


Bacterial Community Composition and Presence of Plasmids in the Endosphere- and Rhizosphere-Associated Microbiota of Sea Fig (Carpobrotus aequilaterus)

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Specimens
2.2. Sampling and DNA Extraction
2.3. Bacterial Community Analyses
2.3.1. Quantification of the Bacteria
2.3.2. DNA Metabarcoding Analysis
2.3.3. DNA Data Analyses
2.4. Occurrence of Plasmids
2.4.1. Detection of Plasmids
2.4.2. Quantification of IncP Plasmids
2.5. Statistical Analysis
3. Results
3.1. Bacterial Community
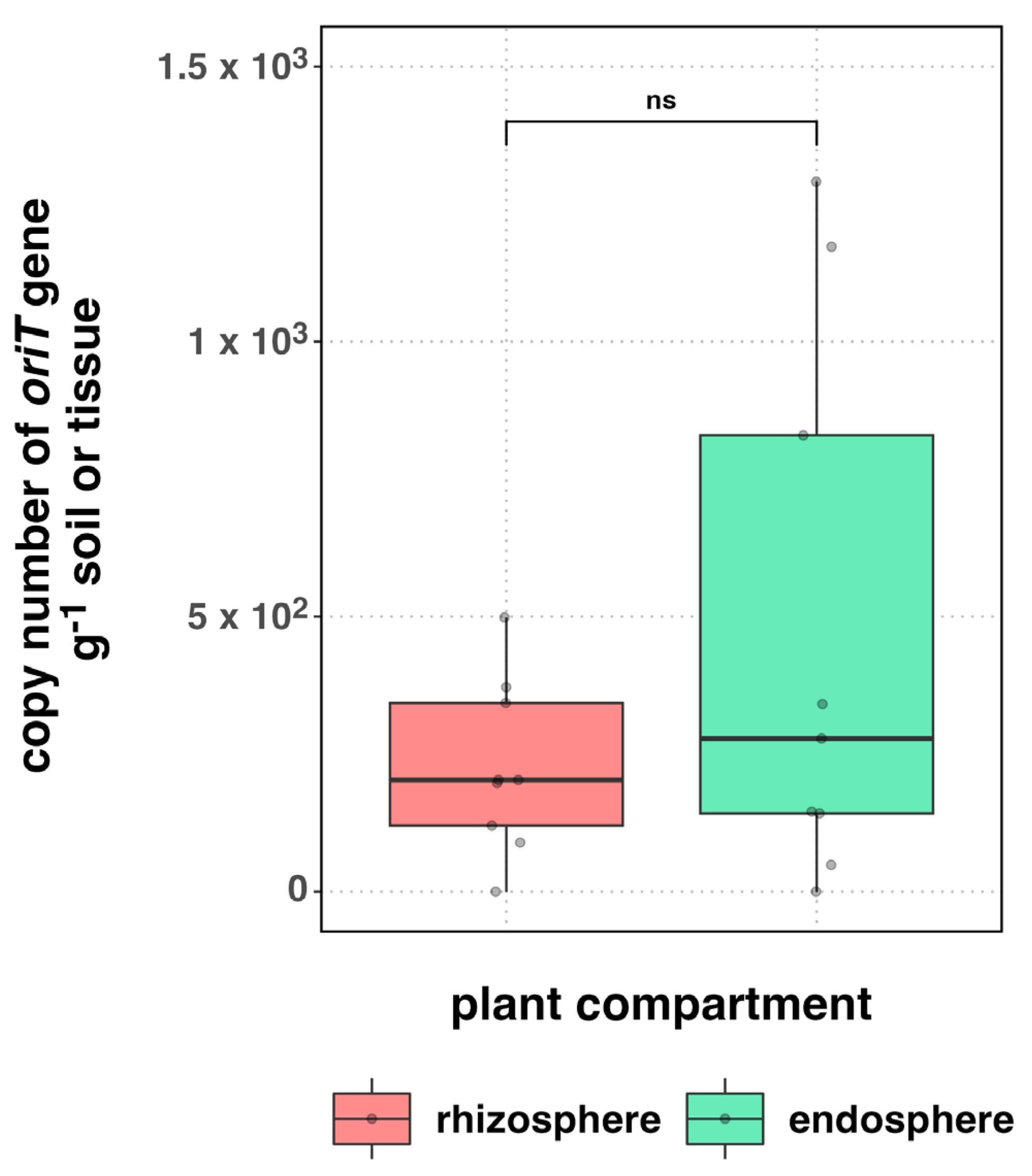
3.2. Occurrence of Plasmids
4. Discussion
4.1. Bacterial Communities
4.2. Occurrence of Plasmids
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Gwenzi, W.; Musiyiwa, K.; Mangori, L. Sources, behavior and health risks of antimicrobial resistance genes in wastewaters: A hotspot reservoir. J. Environ. Chem. Eng. 2020, 8, 102220. [Google Scholar] [CrossRef]
- Marcoleta, A.E.; Varas, M.A.; Costa, J.; Rojas-Salgado, J.; Arros, P.; Berríos-Pastén, C.; Tapia, S.; Silva, D.; Fierro, J.; Canales, N.; et al. Mapping the microbial diversity and natural resistome of North Antarctica soils. bioRxiv 2021. [Google Scholar] [CrossRef]
- Riva, V.; Patania, G.; Riva, F.; Vergani, L.; Crotti, E.; Mapelli, F. Acinetobacter baylyi Strain BD413 Can Acquire an Antibiotic Resistance Gene by Natural Transformation on Lettuce Phylloplane and Enter the Endosphere. Antibiotics 2022, 11, 1231. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Antimicrobial Resistance: Global Report on Surveillance; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Chen, C.Y.; Fuqua, C.; Jackson, C.R.; Kadlec, K.; Top, E.M. Plasmid transfer-mechanisms, ecology, evolution and applications. Front. Microbiol. 2022, 13, 993628. [Google Scholar]
- Shintani, M.; Sanchez, Z.K.; Kimbara, K. Genomics of microbial plasmids: Classification and identification based on replication and transfer systems and host taxonomy. Front. Microbiol. 2015, 6, 242. [Google Scholar] [CrossRef]
- Li, X.; Stokholm, J.; Brejnrod, A.; Vestergaard, G.A.; Russel, J.; Trivedi, U.; Thorsen, J.; Gupta, S.; Hjelmsø, M.H.; Shah, S.A.; et al. The infant gut resistome associates with E. coli, environmental exposures, gut microbiome maturity, and asthma-associated bacterial composition. Cell Host Microbe 2021, 29, 975–987. [Google Scholar] [CrossRef] [PubMed]
