
The Grassland Fragmentation Experiment in the Swiss Jura Mountains: A Synthesis


Abstract
:1. Introduction
2. A Replicated, Controlled 7-Year Fragmentation Experiment
2.1. Grasslands Examined
2.2. Fragmentation Experiment
2.3. Weather Conditions during the 7-Year Study
2.4. Methodology of the Surveys
2.5. Procedure of this Synthesis
3. Findings
3.1. High Species Richness in the Studied Grasslands
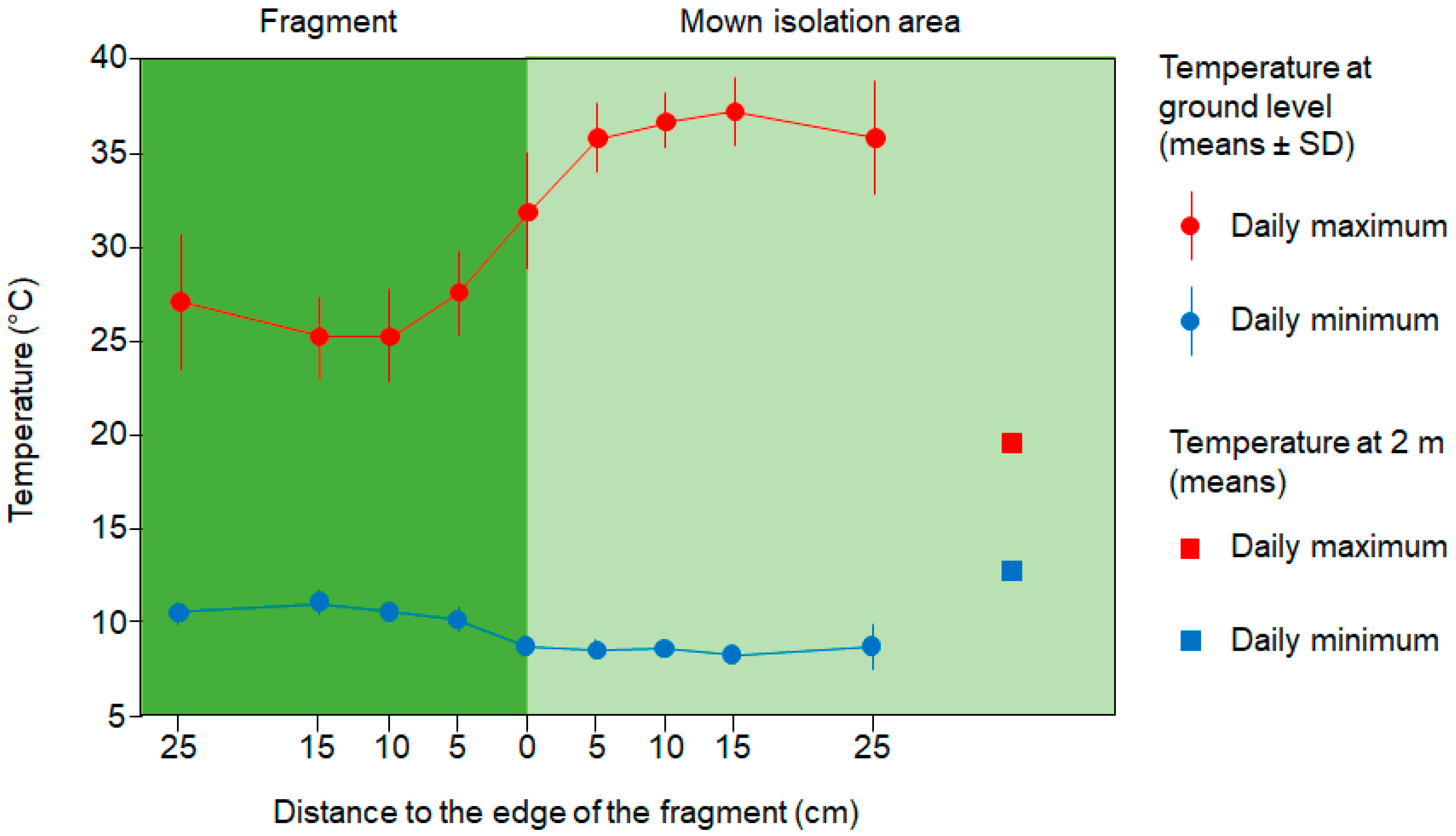
3.2. Increased Primary Productivity in the Edge Zone of Fragments
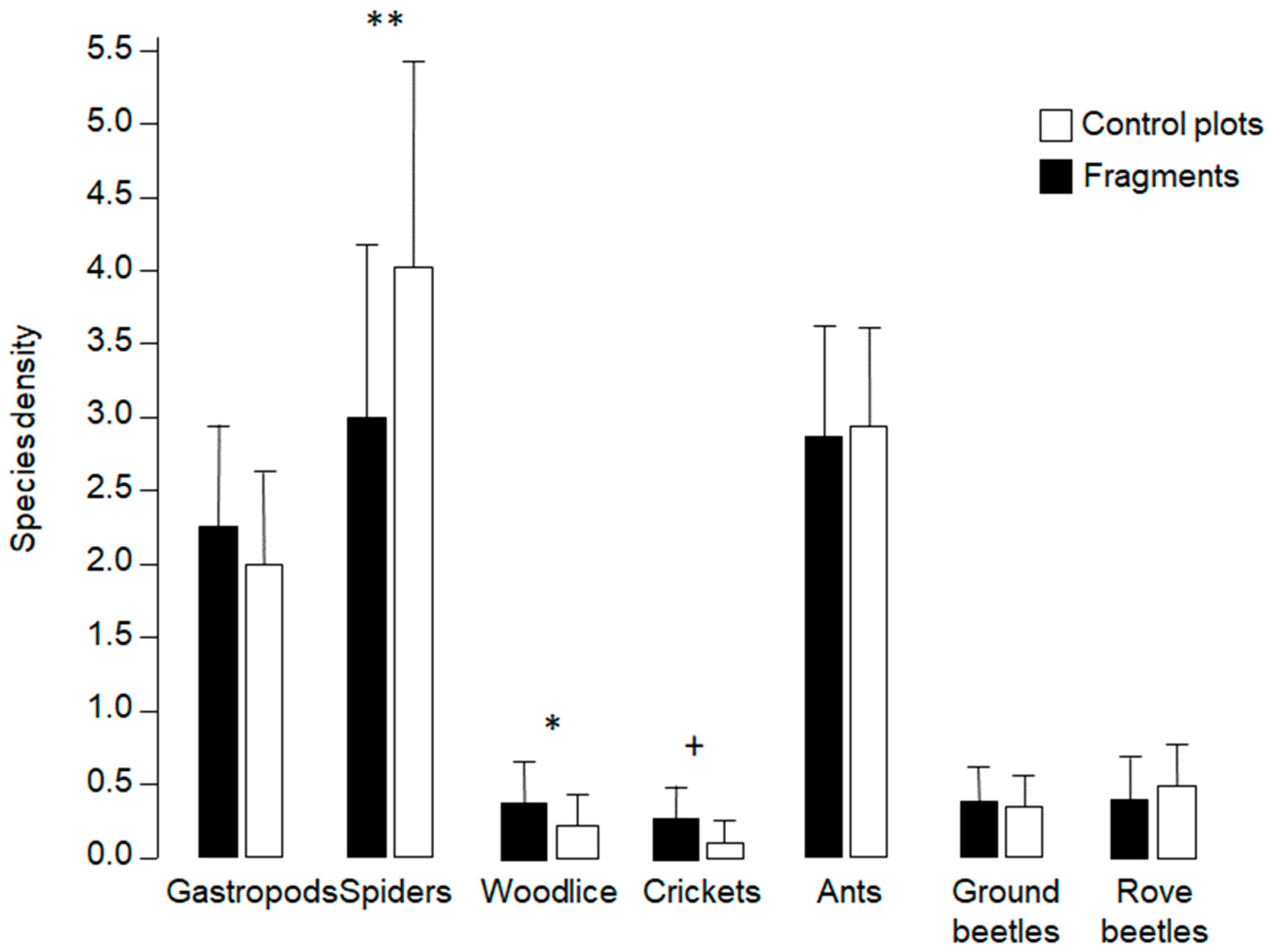
3.3. Differing Responses by Taxonomic Groups to Altered Conditions in Mown Isolation Area
3.4. Contrasting Effects on Species Richness and Diversity
3.4.1. Effects on Species Richness
3.4.2. Few Effects on Shannon Diversity and Species Turnover
3.5. Little Influence of Fragmentation on Species Composition but Huge Variation among Sites
3.6. Several Species Had Higher Abundance in Fragments
3.7. Fragmentation Affected Colonization and Extinction Rates in Some Species
3.8. Delayed Reduction in Functional Dispersion in Fragments
3.9. Species Interactions Responded Strongly to Fragmentation
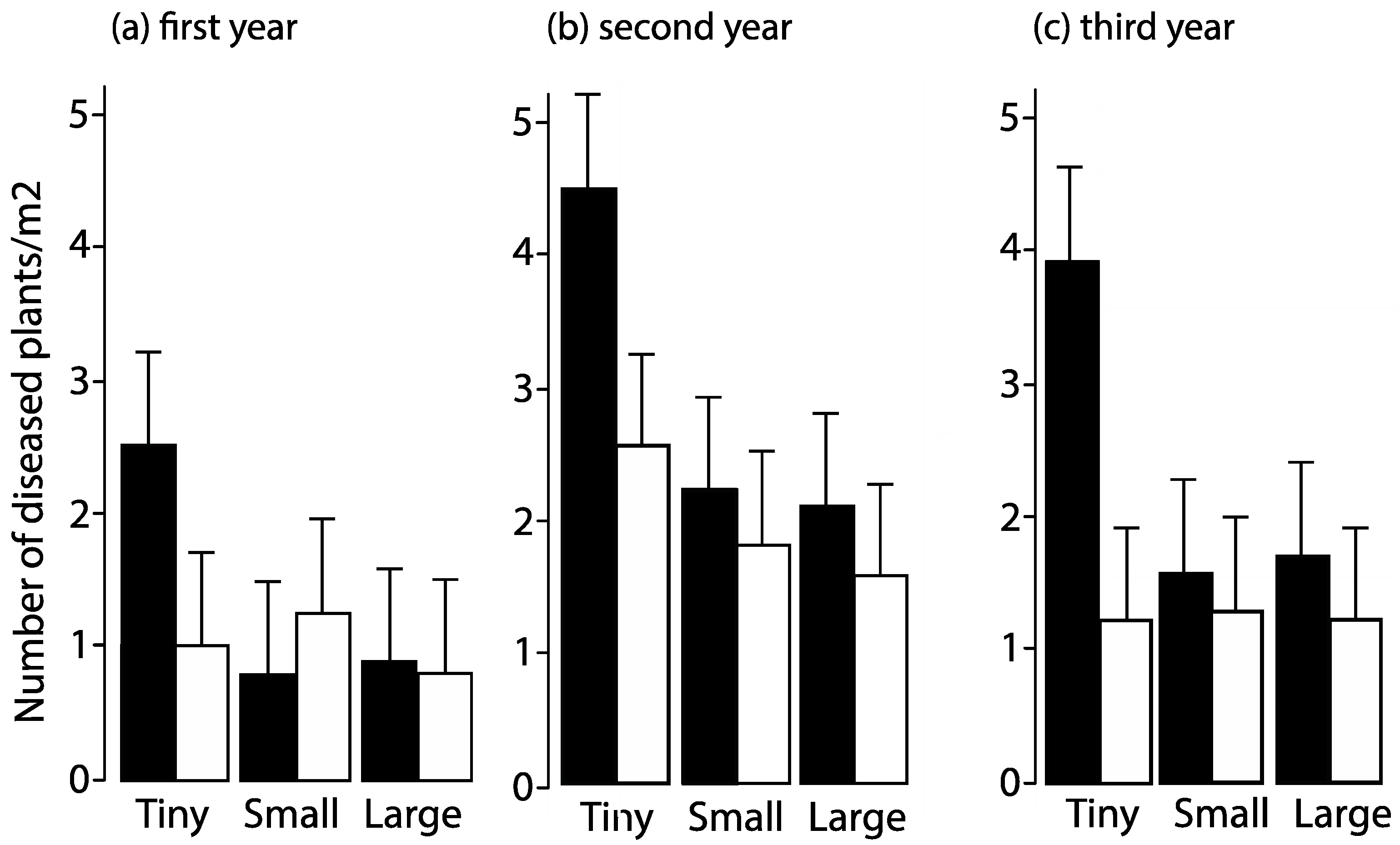
3.9.1. Changes in Herbivore Abundance Altered Extent of Plant Damage
3.9.2. Fragmentation Affected the Prevalence of a Fungal Disease
3.9.3. Altered Mutualistic and Competitive Interactions in Fragments
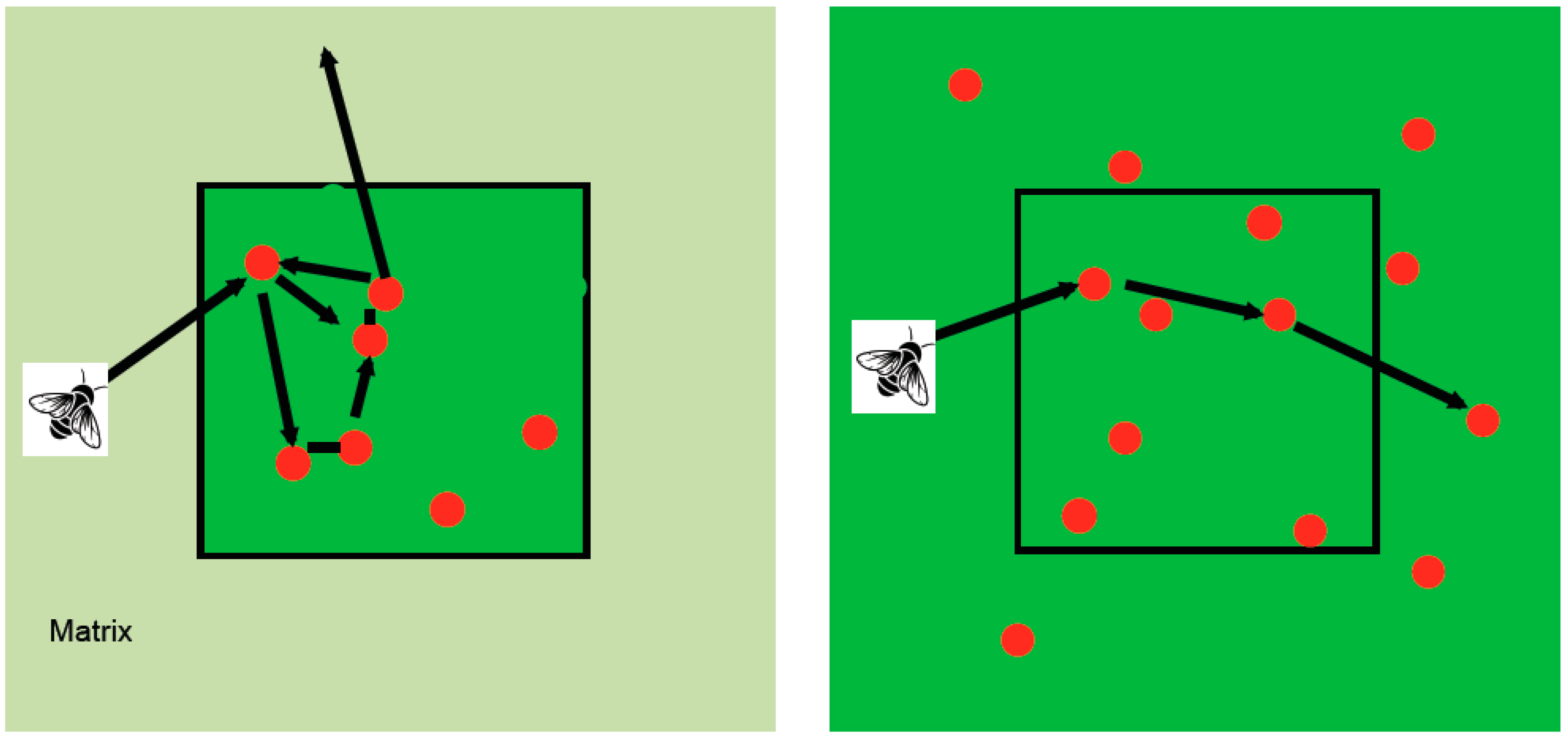
3.9.4. Fragmentation-Related Changes in Pollinator Behavior
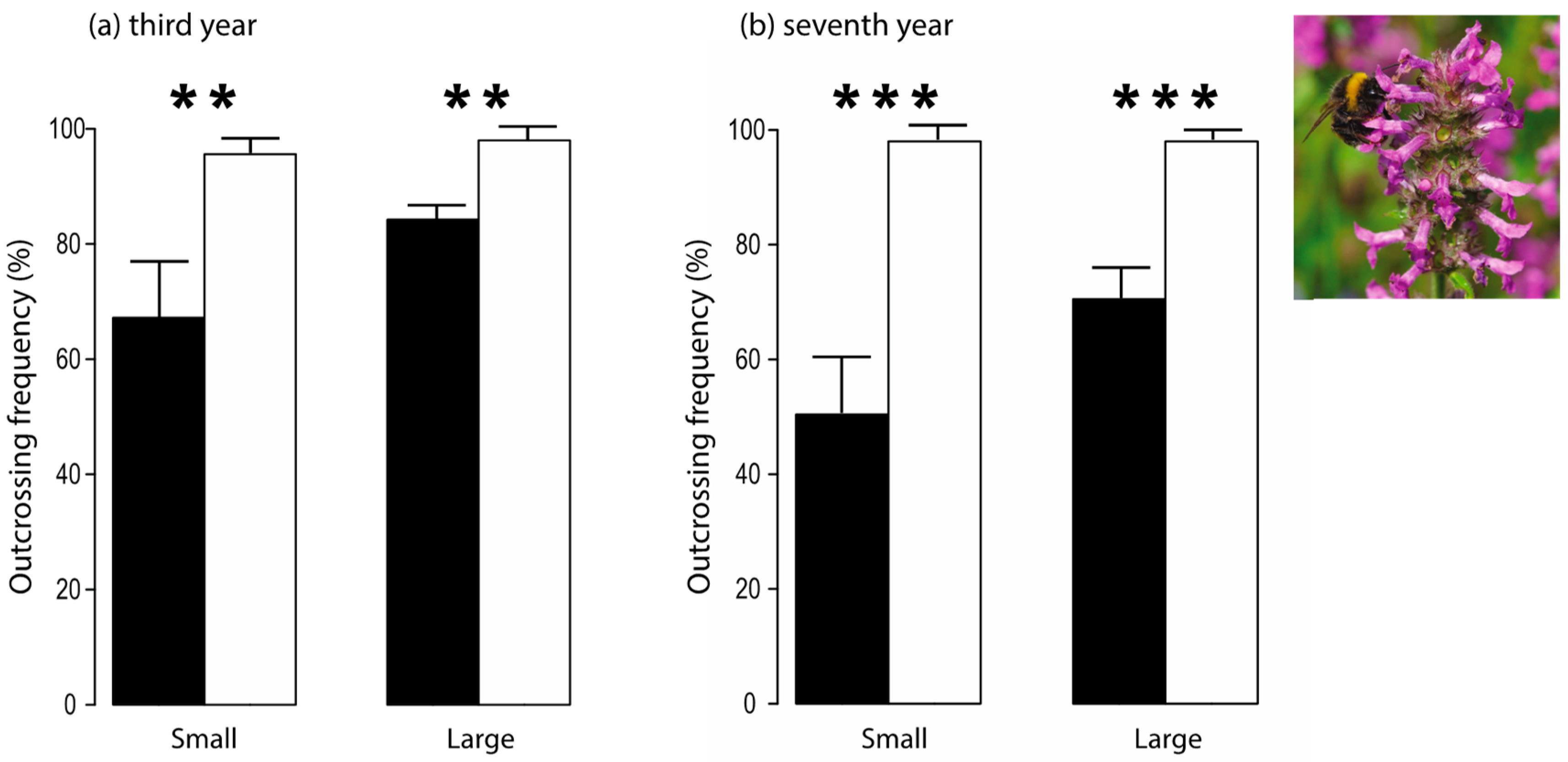
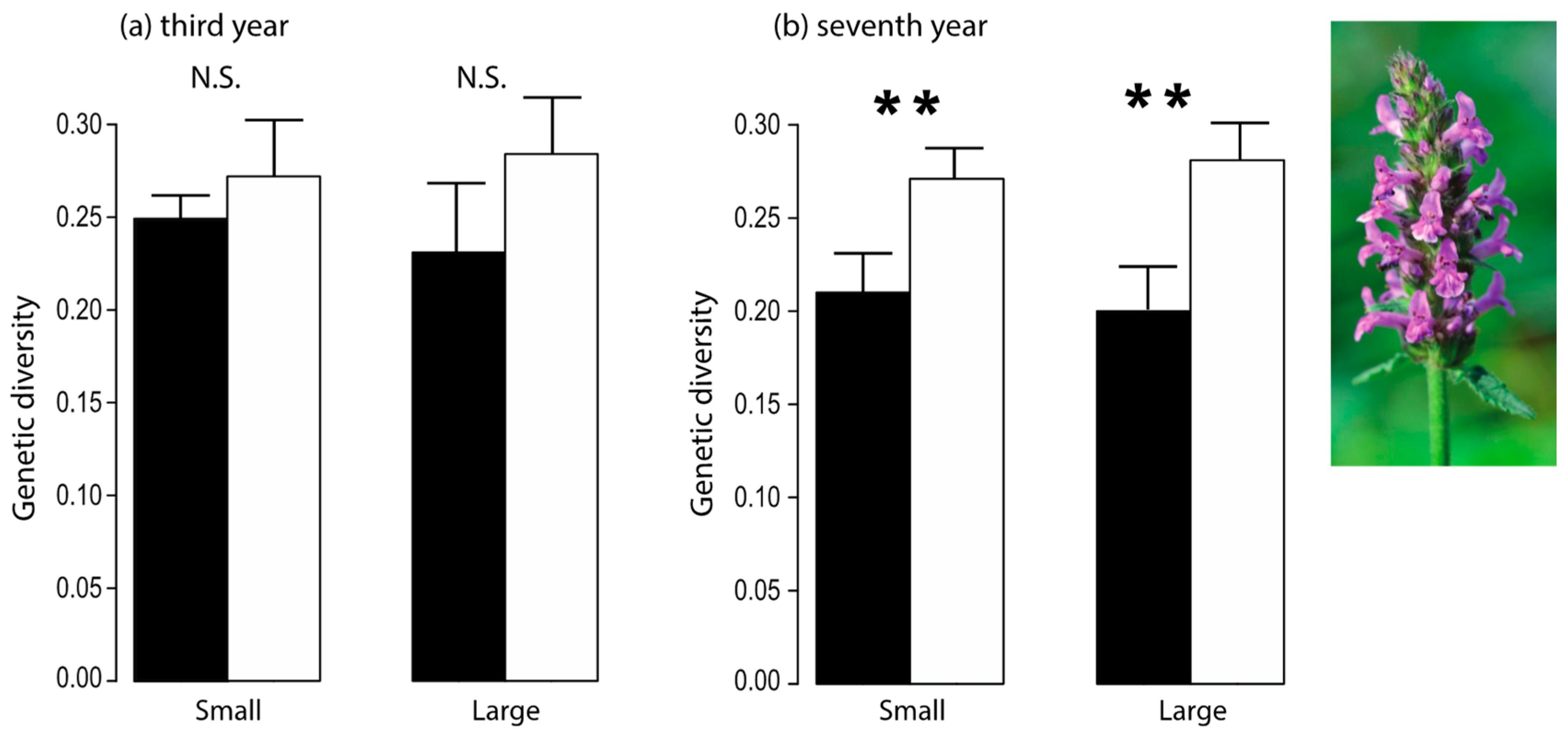
3.10. Fragmentation-Related Changes in Pollinator Behavior Led to Reduced Genetic Diversity in a Plant Species
3.11. Differing Responses to Fragmentation Can Be Explained by Species Traits
3.11.1. Smaller Species Were More Frequently Adversely Affected by Fragmentation
3.11.2. Species with Certain Life-History Traits Were More Abundant in Fragments
3.12. Species-Specific Humidity Preferences Could Hardly Explain Fragmentation Responses
3.13. Habitat Specialists Were Generally More Adversely Affected by Fragmentation
