
Ecosystem Resilience of a South African Mesic Grassland with Change from Rotational to Continuous Grazing


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Land Use History
2.3. Data Collection
2.3.1. Study Setup
2.3.2. Soil Physical Properties
2.3.3. Soil Chemical Properties
2.3.4. Forage Quality
2.3.5. Veld Condition Assessment
2.3.6. Plant Species Composition, Diversity, and Functional Diversity
2.4. Statistical Analyses
3. Results
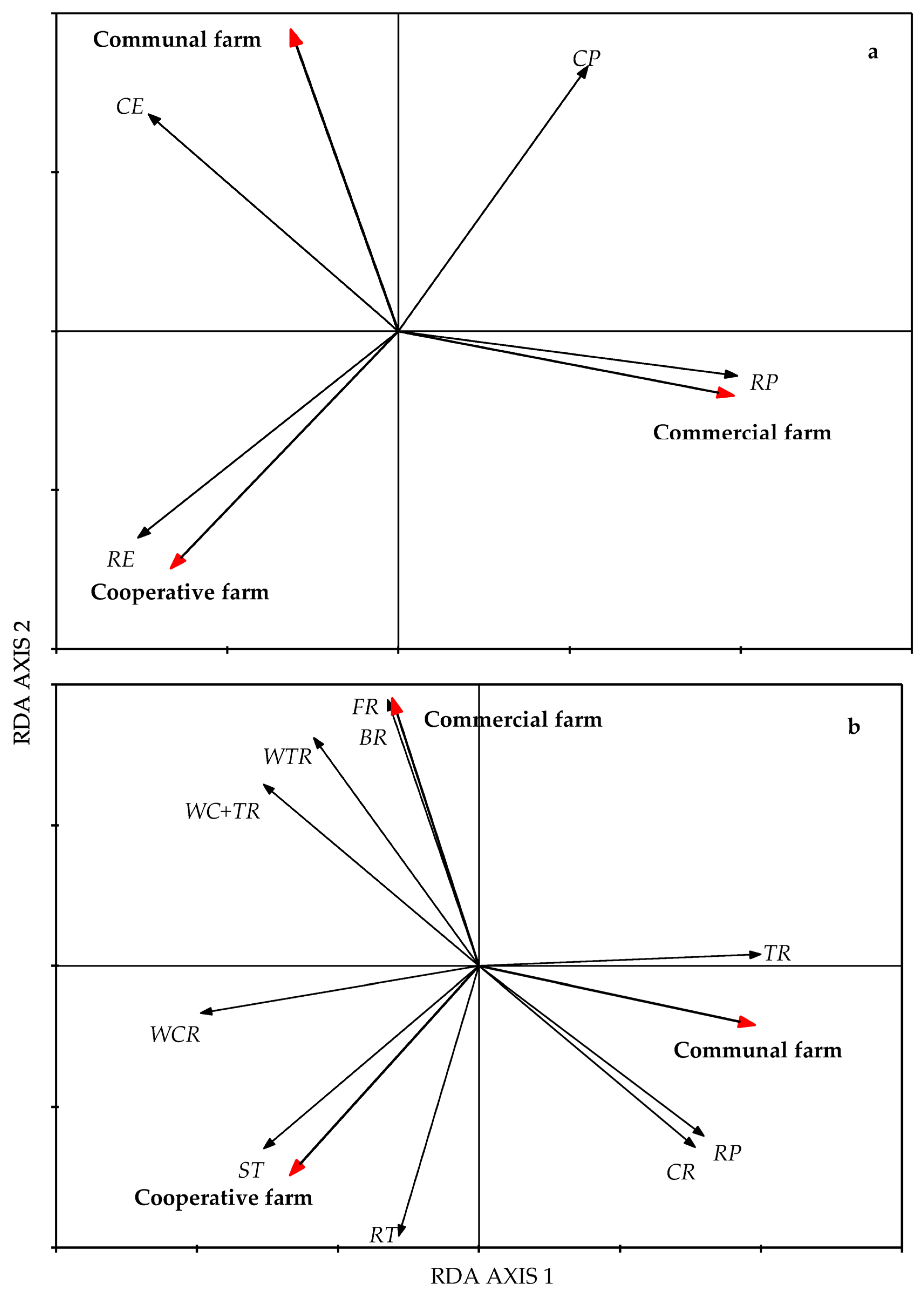
3.1. The Effect of Livestock Grazing on Soil Physical and Chemical Properties, Forage Quality, and Veld Condition
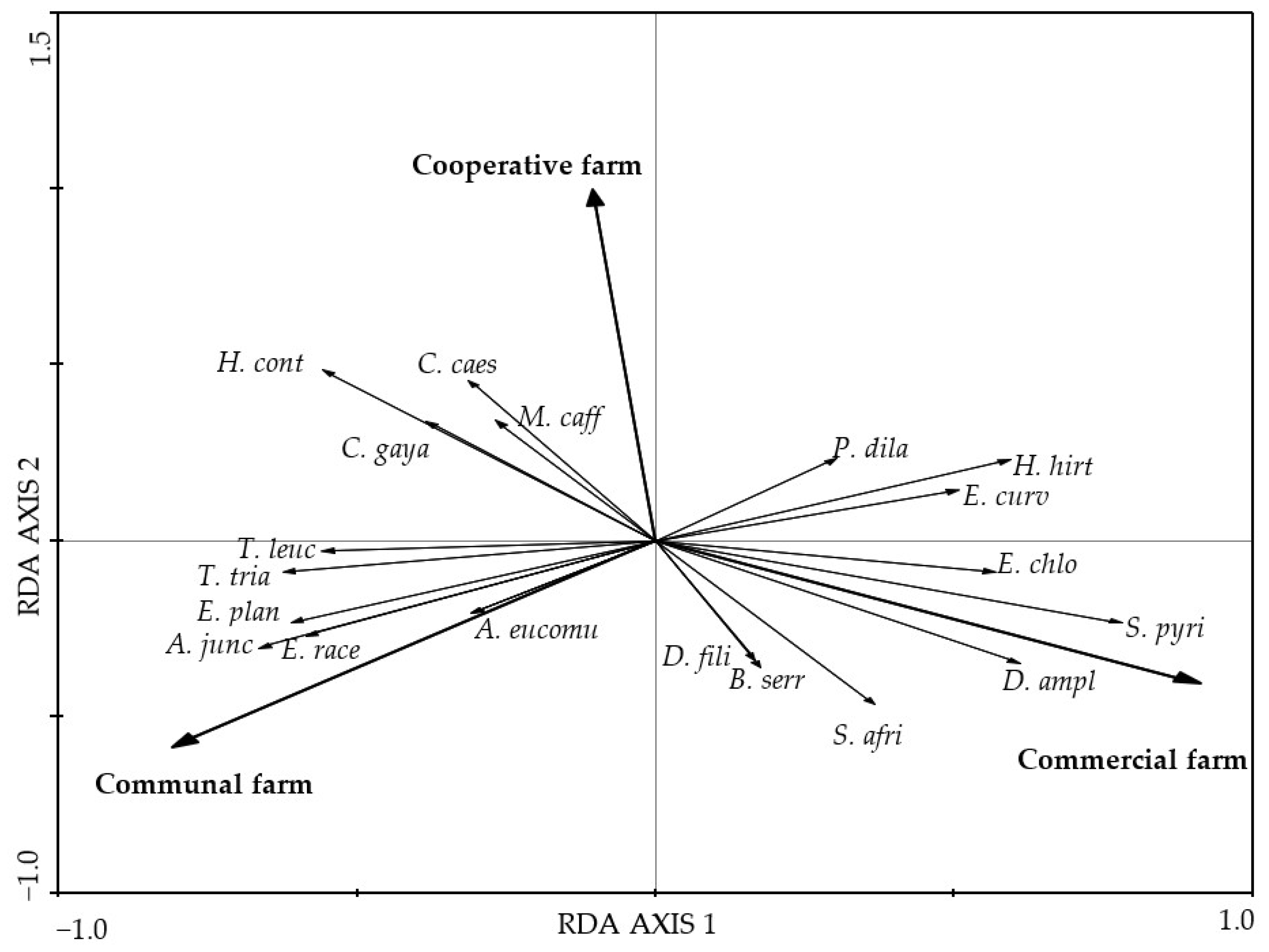
3.2. The Effect of Grazing on Plant Species Composition, Diversity, and Functional Diversity
3.3. Growth Forms of Forbs above Ground and Below Ground
4. Discussion
4.1. Effects of Grazing Systems on Soil Physical and Chemical Properties
4.2. Effects of Livestock Grazing on Forage Quality
4.3. Effects of Livestock Grazing on Plant Species Diversity and Functional Diversity
4.4. Functional Diversity of Forbs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cadman, M.; de Villiers, C.; Lechmere-Oertel, R.; McCulloch, D.; South African National Biodiversity Institute. Grasslands Ecosystem Guidelines: Landscape Interpretation for Planners & Managers; South African National Biodiversity Institute: Pretoria, South Africa, 2013. [Google Scholar]
- Blair, J.; Nippert, J.; Briggs, J. Grassland Ecology. In Ecology and the Environment; Monson, R.K., Ed.; The Plant Sciences: New York, NY, USA, 2014; pp. 389–423. [Google Scholar]
- Scott-Shaw, R.; Morris, C.D. Grazing depletes forb species diversity in the mesic grasslands of KwaZulu-Natal, South Africa. Afri. J. Range. For. Sci. 2015, 32, 21–31. [Google Scholar] [CrossRef]
- Fynn, R.W.; O’Connor, T.G. Determinants of community organization of a South African mesic grassland. J. Veg. Sci. 2005, 16, 93–102. [Google Scholar] [CrossRef]
- Bachinger, L.M.; Brown, L.R.; van Rooyen, M.W. The effects of firebreaks on plant diversity and species composition in the grasslands of the Loskop Dam Nature Reserve, South Africa. Afr. J. Ran. For. Sci. 2016, 33, 21–32. [Google Scholar] [CrossRef]
- Watkinson, A.R.; Ormerod, S.J. Grasslands, grazing and biodiversity: Editors introduction. J. Appl. Ecol. 2001, 38, 233–237. [Google Scholar]