- Macedo, G.; Olesen, A.K.; Maccario, L.; Hernandez Leal, L.; Maas, P.v.d.; Heederik, D.; Mevius, D.; Sørensen, S.J.; Schmitt, H. Horizontal gene transfer of an IncP1 plasmid to soil bacterial community introduced by Escherichia coli through manure amendment in soil microcosms. Environ. Sci. Technol. 2022, 56, 11398–11408. [Google Scholar] [CrossRef] [PubMed]
- Olesen, A.K.; Pinilla-Redondo, R.; Hansen, M.F.; Russel, J.; Dechesne, A.; Smets, B.F.; Madsen, J.S.; Nesme, J.; Sørensen, S.J. IncHI1A plasmids potentially facilitate horizontal flow of antibiotic resistance genes to pathogens in microbial communities of urban residential sewage. Mol. Ecol. 2022, 31, 1595–1608. [Google Scholar] [CrossRef]
- Xu, H.; Chen, Z.; Huang, R.; Cui, Y.; Li, Q.; Zhao, Y.; Wang, X.; Mao, D.; Luo, Y.; Ren, H. Antibiotic resistance gene-carrying plasmid spreads into the plant endophytic bacteria using soil bacteria as carriers. Environ. Sci. Technol. 2021, 55, 10462–10470. [Google Scholar] [CrossRef]
- Pilosof, S. Conceptualizing microbe–plasmid communities as complex adaptive systems. Trends Microbiol. 2023, 31, 672–680. [Google Scholar] [CrossRef]
- Taghavi, S.; Barac, T.; Greenberg, B.; Borremans, B.; Vangronsveld, J.; van der Lelie, D. Horizontal gene transfer to endogenous endophytic bacteria from poplar improves phytoremediation of toluene. Appl. Environ. Microbiol. 2005, 71, 8500–8505. [Google Scholar] [CrossRef]
- Wang, Y.; Xiao, M.; Geng, X.; Liu, J.; Chen, J. Horizontal transfer of genetic determinants for degradation of phenol between the bacteria living in plant and its rhizosphere. Appl. Microbiol. Biotechnol. 2007, 77, 733–739. [Google Scholar] [CrossRef]
- Wang, Y.; Li, H.; Zhao, W.; He, X.; Chen, J.; Geng, X.; Xiao, M. Induction of toluene degradation and growth promotion in corn and wheat by horizontal gene transfer within endophytic bacteria. Soil Biol. Biochem. 2010, 42, 1051–1057. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.; Bakker, P.A. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Haney, C.H.; Samuel, B.S.; Bush, J.; Ausubel, F.M. Associations with rhizosphere bacteria can confer an adaptive advantage to plants. Nat. Plants 2015, 1, 15051. [Google Scholar] [CrossRef] [PubMed]
