

Modeling of the Elements Ca2+, Mg2+ and Si in the Sediments and the Body Walls of Sea Cucumbers in the Tropical Seagrass Meadows

Abstract
:1. Introduction
2. Materials and Methods
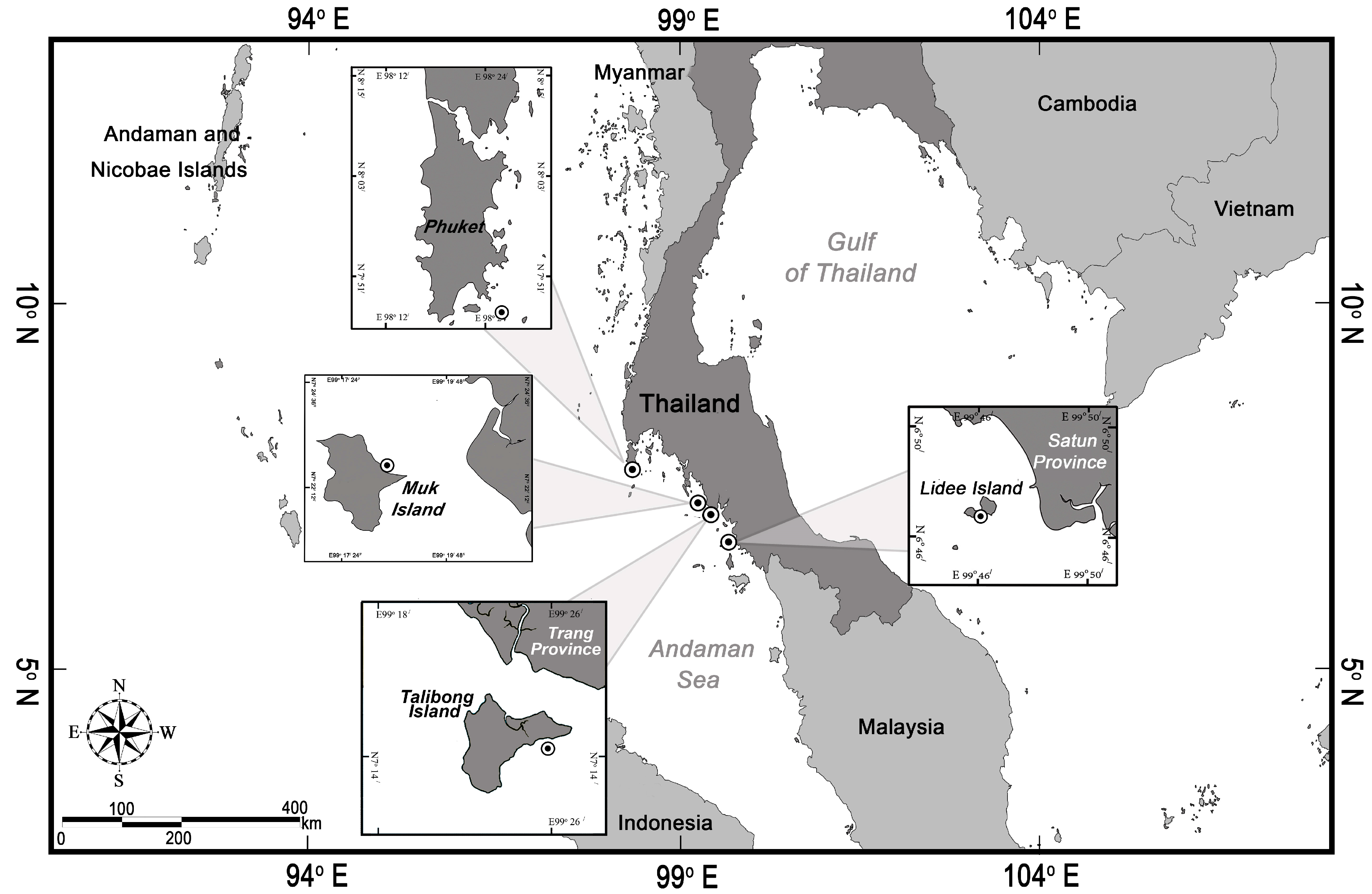
2.1. Study Sites
2.2. Sampling and Processing of Samples
2.3. Statistical Analysis
3. Results
3.1. Comparison among Species and Locations
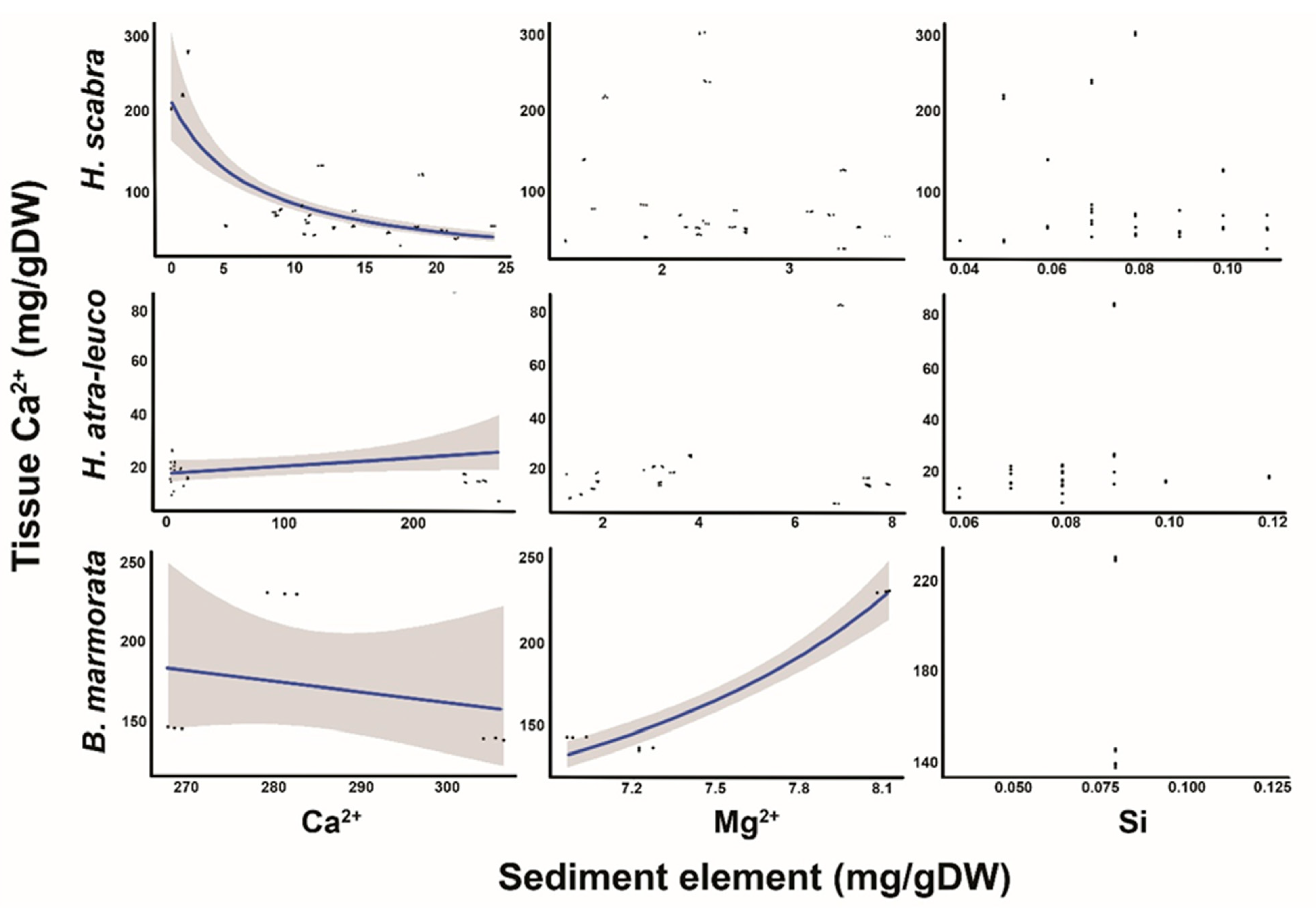
3.2. Relationship between Tissue Ca2+ and Sediment Ca2+, Mg2+ and Si
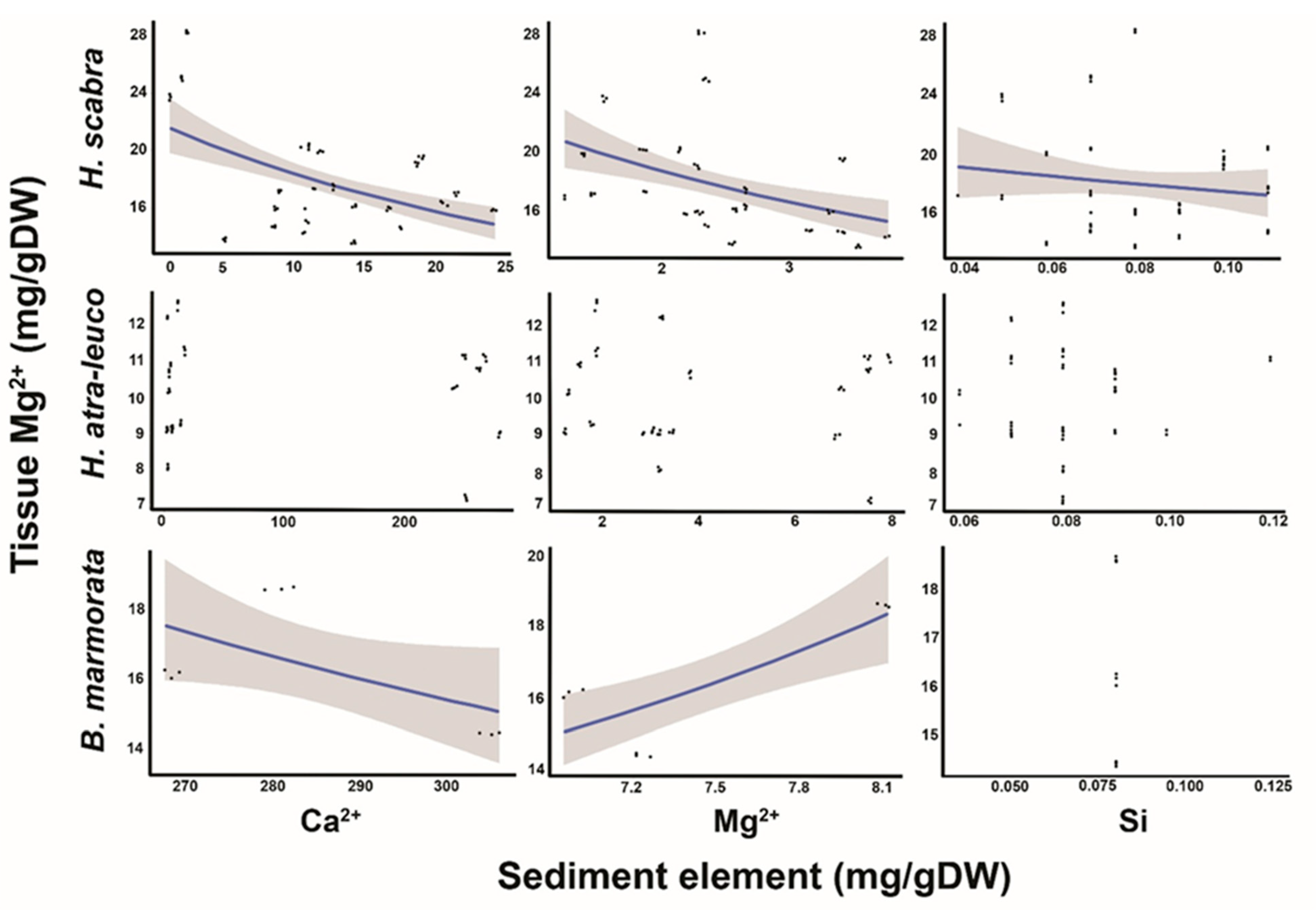
3.3. Relationship between Tissue Mg2+ and Sediment Ca2+, Mg2+ and Si
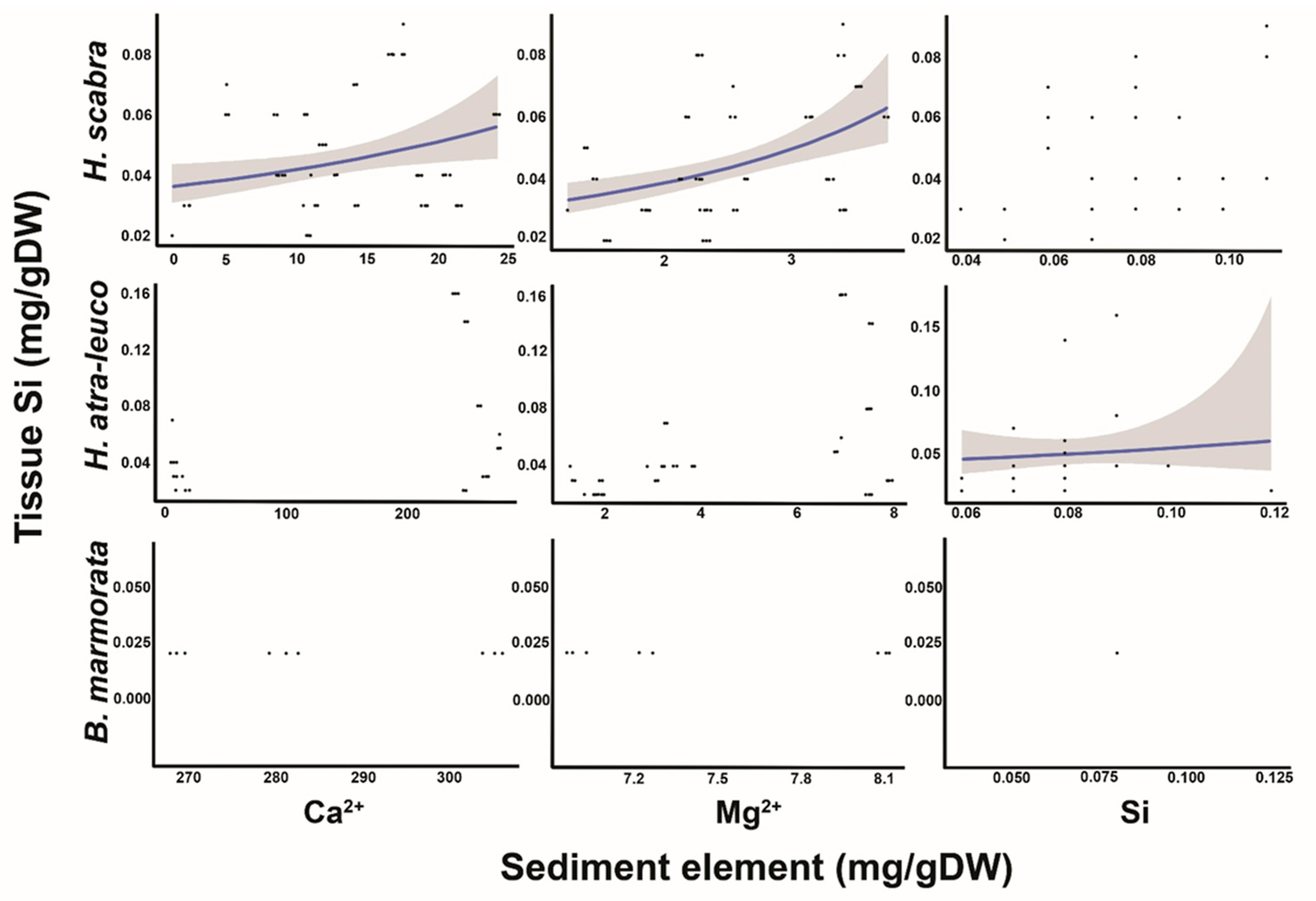
3.4. Relationship between Tissue Si and Sediment Ca2+, Mg2+ and Si
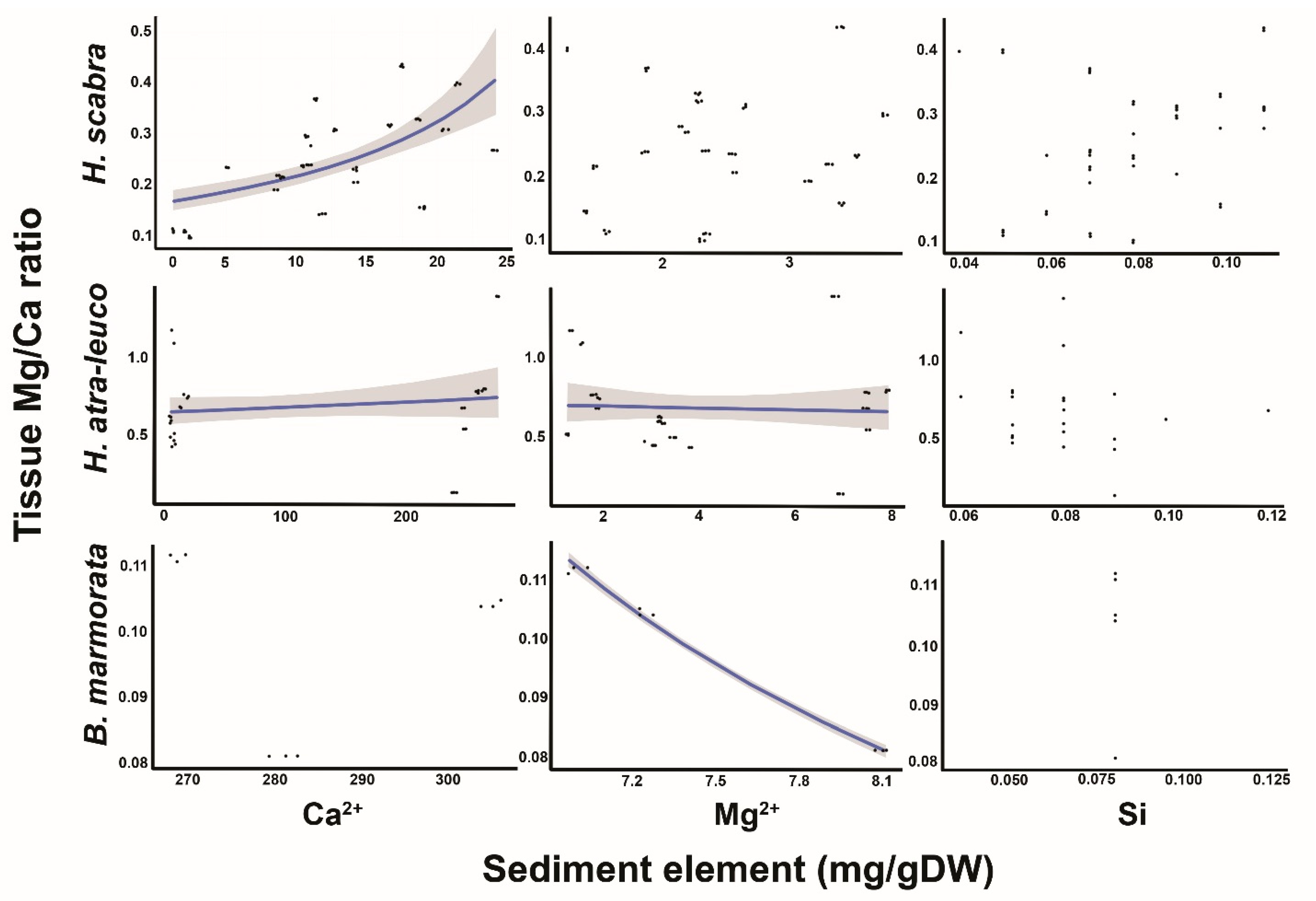
3.5. Relationship between Tissue Mg/Ca and Sediment Ca2+, Mg2+, Si and Mg/Ca
4. Discussion
4.1. Comparison among Species and Locations
4.2. Relationship between Tissue Ca2+ and Sediment Ca2+, Mg2+ and Si
4.3. Relationship between Tissue Mg2+ and Sediment Ca2+, Mg2+ and Si
4.4. Relationship between Tissue Si and Sediment Ca2+, Mg2+ and Si
4.5. Relationship between Tissue Mg:Ca Ratio and Sediment Ca2+, Mg2+ and Si
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guerrero, A.; Rodríguez Forero, A. Histological characterization of skin and radial bodies of two species of genus Isostichopus (Echinodermata: Holothuroidea). Egypt. J. Aquat. Res. 2018, 44, 155–161. [Google Scholar] [CrossRef]
