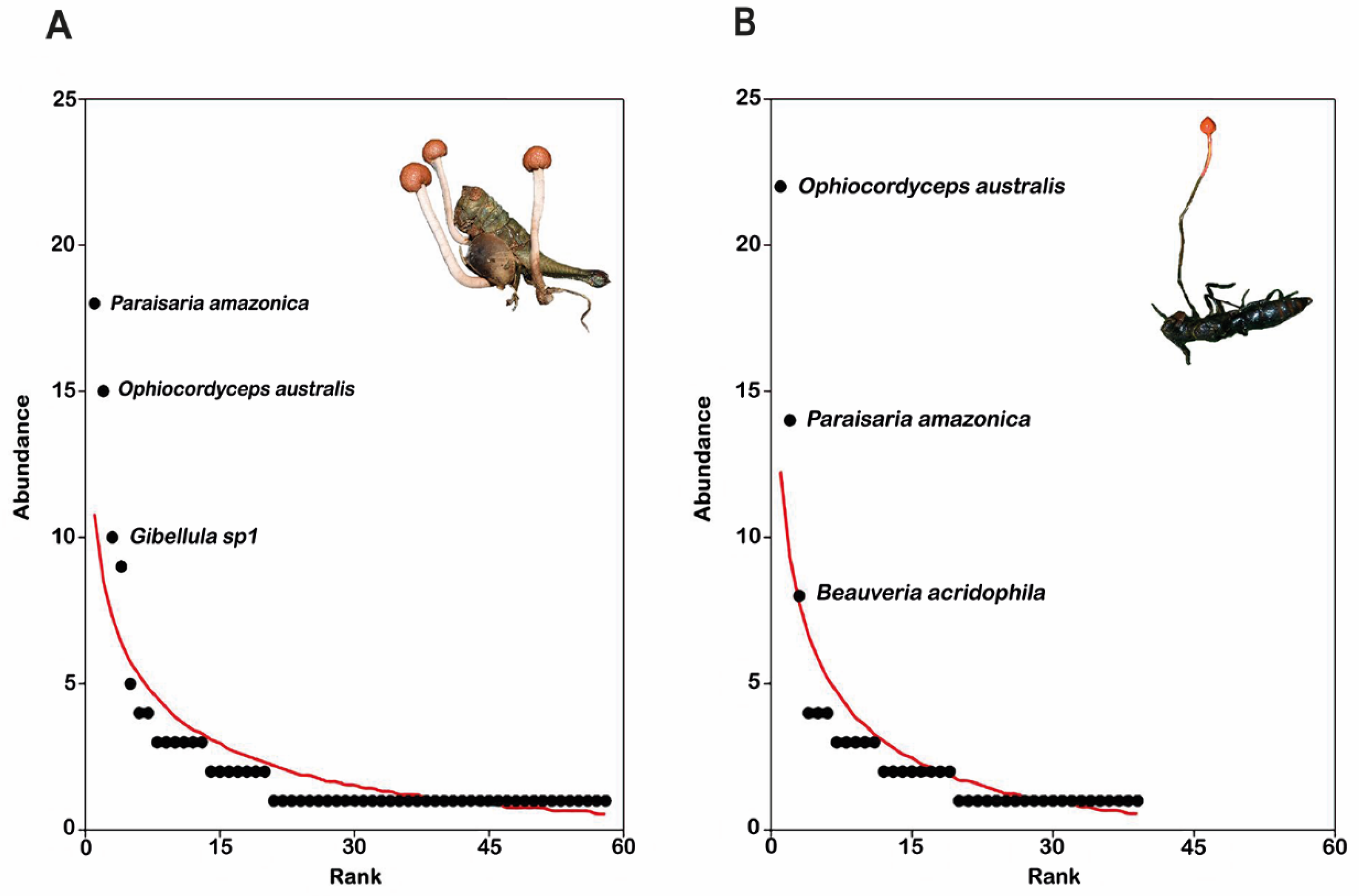
3.2. Structure
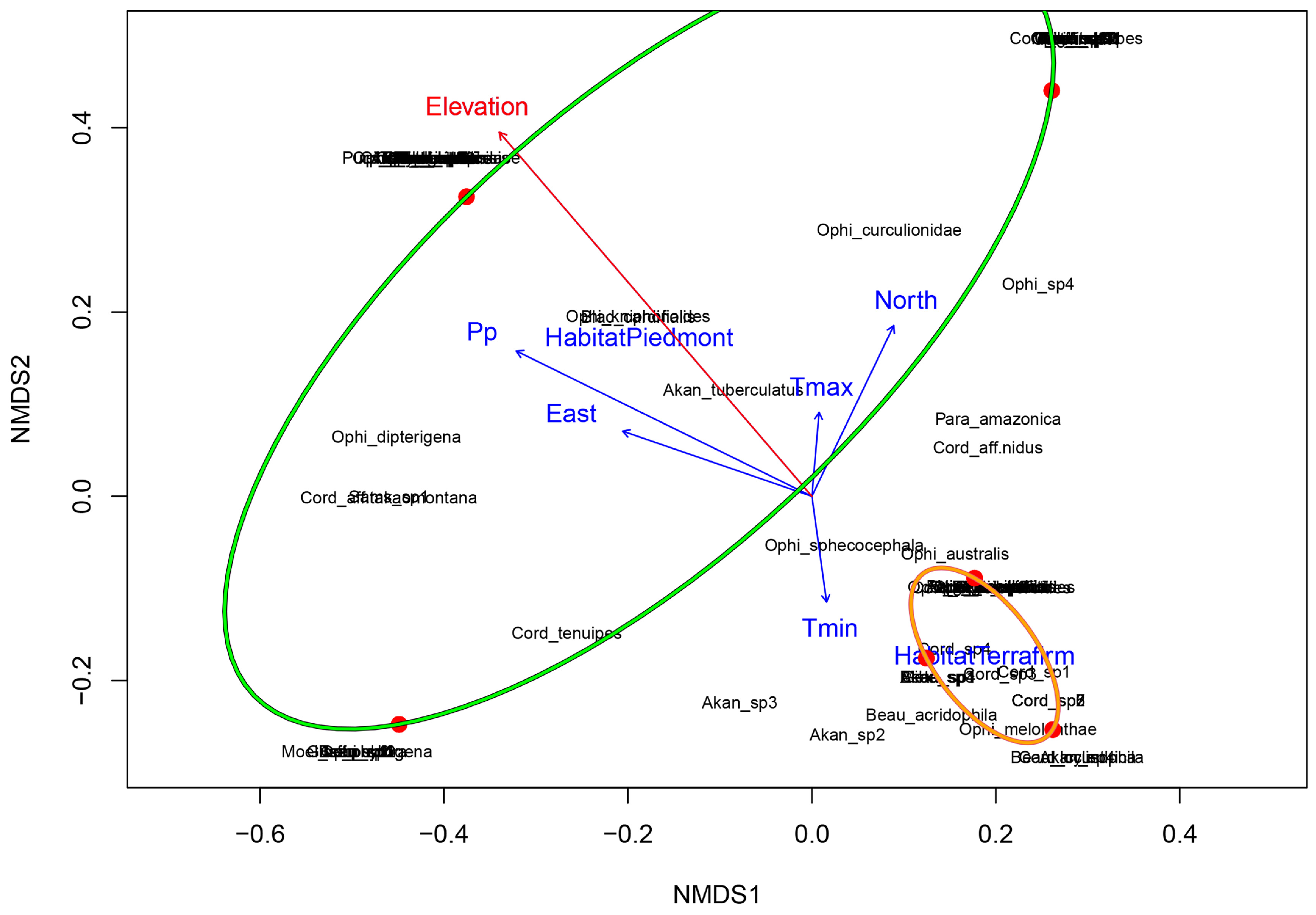
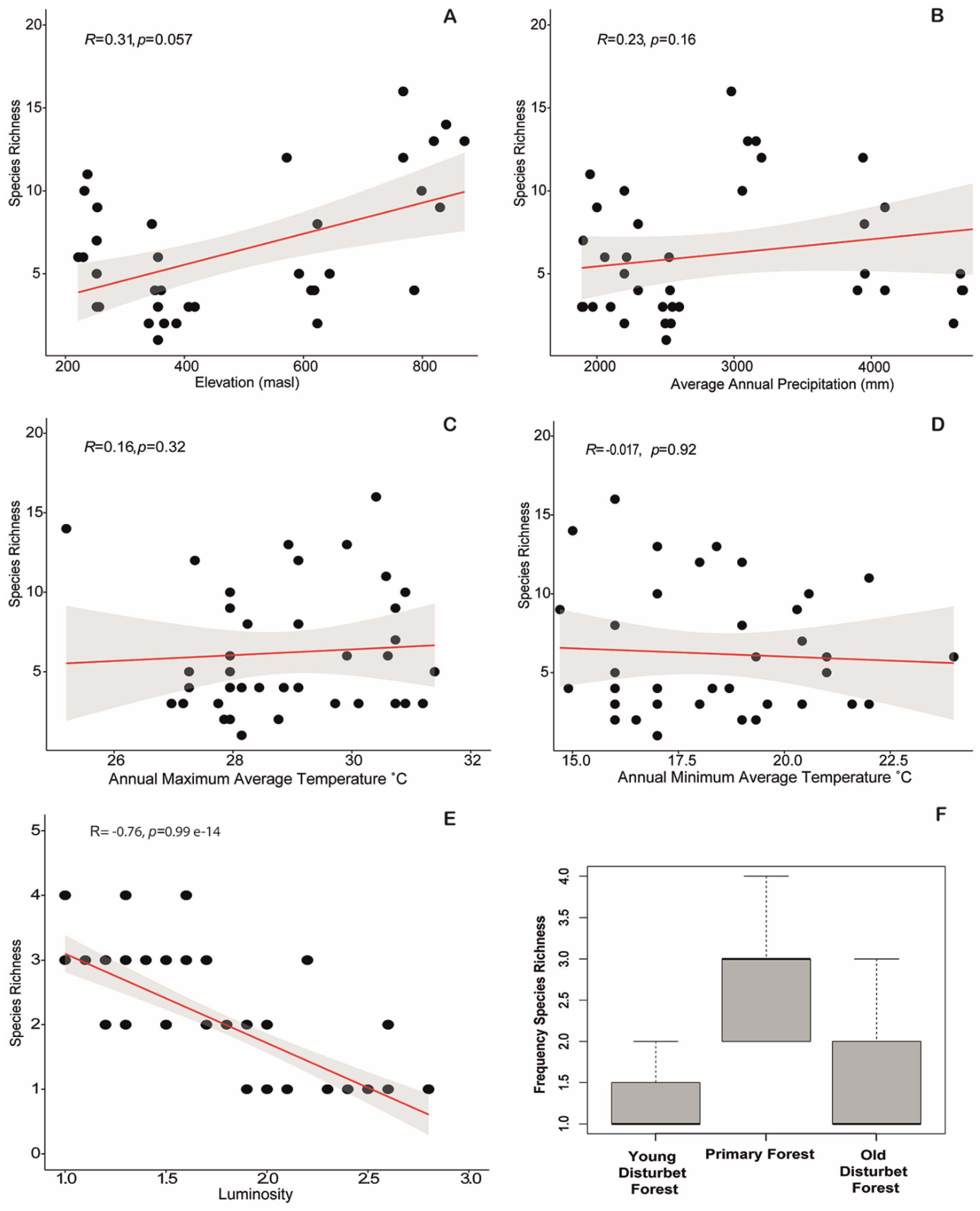
3.3. Environmental Relationships
4. Discussion
4.1. Patterns of Diversity and Composition
4.2. Dominance Patterns at the Regional Scale
4.3. Influence of Local Environmental Conditions on Arthropod-Pathogen Fungi
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nilsson, R.H.; Anslan, S.; Bahram, M.; Wurzbacher, C.; Baldrian, P. Mycobiome diversity: High-throughput sequencing and identification of fungi. Nat. Rev. Microbiol. 2019, 17, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Butt, T.M.; Coates, C.J.; Dubovskiy, I.M.; Ratcliffe, N.A. Entomopathogenic fungi: New insights into host–pathogen interactions. Adv. Genet. 2016, 94, 307–634. [Google Scholar]
- Blackwell, M. Fungal Evolution and Taxonomy. In BioControl; Springer Nature: Cham, Switzerland, 2010; Volume 55, pp. 7–16. [Google Scholar]
- Barrios-Gonzales, J.; Mejía, A. Production of secondary metabolites by solid-state fermentation. Biotechnol. Annu. Rev. 1996, 2, 85–121. [Google Scholar]
- Das, S.K.; Masuda, M.; Sakurai, A.; Sakakibara, M. Medicinal uses of the mushroom Cordyceps militaris: Current state and prospects. Fitoterapia 2010, 81, 961–968. [Google Scholar] [CrossRef]
- Catalogue of Life Homepage. 2023. Available online: https://www.catalogueoflife.org/Checklist (accessed on 9 September 2023).