3.14. Long-Term Research Allowed Serendipitous Findings
3.14.1. Extending Biological Knowledge and Testing Methods
3.14.2. Expanding Faunistic Knowledge
4. Advantages and Limitations of Grassland Fragmentation Experiments
5. Lessons for Conservation
5.1. Even Mild Fragmentation Results in Noticeable Effects
5.2. Even a Small-Scale Fragmentation Matters
5.3. Assessment Shortly after Environmental Change Underestimates Long-Term Effects
5.4. Conservation Decisions Should Not Be Based on a Single Taxonomical Group
5.5. Biodiversity of a Grassland Does Not Substitute Those of Other Grasslands
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilson, M.C.; Chen, X.-Y.; Corlett, R.T.; Didham, R.K.; Ding, P.; Holt, R.D.; Holyoak, M.; Hu, G.; Hughes, A.C.; Jiang, L.; et al. Habitat fragmentation and biodiversity conservation: Key findings and future challenges. Landsc. Ecol. 2016, 31, 219–227. [Google Scholar] [CrossRef] [Green Version]
- Johnson, C.; Balmford, A.; Brook, B.; Buettel, J.; Galetti, M.; Lei, G.; Wilmshurst, J. Biodiversity losses and conservation responses in the Anthropocene. Science 2017, 356, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Quinn, J.F.; Hastings, A. Extinction in subdivided habitats. Conserv. Biol. 1987, 1, 198–208. [Google Scholar] [CrossRef]
- Saunders, D.A.; Hobbs, R.J.; Margules, C.R. Biological consequences of ecosystem fragmentation: A review. Conserv. Biol. 1991, 5, 18–32. [Google Scholar] [CrossRef]
- Margules, C.R.; Milkovits, G.A. Contrasting effects of habitat fragmentation on the scorpion Cercophonius squama and an amphipod. Ecology 1994, 75, 2033–2042. [Google Scholar] [CrossRef]
- Rossetti, M.R.; Tscharntke, T.; Aguilar, R.; Batáry, P. Responses of insect herbivores and herbivory to habitat fragmentation: A hierarchical meta-analysis. Ecol. Lett. 2017, 20, 264–272. [Google Scholar] [CrossRef] [Green Version]
- Davies, K.F.; Margules, C.R. Effects of habitat fragmentation on carabid beetles: Experimental evidence. J. Anim. Ecol. 1998, 67, 460–471. [Google Scholar] [CrossRef]
