
Host Migration and Size Do Not Influence the Prevalence of Most Chewing Lice (Phthiraptera: Amblycera and Ischnocera) on Shorebirds (Aves: Charadriiformes) across the World

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection and Identification of Lice
2.2. Literature Survey
2.3. Data Analysis
3. Results
3.1. Louse Identification
3.2. Literature Survey
3.3. Host Migration and Prevalence of Lice
4. Discussion
4.1. Flyways
4.2. Host Body Size
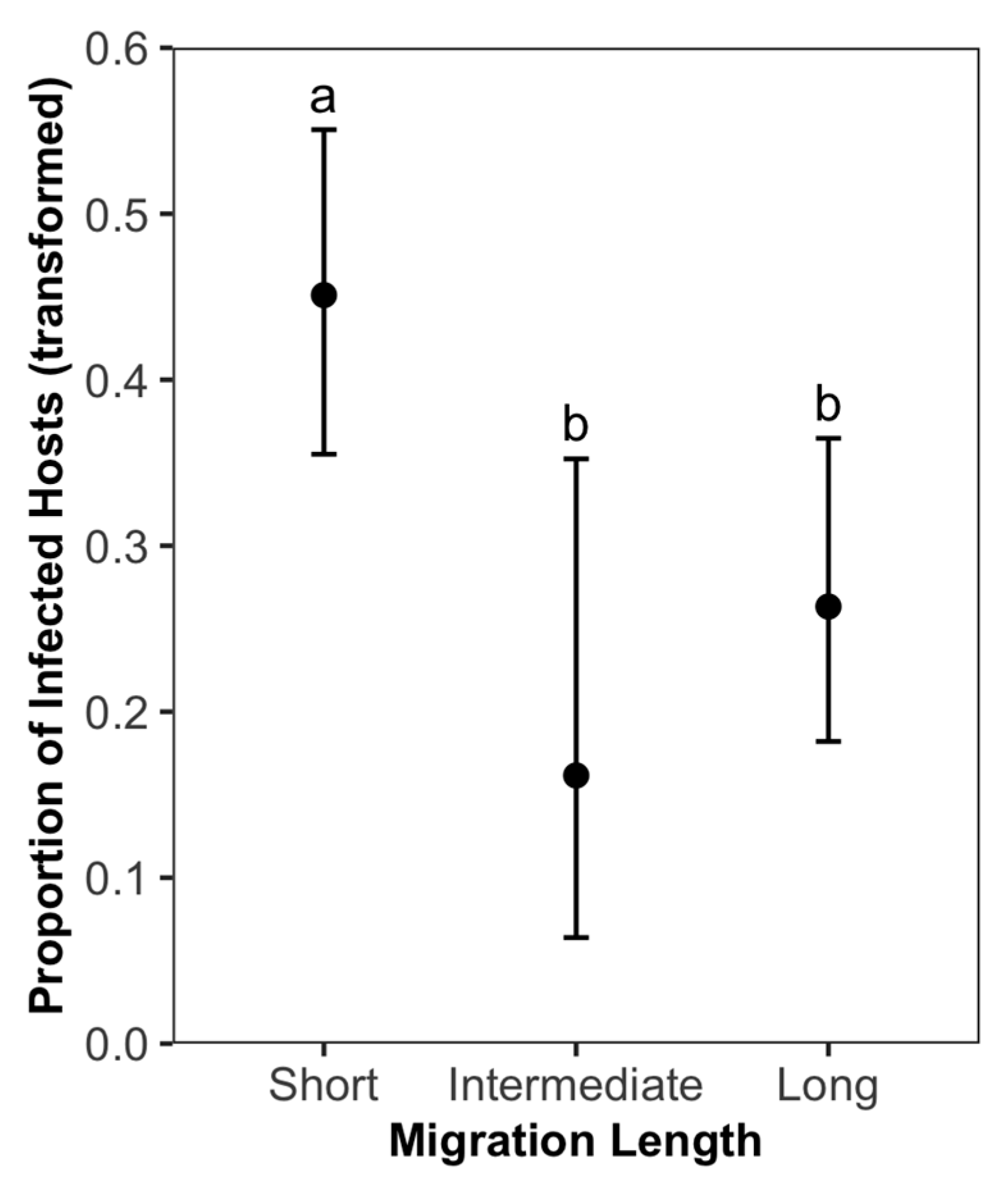
4.3. Migration Length
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gill, R.E., Jr.; Tibbitts, T.L.; Douglas, D.C.; Handel, C.M.; Molcahy, D.M.; Gottschalck, J.C.; Warnock, N.; McCaffery, B.J.; Battley, P.F.; Piersma, T. Extreme endurance flights by landbirds crossing the Pacific Ocean: Ecological corridor rather than barrier? Proc. R. Soc. B 2009, 276, 447–457. [Google Scholar] [CrossRef]
- Egevang, C.; Stenhouse, I.J.; Philips, R.A.; Petersen, A.; Fox, J.W.; Silk, J.R.D. Tracking of Arctic terns Sterna paradisaea reveals longest animal migration. Proc. Natl. Acad. Sci. USA 2010, 107, 2078–2081. [Google Scholar] [CrossRef] [PubMed]
- Scott, G.R.; Egginton, S.; Richards, J.G.; Milson, W.K. Evolution of muscle phenotype for extreme high altitude flight in the bar-headed goose. Proc. R. Soc. B 2009, 276, 3645–3653. [Google Scholar] [CrossRef] [PubMed]
- Newton, I. The Migration Ecology of Birds; Academic Press: London, UK, 2008; pp. viii + 976. [Google Scholar]
- Jehl, J.R.; Henry, A.E.; Ellis, H.I. Optimizing migration in a reluctant and inefficient flier: The Eared Grebe. In Avian Migration; Berthold, P., Gwinner, E., Sonnenschein, E., Eds.; Springer-Verlag: Berlin, Germany, 2003; pp. XVI + 610. [Google Scholar]
- Munro, U. Life-history and ecophysiological adaptations to migration in Australian birds. In Avian Migration; Berthold, P., Gwinner, E., Sonnenschein, E., Eds.; Springer-Verlag: Berlin, Germany, 2003; pp. XVI + 610. [Google Scholar]
- Carlisle, J.D.; Kaltenecker, G.S.; Swanson, D.L. Molt strategies and age differences in migration timing among autumn landbird migrants in Southwestern Idaho. Auk 2005, 122, 1070–1085. [Google Scholar] [CrossRef]
- Barta, Z.; McNamara, J.M.; Houston, A.I.; Weber, T.P.; Hedenström, A.; Feró, O. Optimal moult strategies in migratory birds. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 363, 211–229. [Google Scholar] [CrossRef]
- Hall, K.S.; Tullberg, B.S. Phylogenetic analyzes of the diversity of moult strategies in Sylviidae in relation to migration. Evol. Ecol. 2004, 18, 85–105. [Google Scholar] [CrossRef]
- Winkler, H.; Leisler, B. On the ecomorphology of migrants. Ibis 1992, 134, 21–28. [Google Scholar] [CrossRef]
- Leisler, B.; Winkler, H. Morphological consequences of migration in passerines. In Avian Migration; Berthold, P., Gwinner, E., Sonnenschein, E., Eds.; Springer-Verlag: Berlin, Germany, 2003; pp. XVI + 610. [Google Scholar]
- Møller, A.P.; de Lope, F.; Saino, N. Parasitism, immunity, and arrival date in a migratory bird, the barn swallow. Ecology 2004, 85, 206–219. [Google Scholar] [CrossRef]
- Garvin, M.C.; Szell, C.C.; Moore, F.R. Blood parasites of Nearctic-Neotropical migrant passerine birds during spring trans-Gulf migration: Impact on host body condition. J. Parasitol. 2006, 92, 990–996. [Google Scholar] [CrossRef]
- Koprivnikar, J.; Leung, T.L.F. Flying with diverse passengers: Greater richness of parasitic nematodes in migratory birds. Oikos 2015, 124, 399–405. [Google Scholar] [CrossRef]