- Takemae, N.; Nakaya, F.; Motokawa, T. Low oxygen consumption and high body content of catch connective tissue contribute to low metabolic rate of sea cucumbers. Biol. Bull. 2009, 216, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Candia Carnevali, M.D. Regenerative response and endocrine disrupters in crinoid echinoderms: An old experimental model, a new ecotoxicological test. Prog. Mol. Subcell. Biol. 2005, 39, 167–200. [Google Scholar] [CrossRef] [PubMed]
- Inoue, M.; Tamori, M.; Motokawa, T. Innervation of holothurian body wall muscle: Inhibitory effects and localization of 5-HT. Zool. Sci. 2002, 19, 1217–1222. [Google Scholar] [CrossRef] [Green Version]
- Matsuno, A.; Motokawa, T. Evidence for calcium translocation in catch connective tissue of the sea cucumber Stichopus chloronotus. Cell Tissue Res. 1992, 267, 307–312. [Google Scholar] [CrossRef]
- Hayashi, Y.; Motokawa, T. Effects of ionic environment on viscosity of catch connective tissue in Holothurian body wall. J. Exp. Biol. 1986, 125, 71–84. [Google Scholar] [CrossRef]
- Ries, J.B. Review: Geological and experimental evidence for secular variation in seawater Mg/Ca (calcite-aragonite seas) and its effects on marine biological calcification. Biogeosciences 2010, 7, 2795–2849. [Google Scholar] [CrossRef] [Green Version]
- Marin, F.; Bundeleva, I.; Takeuchi, T.; Immel, F.; Medakovic, D. Organic matrices in metazoan calcium carbonate skeletons: Composition, functions, evolution. J. Struct. Biol. 2016, 196, 98–106. [Google Scholar] [CrossRef]
- Clark, M.S. Molecular mechanisms of biomineralization in marine invertebrates. J. Exp. Biol. 2020, 223, jeb206961. [Google Scholar] [CrossRef]