- Gaines, M.S.; Diffendorfer, J.E.; Tamarin, R.H.; Whittam, T.S. The effects of habitat fragmentation on the genetic structure of small mammal populations. J. Hered. 1997, 88, 294–304. [Google Scholar] [CrossRef]
- Lei, G.-C.; Hanski, I. Metapopulation structure of Cotesia melitaearum, a specialist parasitoid of the butterfly Melitaea cinxia. Oikos 1997, 78, 91–100. [Google Scholar] [CrossRef]
- Robinson, G.R.; Holt, R.D.; Gaines, M.S.; Hamburg, S.P.; Johnson, M.L.; Fitch, H.S.; Martinko, E.A. Diverse and contrasting effects of habitat fragmentation. Science 1992, 257, 524–526. [Google Scholar] [CrossRef]
- Simberloff, D. The contribution of population and community biology to conservation science. Annu. Rev. Ecol. Syst. 1988, 19, 473–511. [Google Scholar] [CrossRef]
- Kareiva, P.; Wennergren, U. Connecting landscape patterns to ecosystem and population processes. Nature 1995, 373, 299–302. [Google Scholar] [CrossRef]
- Wiens, J.A. Spatial scaling in ecology. Funct. Ecol. 1989, 3, 385–397. [Google Scholar] [CrossRef]
- Forman, R.T.T.; Godron, M. Landscape Ecology; Wiley: New York, NY, USA, 1986; p. 640. [Google Scholar]
- Wiens, J.A. Habitat fragmentation: Island versus landscape perspectives on bird conservation. Ibis 1994, 137, 97–104. [Google Scholar] [CrossRef]
- Tilman, D. Competition and biodiversity in spatially structured habitats. Ecology 1994, 75, 2–16. [Google Scholar] [CrossRef]
- Baur, B.; Erhardt, A. Habitat fragmentation and habitat alteration: Principal threats to most animal and plant species. GAIA 1995, 4, 221–226. [Google Scholar] [CrossRef]
- Schlaepfer, D.R.; Braschler, B.; Rusterholz, H.-P.; Baur, B. Genetic effects of anthropogenic habitat fragmentation on remnant animal and plant populations: A meta-analysis. Ecosphere 2018, 9, e02488. [Google Scholar] [CrossRef]
- Debinski, D.M.; Holt, R.D. A survey and overview of habitat fragmentation experiments. Conserv. Biol. 2000, 14, 342–355. [Google Scholar] [CrossRef]