- Gutiérrez, J.S.; Piersma, T.; Thieltges, D.W. Micro- and macroparasites species richness in birds: The role of host life history and ecology. J. Anim. Ecol. 2019, 88, 1226–1239. [Google Scholar] [CrossRef] [PubMed]
- Altizer, S.; Bartel, R.; Han, B.A. Animal migration and infectious disease risk. Science 2011, 331, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Shaw, A.K.; Binning, S.A. Migratory recovery from infection as a selective pressure for the evolution of migration. Am. Nat. 2016, 187, 491–501. [Google Scholar] [CrossRef]
- Sychra, O.; Literák, I.; Podzemny, P.; Harmat, P.; Hrabák, R. Insect ectoparasites on wild birds in the Czech Republic during the pre-breeding season. Parasite 2011, 18, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Dietsch, T.V. Seasonal variation of infestation by ectoparasitic chigger mite larvae (Acarina: Trombiculidae) on resident and migratory birds in coffee agroecosystems of Chiapas, Mexico. J. Parasitol. 2005, 91, 1294–1303. [Google Scholar] [CrossRef]
- Poulin, R.; de Angeli Dutra, D. Animal migrations and parasitism: Reciprocal effects within a unified framework. Biol. Rev. 2021, 96, 1331–1348. [Google Scholar] [CrossRef]
- Figuerola, J. Ecological correlates of feather mite prevalence in passerines. J. Avian Biol. 2000, 31, 489–494. [Google Scholar] [CrossRef]
- Chu, X.; Dik, B.; Gustafsson, D.R.; Che, X.; Zhang, Q.; Zou, F. The influence of host body size and food guild on prevalence and mean intensity of chewing lice (Phthiraptera) on birds in southern China. J. Parasitol. 2019, 105, 334–344. [Google Scholar] [CrossRef]
- Gustafsson, D.R.; Lei, L.; Luo, K.; Chu, X.; Zhao, X.; Zhang, Q.; Zou, F. Chewing lice from high-altitude and migrating birds in Yunnan, China, with descriptions of two new species of Guimaraesiella. Med. Vet. Entomol. 2019, 33, 407–419. [Google Scholar] [CrossRef]
- Ferris, G.F. Some generic groups in the mallophagan family Menoponidae. Can. Entomol. 1916, 48, 301–311. [Google Scholar] [CrossRef]
- Bedford, G.A.H. Notes on Menoponidae (Mallophaga) with descriptions of new genera and species. Onderstepoort J. Vet. Sci. Anim. Ind. 1939, 12, 121–152. [Google Scholar]
- Clay, T.; Meinertzhagen, R. Three new genera of Mallophaga from Charadriiformes. Ann. Mag. Nat. Hist. 1939, 4, 450–454. [Google Scholar] [CrossRef]
- Timmermann, G. Saemundssonia nov. gen., ein neues Mallophagengenus, aufgestellt für Philopterus gonothorax (Giebel) und verwandte Arten. Zool. Anz 1936, 114, 97–100. [Google Scholar]
- Johnson, K.P.; Shreve, S.M.; Smith, V.S. Repeated adaptive divergence of microhabitat specialization in avian feather lice. BMC Biology 2012, 10, 52. [Google Scholar] [CrossRef]
- Villa, S.M.; Goodman, G.B.; Ruff, J.S.; Clayton, D.H. Does allopreening control avian ectoparasites? Biol. Lett. 2016, 12, 20160362. [Google Scholar] [CrossRef]
- Goodman, G.B.; Klingensmith, M.C.; Bush, S.E.; Clayton, D.H. The role of scratching in the control of ectoparasites on birds. Auk 2020, 137, ukaa010. [Google Scholar] [CrossRef]
- Price, R.D.; Leibovitz, L. A new species of Actornithophilus (Mallophaga: Menoponidae) from the know. Can. Entomol. 1969, 101, 997–999. [Google Scholar] [CrossRef]
- Taylor, A.L. Adventitious moult in red knot possibly caused by Actornithophilus (Mallophaga, Menoponidae). J. Field Ornithol. 1981, 52, 241. [Google Scholar]
- The eBird/Clements Checklist of Birds of the World: v2021. Available online: http://www.birds.cornell.edu/clementschecklist/download/ (accessed on 3 March 2022).