- Martindale, G.J. Influence of Livestock Grazing on Plant Diversity of Highland Sourveld Grassland in KwaZulu-Natal. Ph.D. Thesis, University of Witwatersrand, Johannesburg, South Africa, 2007. [Google Scholar]
- Loucougaray, G.; Bonis, E.; Bouzille, B. Effects of grazing by horses and/or cattle on the diversity of coastal grasslands in western France. Biol. Cons. 2004, 11, 59–71. [Google Scholar] [CrossRef]
- Pavlu, V.; Hejcman, M.; Pavlu, L.; Gaisler, J. Effect of rotational and continuous grazing on vegetation of an upland grassland in the Jizerské Hory Mts., Czech Republic. Folia Geobot. 2003, 38, 21–34. [Google Scholar] [CrossRef]
- Hickman, K.R.; Hartnett, D.C.; Cochran, R.C.; Owensby, C.E. Grazing management effects on plant species diversity in tallgrass prairie. J. Range Manag. 2004, 57, 58–65. [Google Scholar] [CrossRef]
- O’Connor, T.G.; Kuyler, P.; Kirkman, K.P.; Corcoran, B. Which grazing management practices are most appropriate for maintaining biodiversity in South African grassland? Afri. J. Range For. Sci. 2010, 27, 67–76. [Google Scholar] [CrossRef]
- Perotti, E.; Probo, M.; Pittarello, M.; Lonati, M.; Lombardi, G. A 5-year rotational grazing changes the botanical composition of sub-alpine and alpine grasslands. Appl. Veg. Sci. 2018, 21, 647–657. [Google Scholar] [CrossRef]
- Folke, C.; Carpenter, S.; Walker, B.; Scheffer, M.; Elmqvist, T.; Gunderson, L.; Holling, C.S. Regime shifts, resilience, and biodiversity in ecosystem management. Ann. Rev. Ecol. Evol. Syste. 2004, 35, 555–581. [Google Scholar] [CrossRef]
- Côté, I.M.; Darling, E.S. Rethinking ecosystem resilience in the face of climate change. PLoS Biol. 2010, 8, e1000438. [Google Scholar] [CrossRef] [PubMed]
- Du, W.C.; Yan, T.; Chang, S.H.; Wang, Z.F.; Hou, F.J. Seasonal hogget grazing as a potential alternative grazing system for the Qinghai-Tibetan Plateau: Weight gain and animal behaviour under continuous or rotational grazing at high or low stocking rates. Rangel. J. 2017, 39, 329–339. [Google Scholar] [CrossRef]
- Boavista, L.D.R.; Trindade, J.P.P.; Overbeck, G.E.; Müller, S.C. Effects of grazing regimes on the temporal dynamics of grassland communities. Appl. Veg. Sci. 2019, 22, 326–335. [Google Scholar] [CrossRef]