- Zoller, H. Die Typen der Bromus erectus-Wiesen des Schweizer Juras, ihre Abhängigkeit von den Standortsbedingungen und wirtschaftlichen Einflüssen und ihre Beziehungen zur ursprünglichen Vegetation. In Series: Beiträge zur Geobotanischen Landesaufnahme der Schweiz, Vol. 33; Hans Huber: Bern, Switzerland, 1954; p. 309. [Google Scholar]
- Baur, B.; Joshi, J.; Schmid, B.; Hänggi, A.; Borcard, D.; Stary, J.; Pedroli-Christen, A.; Thommen, G.H.; Luka, H.; Rusterholz, H.-P.; et al. Variation in species richness of plants and diverse groups of invertebrates in three calcareous grasslands of the Swiss Jura mountains. Rev. Suisse Zool. 1996, 103, 801–833. [Google Scholar] [CrossRef]
- Veen, P.; Jefferson, R.; de Smidt, J.; van der Straaten, J. Grasslands in Europe of High Nature Value; KNNV Publishing: Zeist, The Netherlands, 2009; p. 320. [Google Scholar]
- Fischer, M.; Stöcklin, J. Local extinctions of plants in remnants of extensively used calcareous grasslands 1950–85. Conserv. Biol. 1997, 11, 727–737. [Google Scholar] [CrossRef]
- Zoller, H.; Bischof, N. Stufen der Kulturintensität und ihr Einfluss auf Artenzahl und Artengefüge der Vegetation. Phytocoenologia 1980, 7, 33–51. [Google Scholar]
- Boschi, C.; Baur, B. Past pasture management affects the land snail diversity in nutrient-poor calcareous grasslands. Basic Appl. Ecol. 2008, 9, 752–761. [Google Scholar] [CrossRef]
- Zoller, H.; Wagner, C.; Frey, V. Nutzungsbedingte Veränderungen in Mesobromion-Halbtrockenrasen in der Region Basel—Vergleich 1950–1980. Abh. Westfäl. Mus. Naturk. 1986, 48, 93–107. [Google Scholar]
- Zoller, H. Studien an Bromus erectus-Trockenrasengesellschaften in der Nordwestschweiz, speziell im Blauengebiet. Ber. Geobot. Inst. ETH 1947, 51–81. [Google Scholar]
- Zschokke, S.; Dolt, C.; Rusterholz, H.-P.; Oggier, P.; Braschler, B.; Thommen, G.H.; Lüdin, E.; Erhardt, A.; Baur, B. Short-term responses of plants and invertebrates to experimental small-scale grassland fragmentation. Oecologia 2000, 125, 559–572. [Google Scholar] [CrossRef] [PubMed]
- MeteoSwiss. IDAWEB 1.3.5.0. Available online: https://gate.meteoswiss.ch/idaweb/login.do (accessed on 19 July 2022).