- Price, R.D.; Hellenthal, R.A.; Palma, R.L.; Johnson, K.P.; Clayton, D.H. The Chewing Lice: World Checklist and Biological Overview; Illinois Natural History Survey Special Publication: Champaign, IL, USA, 2003; pp. 24. x + 501. [Google Scholar]
- Gustafsson, D.R.; Olsson, U. Flyway homogenization or differentiation? Insights from the phylogeny of the sandpiper (Charadriiformes: Scolopacidae: Calidrinae) wing louse genus Lunaceps (Phthiraptera: Ischnocera). Int. J. Parasitol. 2012, 42, 93–102. [Google Scholar] [CrossRef]
- Eduardo, S.L. A new species of Saemundssonia and new records of Quadraceps species (Phthiraptera: Ischnocera: Philopteridae) found on some Philippine charadriiform birds. Philipp. J. Vet. Med. 2018, 55, 115–126. [Google Scholar]
- Clay, T. A key to the species of Actornithophilus Ferris with notes and descriptions of new species. Bull. Mus. Hist. Entomol. 1962, 11, 189–244. [Google Scholar]
- Clay, T. A key to the species of Austromenopon Bedford (Mallophaga) parasitic on the Charadriiformes. Proc. R. Entomol. Soc. Lond. 1959, 28, 157–168. [Google Scholar] [CrossRef]
- Timmermann, G. A revision of the genus Carduiceps Clay & Meinertzhagen, 1939 (Mallophaga). Ann. Mag. Nat. Hist. 1954, 7, 40–48. [Google Scholar]
- Gustafsson, D.R.; Olsson, U. The “very thankless task”: Revision of Lunaceps Clay & Meinertzhagen, 1939 (Insecta: Phthiraptera: Ischnocera: Philopteridae), with descriptions of six new species and one new subspecies. Zootaxa 2012, 3377, 1–85. [Google Scholar]
- Timmermann, G. Beiträge zur Kenntnis der Ektoparasitenfauna isländischer Säugetiere und Vögel. 2. Mitteilung.Das Mallophagengenus Koeniginirmus Eichler, 1940. Visindafel. Isl. 1949, 2, 83–88. [Google Scholar]
- Timmermann, G. Beiträge zur Kenntnis der Ektoparasitenfauna isländischer Säugetiere und Vögel. 4. Mitteilung. Die Gattung Quadraceps Clay & Meinertzhagen, 1939 und verwandte Genera Mallophagorum. Fauna Isl. Misc. Pap. Icel. Zool. 1950, 2, 1–8. [Google Scholar]
- Timmermann, G. Mallophagologische Kollektaneen, 2. Senckenberg. Biol. 1971, 52, 41–47. [Google Scholar]
- Hopkins, G.H.E.; Timmermann, G. A revision of the species of Quadraceps (Mallophaga) parasitic on Tringinae. Trans. R. Entomol. Soc. Lond. 1954, 105, 131–150. [Google Scholar] [CrossRef]
- Clay, T. Species of the genus Saemundssonia (Mallophaga) from the Sterninae. Am. Mus. Novit. 1949, 1409, 1–25. [Google Scholar]
- Timmermann, G. Beiträge zur Kenntnis der Ektoparasitenfauna isländischer Säugetiere und Vögel. 1. Mitteilung. Das Mallophagengenus Saemundssonia Timmermann, 1936. Visindafel. Isl. 1949, 2, 1–32. [Google Scholar]
- Ward, R.A. Biting lice of the genus Saemundssonia (Mallophaga: Philopteridae) occurring on terns. Proc. United States Natl. Mus. 1955, 105, 83–100. [Google Scholar] [CrossRef]
- Tavera, E.A.; Minaya, D.; Lopez, E.O.; Iannacone, J.; Lank, D.B. Chewing lice richness and occurrence in non-breeding shorebirds in Paracas, Perú. Wader Study 2019, 126, 190–199. [Google Scholar]
- Literák, I. Ektoparazité na ptácich (Passeriformes, Charadriiformes) protahujícich Moravskou bránou. Vet. Med. 1983, 28, 737–744. [Google Scholar]
- Billerman, S.M.; Keeney, B.K.; Rodewald, P.G.; Schulenberg, T.S. Birds of the World, Cornell Laboratory of Ornithology. 2022. Available online: https://birdsoftheworld.org/bow/home (accessed on 1 April 2022).
- Hayman, P. Shorebirds, an Identification Guide to the Waders of the World; Christopher Helm, A&C Black: London, UK, 1986; p. 412. [Google Scholar]
- Message, S.; Taylor, D. Waders of Europe, Asia and North America; Christopher Helm: London, UK, 2014; p. 230. [Google Scholar]
- Madge, S.; McGowan, P. Pheasants, Partridges & Grouse, Including Buttonquail, Sandgrouse and Allies; Christopher Helm: London, UK, 2002; p. 488. [Google Scholar]
- Olsen, K.M. Gulls of the World, a Photographic Guide; Christopher Helm: London, UK, 2018; p. 368. [Google Scholar]
- Menkhorst, P.; Rogers, D.; Clarke, R.; Davies, J.; Marsack, P.; Franklin, K. The Australian Bird Guide; Christopher Helm: London, UK, 2017; pp. x + 566. [Google Scholar]
- Svensson, L.; Mullarney, K.; Zetterström, D. Collins Bird Guide, 2nd ed.; HarperCollins: London, UK, 2009; p. 445. [Google Scholar]
- Smithson, M.; Verkuilen, J. A better lemon squeezer? Maximum-likelihood regression with beta distributed dependent variables. Psychol. Methods 2006, 11, 54–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Maechler, M.; Bolker, B.M. glmmTMB Balances Speed and Flexibility Among Packages for Zero-inflated Generalized Linear Mixed Modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef]
- RStudio Team. RStudio: Integrated Development Environment for R; RStudio, PBC: Boston, MA, USA, 2021; Available online: http://www.rstudio.com/ (accessed on 1 July 2022).
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level / Mixed) Regression Models. R Package Version 0.4.5. 2022. Available online: https://CRAN.R-project.org/package=DHARMa (accessed on 1 July 2022).
- Lüdecke, D. Ggeffects: Tidy Data Frames of Marginal Effects from Regression Models. J. Open Source Softw. 2018, 3, 772. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer-Verlag: New York, NY, USA, 2016. [Google Scholar]
- Lenth, R.V. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. R Package Version 1.7.4-1. 2022. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 1 July 2022).
- Gustafsson, D.R.; DiBlasi, E.; Olsson, U.; Najer, T.; Sychra, O.; Bush, S.E. Checklist and key to the lice (Insecta: Phthiraptera) of Sweden. Entomol. Tidskr. 2019, 139, 205–394. [Google Scholar]