- Van Elsas, J.D.; Turner, S.; Bailey, M.J. Horizontal gene transfer in the phytosphere. New Phytol. 2003, 157, 525–537. [Google Scholar] [CrossRef]
- Hassani, M.; Durán, P.; Hacquard, S. Microbial interactions within the plant holobiont. Microbiome 2018, 6, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Mølbak, L.; Kroer, N. Root growth and exudate production define the frequency of horizontal plasmid transfer in the rhizosphere. FEMS Microbiol. Ecol. 2007, 59, 167–176. [Google Scholar] [CrossRef]
- Zhu, H.; Zhang, L.; Li, S.; Wang, Y.; Sun, S.; Chen, J.; Kou, S.; Jin, Q.; Xiao, M. The rhizosphere and root exudates of maize seedlings drive plasmid mobilization in soil. Appl. Soil Ecol. 2018, 124, 194–202. [Google Scholar] [CrossRef]
- Carrión, V.J.; Perez-Jaramillo, J.; Cordovez, V.; Tracanna, V.; de Hollander, M.; Ruiz-Buck, D.; Mendes, L.W.; van Ijcken, W.F.J.; Gomez-Exposito, R.; Elsayed, S.S.; et al. Pathogen-induced activation of disease-suppressive functions in the endophytic root microbiome. Science 2019, 366, 606–612. [Google Scholar] [CrossRef]
- Sánchez-Salazar, A.M.; Taparia, T.; Olesen, A.K.; Acuña, J.J.; Sørensen, S.J.; Jorquera, M.A. An overview of plasmid transfer in the plant microbiome. Plasmid 2023, 127, 102695. [Google Scholar] [CrossRef] [PubMed]
- Earnshaw, M.J.; Carver, K.A.; Charlton, W.A. Leaf anatomy, water relations and crassulacean acid metabolism in the chlorenchyma and colourless internal water-storage tissue of Carpobrotus edulis and Senecio? mandraliscae. Planta 1987, 170, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Novoa, A.; Keet, J.H.; Lechuga-Lago, Y.; Pyšek, P.; Roux, J.J.L. Urbanization and Carpobrotus edulis invasion alter the diversity and composition of soil bacterial communities in coastal areas. FEMS Microbiol. Ecol. 2020, 96, fiaa106. [Google Scholar] [CrossRef]
- Campoy, J.G.; Acosta, A.T.R.; Affre, L.; Barreiro, R.; Brundu, G.; Buisson, E.; González, L.; Lema, M.; Novoa, A.; Retuerto, R.; et al. Monographs of invasive plants in Europe: Carpobrotus. Bot. Lett. 2018, 165, 440–475. [Google Scholar] [CrossRef]
- GISD. Global Invasive Species Database, ISSG, Species Profile: Carpobrotus edulis. 2018. Available online: http://www.iucngisd.org/gisd/ (accessed on 23 July 2023).