- Groppe, K.; Steinger, T.; Schmid, B.; Baur, B.; Boller, T. Effects of habitat fragmentation on choke disease (Epichloë bromicola) in the grass Bromus erectus. J. Anim. Ecol. 2001, 89, 247–255. [Google Scholar] [CrossRef]
- Goverde, M.; Schweizer, K.; Baur, B.; Erhardt, A. Small-scale fragmentation affects pollinator behaviour: Experimental evidence from the bumblebee Bombus veteranus on calcareous grasslands. Biol. Conserv. 2002, 104, 293–299. [Google Scholar] [CrossRef]
- Ledergerber, S.; Dolt, C.; Zschokke, S.; Baur, B. Effects of experimental small-scale grassland fragmentation on the extent of grazing damage in Trifolium repens seedlings. Acta Oecol. 2002, 23, 329–336. [Google Scholar] [CrossRef]
- Braschler, B.; Baur, B. Effects of experimental small-scale grassland fragmentation on spatial distribution, density, and persistence of ant nests. Ecol. Entomol. 2003, 28, 651–658. [Google Scholar] [CrossRef]
- Braschler, B.; Lampel, G.; Baur, B. Experimental small-scale grassland fragmentation alters aphid population dynamics. Oikos 2003, 100, 581–591. [Google Scholar] [CrossRef] [Green Version]
- Braschler, B.; Baur, B. Experimental small-scale grassland fragmentation alters competitive interactions among ant species. Oecologia 2005, 143, 291–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolt, C.; Goverde, M.; Baur, B. Effects of experimental small-scale habitat fragmentation on above-ground and below-ground plant biomass in calcareous grasslands. Acta Oecol. 2005, 27, 49–56. [Google Scholar] [CrossRef]
- Stoll, P.; Dolt, C.; Goverde, M.; Baur, B. Experimental habitat fragmentation and invertebrate grazing in a herbaceous grassland species. Basic Appl. Ecol. 2006, 7, 307–319. [Google Scholar] [CrossRef]
- Joshi, J.; Stoll, P.; Rusterholz, H.-P.; Schmid, B.; Dolt, C.; Baur, B. Small-scale experimental habitat fragmentation reduces colonization rates in species-rich grasslands. Oecologia 2006, 148, 144–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braschler, B.; Marini, L.; Thommen, G.H.; Baur, B. Effects of small-scale grassland fragmentation and frequent mowing on population density and species diversity of orthopterans: A long-term study. Ecol. Entomol. 2009, 34, 321–329. [Google Scholar] [CrossRef]
- Stoll, P.; Oggier, P.; Baur, B. Population dynamics of six land snail species in experimentally fragmented grassland. J. Anim. Ecol. 2009, 78, 236–246. [Google Scholar] [CrossRef]
- Rusterholz, H.-P.; Baur, B. Delayed response in a plant-pollinator system to experimental grassland fragmentation. Oecologia 2010, 163, 141–152. [Google Scholar] [CrossRef]
- Braschler, B.; Baur, B. Diverse effects of a seven-year experimental grassland fragmentation on major invertebrate groups. PLoS ONE 2016, 11, e0149567. [Google Scholar] [CrossRef]
- Braschler, B.; Oggier, P.; Baur, B. Subtle effects of experimental grassland fragmentation on density, species composition and functional dispersion of gastropods. Diversity 2022, 14, 474. [Google Scholar] [CrossRef]
- Tilman, D.; Reich, P.B.; Knops, J.; Wedin, D.; Mielke, T.; Lehman, C. Diversity and productivity in a long-term grassland experiment. Science 2001, 294, 843–845. [Google Scholar] [CrossRef] [Green Version]
- Braschler, B.; Zschokke, S.; Dolt, C.; Thommen, G.H.; Oggier, P.; Baur, B. Grain-dependent relationships between plant productivity and invertebrate species richness and biomass in calcareous grasslands. Basic Appl. Ecol. 2004, 5, 15–24. [Google Scholar] [CrossRef]
- Chase, J.M.; Leibold, M.A. Spatial scale dictates the productivity-biodiversity relationship. Nature 2002, 416, 427–430. [Google Scholar] [CrossRef] [PubMed]
- Ricketts, T.H. The matrix matters: Effective isolation in fragmented landscapes. Am. Nat. 2001, 158, 87–99. [Google Scholar] [CrossRef]
- Laurance, W.F.; Camargo, J.L.C.; Luizao, R.C.C.; Laurance, S.G.; Pimm, S.L.; Bruna, E.M.; Stouffer, P.C.; Willamson, G.B.; Benitez-Malvido, J.; Vasconcelos, H.L.; et al. The fate of Amazonian forest fragments: A 32-year investigation. Biol. Conserv. 2011, 144, 56–67. [Google Scholar] [CrossRef]
- Fahrig, L. Ecological response to habitat fragmentation per se. Annu. Rev. Ecol. Syst. 2017, 48, 1–23. [Google Scholar] [CrossRef]
- Gallé, R.; Tolgyesi, C.; Torma, A.; Bátori, B.; Lörinczi, G.; Szilassi, P.; Gallé-Szpisjak, N.; Kaur, H.; Makra, T.; Módra, G.; et al. Matrix quality and habitat type drive the diversity pattern of forest steppe fragments. Perspect. Ecol. Conserv. 2022, 20, 60–68. [Google Scholar] [CrossRef]
- Rusterholz, H.-P. Flower Densities and Bumblebee Foraging Behavior; University of Basel: Basel, Switzerland, 2010; unpublished work. [Google Scholar]
- Hudson, L.N.; Newbold, T.; Contu, S.; Hill, S.L.L.; Lysenko, I.; De Palma, A.; Phillips, H.R.P.; Alhusseini, T.I.; Bedford, F.E.; Bennett, D.J.; et al. The database of the PREDICTS (Projecting Responses of Ecological Diversity in Changing Terrestrial Systems) project. Ecol. Evol. 2017, 7, 145–188. [Google Scholar] [CrossRef] [Green Version]
- Magurran, A.E. Measuring Biological Diversity; Wiley-Blackwell: Hoboken, NJ, USA, 2013; p. 272. [Google Scholar]
- Southwood, T.R.E.; Henderson, P.A. Ecological Methods, 3rd ed.; Blackwell Science Ltd.: Oxford, UK, 2000; p. 575. [Google Scholar]
- Hanski, I. Metapopulation Ecology; Oxford University Press: Oxford, UK, 1999; p. 313. [Google Scholar]
- Fahrig, L. Effect of habitat fragmentation on the extinction threshold: A synthesis. Ecol. Appl. 2002, 12, 346–353. [Google Scholar]
- Koleff, P.; Gaston, K.J.; Lennon, J.J. Measuring beta diversity for presence-absence data. J. Anim. Ecol. 2003, 72, 367–382. [Google Scholar] [CrossRef] [Green Version]
- Wallis DeVries, M.F.; Poschlod, P.; Willems, J.H. Challenges for the conservation of calcareous grasslands in northwestern Europe: Integrating the requirements of flora and fauna. Biol. Conserv. 2002, 104, 265–273. [Google Scholar] [CrossRef]
- Poschlod, P. Geschichte der Kulturlandschaft; Ulmer Verlag: Stuttgart, Germany, 2015; p. 320. [Google Scholar]
- Hölldobler, B.; Wilson, E.O. The Ants; Springer: Berlin, Germany, 1990; p. 732. [Google Scholar]
- New, T.R. An Introduction to Invertebrate Conservation Biology; Oxford University Press: Oxford, UK, 1995; p. 194. [Google Scholar]
- Samways, M.J. Insect Diversity Conservation; Cambridge University Press: Cambridge, UK, 2005; p. 342. [Google Scholar]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967; p. 203. [Google Scholar]
- Oggier, P. Circadian and year-round activity of the land snails Candidula unifasciata and Helicella itala in grasslands of the Swiss Jura mountains (Gastropoda: Stylommatophora: Helicidae). Malakol. Abh. 1998, 19, 89–101. [Google Scholar]
- Laliberté, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef]
- Gossner, M.M.; Simons, N.K.; Achtziger, R.; Blick, T.; Dorow, W.H.O.; Dziock, F.; Köhler, F.; Rabitsch, W.; Weisser, W.W. A summary of eight traits of Coleoptera, Hemiptera, Orthoptera and Araneae, occurring in grasslands in Germany. Sci. Data 2015, 2, 150013. [Google Scholar] [CrossRef] [PubMed]
- Orthoptera.ch. Orthoptera.ch. Available online: https://www.orthoptera.ch (accessed on 21 November 2022).