- Murray, M.D.; Palma, R.L.; Pilgrim, R.L.C. Appendix I. Ectoparasites of Australian, New Zealand and Antarctic birds. In Handbook of Australian, New Zealand and Antarctic Birds. Raptors to Lapwings; Marchant, S., Higgins, P.J., Eds.; Oxford University Press: Oxford, UK, 1993; Volume 2, pp. 959–962. [Google Scholar]
- Murray, M.D.; Palma, R.L.; Pilgrim, R.L.C. Appendix II. Ectoparasites of Australian, New Zealand and Antarctic birds. In Handbook of Australian, New Zealand and Antarctic Birds. Boatbills to Starlings; Higgins, P.J., Peter, J., Cowling, S., Eds.; Oxford University Press: Oxford, UK, 2006; Volume 2, pp. 1964–1966. [Google Scholar]
- Bechet, I. Specii de Lunaceps (Insecta, Mallophaga) din Fauna României. Comunicări Zool. Soc. Ştiinţe Biol. București 1968, 6, 125–129. [Google Scholar]
- Blagoveshtchensky, D.I. Mallophaga of Tadzhikistan. Mag. Parasitol. L’institut Zool. L’academie Sci. L’ussr 1951, 13, 272–327. (In Russian) [Google Scholar]
- Mjöberg, E. Studien über Mallophagen und Anopluren. Ark. Zool. 1910, 6, 1–296. [Google Scholar] [CrossRef]
- Gustafsson, D.R.; Lei, L.; Chu, X.; Zhao, X.; Zou, F. Chewing lice (Phthiraptera) of the grey-headed lapwing Vanellus cinereus in China. Wader Study 2019, 126, 217–227. [Google Scholar]
- Schrank, F.d.P. Fauna Boica: Durchgedachte Geschichte der in Baiern Einheimischen und Zahmen Thiere. Part 1; Philipp Krüll: Landshut, Germany, 1803; Volume 3, pp. viii + 272. [Google Scholar]
- Timmermann, G. Studies on the Mallophaga from the collections of the British Museum (Nat. Hist.) London. I. A preliminary survey of the genus Lunaceps (Clay & Meinertzhagen), 1939. Ann. Mag. Nat. Hist. 1954, 7, 623–637. [Google Scholar]
- Denny, H. Monographia Anoplurorum Britanniae; Henry, G., Ed.; Bohn: London, UK, 1842; Volume 842, pp. xxvi + 262. [Google Scholar]
- Timmermann, G. Neue und wenig bekannte Kletterfederlinge von charadriiformes Wirten. Zool. Anz. 1954, 152, 163–177. [Google Scholar]
- Giebel, C.G. Die im zoologischen Museum der Universität Halle aufgestellten Epizoen nebst Beobachtungen über dieselben. Z. Die Gesammten Nat. Halle 1866, 28, 353–397. [Google Scholar]
- Waterston, J. On some ectoparasites in the South African Museum, Cape Town. Ann. S. Afr. Mus. 1914, 10, 271–324. [Google Scholar] [CrossRef]
- Burmeister, K.H.K. Mallophaga Nitzsch. In Handbuch der Entomologie. Zweiter Band. Besondere Entomologie. Zweite Abteilung. Lauskerfe. Gymnognatha. (Zweiter Ha¨lfte; vulgo Neuroptera); Theodor Christian Friedrich Enslin: Berlin, Germany, 1838; Volume 2, p. 293. [Google Scholar]
- Kellogg, V.L.; Kuwana, S.I. Papers from the Hopkins Stanford Galapagos Expedition, 1898–1899. X. Entomological results (8). Mallophaga from birds. J. Wash. Acad. Sci. 1902, 4, 457–499. [Google Scholar]
- Séguy, E. Pédiculines nouvelles de Madagascar. Mém. Inst. Sci. Madag. Ser. A 1949, 3, 165–170. [Google Scholar]
- Timmermann, G. Die Quadraceps-Arten (Mallophaga) der Kiebitze. Z. Parasitenkd. 1954, 16, 195–208. [Google Scholar] [CrossRef]
- Fabricius, O. Fauna Groenlandica: Systematice Sistens Animalia Groenlandiae Occidentalis Hactenus Indagata, Quoad Nomen Specificum, Triuiale, Vernaculumque: Synonyma Auctorum Plurium, Descriptionem, Locum, Victum, Generationem, Mores, Vsum, Capturamque Singuli, Prout Detegendi Occasio Fuit; Impensis Ioannis Gottlob Rothe: Copenhagen, Denmark, 1780; pp. xvi + 452. [Google Scholar]
- Martens, J.M. Zur Taxonomie der Gattung Saemundssonia Timmermann (Mallophaga: Ischnocera) auf Schnepfen (Scolopacinae) und Strandläufern (Eroliinae). Mitt. Hamb. Zool. Mus. Inst. 1974, 70, 119–163. [Google Scholar]
- Kellogg, V.L.; Chapman, B.L. Mallophaga from Birds of California. Occas. Pap. Calif. Acad. Sci. 1899, 6, 53–143. [Google Scholar]
- Piaget, E. Quelques pediculines Nouvelles. Tijdschr. Entomol. 1890, 33, 223–259. [Google Scholar]
- Touleshkov, K. Two new mallophag (sic) species on the species of the Larinae family. Izv. Zool. Inst. Comptes Rendus L’academe Bulg. Sci. 1959, 12, 557–559. [Google Scholar]
- Kellogg, V.L.; Chapman, B.L. Mallophaga from birds of the Hawaiian Islands. J. N. Y. Entomol. Soc. 1902, 10, 155–169. [Google Scholar]
- Grube, E. Beschreibung der auf A. Th. v. Middeondorff’s Sibirischer Reise gesammelten Parasiten. In Reise in den Aussersten Norden und Osten Sibiriens Während der Jahre 1843–1844: Zoologie, Band II, Theil 1; von Middendorff, T.A., Ed.; Buchdrückerei der Kaiserlischen Akademie der Wissenschaften: St. Petersburg, Russia, 1851; pp. 39 + 2 plates. [Google Scholar]
- Gustafsson, D.R.; Olsson, U. Unexpected distribution patterns of Carduiceps feather lice (Phthiraptera: Ischnocera: Philopteridae) on sandpipers (Aves: Charadriiformes: Scolopacidae). Syst. Entomol. 2017, 42, 509–522. [Google Scholar] [CrossRef]
- Bueter, C.; Weckstein, J.; Johnson, K.P.; Bates, J.M.; Gordon, C.E. Comparative phylogenetic histories of two louse genera found on Catharus thrushes and other birds. J. Parasitol. 2009, 95, 295–307. [Google Scholar] [CrossRef]
- Sweet, A.D.; Boyd, B.M.; Johnson, K.P. Cophylogenetic patterns and uncorrelated between two lineages of parasites on the same hosts. Biol. J. Linn. Soc. 2006, 118, 813–828. [Google Scholar] [CrossRef]
- Bush, S.E.; Harbison, C.W.; Slager, D.L.; Peterson, A.T.; Price, R.D.; Clayton, D.H. Geographic variation in the community structure of lice on western scrub-jays. J. Parasitol. 2009, 95, 10–13. [Google Scholar] [CrossRef] [PubMed]