- Tapia, Y.; Bustos, P.; Salazar, O.; Casanova, M.; Castillo, B.; Acuña, E.; Masaguer, A. Phytostabilization of Cu in mine tailings using native plant Carpobrotus aequilaterus and the addition of potassium humates. J. Geochem. Explor. 2017, 183, 102–113. [Google Scholar] [CrossRef]
- Govindasamy, V.; George, P.; Ramesh, S.V.; Sureshkumar, P.; Rane, J.; Minhas, P.S. Characterization of root-endophytic actinobacteria from cactus (Opuntia ficus-indica) for plant growth promoting traits. Arch. Microbiol. 2022, 204, 150. [Google Scholar] [CrossRef] [PubMed]
- Lagos, L.M.; Navarrete, O.U.; Maruyama, F.; Crowley, D.E.; Cid, F.P.; Mora, M.L.; Jorquera, M.A. Bacterial community structures in rhizosphere microsites of ryegrass (Lolium perenne var. Nui) as revealed by pyrosequencing. Biol. Fertil. Soils 2014, 50, 1253–1266. [Google Scholar] [CrossRef]
- Jorquera, M.A.; Martínez, O.A.; Marileo, L.G.; Acuna, J.J.; Saggar, S.; Mora, M.L. Effect of nitrogen and phosphorus fertilization on the composition of rhizobacterial communities of two Chilean Andisol pastures. World J. Microbiol. Biotechnol. 2014, 30, 99–107. [Google Scholar] [CrossRef]
- Whelan, J.A.; Russell, N.B.; Whelan, M.A. A method for the absolute quantification of cDNA using real-time PCR. J. Immunol. Methods 2003, 278, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Acuña, J.J.; Lagos, L.M.; Ogram, A.; Jorquera, M.A. Bacterial alkaline phosphatase in the rhizosphere of plants grown in Chilean extreme environments. Biol. Fertil. Soils 2016, 52, 763–773. [Google Scholar] [CrossRef]
- Herlemann, D.P.; Labrenz, M.; Jürgens, K.; Bertilsson, S.; Waniek, J.J.; Andersson, A.F. Transitions in bacterial communities along the 2000 km salinity gradient of the Baltic Sea. ISME J. 2011, 5, 1571–1579. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Peter, R.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; et al. Vegan: Community Ecology Package (Version 2.5-6); The Comprehensive R Archive Network: Vienna, Austria, 2016. [Google Scholar]
- Louca, S.; Parfrey, L.W.; Doebeli, M. Decoupling function and taxonomy in the global ocean microbiome. Science 2016, 353, 1272–1277. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Acuña, J.J.; Inostroza, N.G.; Duran, P.; Mora, M.L.; Sadowsky, M.J.; Jorquera, M.A. Niche differentiation in the composition, predicted function, and co-occurrence networks in bacterial communities associated with Antarctic vascular plants. Front. Microbiol. 2020, 11, 1036. [Google Scholar] [CrossRef]
- Zhang, T.; Zhang, X.X.; Ye, L. Plasmid metagenome reveals high levels of antibiotic resistance genes and mobile genetic elements in activated sludge. PLoS ONE 2011, 6, e26041. [Google Scholar] [CrossRef] [PubMed]
- Götz, A.; Pukall, R.; Smit, E.; Tietze, E.; Prager, R.; Tschäpe, H.; van Elsas, J.D.; Smalla, K. Detection and characterization of broad-host-range plasmids in environmental bacteria by PCR. Appl. Environ. Microbiol. 1996, 62, 2621–2628. [Google Scholar] [CrossRef]