- Sun, N.; Wu, H.; Du, M.; Tang, Y.; Liu, H.; Fu, Y.; Zhu, B. Food protein-derived calcium chelating peptides: A review. Trends Food Sci. Technol. 2006, 58, 140–148. [Google Scholar] [CrossRef]
- Vrieling, E.G.; Beelen, T.P.M.; Sun, Q.; Hazelaar, S.; van Santen, R.A.; Gieskes, W.W.C. Ultrasmall, Small, and Wide Angle X-ray Scattering analysis of diatom biosilica: Interspecific differences in fractal properties. J. Mater. Chem. 2004, 14, 1970–1975. [Google Scholar] [CrossRef]
- Rondanelli, M.; Faliva, M.A.; Peroni, G.; Gasparri, C.; Perna, S.; Riva, A.; Petrangolini, G.; Tartara, A. Silicon: A neglected micronutrient essential for bone health. Exp. Biol. Med. 2021, 246, 1500–1511. [Google Scholar] [CrossRef] [PubMed]
- Jugdaohsingh, R. Silicon and bone health. J. Nutr. Health Aging 2007, 11, 99–110, PMCID:PMC2658806. [Google Scholar] [PubMed]
- Zioła-Frankowska, A.; Kubaszewski, Ł.; Dąbrowski, M.; Frankowski, M. Interrelationship between silicon, aluminum, and elements associated with tissue metabolism and degenerative processes in degenerated human intervertebral disc tissue. Environ. Sci. Pollut. Res. 2017, 24, 19777–19784. [Google Scholar] [CrossRef] [Green Version]
- Lowenstam, H.A.; Rossman, G.R. Amorphous, hydrous, ferric phosphatic dermal granules in Molpadia (Holothuroidea): Physical and chemical characterization and ecologic implications of the bioinorganic fraction. Chem. Geol. 1975, 15, 15–51. [Google Scholar] [CrossRef]