- Meinertzhagen, R. A visit to Magadi, in Kenya, in March 1949. Ibis 1950, 92, 148–149. [Google Scholar]
- Campos, A.R.; Catry, P.; de Rojas, M.; Bearhop, S.; Ramos, J.; Newton, J. Winter habitat influences the number of feather mites of two species living on European robins Erithacus Rubecula. Ardeola 2011, 58, 103–111. [Google Scholar] [CrossRef]
- Gutiérrez, J.S.; Rakhimberdiev, E.; Persma, T.; Thieltges, D.W. Migration and parasitism: Habitat use, not migration distance, influences helminth species richness in charadriiform birds. J. Biogeogr. 2017, 44, 1137–1147. [Google Scholar] [CrossRef]
- Rózsa, L. Patterns in the abundance of avian lice (Phthiraptera: Amblycera, Ischnocera). J. Avian Biol. 1997, 28, 249–254. [Google Scholar] [CrossRef]
- Clayton, D.H.; Walther, B.A. Influence of host ecology and morphology on the diversity of Neotropical bird lice. Oikos 2001, 94, 455–467. [Google Scholar] [CrossRef]
- Galloway, T.D.; Lamb, R.J. Abundance of chewing lice (Phthiraptera: Amblycera and Ischnocera) increases with the body size of their host woodpeckers and sapsuckers (Aves: Piciformes: Picidae). Can. Entomol. 2017, 149, 473–481. [Google Scholar] [CrossRef]
- Durkin, E.S.; Luong, L.T.; Bird, J. Mechanisms underlying parasite infection: Influence of host body mass and age on chewing louse distribution among brown-headed cowbirds. Parasitol. Res. 2015, 114, 4169–4174. [Google Scholar] [CrossRef]
- Galloway, T.D.; Lamb, R.J.; Rempel, Z.D.L. Infestation parameters for chewing lice (Phthiraptera: Amblycera, Ischnocera) parasitising true thrushes (Aves: Passeriformes: Turdidae) in Manitoba, Canada. Can. Entomol. 2021, 151, 608–620. [Google Scholar] [CrossRef]
- Folmer, E.O.; Olff, H.; Piersma, T. How well do food distributions predict spatial distributions of shorebirds with different degrees of self-organization? J. Anim. Ecol. 2010, 79, 747–756. [Google Scholar] [CrossRef]
- Clayton, D.H.; Cotgreave, P. Relationship of bill morphology to grooming behavior in birds. Anim. Behav. 1994, 47, 195–201. [Google Scholar] [CrossRef]
- Ash, J.S. A study of the Mallophaga of birds with particular reference to their ecology. Ibis 1960, 102, 93–110. [Google Scholar] [CrossRef]
- Sychra, O.; Literák, I.; Podzemny, P.; Benedikt, V. Insect ectoparasites from wild passerine birds in the Czech Republic. Parasite 2008, 15, 599–604. [Google Scholar] [CrossRef] [Green Version]
- Clayton, D.H.; Moyer, B.R.; Bush, S.E.; Jones, T.G.; Gardiner, D.W.; Rhodes, B.B.; Goller, F. Adaptive significance of avian beak morphology for ectoparasite control. Proc. R. Soc. B 2005, 272, 811–817. [Google Scholar] [CrossRef]
- Gerritsen, A.F.C.; Meiboom, A. The role of touch in prey density estimation by Calidris alba. Neth. J. Zool. 1986, 36, 530–562. [Google Scholar] [CrossRef]
- Piersma, T.; van Aelst, R.; Kurk, K.; Berkhoudt, H.; Maas, L.R.M. A new pressure sensory mechanism for prey detection in birds: The use of principles of seabed dynamics? Proc. R. Soc. B 1998, 265, 1377–1383. [Google Scholar] [CrossRef]
- Nebel, S.; Jackson, D.L.; Elner, R.W. Functional association of bill morphology and foraging behavior in calidrid sandpipers. Anim. Biol. 2005, 55, 235–243. [Google Scholar] [CrossRef]
- Elner, R.W.; Beninger, P.G.; Jackson, D.L.; Potter, T.M. Evidence of a new feeding mode in western sandpiper (Calidris mauri) and dunlin (Calidris alpina) based on bill and tongue morphology and ultrastructure. Mar. Biol. 2005, 146, 1223–1234. [Google Scholar] [CrossRef]
- Delius, J.D. Preening and associated comfort behavior in birds. Ann. N. Y. Acad. Sci. 1988, 525, 40–55. [Google Scholar] [CrossRef]
- Cotgreave, P.; Clayton, D.H. Comparative analysis of time spent grooming by birds in relation to parasite load. Behaviour 1994, 131, 171–187. [Google Scholar]
- Burger, J.; Niles, L.; Clark, K.E. Importance of beach, mudflat and marsh habitats to migrant shorebirds on Delaware Bay. Biol. Conserv. 1997, 79, 283–292. [Google Scholar] [CrossRef]
- Beg, S.; Kumar, S.; Gupta, N.; Khan, V.; Saxena, A.K. Life table of Philopterus lahorensis (Phthiraptera: Mallophaga) infesting crows. Rev. Iber. Parasitol. 2005, 65, 63–66. [Google Scholar]
- Arya, G.; Ahman, A.; Bansal, N.; Rashmi, A.; Saxena, A.K. Population expansion of the common baya louse, Brueelia plocea (Lakshminarayana, 1968 (Phthiraptera: Ischnocera). Rev. Iero-Lat. Parasitol. 2009, 68, 192–195. [Google Scholar]
- Ahmad, A. Intrinsic rate of natural increase of an ischnoceran louse, Goniocotes jifrufti (Ansari, 1947) (Phthiraptera: Insecta). Am. J. Sci. 2020, 16, 63–67. [Google Scholar]
- Birkhead, T.R.; Hudson, P.J. Population parameters for the common guillemot Uria aalge. Ornis Scand. 1977, 8, 145–154. [Google Scholar] [CrossRef]
- Piatt, J.F.; McLagan, R.L. Common murre (Uria aalge) attendance patterns at Cape St. Mary’s, Newfoundland. Can. J. Zool. 1987, 65, 1530–1534. [Google Scholar] [CrossRef]
- Morris, R.D.; Chardine, J.W. The effects of ice cover over the colony site on reproductive activities of herring gulls. Can. J. Zool. 1985, 63, 607–611. [Google Scholar] [CrossRef]
- Hatch, S.A.; Hatch, M.A. Breeding seasons of oceanic birds in a subarctic colony. Can. J. Zool. 1990, 68, 1664–1679. [Google Scholar] [CrossRef]
- Merkel, B.; Descamps, S.; Yoccoz, N.G.; Danielsen, J.; Daunt, F.; Erikstad, K.E.; Ezhov, A.V.; Grémillet, D.; Gavrilo, M.; Lorentsen, S.-H.; et al. Earlier colony arrival but no trend in hatching timing in two congeneric seabirds (Uria spp.) across the North Atlantic. Biol. Lett. 2019, 15, 20190634. [Google Scholar] [CrossRef] [PubMed]