- Carattoli, A.; Bertini, A.; Villa, L.; Falbo, V.; Hopkins, K.L.; Threlfall, E.J. Identification of plasmids by PCR-based replicon typing. J. Microbiol. Methods 2005, 63, 219–228. [Google Scholar] [CrossRef]
- Jorquera, M.; Yamaguchi, N.; Tani, K.; Nasu, M. A combination of direct viable counting, fluorescence in situ hybridization, and green fluorescent protein gene expression for estimating plasmid transfer at the single cell level. Microbes Environ. 2006, 21, 101–111. [Google Scholar] [CrossRef]
- Zhang, Q.; Acuña, J.J.; Inostroza, N.G.; Mora, M.L.; Radic, S.; Sadowsky, M.J.; Jorquera, M.A. Endophytic bacterial communities associated with roots and leaves of plants growing in Chilean extreme environments. Sci. Rep. 2019, 9, 4950. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Li, Y.; Tang, L.; Ikenaga, M.; Liu, R.; Xu, G. Soil microbial community shifts explain habitat heterogeneity in two Haloxylon species from a nutrient perspective. Ecol. Evol. 2023, 13, e9727. [Google Scholar] [CrossRef] [PubMed]
- Xing, R.; Gao, Q.B.; Zhang, F.Q.; Wang, J.L.; Chen, S.L. Large-scale distribution of bacterial communities in the Qaidam Basin of the Qinghai-Tibet Plateau. MicrobiologyOpen 2019, 8, e909. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, W.; Li, Q.; Cui, R.; Wang, Z.; Wang, Y.; Zhang, Y.-Z.; Ding, W.; Shen, X. Deciphering the root endosphere microbiome of the desert plant Alhagi sparsifolia for drought resistance-promoting bacteria. Appl. Environ. Microbiol. 2020, 86, e02863-19. [Google Scholar] [CrossRef] [PubMed]
- Mandakovic, D.; Aguado-Norese, C.; García-Jiménez, B.; Hodar, C.; Maldonado, J.E.; Gaete, A.; Latorre, M.; Wilkinson, M.D.; Gutiérrez, R.; Cavieres, L.A.; et al. Testing the stress gradient hypothesis in soil bacterial communities associated with vegetation belts in the Andean Atacama Desert. Environ. Microbiome 2023, 18, 1–17. [Google Scholar] [CrossRef]
- Fonseca-García, C.; Desgarennes, D.; Flores-Núñez, V.M.; Partida-Martínez, L.P. The Microbiome of Desert CAM Plants: Lessons from Amplicon Sequencing and Metagenomics; Elsevier Inc.: Amsterdam, The Netherlands, 2018. [Google Scholar] [CrossRef]
- Griffiths, H.; Males, J. Succulent plants. Curr. Biol. 2017, 27, R890–R896. [Google Scholar] [CrossRef]
- Gao, J.; Luo, Y.; Wei, Y.; Huang, Y.; Zhang, H.; He, W.; Sheng, H.; An, L. Effect of aridity and dune type on rhizosphere soil bacterial communities of Caragana microphylla in desert regions of northern China. PLoS ONE 2019, 14, e0224195. [Google Scholar] [CrossRef]
- Chen, J.; Xu, D.; Chao, L.; Liu, H.; Bao, Y. Microbial assemblages associated with the rhizosphere and endosphere of an herbage, Leymus chinensis. Microb. Biotechnol. 2020, 13, 1390–1402. [Google Scholar] [CrossRef]
- Dong, C.J.; Wang, L.L.; Li, Q.; Shang, Q.M. Bacterial communities in the rhizosphere, phyllosphere and endosphere of tomato plants. PLoS ONE 2019, 14, e0223847. [Google Scholar] [CrossRef]