- Barzkar, N.; Fariman, G.A.; Taheri, A. Proximate composition and mineral contents in the body wall of two species of sea cucumber from Oman Sea. Environ. Sci. Pollut. Res. 2017, 24, 18907–18911. [Google Scholar] [CrossRef]
- Iglikowska, A.; Najorka, J.; Voronkov, A.; Chełchowski, M.; Kuklinski, P. Variability in magnesium content in Arctic echinoderm skeletons. Mar. Environ. Res. 2017, 129, 207–218. [Google Scholar] [CrossRef]
- Thomson, T. Silicon. In Methods in Geochemistry and Geophysics; Easton, A.J., Ed.; Elsevier Publishing Company: Amsterdam, The Netherlands, 1972; Volume 6, pp. 71–94. [Google Scholar] [CrossRef]
- Kumar, S.; Natalio, F.; Elbaum, R. Protein-driven biomineralization: Comparing silica formation in grass silica cells to other biomineralization processes. J. Struct. Biol. 2021, 213, 107665. [Google Scholar] [CrossRef]
- Conley, D.J.; Chelske, C.L.; Stoermer, E.F. Modification of the biogeochemical cycle of silica with eutrophication. Mar. Ecol. Prog. Ser. 1993, 101, 179–192. [Google Scholar] [CrossRef]
- Tambutté, S.; Holcomb, M.; Ferrier-Pagès, C.; Reynaud, S.; Tambutté, É.; Zoccola, D.; Allemand, D. Coral biomineralization: From the gene to the environment. J. Exp. Mar. Biol. Ecol. 2011, 408, 58–78. [Google Scholar] [CrossRef]
- Smith, A.B. Echinoderm Larvae and Phylogeny. Annu. Rev. Ecol. Syst. 1997, 28, 219–241. [Google Scholar] [CrossRef]
- Hammond, L.S. An analysis of grain size modification in biogenic carbonate sediments by deposit-feeding holothurians and echinoids. Limnol. Oceanogr. 1981, 26, 898–906. [Google Scholar] [CrossRef]