- Székely, T.; Reynolds, J.D. Evolutionary transitions in parental care in shorebirds. Proc. R. Soc. B 1995, 262, 57–64. [Google Scholar]
- McKinnon, L.; Schmaltz, L.; Aubry, Y.; Rochepault, Y.; Buidin, C.; Juillet, C. Female migration phenology and climate conditions explain juvenile red know (Calidris canutus rufa) counts during fall migration. Avian Conserv. Ecol. 2022, 17, 9. [Google Scholar] [CrossRef]
- Rózsa, L.; Rékási, J.; Reiczigel, J. Relationship of host coloniality to the population ecology of avian lice (Insecta: Phthiraptera). J. Anim. Ecol. 1996, 65, 242–248. [Google Scholar] [CrossRef]
- Gaston, A.J.; Hashimoto, Y.; Wilson, L. Post-breeding movements of ancient murrelet Synthliboramphus antiquus family groups, subsequent migration of adults and implications for management. PLoS ONE 2017, 12, e0171726. [Google Scholar] [CrossRef]
- Garcia, C.A.; Canaris, A.G. Metazoan parasites of Recurvirostra americana Gmelin (Aves), from southwestern Texas and Monte Vista National Wildlife Refuge, Colorado, with a checklist of helminth parasites hosted by this species in North America. Southwest. Nat. 1987, 32, 85–91. [Google Scholar] [CrossRef]
- Hinojos, J.G.; Canaris, A.G. Metazoan parasites of Himantopus mexicanus Muller (Aves) from southwestern Texas, with a checklist of helminth parasites from North America. J. Parasitol. 1988, 74, 326–331. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Host Family | Actornithophilus | Austromenopon | Other Amblycera | Quadraceps | Saemundssonia | Other Ischnocera |
|---|---|---|---|---|---|---|
| Alcidae | – | X | – | X | X | Craspedonirmus |
| Burhinidae | X | – | – | X | – | – |
| Charadriidae | X | X | – | X | X | – |
| Chionidae | X | – | – | X | X | – |
| Dromadidae | X | – | – | X | – | – |
| Glareolidae | X | X | Rediella | X | – | – |
| Haematopodidae | X | X | – | X | X | – |
| Ibidorhynchidae | X | – | – | X | – | – |
| Jacanidae | – | – | Pseudomenopon | – | X | Rallicola |
| Laridae | X | X | – | X | X | – |
| Pedionomidae | – | – | – | – | – | – |
| Pluvianellidae | – | – | – | – | – | – |
| Pluvianidae | – | – | – | X | – | – |
| Recurvirostridae | X | X | – | X | X | Cirrophthirius |
| Rostratulidae | X | – | Pseudomenopon | X | X | – |
| Scolopacidae | X | X | – | X | X | Carduiceps, Cummingsiella, Lunaceps, Rhynonirmus, Rotundiceps |
| Stercorariidae | – | X | – | X | X | Haffneria |
| Thinocoridae | – | – | – | X | – | – |
| Turnicidae | – | – | – | – | – | Turnicola |
| Host | NE | Louse | NI (P) | M | F | N | ID |
|---|---|---|---|---|---|---|---|
| Calidris alpina | 24 | Austromenopon sp. | 1 (4.1%) | 0 | 0 | 2 | Nymphs, unidentifiable |
| Carduiceps meinertzhageni | 8 (33.3%) | 11 | 11 | 4 | Timmermann (1954) [39] | ||
| Lunaceps schismatus | 18 (75.0%) | 53 | 89 | 44 | Gustafsson and Olsson (2012) [40] | ||
| Calidris canutus | 1 | Actornithophilus canuti | 1 (100%) | 0 | 1 | 0 | Price and Leibovitz (1969) [31] |
| Lunaceps drosti | 1 (100%) | 0 | 2 | 0 | Gustafsson and Olsson (2012) [40] | ||
| Calidris pygmeus | 2 | Lunaceps falcinellus | 2 (100%) | 0 | 7 | 0 | Gustafsson and Olsson (2012) [40] |
| Calidris ruficollis | 2 | Lunaceps falcinellus | 1 (50%) | 0 | 1 | 0 | Gustafsson and Olsson (2012) [40] |
| Calidris temminckii | 3 | --- | 0 | 0 | 0 | 0 | |
| Charadrius alexandrinus | 12 | Lunaceps falcinellus * | 2 (16.7%) | 1 | 1 | 0 | Gustafsson and Olsson (2012) [40] |
| Quadraceps macrocephalus | 9 (75%) | 22 | 15 | 1 | Specimen comparisons | ||
| Charadrius leschenaultii | 2 | Quadraceps ptyadis | 1 (50%) | 3 | 7 | 3 | Specimen comparisons |
| Charadrius mongolus | 3 | Quadraceps ptyadis | 3 (100%) | 15 | 20 | 4 | Specimen comparisons |
| Chlidonias hybrida | 1 | Quadraceps anagrapsus | 1 (100%) | 1 | 0 | 0 | Specimen comparisons |
| Gallinago gallinago | 13 | Quadraceps obscurus * | 1 (7.7%) | 1 | 0 | 0 | Specimen comparisons |
| Rhynonirmus scolopacis | 1 (7.7%) | 2 | 1 | 2 | Specimen comparisons | ||
| Himantopus himantopus | 1 | Actornithophilus himantopi | 1 (100%) | 5 | 7 | 1 | Clay (1962) [37] |
| Limosa lapponica | 1 | Lunaceps limosae | 1 (100%) | 2 | 5 | 0 | Gustafsson and Olsson (2012) [40] |
| Pluvialis squatarola | 1 | --- | 0 | 0 | 0 | 0 | |
| Saundersilarus saundersi | 1 | Saemundssonia lari | 1 (100%) | 1 | 1 | 0 | Specimen comparisons |
| Tringa stagnatilis | 10 | Actornithophilus totani | 5 (50%) | 1 | 0 | 10 | Clay (1962) [37] |
| Austromenopon sp. | 1 (10%) | 0 | 1 | 0 | Clay (1959) [38] | ||
| Carduiceps meinertzhageni * | 1 (10%) | 0 | 1 | 0 | Timmermann (1954) [39]; Specimen comparisons | ||
| Lunaceps sp. * | 1 (10%) | 0 | 1 | 0 | Unidentifiable specimen | ||
| Quadraceps obscurus | 9 (90%) | 17 | 25 | 9 | Hopkins and Timmermann (1954) [44]; Specimen comparisons | ||
| Tringa totanus | 5 | Actornithophilus totani | 1 (20%) | 1 | 2 | 8 | Clay (1962) [37] |
| Quadraceps obtusus | 4 (80%) | 3 | 7 | 0 | Hopkins and Timmermann (1954) [44]; Specimen comparisons |
| Louse Species | Host Species in China | Known Range in China | Source |
|---|---|---|---|
| Amblycera | |||
| Actornithophilus canuti Price & Leibovitz, 1969 [31] | Calidris canutus | Guangdong | New record in this study |
| Actornithophilus himantopi Blagoveshtchensky, 1951 [68] | Himantopus himantopus | Guangdong | New record in this study |
| Actornithophilus hoplopteri (Mjöberg, 1910) [69] | Vanellus cinereus | Yunnan | Gustafsson et al. [23,70] |