- Karami, P.; Bandak, I.; Karaji, M.G. Comparing the effects of continuous grazing and long term exclosure on floristic composition and plant diversity in rangeland ecosystems of Saral, Iran. Int. J. Environ. Sci. Tech. 2019, 16, 7769–7776. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, J.; Niu, S. Toward a sustainable grazing management based on biodiversity and ecosystem multifunctionality in drylands. Curr. Opin. Environ. Sustain. 2021, 48, 36–43. [Google Scholar] [CrossRef]
- Ndandani, A. Range Condition Assessment to Document the Extent of Degradation on Selected Semi-Arid Rangelands of the Eastern Cape, South Africa. Master’s Thesis, University of Fort Hare, Alice, South Africa, 2016. [Google Scholar]
- Short, A. Can we use the ordered-distance method to estimate basal cover in tufted grasslands? Grassroots 2010, 10, 15–18. [Google Scholar]
- Kotzé, E.; Sandhage-Hofmann, A.; Meinel, J.A.; Du Preez, C.C.; Amelung, W. Rangeland management impacts on the properties of clay soils along grazing gradients in the semi-arid grassland biome of South Africa. J. Arid Envi. 2013, 97, 220–229. [Google Scholar] [CrossRef]
- Shezi, T.A.; O’Connor, T.G.; Witkowski, E.T.T. Impact of livestock grazing intensity on plant diversity of montane grassland in the northern Drakensberg, South Africa. Afri. J. Range For. Sci. 2021, 38, 67–79. [Google Scholar] [CrossRef]
- Ma, S.; Zhou, Y.; Gowda, P.H.; Chen, L.; Starks, P.J.; Steiner, J.L.; Neel, J.P.S. Evaluating the impacts of continuous and rotational grazing on tallgrass prairie landscape using high-spatial-resolution imagery. Agronomy 2019, 9, 238. [Google Scholar] [CrossRef]
- McDonald, S.E.; Reid, N.; Smith, R.; Waters, C.M.; Hunter, J.; Rader, R. Rotational grazing management achieves similar plant diversity outcomes to areas managed for conservation in a semi-arid rangeland. Rangel. J. 2019, 41, 135–145. [Google Scholar] [CrossRef]
- Venter, Z.S.; Cramer, M.D.; Hawkins, H.J. Rotational grazing management has little effect on remotely sensed vegetation characteristics across farm fence-line contrasts. Agric. Ecosyt. Environ. 2019, 282, 40–48. [Google Scholar] [CrossRef]
- Derner, J.D.; Hart, R.H. Grazing-induced modifications to peak standing crop in northern mixed-grass prairie. Rangel. Ecol. Manag. 2007, 60, 270–276. [Google Scholar] [CrossRef]