- Sung, G.H.; Hywel-Jones, N.L.; Sung, J.M.; Luangsa-ard, J.J.; Shrestha, B.; Spatafora, J.W. Phylogenetic classification of Cordyceps and the Clavicipitaceous fungi. Stud. Mycol. 2007, 57, 5–59. [Google Scholar] [CrossRef]
- Aung, O.M.; Soytong, K.; Hyde, K.D. Diversity of entomopathogenic fungi in rainforest of Chiang Mai Province, Thailand. Fungal Divers. 2008, 30, 15–20. [Google Scholar]
- Sanjuan, T.; Henao, L.; Amat, G. Distribución espacial de Cordyceps spp. (Ascomycotina: Clavicipitaceae) y su impacto sobre las hormigas en selvas del piedemonte amazónico de Colombia. Rev. de Biol. Trop. 2001, 49, 945–955. [Google Scholar]
- Evans, C.H. Natural control of arthropods, with special reference to ants (Formicidae) by fungi in the tropical high forest of Ghana. J. Appl. Ecol. 1974, 11, 37–49. [Google Scholar] [CrossRef]
- Gazis, R. Evaluation of the Macrofungal Community at Los Amigos Biological Station, Madre de Dios, Peru. Master’s Thesis, Texas Christian University, Fort Worth, TX, USA, 2007. [Google Scholar]
- García, R.M. Contribución al Conocimiento de los Macrohongos en la Provincia de Tambopata, Madre de Dios. Ph.D. Thesis, Universidad Politécnica de Madrid, España, Madrid, Spain, 2015. [Google Scholar]
- Josse, C.; Navarro, G.; Encarnación, F.; Tovar, A.; Comer, P.; Ferreira, W.; Rodríguez, F.; Saito, J.; Sanjurjo, J.; Dyson, J.; et al. Sistemas Ecológicos de la Cuenca Amazónica de Perú y Bolivia: Clasificación y Mapeo; NatureServe: Arlington, VA, USA, 2007. [Google Scholar]
- Instituto Nacional de Desarrollo. Mesozonificación Ecológica—Económica del Corredor Interoceánico Sur, tramo Iñapari—Inambari; Madre de Dios, Perú; 2007. Available online: https://sigrid.cenepred.gob.pe/docs/PARA%20PUBLICAR/OTROS/GORE_MDD_EstudioMezoZZE_CorredorInteroceanico_SurTramoInapariInambiri_INADE_2007.pdf (accessed on 24 April 2005).
- Mains, E.B. North American entomogenous species of Cordyceps. Mycologia 2015, 20, 169–222. [Google Scholar] [CrossRef]
- Kobayasi, Y. Keys to the taxa of the genera Cordyceps and Torrubiella. Trans. Mycol. Soc. Jpn. 1982, 23, 329–364. [Google Scholar]
- Phillips, O.; Miller, P. Global Patterns of Plant Diversity: Alwyn H. Gentry’s Forest Transect Data Set; Monographs in Systematic Botany from the Missouri Botanical Garden: St. Louis, MO, USA, 2002. [Google Scholar]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: An R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- R Development Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; ISBN 3-900051-07-0. [Google Scholar]
- Chao, A. Nonparametric estimation of the number of classes in a population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- Chao, A. Estimating the population size for capture-recapture data with unequal catchability. Biometrics 1987, 43, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Lejendre, P.; Minchin, P.; Hara, R.; Solymos, P.; Stevens, H.; Szöcs, E.; et al. Vegan Community Ecology Package; R Package Version 2.6–2; The Comprehensive R Archive Network: Vienna, Austria, 2022; Available online: https://CRAN.R-project.org/package=vegan (accessed on 12 October 2022).