| Actornithophilus totani (Schrank, 1803) [71] | Tringa totanus | Guangdong | New record in this study |
| Ischnocera | |||
| Carduiceps meinertzhageni Timmermann, 1954 [39] | Calidris alpina | Guangdong | New record in this study |
| Lunaceps drosti Timmermann, 1954 [72] | Calidris canutus | Guangdong | New record in this study |
| Lunaceps falcinellus Timmermann, 1954 [72] | Calidris pygmaea | Guangdong | New record in this study |
| Lunaceps numenii phaeopi (Denny, 1842) [73] | Ibidorhynchus struthersii 1 | Tibet | Gustafsson and Olsson [40] |
| Lunaceps schismatus Gustafsson & Olsson, 2012 [40] | Calidris alpina | Guangdong | New record in this study |
| Quadraceps altoasiaticus Timmermann, 1954 [74] | Ibidorhynchus struthersii | Tibet | Timmermann [74] |
| Quadraceps anagrapsus (Nitzsch [in Giebel], 1866) [75] | Chlidonias hybridus | Guangdong | New record in this study |
| Quadraceps macrocephalus (Waterston, 1914) [76] | Charadrius alexandrinus | Guangdong | New record in this study |
| Quadraceps obscurus (Burmeister, 1838) [77] | Tringa stagnatilis | Guangdong | New record in this study |
| Quadraceps obtusus (Kellogg & Kuwana, 1902) [78] | Tringa totanus | Guangdong | New record in this study |
| Quadraceps ptyadis (Séguy, 1949) [79] | Charadrius leschenaltii, Charadrius mongolus | Guangdong | New record in this study |
| Quadraceps sinensis Timmermann, 1954 [80] | Vanellus cinereus | Yunnan | Gustafsson et al. [23,39] |
| Rhynonirmus scolopacis (Denny, 1842) [73] | Gallinago gallinago | Guangdong | New record in this study |
| Saemundssonia tringae (Fabricius, 1780) [81] | Calidris pygmaea | “China” | Martens [82] |
| Saemundssonia weidneri Martens, 1974 [82] | Gallinago megala | “Atchang, China” | Martens [82] |
| Saemundssonia lari (Fabricius, 1780) [81] | Saundersilarus saundersi | Guangdong | New record in this study |
| Host Species | NE | Louse Species | NI | #SP | ID |
|---|---|---|---|---|---|
| Calidris alba | 5 | Lunaceps actophilus (Kellogg & Chapman, 1899) [83] | 4 (80%) | 11 | Gustafsson and Olsson [40] |
| Charadrius leschenaultii | 14 | Quadraceps ptyadis Séguy, 1949 [79] | 11 (78.6%) | 108 | Séguy [79] |
| Charadrius veredus | 8 | Quadraceps assimilis (Piaget, 1890) [84] | 6 (75%) | 24 | Specimen comparisons |
| Gelochelidon nilotica | 5 | Quadraceps baliola (Blagoveshtchensky, 1951) [68] | 2 (40%) | 3 | Specimen comparisons |
| Saemundssonia gelochelidoni Touleshkov, 1959 [85] | 2 (40%) | 8 | Touleshkov [85] 1 | ||
| Tringa brevipes | 9 | Actornithophilus kilauensis (Kellogg & Chapman, 1902) [86] | 5 (55.6%) | 19 | Clay [37] |
| Quadraceps impar Hopkins & Timmermann, 1954 [44] 2 | 4 (44.4%) | 382 | Hopkins and Timmermann [44] | ||
| Tringa nebularia | 4 | Actornithophilus paludosus Clay, 1962 [37] | 2 (50%) | 2 | Clay [37] |
| Quadraceps similis (Giebel, 1866) [75] | 3 (75%) | 9 | Hopkins and Timmermann [44] | ||
| Tringa stagnatilis | 7 | Actornithophilus totani (Schrank, 1803) [71] | 2 (28.6%) | 3 | Clay [37] |
| Quadraceps obscurus (Burmeister, 1838) [77] | 6 (85.7%) | 50 | Hopkins and Timmermann [44] | ||
| Xenus cinereus | 12 | Carduiceps fulvofasciatus (Grube, 1851) [87] | 4 (33.3%) | 9 | Specimen comparisons |
| Response | χ2 | df | p-Value |
|---|---|---|---|
| Actornithophilus | 3.253 | 3 | 0.354 |
| Austromenopon | 1.769 | 3 | 0.622 |
| Quadraceps | 1.196 | 3 | 0.754 |
| Scolopacidae | 2.875 | 3 | 0.411 |
| Response | n | Predictor | Coefficient | Standard Error | z-Value | p-Value |
|---|---|---|---|---|---|---|
| Actornithophilus | 35 | Intercept [Flyway (African-Eurasian)] | −4.416 | 2.331 | −1.894 | 0.058 * |
| Flyway (Americas) | 0.291 | 0.304 | 0.960 | 0.337 | ||
| Flyway (East Asian-Australian) | 0.485 | 0.410 | 1.183 | 0.237 | ||
| Median Host Length | 0.604 | 0.425 | 1.419 | 0.156 | ||
| Austromenopon | 25 | Intercept [Flyway (African-Eurasian)] | −4.361 | 3.957 | −1.102 | 0.270 |
| Flyway (Americas) | −0.231 | 0.501 | −0.460 | 0.645 | ||
| Flyway (East Asian-Australian) | −0.552 | 0.634 | −0.870 | 0.384 | ||
| Median Host Length | 0.582 | 0.721 | 0.807 | 0.419 | ||
| Quadraceps | 29 | Intercept [Flyway (African-Eurasian)] | −3.174 | 5.758 | −0.551 | 0.581 |
| Flyway (Americas) | −0.700 | 0.713 | −0.982 | 0.326 | ||
| Flyway (East Asian-Australian) | 0.282 | 0.657 | 0.429 | 0.668 | ||
| Median Host Length | 0.630 | 1.046 | 0.602 | 0.547 | ||
| Scolopacidae | 125 | Intercept [Flyway (African-Eurasian)] | −3.385 | 2.135 | −1.585 | 0.113 |
| Flyway (Americas) | 0.057 | 0.233 | 0.246 | 0.805 | ||
| Flyway (East Asian-Australian) | 0.276 | 0.253 | 1.088 | 0.276 | ||
| Median Host Length | 0.509 | 0.394 | 1.292 | 0.196 |
| Response | χ2 | df | p-Value |
|---|---|---|---|
| Actornithophilus | 3.317 | 3 | 0.345 |
| Austromenopon | 1.525 | 3 | 0.677 |
| Quadraceps | 0.884 | 3 | 0.829 |
| Scolopacidae | 2.878 | 3 | 0.411 |
| Response | n | Predictor | Coefficient | Standard Error | z-Value | p-Value |
|---|---|---|---|---|---|---|
| Actornithophilus | 35 | Intercept [Flyway (African-Eurasian)] | −2.463 | 0.955 | −2.580 | 0.009 * |
| Flyway (Americas) | 0.344 | 0.308 | 1.116 | 0.264 | ||
| Flyway (East Asian-Australian) | 0.478 | 0.410 | 10167 | 0.243 | ||
| Median Host Weight | 0.294 | 0.204 | 1.441 | 0.150 | ||
| Austromenopon | 25 | Intercept [Flyway (African-Eurasian)] | −2.109 | 1.512 | −1.395 | 0.163 |
| Flyway (Americas) | −0.124 | 0.457 | −0.270 | 0.787 | ||
| Flyway (East Asian-Australian) | −0.542 | 0.644 | −0.842 | 0.400 | ||
| Median Host Weight | 0.203 | 0.325 | 0.625 | 0.532 | ||