- Schneider, K.; Silverman, J.; Kravitz, B.; Rivlin, T.; Schneider-Mor, A.; Barbosa, S.; Byrne, M.; Caldeira, K. Inorganic carbon turnover caused by digestion of carbonate sands and metabolic activity of holothurians. Estuar. Coast. Shelf Sci. 2013, 133, 217–223. [Google Scholar] [CrossRef]
- Floren, A.S.; Hayashizaki, K.I.; Tuntiprapas, P.; Prathep, A. Contributions of seagrasses and other sources to sea cucumber diets in a tropical seagrass ecosystem. Chiang Mai J. Sci. 2021, 48, 1259–1270. [Google Scholar]
- Yuan, X.; McCoy, S.J.; Du, Y.; Widdicombe, S.; Hall-Spencer, J.M. Physiological and behavioral plasticity of the sea cucumber Holothuria forskali (Echinodermata, Holothuroidea) to acidified seawater. Front. Physiol. 2018, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Mayer, L.M.; Weston, D.P.; Bock, M.J.; Jumars, P.A. Inhibition of digestive enzyme activities by copper in the guts of various marine benthic invertebrates. Environ. Toxicol. Chem. 2002, 21, 1243–1248. [Google Scholar] [CrossRef]
- Dickson, J.A.D. Fossil echinoderms as monitor of the Mg/Ca ratio of Phanerozoic oceans. Science 2002, 298, 1222–1224. [Google Scholar] [CrossRef]
- Stanley, S.M. Influence of seawater chemistry on biomineralization throughout phanerozoic time: Paleontological and experimental evidence. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2006, 232, 214–236. [Google Scholar] [CrossRef]