- Briske, D.D.; Derner, J.D.; Brown, J.R.; Fuhlendorf, S.D.; Teague, W.R.; Havstad, K.M.; Gillen, R.L.; Ash, A.J.; Willms, W.D. Rotational grazing on rangelands: Reconciliation of perception and experimental evidence. Rangel. Ecol. Manag. 2008, 61, 3–17. [Google Scholar] [CrossRef]
- Dent, M.C.; Lynch, S.; Schulze, R.E. (Eds.) Mapping Mean Annual and Other Rainfall Statistics Over Southern Africa; Water Research Commission Report No. 109/1/89; University of Natal: Durban, South Africa, 1987. [Google Scholar]
- Mucina, L.; Rutherford, M.C. (Eds.) The Vegetation of South Africa, Lesotho and Swaziland; South African National Biodiversity Institute: Pretoria, South Africa, 2006. [Google Scholar]
- Trollope, W.S.W.; Trollope, L.A.; Bosch, O.J.H. Veld and pasture management terminology in southern Africa. J. Grassl. Socie. South. Africa 1990, 7, 52–61. [Google Scholar] [CrossRef]
- Ramage, B.S.; Sheil, D.; Salim, H.M.; Fletcher, C.; Mustafa, N.Z.A.; Luruthusamay, J.C.; Harrison, R.D.; Butod, E.; Dzulkiply, A.D.; Kassim, A.R.; et al. Pseudoreplication in tropical forests and the resulting effects on biodiversity conservation. Conserv. Biol. 2013, 27, 364–372. [Google Scholar] [CrossRef]
- Davies, G.M.; Gray, A. Don’t let spurious accusations of pseudoreplication limit our ability to learn from natural experiments (and other messy kinds of ecological monitoring). Ecol. Evol. 2015, 5, 5295–5304. [Google Scholar] [CrossRef]
- Hulbert, S.H. Pseudoreplication and the design of ecological field experiments. Ecol. Monogr. 1984, 54, 187–211. [Google Scholar] [CrossRef]
- Rutherford, M.C.; Powrie, L.W. Severely degraded rangeland: Implications for plant diversity from a case study in Succulent Karoo, South Africa. J. Arid. Environ. 2010, 74, 692–701. [Google Scholar] [CrossRef]
- Herrick, J.E.; Jones, T.L. A dynamic cone penetrometer for measuring soil penetration resistance. Soil Sci. Soc. Am. J. 2011, 66, 1320–1324. [Google Scholar] [CrossRef]
- Laycock, W.A.; Conrad, P.W. Effect of grazing on soil compaction as measured by bulk density on a high elevation cattle range. Rangel. Ecol. Manag. 1967, 20, 136–140. [Google Scholar] [CrossRef]
- Manson, A.D.; Bainbridge, S.H.; Thibaud, G.R. Methods Used for Analysis of Soils and Plant Material by Analytical Services at Cedara; KZN Agri-Report No. N/A/2020/07; KwaZulu-Natal Department of Agriculture and Rural Development: Cedara, South Africa, 2020.