- Kareiva, P. Habitat fragmentation and the stability of predator-prey interactions. Nature 1987, 326, 388–390. [Google Scholar] [CrossRef]
- Kruess, A.; Tscharntke, T. Habitat fragmentation, species loss, and biological control. Science 1994, 264, 1581–1584. [Google Scholar] [CrossRef] [PubMed]
- Grez, A.A.; Zaviezo, T.; Diaz, S.; Camousseigt, B.; Cortés-Hinojosa, G.A. Effects of habitat loss and fragmentation on the abundance and species richness of aphidophageous beetles and aphids in experimental alfalfa landscapes. Eur. J. Entomol. 2008, 105, 411–420. [Google Scholar] [CrossRef] [Green Version]
- Dubbert, M.; Tscharntke, T.; Vidal, S. Stem-boring insects of fragmented Calamagrostis habitats: Herbivore-parasitoid community structure and the unpredictability of grass shoot abundance. Ecol. Entomol. 1998, 23, 271–280. [Google Scholar] [CrossRef]
- Bitters, M.E.; Meyers, J.; Resasco, J.; Sarre, S.D.; Tuff, K.T.; Davies, K.F. Experimental habitat fragmentation disrupts host-parasite interaction over decades via life-cycle bottlenecks. Ecology 2022, 103, e3758. [Google Scholar] [CrossRef]
- LaSalle, J.; Gauld, I.D. Parasitic hymenoptera and the biodiversity crisis. Redia 1991, 74, 315–334. [Google Scholar]
- Marjakangas, E.-L.; Abrego, N.; Grøtan, V.; de Lima, A.F.; Bello, C.; Bovendorp, R.S.; Culot, L.; Hasui, É.; Lima, F.; Muylaert, R.L.; et al. Fragmented tropical forests lose mutualistic plant-animal interactions. Divers. Distrib. 2019, 26, 154–168. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, J.; Berens, D.G.; Jaroszewicz, B.; Selva, N.; Brandl, R.; Farwig, N. Correlated loss of ecosystem services in coupled mutualistic networks. Nat. Commun. 2014, 5, 3810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herbers, J.M. Seasonal structuring of a north temperate ant community. Insectes Soc. 1985, 32, 224–240. [Google Scholar] [CrossRef]
- Herbers, J.M. Community structure in north temperate ants: Temporal and spatial variation. Oecologia 1989, 81, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Seifert, B. Die Ameisen Mittel- und Nordeuropas; Lutra—Verlags- und Vertriebsgemeinschaft: Tauer, Germany, 2007; p. 368. [Google Scholar]
- Traniello, J.F.A. Foraging strategies of ants. Ann. Rev. Entomol. 1989, 34, 191–210. [Google Scholar] [CrossRef]
- Potts, S.G.; Bismeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 6, 345–353. [Google Scholar] [CrossRef]
- Vanbergen, A.J. Threats to an ecosystem service: Pressures on pollinators. Front. Ecol. Environ. 2013, 11, 251–259. [Google Scholar] [CrossRef]
- Aguilar, R.; Cristobal-Perez, E.J.; Balvino-Olvera, F.J.; Aguilar-Aguilar, M.D.; Aquirre-Acosta, N.; Ashworth, L.; Lobo, J.A.; Marten-Rodriguez, S.; Fuchs, E.J.; Sanchez-Montoya, G.; et al. Habitat fragmentation reduces plant progeny quality: A global analysis. Ecol. Lett. 2019, 7, 1163–1173. [Google Scholar] [CrossRef] [Green Version]
- Zimmerman, M. Optimal foraging, plant density and the marginal value theorem. Oecologia 1981, 49, 148–153. [Google Scholar] [CrossRef]
- Chittka, L.; Thomson, J.D. Sensori-motor learning and its relevance for task specialization in bumble bees. Behav. Ecol. Sociobiol. 1997, 41, 385–398. [Google Scholar] [CrossRef]
- Cresswell, J.E. A comparison of bumblebees’ movements in uniform and aggregated distribution of their forage plant. Ecol. Entomol. 2000, 25, 19–25. [Google Scholar] [CrossRef]
- Jennersten, O. Pollination of Dianthus deltoides (Caryophyllaceae), effects of habitat fragmentation on visitation and seed set. Conserv. Biol. 1988, 2, 359–366. [Google Scholar] [CrossRef]
- Kearns, C.A.; Inouye, D.W.; Waser, N.M. Endangered mutualisms, the conservation of plant–pollinator interactions. Ann. Rev. Ecol. Syst. 1998, 29, 83–112. [Google Scholar] [CrossRef]
- Aizen, M.A.; Ashworth, L.; Galetto, L. Reproductive success in fragmented habitats: Do compatibility system and pollination specialization matter? J. Veg. Sci. 2002, 13, 885–892. [Google Scholar] [CrossRef]
- Aguilar, R.; Ashworth, L.; Galetto, L.; Aizen, M.A. Plant reproductive susceptibility to habitat fragmentation: Review and synthesis through a meta-analysis. Ecol. Lett. 2006, 9, 968–980. [Google Scholar] [CrossRef]
- Reed, D.; Lowe, E.; Briscoe, D.; Frankham, R. Inbreeding and extinction: Effects of rate of inbreeding. Conserv. Genet. 2003, 4, 405–410. [Google Scholar] [CrossRef]
- Lowe, A.J.; Boshier, D.; Ward, M.; Bacles, C.F.E.; Navarro, C. Genetic resource impacts of habitat loss and degradation; reconciling empirical evidence and predicted theory for neotropical trees. Heredity 2005, 95, 255–273. [Google Scholar] [CrossRef]
- Leimu, R.; Mutikainen, P.; Koricheva, J.; Fischer, M. How general are positive relationships between plant population size, fitness and genetic variation? J. Ecol. 2006, 94, 942–952. [Google Scholar] [CrossRef]
- Charlesworth, D.; Willis, J.H. The genetics of inbreeding depression. Nat. Rev. Genet. 2009, 10, 783–796. [Google Scholar] [CrossRef]
- Jump, A.S.; Marchant, R.; Penuelas, P. Environmental change and the option value of genetic diversity. Trends Plant Sci. 2009, 14, 51–58. [Google Scholar] [CrossRef]
- Manel, S.; Holderegger, R. Ten years of landscape genetics. Trends Ecol. Evol. 2013, 28, 614–621. [Google Scholar] [CrossRef]
- Gonzalez, A.V.; Gomez-Silva, V.; Ramirez, M.J.; Fonturbel, F.E. Meta-analysis of the differential effects of habitat fragmentation and degradation on plant genetic diversity. Conserv. Biol. 2020, 34, 711–720. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, R.; Quesada, M.; Ashworth, L.; Herrerias-Diego, Y.; Lobo, A. Genetic consequences of habitat fragmentation in plant populations: Susceptible signals in plant traits and methodological approaches. Mol. Ecol. 2008, 17, 5177–5188. [Google Scholar] [CrossRef] [PubMed]
- Eckert, C.G.; Kalisz, S.; Geber, M.A.; Sargent, R.; Elle, E.; Cheptou, P.-O.; Goodwillie, C.; Johnston, M.O.; Kelly, J.K.; Moeller, D.A.; et al. Plant mating systems in a changing world. Trends Ecol. Evol. 2010, 25, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Nei, M. Analysis of gene diversity in subdivided populations. Proc. Natl. Acad. Sci. USA 1973, 70, 3321–3323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tscharntke, T.; Brandl, R. Plant-insect interactions in fragmented landscapes. Annu. Rev. Entomol. 2004, 49, 405–430. [Google Scholar] [CrossRef] [PubMed]