- Zhang, Q.; White, J.F. Bioprospecting desert plants for endophytic and biostimulant microbes: A strategy for enhancing agricultural production in a Hotter, drier future. Biology 2021, 10, 961. [Google Scholar] [CrossRef] [PubMed]
- Karray, F.; Gargouri, M.; Chebaane, A.; Mhiri, N.; Mliki, A.; Sayadi, S. Climatic aridity gradient modulates the diversity of the rhizosphere and endosphere bacterial microbiomes of Opuntia ficus-indica. Front. Microbiol. 2020, 11, 1622. [Google Scholar] [CrossRef] [PubMed]
- Astorga-Eló, M.; Zhang, Q.; Larama, G.; Stoll, A.; Sadowsky, M.J.; Jorquera, M.A. Composition, predicted functions and co-occurrence networks of rhizobacterial communities impacting flowering desert events in the Atacama Desert, Chile. Front. Microbiol. 2020, 11, 571. [Google Scholar] [CrossRef]
- Zhang, W.; Bahadur, A.; Zhang, G.; Zhang, B.; Wu, X.; Chen, T.; Liu, G. Diverse Bacterial Communities from Qaidam Basin of the Qinghai–Tibet Plateau: Insights into Variations in Bacterial Diversity Across Different Regions. Front. Microbiol. 2020, 11, 554105. [Google Scholar] [CrossRef]
- Caravaca, F.; Torres, P.; Díaz, G.; Roldán, A. Elevated functional versatility of the soil microbial community associated with the invader Carpobrotus edulis across a broad geographical scale. Sci. Total Environ. 2022, 813, 152627. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Caballero, G.; Caravaca, F.; Díaz, G.; Torres, P.; Roldán, A. The invader Carpobrotus edulis promotes a specific rhizosphere microbiome across globally distributed coastal ecosystems. Sci. Total Environ. 2020, 719, 137347. [Google Scholar] [CrossRef] [PubMed]
- Menicagli, V.; Balestri, E.; Giommoni, F.; Vannini, C.; Lardicci, C. Plastic litter changes the rhizosphere bacterial community of coastal dune plants. Sci. Total Environ. 2023, 880, 163293. [Google Scholar] [CrossRef]
- Liu, J.; Sun, X.; Zuo, Y.; Hu, Q.; He, X. Plant species shape the bacterial communities on the phyllosphere in a hyper-arid desert. Microbiol. Res. 2023, 269, 127314. [Google Scholar] [CrossRef]
- Zhang, M.; Li, X.; Shi, C.; Qiu, Z.; Han, J.; Wang, K.; Zheng, P.; Shi, F. Driving factors, co-occurrence networks, and metabolic profiles of soil bacterial communities within the root proximity of Amaranthus palmeri. J. Soil Sci. Plant Nutr. 2022, 22, 1928–1941. [Google Scholar] [CrossRef]
- Vymazal, J. Removal of nutrients in various types of constructed wetlands. Sci. Total Environ. 2007, 380, 48–65. [Google Scholar] [CrossRef]
- Balkwill, D.L.; Fredrickson, J.K.; Romine, M.F. Sphingomonas and related genera. In The Prokaryotes: Volume 7: Proteobacteria: Delta, Epsilon Subclass; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 605–629. [Google Scholar]
- Stolz, A. Molecular characteristics of xenobiotic-degrading sphingomonads. Appl. Microbiol. Biotechnol. 2009, 81, 793–811. [Google Scholar] [CrossRef]
- Smit, E.; Wolters, A.; van Elsas, J.D. Self-transmissible mercury resistance plasmids with gene-mobilizing capacity in soil bacterial populations: Influence of wheat roots and mercury addition. Appl. Environ. Microbiol. 1998, 64, 1210–1219. [Google Scholar] [CrossRef] [PubMed]