| Quadraceps | 29 | Intercept [Flyway (African-Eurasian)] | 0.760 | 20237 | 0.340 | 0.734 |
| Flyway (Americas) | −0.448 | 0.671 | −0.668 | 0.504 | ||
| Flyway (East Asian-Australian) | 0.183 | 0.669 | 0.274 | 0.784 | ||
| Median Host Weight | −0.101 | 0.475 | −0.213 | 0.831 | ||
| Scolopacidae | 125 | Intercept [Flyway (African-Eurasian)] | −1.597 | 0.755 | −2.115 | 0.034 * |
| Flyway (Americas) | 0.077 | 0.239 | 0.324 | 0.746 | ||
| Flyway (East Asian-Australian) | 0.280 | 0.253 | 1.105 | 0.269 | ||
| Median Host Weight | 0.215 | 0.166 | 1.296 | 0.195 |
| Response | χ2 | df | p-Value |
|---|---|---|---|
| Actornithophilus | 4.136 | 3 | 0.182 |
| Austromenopon | 3.932 | 3 | 0.269 |
| Saemundssonia | 13.334 | 3 | 0.004 * |
| Quadraceps | 1.842 | 3 | 0.606 |
| Scolopacidae | 4.221 | 3 | 0.239 |
| Response | n | Predictor | Coefficient | Standard Error | z-Value | p-Value |
|---|---|---|---|---|---|---|
| Actornithophilus | 46 | Intercept [Migration Length (Intermediate)] | −6.228 | 3.534 | −1.762 | 0.078 |
| Migration Length (Long) | 0.509 | 0.595 | 0.856 | 0.392 | ||
| Migration Length (Short) | 0.899 | 0.675 | 1.287 | 0.198 | ||
| Median Host Length | 0.858 | 0.603 | 1.421 | 0.155 | ||
| Austromenopon | 49 | Intercept [Migration Length (Intermediate)] | −0.424 | 2.782 | −0.152 | 0.879 |
| Migration Length (Long) | 0.381 | 0.398 | 0.956 | 0.339 | ||
| Migration Length (Short) | 0.815 | 0.464 | 1.757 | 0.079 | ||
| Median Host Length | −0.178 | 0.508 | −0.350 | 0.726 | ||
| Saemundssonia | 52 | Intercept [Migration Length (Intermediate)] | −2.594 | 2.257 | −1.149 | 0.251 |
| Migration Length (Long) | 0.618 | 0.548 | 1.127 | 0.260 | ||
| Migration Length (Short) | 1.450 | 0.578 | 2.510 | 0.012 * | ||
| Median Host Length | 0.162 | 0.394 | 0.411 | 0.681 | ||
| Quadraceps | 55 | Intercept [Migration Length (Intermediate)] | −1.968 | 3.293 | −0.598 | 0.550 |
| Migration Length (Long) | 0.345 | 0.565 | 0.610 | 0.542 | ||
| Migration Length (Short) | 0.567 | 0.569 | 0.997 | 0.319 | ||
| Median Host Length | 0.311 | 0.555 | 0.559 | 0.576 | ||
| Scolopacidae | 133 | Intercept [Migration Length (Intermediate)] | −2.810 | 1.828 | −1.537 | 0.124 |
| Migration Length (Long) | 0.291 | 0.226 | 1.289 | 0.197 | ||
| Migration Length (Short) | −0.212 | 0.422 | −0.502 | 0.615 | ||
| Median Host Length | 0.366 | 0.344 | 1.064 | 0.287 |
| Response | χ2 | df | p-Value |
|---|---|---|---|
| Actornithophilus | 2.865 | 3 | 0.413 |
| Austromenopon | 5.065 | 3 | 0.167 |
| Saemundssonia | 15.032 | 3 | 0.002 * |
| Quadraceps | 2.442 | 3 | 0.486 |
| Scolopacidae | 4.085 | 3 | 0.252 |
| Response | n | Predictor | Coefficient | Standard Error | z Value | p-Value |
|---|---|---|---|---|---|---|
| Actornithophilus | 46 | Intercept [Migration Length (Intermediate)] | −1.789 | 1.491 | −1.99 | 0.230 |
| Migration Length (Long) | 0.317 | 0.591 | 0.536 | 0.592 | ||
| Migration Length (Short) | 0.985 | 0.678 | 1.453 | 0.146 | ||
| Median Host Weight | 0.092 | 0.245 | 0.348 | 0.728 | ||
| Austromenopon | 49 | Intercept [Migration Length (Intermediate)] | −2.471 | 1.020 | −2.423 | 0.015 * |
| Migration Length (Long) | 0.243 | 0.393 | 0.617 | 0.537 | ||
| Migration Length (Short) | 0.388 | 0.500 | 0.776 | 0.438 | ||
| Median Host Weight | 0.241 | 0.214 | 1.128 | 0.260 | ||
| Saemundssonia | 52 | Intercept [Migration Length (Intermediate)] | −2.655 | 0.875 | −3.036 | 0.002 * |
| Migration Length (Long) | 0.651 | 0.549 | 1.166 | 0.244 | ||
| Migration Length (Short) | 1.304 | 0.571 | 2.286 | 0.022 * | ||
| Median Host Weight | 0.193 | 0.139 | 1.391 | 0.164 | ||
| Quadraceps | 55 | Intercept [Migration Length (Intermediate)] | −1.330 | 1.327 | −1.002 | 0.316 |
| Migration Length (Long) | 0.364 | 0.552 | 0.659 | 0.510 | ||
| Migration Length (Short) | 0.409 | 0.594 | 0.688 | 0.510 | ||
| Median Host Weight | 0.228 | 0.236 | 0.964 | 0.335 | ||
| Scolopacidae | 133 | Intercept [Migration Length (Intermediate)] | −1.464 | 0.623 | −2.352 | 0.019 * |
| Migration Length (Long) | 0.282 | 0.228 | 10239 | 0.215 | ||
| Migration Length (Short) | −0.241 | 0.433 | −0.556 | 0.578 | ||
| Median Host Weight | 0.144 | 0.144 | 0.999 | 0.318 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grossi, A.A.; Lee, M.-B.; Tian, C.; Zou, F.; Choi, C.-Y.; Gustafsson, D.R. Host Migration and Size Do Not Influence the Prevalence of Most Chewing Lice (Phthiraptera: Amblycera and Ischnocera) on Shorebirds (Aves: Charadriiformes) across the World. Diversity 2023, 15, 200. https://doi.org/10.3390/d15020200
Grossi AA, Lee M-B, Tian C, Zou F, Choi C-Y, Gustafsson DR. Host Migration and Size Do Not Influence the Prevalence of Most Chewing Lice (Phthiraptera: Amblycera and Ischnocera) on Shorebirds (Aves: Charadriiformes) across the World. Diversity. 2023; 15(2):200. https://doi.org/10.3390/d15020200
Chicago/Turabian StyleGrossi, Alexandra Ashley, Myung-Bok Lee, Chunpo Tian, Fasheng Zou, Chi-Yeung Choi, and Daniel Roland Gustafsson. 2023. "Host Migration and Size Do Not Influence the Prevalence of Most Chewing Lice (Phthiraptera: Amblycera and Ischnocera) on Shorebirds (Aves: Charadriiformes) across the World" Diversity 15, no. 2: 200. https://doi.org/10.3390/d15020200
APA StyleGrossi, A. A., Lee, M. -B., Tian, C., Zou, F., Choi, C. -Y., & Gustafsson, D. R. (2023). Host Migration and Size Do Not Influence the Prevalence of Most Chewing Lice (Phthiraptera: Amblycera and Ischnocera) on Shorebirds (Aves: Charadriiformes) across the World. Diversity, 15(2), 200. https://doi.org/10.3390/d15020200