- Grass, I.; Jauker, B.; Steffan-Dewenter, I.; Tscharntke, T.; Jauker, F. Past and potential future effects of habitat fragmentation on structure and stability of plant-pollinator and host-parasitoid networks. Nat. Ecol. Evol. 2018, 2, 1408–1417. [Google Scholar] [CrossRef]
- Greenleaf, S.S.; Williams, N.M.; Winfree, R.; Kremen, C. Bee foraging ranges and their relationship to body size. Oecologia 2007, 153, 589–596. [Google Scholar] [CrossRef]
- Oggier, P.; Zschokke, S.; Baur, B. A comparison of three methods for assessing the gastropod community in dry grasslands. Pedobiologia 1998, 42, 348–357. [Google Scholar]
- Mittelbach, G.G.; Steiner, C.F.; Scheiner, S.M.; Gross, K.L.; Reynolds, H.L.; Waide, R.B.; Willig, M.R.; Dodson, S.I.; Gough, L. What is the observed relationship between species richness and productivity. Ecology 2001, 82, 2381–2396. [Google Scholar] [CrossRef]
- Niemelä, J.; Baur, B. Threatened species in a vanishing habitat: Plants and invertebrates in calcareous grasslands in the Swiss Jura mountains. Biodivers. Conserv. 1998, 7, 1407–1416. [Google Scholar] [CrossRef]
- Borcard, Y.; Borcard, D.; Vernier, R. Epitritus argiolus Emery, 1869 (Hymenoptera, Formicidae) dans une prairie maigre du Jura. Mitt. Schweiz. Entomol. Ges. 1997, 70, 265–269. [Google Scholar]
- Baroni Urbani, C. Strumigenys baudueri (Emery): Espèce nouvelle pour la Suisse (Hymenoptera: Formicidae). Mitt. Schweiz. Entomol. Ges. 1998, 71, 163–164. [Google Scholar]
- Braschler, B. Neue Aspekte zur Verbreitung von Pyramica baudueri (Emery, 1875) (Hymenoptera, Formicidae). Mitt. Entomol. Ges. Basel 2002, 52, 139–142. [Google Scholar]
- Doak, D.F.; Marino, P.C.; Kareiva, P.M. Spatial scale mediates the influence of habitat fragmentation on dispersal success: Implications for conservation. Theor. Popul. Biol. 1992, 41, 315–336. [Google Scholar] [CrossRef]
- Cattarino, L.; McAlpine, C.A.; Rhodes, J.R. Spatial scale and movement behaviour traits control the impacts of habitat fragmentation on individual fitness. J. Anim. Ecol. 2016, 85, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Baur, A.; Baur, B. Are roads barriers to dispersal in the land snail Arianta arbustorum? Can. J. Zool. 1990, 68, 613–617. [Google Scholar] [CrossRef]
- Wirth, T.; Oggier, P.; Baur, B. Effect of road width on dispersal and genetic population structure in the land snail Helicella itala. Z. Ökol. Natursch. 1999, 8, 23–29. [Google Scholar]
- Bhattacharya, M.; Primack, R.B.; Gerwein, J. Are roads and railroads barriers to bumblebee movement in a temperate suburban conservation area? Biol. Conserv. 2003, 109, 37–45. [Google Scholar] [CrossRef]
- Ferraz, G.; Russell, G.J.; Stouffer, P.C.; Bierregaard, R.O.; Pimm, S.L.; Lovejoy, T.E. Rates of species loss from Amazonian forest fragments. Proc. Natl. Acad. Sci. USA 2003, 100, 14069–14073. [Google Scholar] [CrossRef] [Green Version]
- Krauss, J.; Bommarco, R.; Guardiola, M.; Heikkinen, R.K.; Helm, A.; Kuussaari, M.; Lindborg, R.; Öckinger, E.; Pärtel, M.; Pino, J.; et al. Habitat fragmentation causes immediate and time-delayed biodiversity loss at different trophic levels. Ecol. Lett. 2010, 15, 597–605. [Google Scholar] [CrossRef] [Green Version]
- Collinge, S.K. Effects of grassland fragmentation on insect species loss, colonization, and movement patterns. Ecology 2000, 81, 2211–2226. [Google Scholar] [CrossRef]
- Tewksbury, J.J.; Levey, D.J.; Haddad, N.M.; Sargent, S.; Orrock, J.L.; Weldon, A.; Danielson, B.J.; Brinkerhoff, J.; Damschen, E.I.; Townsend, P. Corridors affect plants, animals, and their interactions in fragmented landscapes. Proc. Nat. Acad. Sci. USA 2002, 99, 12923–12926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinhardt, K.; Köhler, G.; Maas, S.; Detzel, P. Low dispersal ability and habitat specificity promote extinctions in rare but not in widespread species: The Orthoptera of Germany. Ecography 2005, 28, 593–602. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Annual Precipitation [mm; % Deviation] | Mean Annual Temperature [°C; % Deviation] | Mean Temperature in Apr–Sept [°C; % Deviation] | Min Temperature in Apr–Sept [°C] | Max Temperature in Apr–Sept [°C] |
|---|---|---|---|---|---|
| 1993 | 881.0 (−4.2) | 9.2 (−6.5) | 14.6 (−4.8) | −1.6 | 33.4 |
| 1994 | 936.8 (+1.9) | 10.7 (+8.7) | 15.5 (+0.5) | −2.6 | 34.6 |
| 1995 | 1118.9 (+21.7) | 9.5 (−3.5) | 14.4 (−6.2) | −1.1 | 34.2 |
| 1996 | 832.9 (−9.4) | 8.4 (−14.7) | 13.9 (−9.3) | −3.8 | 30.9 |
| 1997 | 815.1 (−11.3) | 9.3 (−5.5) | 14.6 (−5.1) | −4.8 | 30.2 |
| 1998 | 925.0 (+0.6) | 9.2 (−6.5) | 14.8 (−3.8) | −3.0 | 35.4 |
| 1999 | 1275.3 (+38.7) | 9.8 (−0.4) | 15.7 (+2.3) | −0.4 | 32.8 |
| Norm a | 919.4 a | 9.8 a | 15.4 a | −6.4 b | 37.7 b |
| Publication | Issue(s) Addressed | Year(s) of Experiment a | Organism Group(s) | Method(s) Applied b |
|---|---|---|---|---|
| Zschokke et al. 2000 [28] | Assessing species richness and species diversity in several taxonomical groups, recording the relative abundance of the 65 most common species | 4 | Grasses, forbs, ants, butterflies, grasshoppers, and gastropods | Classical plant survey, nest counting in ants, direct census of butterflies and grasshoppers, wet cardboard sheets as traps for gastropods |
| Groppe et al. 2001 [30] | Temporal and among-site variation in fungal infestation of a grass species | 1–3 | Target species: Bromus erectus; Fungus: Epichloë bromicola | Mapping Bromus erectus inflorescences and assessing the fungal infestation of the inflorescences |
| Goverde et al. 2002 [31] | Flower visitation rate, taxonomic composition of flower visitors, and bumblebee foraging behavior | 6 | Target species: Stachys officinalis; Pollinator: mainly Bombus veteranus but also other flower visitors | Mapping the spatial distribution of S. officinalis plants in small fragments and control plots, recording frequency of different flower visitors, and foraging behavior of B. veteranus in three blocks at two study sites |
| Ledergerber et al. 2002 [32] | Quantifying the extent of grazing damage to an herb | 4 | Target species: Trifolium repens, potential grazers: 12 gastropod species, 15 grasshopper species | Provision of standardized food (seedlings of Trifolium repens) in two short periods |