- Kirkman, K.P. Factors Affecting the Seasonal Variation of Veld Quality in South Africa. Master’s Thesis, University of Natal, Pietermaritzburg, South Africa, 1988. [Google Scholar]
- Van Oudtshoorn, F. Guide to the Grasses of Southern Africa; Briza Publications: Pretoria, South Africa, 1999. [Google Scholar]
- Danckwerts, J.E.; O’Reagain, P.J.; O’Connor, T.G. Range management in a changing environment: A southern African perspective. Rangel. J. 1993, 15, 133–144. [Google Scholar] [CrossRef]
- Rowntree, K.; Duma, M.; Kakembo, V.; Thornes, J. Debunking the myth of overgrazing and soil erosion. Land Degr. Dev. 2004 15, 203–214. [CrossRef]
- Lesoli, M.S. Vegetation and Soil Status and Human Perceptions on the Condition of Communal Rangelands of the Eastern Cape, South Africa. Master’s Thesis, University of Fort Hare, Alice, South Africa, 2008. [Google Scholar]
- Oztas, T.; Koc, A.; Comakli, B. Changes in vegetation and soil properties along a slope on overgrazed and eroded rangelands. J. Arid Environ. 2004, 55, 93–100. [Google Scholar] [CrossRef]
- Hardy, M.B.; Tainton, N.M. Towards a technique for determining basal cover in tufted grasslands. Afr. J. Range Forage Sci. 1993, 10, 77–81. [Google Scholar] [CrossRef]
- Chamane, S.; Kirkman, K.P.; Morris, C.; O’Connor, T. Does high-density stocking affect perennial forbs in mesic grassland? Afr. J. Range Forage Sci. 2017, 34, 133–142. [Google Scholar] [CrossRef]
- Angert, A.L. Demography of central and marginal populations of monkey flowers (Mimulus cardinalis and M. lewisii). Ecology 2006, 87, 2014–2025. [Google Scholar] [CrossRef] [PubMed]
- Díaz, S.; Acosta, A.; Cabido, M. Community structure in montane grasslands of central Argentina in relation to land use. J. Veg. Sci. 1994, 5, 483–488. [Google Scholar] [CrossRef]
- Uys, R.G. Patterns of Plant Diversity and Their Management Across South African Rangelands. Ph.D. Thesis, University of Cape Town, Cape Town, South Africa, 2006. [Google Scholar]
- Jaccard, P. The distribution of the flora in the alpine zone. New Phyto. 1912, 2, 37–50. [Google Scholar] [CrossRef]
- Ter Braak, C.J.; Smilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5). 2002. Available online: www.canoco.com (accessed on 23 September 2021).