- Magurran, A.E. Ecological Diversity and Its Measurement; Princeton University Press: Princeton, NJ, USA, 1988. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–4. [Google Scholar]
- Pitman, N.; Terborgh, J.; Silman, M.; Nuñez, P.; Neill, D.; Cerón, C.; Palacios, W.; Aulestia, M. Dominance and distribution of tree species in upper Amazonian terra firme forests. Ecology 2001, 82, 2101–2117. [Google Scholar] [CrossRef]
- Sanjuan, T. Toward review of the genus Cordyceps sensu lato in the Neotropical region. In Proceedings of the XI International Mycological Congress, San Juan de, Puerto Rico, Puerto Rico, 16–21 July 2018. [Google Scholar]
- Serrano, A.Y.L. Diversidad de hongos entomopatógenos del género Cordyceps s.l. (Hypocreales: Clavicipitaceae) en el Ecuador. Bachelor’s Thesis, Pontificia Universidad Católica del Ecuador, Quito, Ecuador, 2016. [Google Scholar]
- Araujo, J.P.M.; Evans, C.H.; Kepler RHughes, D. Zombie-ant fungi across continents: 15 new species and new combinations within Ophiocordyceps. I. Myrmecophilous hirsutelloid species. Stud. Mycol. 2018, 90, 119–160. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Villamares, J.C.; Burrola-Aguilar, C.; Aguilar-Miguel, X.; Sanjuan, T.; Jiménez-Sánchez, E. Nuevos registros de hongos entomopatógenos del género Cordyceps s. l. (Ascomycota: Hypocreales) del Estado de México. Rev. Mex. de Biodivers. 2017, 88, 773–783. [Google Scholar] [CrossRef]
- León, B.; Roque, J.; Ulloa, C.; Pitman, N.; Jorgensen, P.; Cano, A. Libro Rojo de las Plantas endémicas del Perú. Rev. Peru. de Biol. Número Espec. 2006, 13, 1–971. [Google Scholar]
- Sanjuan, T.; Tabima, J.; Restrepo, S.; Laessoe, T.; Spatafora, J.W.; Franco-Molano, A.E. Entomopathogens of Amazonian stick insects and locusts are members of the Beauveria species complex (Cordyceps sensu stricto). Mycologia 2014, 106, 260–275. [Google Scholar] [CrossRef]
- Steege, H.; Pitman, N.; Sabatier, D.; Baraloto, C.; Salomão, R.; Guevara, J.; Phillips, O.; Castilho, C.; Magnusson, W.E.; Molino, J.-F.; et al. Hyperdominance in the Amazonian tree flora. Science 2013, 342, 1243092. [Google Scholar] [CrossRef]
- Sanjuan, T.I.; Franco-Molano, A.E.; Kepler, R.M.; Spatafora, J.W.; Tabima, J.; Vasco-Palacios, A.M.; Restrepo, S. Five new species of entomopathogenic fungi from the Amazon and evolution of neotropical Ophiocordyceps. Fungal Biol. 2015, 119, 901–916. [Google Scholar] [CrossRef] [PubMed]
- Velandia, L.M. Diversidad de Hongos Artrópodo-Patógenos en Bosques de Piedemonte y Galería, en Yopal-Casanare, Colombia. Bachelor’s Thesis, Universidad Internacional del Trópico Americano, Casanare, Colombia, 2022. [Google Scholar]
- Chaverri, P.; Liu, M.; Hodge, K.T. A monograph of the entomopathogenic genera Hypocrella, Moelleriella, and Samuelsia gen. nov. (Ascomycota, Hypocreales, Clavicipitaceae), and their aschersonia-like anamorphs in the Neotropics. Stud. Mycol. 2008, 60, 1–66. [Google Scholar] [CrossRef] [PubMed]
- Hudson, P.J.; Dobson, A.P.; Lafferty, K.D. Is a healthy ecosystem one that is rich in parasites? Trends Ecol. Evol. 2006, 21, 381–385. [Google Scholar] [CrossRef]
- Amédégnato, C.; Descamps, M. Dispersal centers of the Amazonian acridids. Acta Amaz. 1982, 12, 155–165. [Google Scholar] [CrossRef]
- Holliday, J.C.; Cleaver, M.P. Medicinal value of the caterpillar fungi species of the genus Cordyceps (Fr.) Link (Ascomycetes). A Review. Int. J. Med. Mushrooms 2008, 10, 219–234. [Google Scholar] [CrossRef]