- Kinkle, B.K.; Schmidt, E.L. Transfer of the pea symbiotic plasmid pJB5JI in nonsterile soil. Appl. Environ. Microbiol. 1991, 57, 3264–3269. [Google Scholar] [CrossRef]
- Mølbak, L.; Licht, T.R.; Kvist, T.; Kroer, N.; Andersen, S.R. Plasmid transfer from Pseudomonas putida to the indigenous bacteria on alfalfa sprouts: Characterization, direct quantification, and in situ location of transconjugant cells. Appl. Environ. Microbiol. 2003, 69, 5536–5542. [Google Scholar] [CrossRef]
- Musovic, S.; Oregaard, G.; Kroer, N.; Sørensen, S.J. Cultivation-independent examination of horizontal transfer and host range of an IncP-1 plasmid among gram-positive and gram-negative bacteria indigenous to the barley rhizosphere. Appl. Environ. Microbiol. 2006, 72, 6687–6692. [Google Scholar] [CrossRef] [PubMed]
- Jorquera, M.A.; Hernández, M.; Martínez, O.; Marschner, P.; de la Luz Mora, M. Detection of aluminum tolerance plasmids and microbial diversity in the rhizosphere of plants grown in acidic volcanic soil. Eur. J. Soil Biol. 2010, 46, 255–263. [Google Scholar] [CrossRef]
- Assinder, S.J.; Williams, P.A. The TOL Plasmids: Determinants of the Catabolism of Toluene and the Xylenes; Advances in Microbial Physiology; Academic Press: Cambridge, MA, USA, 1990; Volume 31, pp. 1–69. [Google Scholar]
- Dealtry, S.; Holmsgaard, P.N.; Dunon, V.; Jechalke, S.; Ding, G.-C.; Krögerrecklenfort, E.; Heuer, H.; Hansen, L.H.; Springael, D.; Zühlke, S.; et al. Shifts in abundance and diversity of mobile genetic elements after the introduction of diverse pesticides into an on-farm biopurification system over the course of a year. Appl. Environ. Microbiol. 2014, 80, 4012–4020. [Google Scholar] [CrossRef] [PubMed]
- Dunon, V.; Sniegowski, K.; Bers, K.; Lavigne, R.; Smalla, K.; Springael, D. High prevalence of IncP-1 plasmids and IS 1071 insertion sequences in on-farm biopurification systems and other pesticide-polluted environments. FEMS Microbiol. Ecol. 2013, 86, 415–431. [Google Scholar] [CrossRef]
- Marcoleta, A.E.; Arros, P.; Varas, M.A.; Costa, J.; Rojas-Salgado, J.; Berríos-Pastén, C.; Tapia-Fuentes, S.; Silva, D.; Fierro, J.; Canales, N.; et al. The highly diverse Antarctic Peninsula soil microbiota as a source of novel resistance genes. Sci. Total Environ. 2022, 810, 152003. [Google Scholar] [CrossRef]
- Compant, S.; Clément, C.; Sessitsch, A. Plant growth-promoting bacteria in the rhizo-and endosphere of plants: Their role, colonization, mechanisms involved and prospects for utilization. Soil Biol. Biochem. 2010, 42, 669–678. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inc Group | Region | Function | Primer Sequence (5′-3′) | Annealing Temperature (°C) |
|---|---|---|---|---|
| IncN | rep 1 | Replication | AGT TCA CCA CCT ACT CGC TCC G | 55 |
| rep 2 | Replication | CAA GTT CTT CTG TTG GGA TTC CG | ||
| oriT1 | Transfer | TTG GGC TTC ATA GTA CCC | 49 | |
| oriT2 | Transfer | GTG TGA TAG CGT GAT TTA TGC | ||