- Asnaghi, V.; Mangialajo, L.; Gattuso, J.-P.; Francour, P.; Privitera, D.; Chiantore, M. Effects of ocean acidification and diet on thickness and carbonate elemental composition of the test of juvenile sea urchins. Mar. Environ. Res. 2014, 93, 78–84. [Google Scholar] [CrossRef]
- Kołbuk, D.; Dubois, P.; Stolarski, J.; Gorzelak, P. Effects of seawater chemistry (Mg2+/Ca2+ ratio) and diet on the skeletal Mg/Ca ratio in the common sea urchin Paracentrotus lividus. Mar. Environ. Res. 2019, 145, 22–26. [Google Scholar] [CrossRef]
- Ricart, A.M.; Dalmau, A.; Perez, M.; Romero, J. Effects of landscape configuration on the exchange of materials in seagrass ecosystems. Mar. Ecol. Prog. Ser. 2015, 532, 89–100. [Google Scholar] [CrossRef]
- Belbachir, N.E.; Lepoint, G.; Mezali, K. Comparison of isotopic niches of four sea cucumbers species (Holothuroidea: Echinodermata) inhabiting two seagrass meadows in the southwestern Mediterranean Sea (Mostaganem, Algeria). Belgian J. Zool. 2019, 149, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Khogkhao, C.; Hayashizaki, K.I.; Tuntiprapas, P.; Prathep, A. Changes in seagrass communities along the runoff gradient of the Trang river, Thailand. ScienceAsia 2017, 43, 339–346. [Google Scholar] [CrossRef]
- Rattanachot, E.; Prathep, A. Temporal variation in growth and reproduction of Enhalus acoroides (L. f.) Royle in a monospecific meadow in Haad Chao Mai National Park, Trang Temporal variation in growth and reproduction of Enhalus acoroides (L. f.) Royle in a monospecific meadow. Bot. Mar. 2015, 54, 201–207. [Google Scholar] [CrossRef]