- Meyling, N.V.; Eilenberg, J. Ecology of the entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae in temperate agroecosystems: Potential for conservation biological control. Biol. Control 2007, 43, 145–155. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Basin | Type of Amazonian Forest | UTM | East Coordinates | North Coordinates | Elevation (msl) | Annual Total Precipitation (mm) | Average Annual Minimum Temperature (°C) | Average Annual Maximal Temperature (°C) |
|---|---|---|---|---|---|---|---|---|---|
| Cusco | Araza | Piedmont | 19L | 334894.07 | 8542052.28 | 600.00 | 4670.00 | 16.00 | 28.83 |
| Madre de Dios | Inambari | Terra firme | 19L | 350424.55 | 8543401.24 | 400.00 | 2600.00 | 19.33 | 28.92 |
| Madre de Dios | Tambopata | Terra firme | 19L | 467402.00 | 8583322.00 | 263.00 | 2160.00 | 20.58 | 30.67 |
| Cusco | Urubamba | Piedmont | 18L | 706345.52 | 8606087.83 | 750.00 | 3200.00 | 21.33 | 31.25 |
| Cusco | Amarumayu | Piedmont | 19L | 242571.88 | 8572773.15 | 650.00 | 3950.00 | 15.00 | 29.17 |
| Madre de Dios | MadreDios | Terra firme | 19L | 486657.46 | 8621633.58 | 250.00 | 1900.00 | 20.42 | 30.83 |
| Basin | Type of Amazonian Forest | Number of Samples Evaluated | Species Richness | Fisher Alpha Diversity | Chao-1 Diversity |
|---|---|---|---|---|---|
| Amarumayu | Piedmont | 6 | 20 | 11.9 | 71.49 |
| Araza | Piedmont | 5 | 16 | 11.8 | 37.84 |
| Inambari | Terra firme | 11 | 15 | 7.162 | 20.15 |
| Madre de Dios | Terra firme | 8 | 28 | 13.29 | 50.51 |
| Tambopata | Terra firme | 4 | 14 | 5.58 | 21.38 |
| Urubamba | Piedmont | 5 | 31 | 17.76 | 35.53 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huamantupa-Chuquimaco, I.; Holgado Rojas, M.E.; Quispe Ordoñez, M.A.; García Roca, M.; Medina, A.C.; Ancco, W.Q.; Poccohuanca-Aguilar, R.O.; Córdova, Z.M.C.; Meza Calvo, J.G.; Sanjuan Giraldo, T.I. Patterns of Diversity, Structure and Local Ecology of Arthropod-Pathogenic Fungi in the Amazonian Forest of Cusco and Madre de Dios Regions, Southern Peru. Diversity 2023, 15, 1122. https://doi.org/10.3390/d15111122
Huamantupa-Chuquimaco I, Holgado Rojas ME, Quispe Ordoñez MA, García Roca M, Medina AC, Ancco WQ, Poccohuanca-Aguilar RO, Córdova ZMC, Meza Calvo JG, Sanjuan Giraldo TI. Patterns of Diversity, Structure and Local Ecology of Arthropod-Pathogenic Fungi in the Amazonian Forest of Cusco and Madre de Dios Regions, Southern Peru. Diversity. 2023; 15(11):1122. https://doi.org/10.3390/d15111122
Chicago/Turabian StyleHuamantupa-Chuquimaco, Isau, María Encarnación Holgado Rojas, Miguel Angel Quispe Ordoñez, Mishari García Roca, Anatoly Cárdenas Medina, Willians Quispe Ancco, Roger Oswaldo Poccohuanca-Aguilar, Zoila Magaly Cuba Córdova, Jackeline Greta Meza Calvo, and Tatiana Ibeth Sanjuan Giraldo. 2023. "Patterns of Diversity, Structure and Local Ecology of Arthropod-Pathogenic Fungi in the Amazonian Forest of Cusco and Madre de Dios Regions, Southern Peru" Diversity 15, no. 11: 1122. https://doi.org/10.3390/d15111122
APA StyleHuamantupa-Chuquimaco, I., Holgado Rojas, M. E., Quispe Ordoñez, M. A., García Roca, M., Medina, A. C., Ancco, W. Q., Poccohuanca-Aguilar, R. O., Córdova, Z. M. C., Meza Calvo, J. G., & Sanjuan Giraldo, T. I. (2023). Patterns of Diversity, Structure and Local Ecology of Arthropod-Pathogenic Fungi in the Amazonian Forest of Cusco and Madre de Dios Regions, Southern Peru. Diversity, 15(11), 1122. https://doi.org/10.3390/d15111122