| IncP | oriT1 | Transfer | CAG CCT CGC AGA GCA GGA T | 57 |
| oriT2 | Transfer | CAG CCG GGC AGG ATA GGT GAA GT | ||
| trfA1 1 | Replication | ATG ACG ACC AAG AAG CG | 57 | |
| trfA1 2 | Replication | AAC CCC CAG CCG GAA CTG | ||
| traG 1 | Transfer | CTG CGT CAC GAT GAA CAG GCT TAC C | 63 | |
| traG 2 | Transfer | ACT TCC AGC GGC GTC TAT GTG G | ||
| IncQ | repB 1 | Replication | TCG TGG TCG CGT TCA AGG TAC G | 62 |
| repB2 | Replication | CTG TAA GTC GAT GAT CTG GGC GTT | ||
| IncW | oriV 1 | Replication | GAC CCG GAA AAC CAA AAA TA | 58 |
| oriV2 | Replication | GTG AGG GTG AGG GTG CTA TC | ||
| oriT 1 | Transfer | TCT GCA TCA TTG TAG CAC C | 51 | |
| oriT2 | Transfer | CCG TAG TGT TAC TGT AGT GG |
| Replication | Transference | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| IncN | IncP | IncQ | IncW | IncN | IncP | IncW | |||
| Sample | rep | trfA1 | repB | oriV | oriT | oriT | traG | oriT | |
| Endosphere | E1.1 | – | + | – | – | + | + | (+) | – |
| E1.2 | – | + | – | – | + | + | (+) | – | |
| E1.3 | – | + | – | – | + | + | (+) | – | |
| E2.1 | – | + | – | – | + | + | (+) | – | |
| E2.2 | – | + | – | – | + | + | (+) | – | |
| E2.3 | – | + | – | – | + | + | (+) | – | |
| E3.1 | – | + | – | – | + | + | (+) | – | |
| E3.2 | – | + | – | – | + | + | (+) | – | |
| E3.3 | – | + | – | – | + | + | (+) | – | |
| Rhizosphere | R1.1 | + | (+) | (+) | + | + | + | (+) | (+) |
| R1.2 | + | (+) | (+) | + | + | + | (+) | (+) | |
| R1.3 | + | (+) | (+) | + | + | + | (+) | (+) | |
| R2.1 | + | (+) | (+) | + | + | + | (+) | (+) | |
| R2.2 | + | (+) | (+) | + | + | + | (+) | (+) | |
| R2.3 | + | (+) | (+) | + | + | + | (+) | (+) | |
| R3.1 | + | (+) | (+) | + | + | + | (+) | (+) | |
| R3.2 | + | (+) | (+) | + | + | + | (+) | (+) | |
| R3.3 | + | (+) | (+) | + | + | + | (+) | (+) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Salazar, A.M.; Acuña, J.J.; Sadowsky, M.J.; Jorquera, M.A. Bacterial Community Composition and Presence of Plasmids in the Endosphere- and Rhizosphere-Associated Microbiota of Sea Fig (Carpobrotus aequilaterus). Diversity 2023, 15, 1156. https://doi.org/10.3390/d15111156
Sánchez-Salazar AM, Acuña JJ, Sadowsky MJ, Jorquera MA. Bacterial Community Composition and Presence of Plasmids in the Endosphere- and Rhizosphere-Associated Microbiota of Sea Fig (Carpobrotus aequilaterus). Diversity. 2023; 15(11):1156. https://doi.org/10.3390/d15111156
Chicago/Turabian StyleSánchez-Salazar, Angela M., Jacquelinne J. Acuña, Michael J. Sadowsky, and Milko A. Jorquera. 2023. "Bacterial Community Composition and Presence of Plasmids in the Endosphere- and Rhizosphere-Associated Microbiota of Sea Fig (Carpobrotus aequilaterus)" Diversity 15, no. 11: 1156. https://doi.org/10.3390/d15111156
APA StyleSánchez-Salazar, A. M., Acuña, J. J., Sadowsky, M. J., & Jorquera, M. A. (2023). Bacterial Community Composition and Presence of Plasmids in the Endosphere- and Rhizosphere-Associated Microbiota of Sea Fig (Carpobrotus aequilaterus). Diversity, 15(11), 1156. https://doi.org/10.3390/d15111156