- Greenwood, K.L.; McKenzie, B.M. Grazing effects on soil physical properties and the consequences for pastures: A review. Aus. J. Exp. Agric. 2001, 4, 1231–1250. [Google Scholar] [CrossRef]
- Xie, Y.; Wittig, R. The impact of grazing intensity on soil characteristics of Stipa grandis and Stipa bungeana steppe in northern China (autonomous region of Ningxia). Acta Oecol. 2004, 25, 197–204. [Google Scholar] [CrossRef]
- Tufa, M.; Melese, A.; Tena, W. Effects of land use types on selected soil physical and chemical properties: The case of Kuyu District, Ethiopia. Eurasian J. Soil Sci. 2019, 8, 94–109. [Google Scholar] [CrossRef]
- Tesk, C.R.M.; Pedreira, B.C.; Pereira, D.H.; Pina, D.D.S.; Ramos, T.A.; Mombach, M.A. Impact of grazing management on forage qualitative characteristics: A review. Sci. Electron. Arch. 2018, 11, 188–197. [Google Scholar] [CrossRef]
- Di Virgilio, A.; Lambertucci, S.A.; Morales, J.M. Sustainable grazing management in rangelands: Over a century searching for a silver bullet. Agric. Ecosyst. Environ. 2019, 283, 106–561. [Google Scholar] [CrossRef]
- Tracy, B.F.; Bauer, R.B. Evaluating mob stocking for beef cattle in a temperate grassland. PLoS ONE 2019, 14, e0226360. [Google Scholar] [CrossRef]
- Moojen, F.G.; Bremm, C.; Laca, E.A.; Machado, D.R.; Savian, J.V.; Carvalho, P.C. Herbage allowance and stocking method effect on grazing systems: Results of a long-term experiment. Grass Forage Sci. 2001, 77, 45–54. [Google Scholar] [CrossRef]
- Collins, M.; Fritz, J.O. Forage quality. An introduction to grassland agriculture. In Forages, Volume 1: An Introduction to Grassland Agriculture, 7th ed.; Barnes, R.F., Nelson, C.J., Collins, M., Moore, K.J., Eds.; Wiley Blackwell: Hoboken, NJ, USA, 2003; pp. 363–390. [Google Scholar]
- Fenetahun, Y.; You, Y.; Fentahun, T.; Xinwen, X.; Yong-Dong, W. Effects of grazing intensity on forage nutritive value of dominant grass species in Borana rangelands of Southern Ethiopia. PeerJ. 2001, 9, e12204. [Google Scholar] [CrossRef] [PubMed]
- Kioko, J.; Kiringe, J.W.; Seno, S.O. Impacts of livestock grazing on a savanna grassland in Kenya. J. Arid Land 2012, 4, 29–35. [Google Scholar] [CrossRef]
- Modernel, P.; Picasso, V.; Carmo, M.; Rossing, W.A.; Corbeels, M.; Soca, P.; Dogliotti, S.; Tittonell, P. Grazing management for more resilient mixed livestock farming systems on native grasslands of southern South America. Grass Forage Sci. 2019, 74, 636–649. [Google Scholar] [CrossRef]
- Hanke, W.; Böhner, J.; Dreber, N.; Jürgens, N.; Schmiedel, U.; Wesuls, D.; Dengler, J. The impact of livestock grazing on plant diversity: An analysis across dryland ecosystems and scales in southern Africa. Ecol. Appl. 2014, 24, 1188–1203. [Google Scholar] [CrossRef]
- Craven, D.; Eisenhauer, N.; Pearse, W.D.; Hautier, Y.; Isbell, F.; Roscher, C.; Manning, P. Multiple facets of biodiversity drive the diversity–stability relationship. Nat. Ecol. Evol. 2018, 2, 1579–1587. [Google Scholar] [CrossRef]