- Rattanachot, E. Effect of Shoot Density on Growth, Recruitment and Reproduction of Enhalus acoroides (L.f.) Royle at Haad Chao Mai National Park, Trang Province, Thailand. Master’s Thesis, Prince of Songkla University, Songkla, Thailand, 2008. [Google Scholar]
- Floren, A.; Hayashizaki, K.I.; Putchakarn, S.; Tuntiprapas, P.; Prathep, A. A Review of Factors Influencing the Seagrass-Sea Cucumber Association in Tropical Seagrass Meadows. Front. Mar. Sci. 2021, 8, 696134. [Google Scholar] [CrossRef]
- Warnau, M.; Dutrieux, S.; Dúbois, P.; Ledent, G.; Rodriguez y Baena, A.M. Heavy metals in the sea cucumber Holothuria tubulosa (echinodermata) from the mediterranean Posidonia oceanica ecosystem: Body compartment, seasonal, geographical and bathymetric variations. Environ. Bioindic. 2006, 1, 268–285. [Google Scholar] [CrossRef]
- Turk Culha, S.; Dereli, H.; Karaduman, F.R.; Culha, M. Assessment of trace metal contamination in the sea cucumber (Holothuria tubulosa) and sediments from the Dardanelles Strait (Turkey). Environ. Sci. Pollut. Res. 2016, 23, 11584–11597. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org (accessed on 8 December 2021).
- Mangiafico, S. Rcompanion: Functions to Support Extension Education Program Evaluation. R Package Version 2.4.1. 2021. Available online: https://CRAN.R-project.org/package=rcompanion (accessed on 8 December 2021).
- Ogle, D.H.; Wheeler, P.; Dinno, A. FSA: Fisheries Stock Analysis. R Package Version 0.8.32. 2021. Available online: https://github.com/droglenc/FSA (accessed on 8 December 2021).