| Braschler & Baur 2003 [33] | Inter- and intraspecific competition; edge effects | 4, 7 | Ants (all species; Lasius paralienus separately) | Spatial distribution of ant nests |
| Braschler et al. 2003 [34] | Infestation rates and abundances of aphids; parasitism rates of aphids; ant-tending rates | 5−7 | Aphids (all species present); plant species infested by aphids; ant species tending aphids | Counting aphids and aphid mummies, only two study sites |
| Braschler & Baur 2005 [35] | Interspecific competition and dominance | 4, 7 | Ants (all species; Lasius paralienus separately) | Counting ants at baits and natural sugar resources; pitfall traps; ant nest density, only small and large fragments |
| Dolt et al. 2005 [36] | Quantifying above- and below-ground plant biomass | 4−6 | Above-ground biomass of grasses and herbs, and overall below-ground plant biomass | Harvesting of plant biomass above a height of 5 cm in 3 years; harvesting of root-biomass using soil cores in the 6th year |
| Stoll et al. 2006 [37] | Extent of grazing damage on leaves | 6 | Target species: Stachys officinalis; potential grazers: gastropods, grasshoppers | Assessment of leaf area consumed in spring, summer, and autumn; two small and large fragments each in two randomly chosen blocks per site |
| Joshi et al. 2006 [38] | Colonization and extinction rate of plants | 1, 2, 3, 4, 7 | Vascular plants | Standardized vegetation survey |
| Braschler et al. 2009 [39] | Effects of fragmentation and frequent mowing on orthopteran species richness, density, and species composition | 1−7 | Orthopterans, also the suborders Ensifera and Caelifera separately | Visual survey in large fragments and control plots, and corresponding plots in the mown matrix; surveys several times per season in each year |
| Stoll et al. 2009 [40] | Quantifying the population dynamics (population sizes, extinction, and recolonization frequencies) of six land snail species | 2−4 | Six gastropod species (Cochlicopa lubrica, Vertigo pygmaea, Pupilla muscorum, Punctum pygmaeum, Helicella itala, and Trichia plebeia) | Mark-release-resight approach |
| Rusterholz & Baur 2010 [41] | Flower visitation rate, foraging behavior of pollinators, plant reproductive characteristics, outcrossing frequency, genetic diversity | 1, 4, 7 | Target species: Stachys officinalis; flower visiting insects, bumblebee species | Counting the number of insects visiting S. officinalis inflorescences; recording the foraging behavior of bumblebees; assessing seed characteristics; determining of outcrossing frequency and genetic diversity of seedlings using RAPD in the fourth and seventh year. |
| Braschler & Baur 2016 [42] | Species richness, density, species composition, body size, and habitat preferences | 7 | Gastropods, ants, ground beetles, rove beetles, Orthoptera, spiders, woodlice | Two pitfall traps per large and small fragment and control plot, emptied every 2 weeks for a total of seven collections from late spring to late summer |
| Braschler et al. 2022 [43] | Temporal and among-site variation in species richness, individual density, species composition, functional dispersion, morphological and life-history traits, and habitat preferences | 4−7 | Gastropods (all gastropods or snails only) | 81 cardboard traps, one night per year in autumn in large plots |
| Taxonomic Group | Movelier | Nenzlingen | Vicques | Overall | Year and Sampling Effort |
|---|---|---|---|---|---|
| Vascular plants | 116 | 111 | 96 | 143 | 1993 and 1994; all fragments and control plots [21] |
| Gastropods | 16 | 21 | 15 | 22 | 1994; all fragments and control plots [21] |
| Gastropods | 15 | 14 | 10 | 19 | 1996–1999; cardboard traps in all large fragments and control plots [43] |
| Gastropods | 18 | 18 | 10 | 22 | 1999; 2 pitfall traps each in all small and large fragments and control plots exposed for 14 weeks [42] |
| Spiders | 63 | 60 | 66 | 108 | 1994; 9 pitfall traps per site in the surroundings of blocks [21] |
| Spiders | 52 | 76 | 58 | 99 | 1999; 2 pitfall traps each in all small and large fragments and control plots exposed for 14 weeks [42] |
| Woodlice | 3 | 4 | 3 | 5 | 1999; 2 pitfall traps each in all small and large fragments and control plots exposed for 14 weeks [42] |
| Orthopterans | 16 | 13 | 10 | 17 | 1994; all fragments and control plots [21] |
| Orthopterans | 21 | 17 | 12 | 19 | 1993–1999; 2 surveys per year in large fragments and control plots and a comparable plot of mown matrix in each block [39] |
| Aphids | NA a | 13 | 17 | 24 | 1998; vouchers were collected from all aphid colonies in tiny fragments and control plots and subsamples in small and large fragments and control plots in Nenzlingen and Vicques [34] |
| Ants | 12 | 11 | 7 | 14 | 1996; nest counts assisted by following foragers attracted to sugar baits [28] |
| Ants | 16 | 20 | 15 | 24 | 1999; 2 pitfall traps each in all small and large fragments and control plots exposed for 14 weeks [42] |
| Butterflies | 46 | 32 | 40 | 46 | 1993 and 1994; three 10 m x 10 m plots per site adjacent to the blocks [21] |
| Butterflies | 26 | 19 | 20 | 29 | 1996; 13 × 30 min observation per block [28] |
| Ground beetles | 19 | 19 | 21 | 38 | 1994; 9 pitfall traps per site in the surroundings of blocks [21] |
| Ground beetles | 19 | 20 | 18 | 33 | 1999; 2 pitfall traps each in all small and large fragments and control plots exposed for 14 weeks [42] |
| Rove beetles | 14 | 25 | 16 | 35 | 1999; 2 pitfall traps each in all small and large fragments and control plots exposed for 14 weeks [42] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baur, B.; Rusterholz, H.-P.; Braschler, B. The Grassland Fragmentation Experiment in the Swiss Jura Mountains: A Synthesis. Diversity 2023, 15, 130. https://doi.org/10.3390/d15020130
Baur B, Rusterholz H-P, Braschler B. The Grassland Fragmentation Experiment in the Swiss Jura Mountains: A Synthesis. Diversity. 2023; 15(2):130. https://doi.org/10.3390/d15020130
Chicago/Turabian StyleBaur, Bruno, Hans-Peter Rusterholz, and Brigitte Braschler. 2023. "The Grassland Fragmentation Experiment in the Swiss Jura Mountains: A Synthesis" Diversity 15, no. 2: 130. https://doi.org/10.3390/d15020130
APA StyleBaur, B., Rusterholz, H. -P., & Braschler, B. (2023). The Grassland Fragmentation Experiment in the Swiss Jura Mountains: A Synthesis. Diversity, 15(2), 130. https://doi.org/10.3390/d15020130