- Muller, M.; Siebert, S.J.; Ntloko, B.R.; Siebert, F. A floristic assessment of grassland diversity loss in South Africa. Both. Afr. Biodiv. Conser. 2021, 51, 1–9. [Google Scholar] [CrossRef]
- Ratovonamana, R.Y.; Rajeriarison, C.; Roger, E.; Kiefer, I.; Ganzhorn, J.U. Impact of livestock grazing on forest structure, plant species composition and biomass in southwestern Madagascar. Scr. Bot. Belg. 2013, 50, 82–92. [Google Scholar]
- Briske, D.D.; Noy-Meir, I. Plant responses to grazing: A comparative evaluation of annual and perennial grasses. In Ecological Basis of Livestock Grazing in Mediterranean Ecosystems; Publications Office of the European Union: Luxembourg, 1998; Volume 13, pp. 26–39. [Google Scholar]
- Fidelis, A.; Appezzato-da-Glória, B.; Pillar, V.D.; Pfadenhauer, J. Does disturbance affect bud bank size and belowground structures diversity in Brazilian subtropical grasslands? Flora-Morphol. Distrib. Funct. Ecol. Plants 2014, 209, 110–116. [Google Scholar] [CrossRef]
- Lamont, B.B.; Downes, K.S. Fire-stimulated flowering among resprouters and geophytes in Australia and South Africa. Plant Ecol. 2011, 212, 2111–2125. [Google Scholar] [CrossRef]
- Ganjurjav, H.; Zhang, Y.; Gornish, E.S.; Hu, G.; Li, Y.; Wan, Y.; Gao, Q. Differential resistance and resilience of functional groups to livestock grazing maintain ecosystem stability in an alpine steppe on the Qinghai-Tibetan Plateau. J. Environ. Manag. 2019, 251, 109–579. [Google Scholar] [CrossRef]
- Diaz, S.; Lavorel, S.; McIntyre, S.; Falczuk, V.; Casanoves, F.; Milchunas, D.G.; Skarpe, C.; Rusch, G.; Sternberg, M.; Noy-Meir, I. Plant trait responses to grazing–a global synthesis. Glob. Change Biol. 2007, 13, 313–341. [Google Scholar] [CrossRef]
- Morris, C.D.; Scott-Shaw, R. Potential grazing indicator forbs for two mesic grasslands in South Africa. Ecol. Indic. 2019, 107, 105–611. [Google Scholar] [CrossRef]
- Fish, L.; Mashau, A.C.; Moeaha, M.J.; Nembudani, M.T. Identification Guide to Southern African Grasses. An Identification Manual with Keys, Descriptions and Distributions. Strelitzia 36. South African Biodiversity Institute, Pretoria. World Flora Online. 2015. Available online: http://www.worldfloraonline.org/ (accessed on 19 November 2023).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CMF | COP | CF | |
|---|---|---|---|
| Total size of grazing area (ha) | 430 | 350 | 412 |
| Mean herd size (AU) * | 160 | 105 | 250 |
| Number of paddocks | 6 | 1 | None |
| Mean paddock size (ha) | 70 | 350 | None |
| Mean period of occupation (days) | 150 | Throughout the year | Throughout the year |
| Mean period of absence (days) | 90 | - | - |
| Soil Chemical Properties | CMF | COP | CF | F | p |
|---|---|---|---|---|---|
| P (mg/kg) | 7.79 ± 1.81 | 9.26 ± 0.85 | 9.87 ± 0.42 | 0.814 | 0.466 |
| K (mg/kg) | 222.01 ± 31.66 | 264.51 | 237.97 ± 26.07 | 0.401 | 0.678 |