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics, 4th ed.; Springer: New York, NY, USA, 2002; ISBN 0-387-95457-0. [Google Scholar]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Duarte, C.M. Submerged aquatic vegetation in relation to different nutrient regimes. Ophelia 1995, 41, 87–112. [Google Scholar] [CrossRef]
- Ries, J.B. Effect of ambient Mg/Ca ratio on Mg fractionation in calcareous marine invertebrates: A record of the oceanic Mg/Ca ratio over the Phanerozoic. Geology 2004, 32, 981–984. [Google Scholar] [CrossRef]
- Ries, J.B. Aragonite production in calcite seas: Effect of seawater Mg/Ca ratio on the calcification and growth of the calcareous alga Penicillus capitatus. Paleobiology 2005, 31, 445–458. [Google Scholar] [CrossRef]
- Stanley, S.M.; Ries, J.B.; Hardie, L.A. Seawater chemistry, coccolithophore population growth, and the origin of Cretaceous chalk. Geology 2005, 33, 593–596. [Google Scholar] [CrossRef] [Green Version]
- Simkiss, K.; Wilbur, K.M. Biomineralization: Cell Biology and Mineral Deposition, 1st ed.; Academic Press: San Diego, CA, USA, 1989. [Google Scholar]
- Chan, N.C.S.; Connolly, S.R. Sensitivity of coral calcification to ocean acidification: A meta-analysis. Glob. Change Biol. 2013, 19, 282–290. [Google Scholar] [CrossRef]
- McClintock, J.B.; Amsler, M.O.; Angus, R.A.; Challener, R.C.; Schram, J.B.; Amsler, C.D.; Mah, C.L.; Cuce, J.; Baker, B.J. The Mg-calcite composition of Antarctic echinoderms: Important implications for predicting the impacts of ocean acidification. J. Geol. 2011, 119, 457–466. [Google Scholar] [CrossRef]
- Hermans, J.; Borremans, C.; Willenz, P.; André, L.; Dubois, P. Temperature, salinity and growth rate dependences of Mg/Ca and Sr/Ca ratios of the skeleton of the sea urchin Paracentrotus lividus (Lamarck): An experimental approach. Mar. Biol. 2010, 157, 1293–1300. [Google Scholar] [CrossRef]
- Carafoli, E. Intracellular calcium homeostasis. Ann. Rev. Biochem. 1987, 56, 395–433. [Google Scholar] [CrossRef]
- Yeagle, P. The Membrane of Cells, 3rd ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 335–378. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Element (mg/g DW) | Sample | Talibong | Muk | Phuket | Lidee | Differences among Species | ||
|---|---|---|---|---|---|---|---|---|
| Hs (n = 24) | Hl (n = 24) | Hs (n = 45) | Ha (n = 15) | Bm (n = 9) | Ha (n = 18) | Kruskal-Wallis (Chi-Squared) | ||
| Ca2+ | Tissue | 129.35 ± 18.44 | 18.83 ± 0.71 | 69.20 ± 4.20 | 12.79 ± 0.95 | 170.89 ± 14.78 | 24.67 ± 6.51 | 92.24 ** |
| Sediment | 6.62 ± 0.90 | 5.56 ± 0.32 | 15.28 ± 0.68 | 11.62 ± 1.33 | 285.14 ± 5.42 | 258.29 ± 2.97 | 64.24 ** | |
| Mg2+ | Tissue | 18.59 ± 1.16 | 9.54 ± 0.25 | 17.41 ± 0.27 | 10.84 ± 0.29 | 16.38 ± 0.61 | 9.90 ± 0.34 | 99.60 ** |
| Sediment | 2.82 ± 0.15 | 3.00 ± 0.15 | 2.25 ± 0.10 | 1.68 ± 0.06 | 7.44 ± 0.17 | 7.36 ± 0.09 | 28.64 ** | |
| Si | Tissue | 0.048 ± 0.004 | 0.043 ± 0.003 | 0.042 ± 0.003 | 0.025 ± 0.001 | 0.019 ± 0.000 | 0.081 ± 0.013 | 22.59 ** |
| Sediment | 0.074 ± 0.003 | 0.080 ± 0.002 | 0.084 ± 0.003 | 0.071 ± 0.002 | 0.081 ± 0.001 | 0.089 ± 0.004 | 0.49 ns | |
| Mg/Ca | Tissue | 0.187 ± 0.014 | 0.516 ± 0.015 | 0.281 ± 0.012 | 0.899 ± 0.055 | 0.099 ± 0.005 | 0.723 ± 0.093 | 92.69 ** |
| Sediment | 0.644 ± 0.076 | 0.596 ± 0.048 | 0.155 ± 0.007 | 0.170 ± 0.016 | 0.026 ± 0.001 | 0.029 ± 0.000 | 69.80 ** | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Floren, A.; Hayashizaki, K.-i.; Tuntiprapas, P.; Prathep, A. Modeling of the Elements Ca2+, Mg2+ and Si in the Sediments and the Body Walls of Sea Cucumbers in the Tropical Seagrass Meadows. Diversity 2023, 15, 146. https://doi.org/10.3390/d15020146
Floren A, Hayashizaki K-i, Tuntiprapas P, Prathep A. Modeling of the Elements Ca2+, Mg2+ and Si in the Sediments and the Body Walls of Sea Cucumbers in the Tropical Seagrass Meadows. Diversity. 2023; 15(2):146. https://doi.org/10.3390/d15020146
Chicago/Turabian StyleFloren, Adonis, Ken-ichi Hayashizaki, Piyalap Tuntiprapas, and Anchana Prathep. 2023. "Modeling of the Elements Ca2+, Mg2+ and Si in the Sediments and the Body Walls of Sea Cucumbers in the Tropical Seagrass Meadows" Diversity 15, no. 2: 146. https://doi.org/10.3390/d15020146
APA StyleFloren, A., Hayashizaki, K. -i., Tuntiprapas, P., & Prathep, A. (2023). Modeling of the Elements Ca2+, Mg2+ and Si in the Sediments and the Body Walls of Sea Cucumbers in the Tropical Seagrass Meadows. Diversity, 15(2), 146. https://doi.org/10.3390/d15020146