| Mg (mg/kg) | 194.31 ± 44.73 a | 314.83 ± 12.17 b | 400.35 ± 18.57 b | 12.89 | 0.001 |
| Ca (mg/kg) | 764.07 ± 116.66 a | 1118.54 ± 128.1 b | 1362.76 ± 84.4 b | 7.321 | 0.008 |
| Soil pH (KCl) | 5.00 ± 0.14 | 5.00 ± 0.09 | 5.00 ± 0.05 | 0.03 | 0.997 |
| Zn (mg/kg) | 1.52 ± 0.50 a | 2.41 ± 0.36 b | 3.88 ± 0.61 c | 6.17 | 0.014 |
| Mn (mg/kg) | 77.66 ± 13.44 | 121.18 ± 23.36 | 117.34 ± 13.42 | 1.92 | 0.189 |
| Cu (mg/kg) | 4.38 ± 0.07 a | 3.63 ± 0.15 b | 3.39 ± 0.26 b | 8.44 | 0.005 |
| C (%) | 3.32 ± 0.26 a | 2.94 ± 0.29 a | 1.34 ± 0.31 b | 8.07 | 0.006 |
| N (%) | 0.12 ± 0.04 a | 0.21 ± 0.02 b | 0.25 ± 0.02 b | 6.52 | 0.012 |
| Forage Quality | CMF | COP | CF | F | p |
|---|---|---|---|---|---|
| T. triandra | |||||
| ADF (%) | 43.99 ± 0.61 | 43.38 ± 0.53 | 45.89 ± 1.99 | 1.1 | 0.392 |
| NDF (%) | 77.81 ± 0.04 | 78.20 ± 0.53 | 82.05 ± 1.94 | 4.05 | 0.077 |
| CP (%) | 4.29 ± 0.29 | 5.02 ± 0.62 | 5.57 ± 0.68 | 1.31 | 0.337 |
| T. leucothrix | |||||
| ADF (%) | 40.87 ± 2.79 | 42.43 ± 1.49 | 40.19 ± 0.18 | 0.394 | 0.69 |
| NDF (%) | 75.77 ± 1.59 | 79.77 ± 2.04 | 75.5 ± 0.17 | 2.541 | 0.159 |
| CP (%) | 7.35 ± 0.49 | 7.47 ± 0.42 | 6.5 ± 0.03 | 1.946 | 0.223 |
| E. muticus | |||||
| ADF (%) | 40.37 ± 1.45 | 38.83 ± 0.38 | 40.06 ± 0.48 | 0.804 | 0.49 |
| NDF (%) | 74.41 ± 2.40 | 75.26 ± 0.39 | 78.20 ± 0.53 | 1.921 | 0.229 |
| CP (%) | 8.19 ± 0.11 a | 6.49 ± 0.08 b | 5.03 ± 0.62 c | 18.87 | 0.003 |
| CMF | COP | CF | F | p | |
|---|---|---|---|---|---|
| Grasses | |||||
| Diversity H′ (m−2) | 2.30 ± 0.05 a | 2.10 ± 0.06 b | 1.80 ± 0.09 c | 14.374 | <0.0001 |
| Evenness J (m−2) | 0.92 ± 0.02 | 0.91 ± 0.01 | 0.88 ± 0.01 | 1.65 | 0.210 |
| Richness (m−2) | 12.20 ± 0.55 a | 10.20 ± 0.51 b | 7.90 ± 0.59 c | 15.22 | <0.0001 |
| Forbs | |||||
| Diversity H′ (m−2) | 2.55 ± 0.02 a | 2.38 ± 0.03 b | 2.22 ± 0.02 c | 42.21 | <0.0001 |
| Evenness J (m−2) | 0.95 ± 0.01 | 0.93 ± 0.01 | 0.93 ± 0.01 | 3.26 | 0.054 |
| Richness (m−2) | 14.80 ± 0.39 a | 12.90 ± 0.43 b | 10.90 ± 0.28 c | 27.46 | <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chonco, N.; Slotow, R.; Tsvuura, Z.; Nkuna, S. Ecosystem Resilience of a South African Mesic Grassland with Change from Rotational to Continuous Grazing. Diversity 2023, 15, 1187. https://doi.org/10.3390/d15121187
Chonco N, Slotow R, Tsvuura Z, Nkuna S. Ecosystem Resilience of a South African Mesic Grassland with Change from Rotational to Continuous Grazing. Diversity. 2023; 15(12):1187. https://doi.org/10.3390/d15121187
Chicago/Turabian StyleChonco, Nomusa, Rob Slotow, Zivanai Tsvuura, and Sindiso Nkuna. 2023. "Ecosystem Resilience of a South African Mesic Grassland with Change from Rotational to Continuous Grazing" Diversity 15, no. 12: 1187. https://doi.org/10.3390/d15121187
APA StyleChonco, N., Slotow, R., Tsvuura, Z., & Nkuna, S. (2023). Ecosystem Resilience of a South African Mesic Grassland with Change from Rotational to Continuous Grazing. Diversity, 15(12), 1187. https://doi.org/10.3390/d15121187