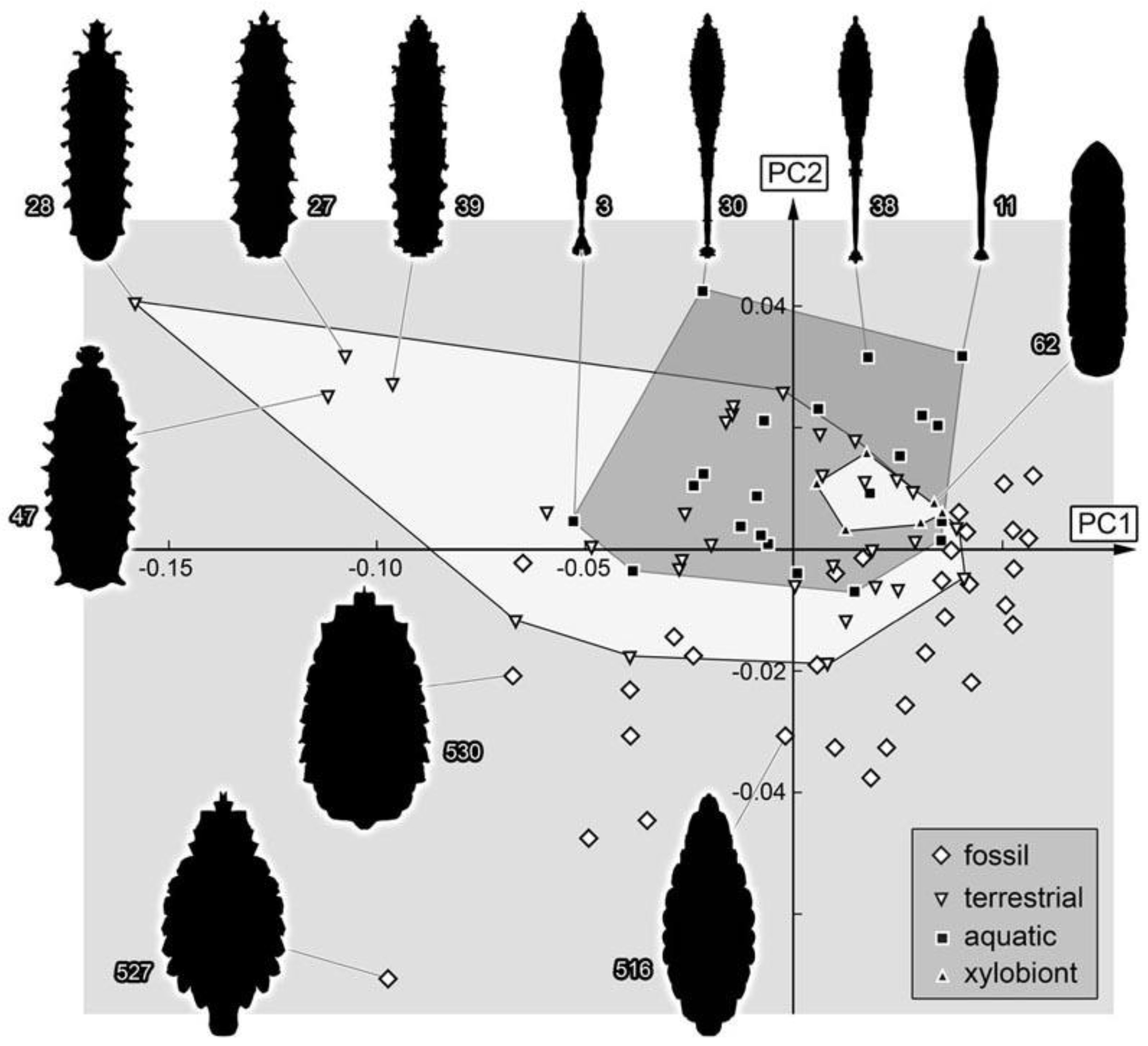
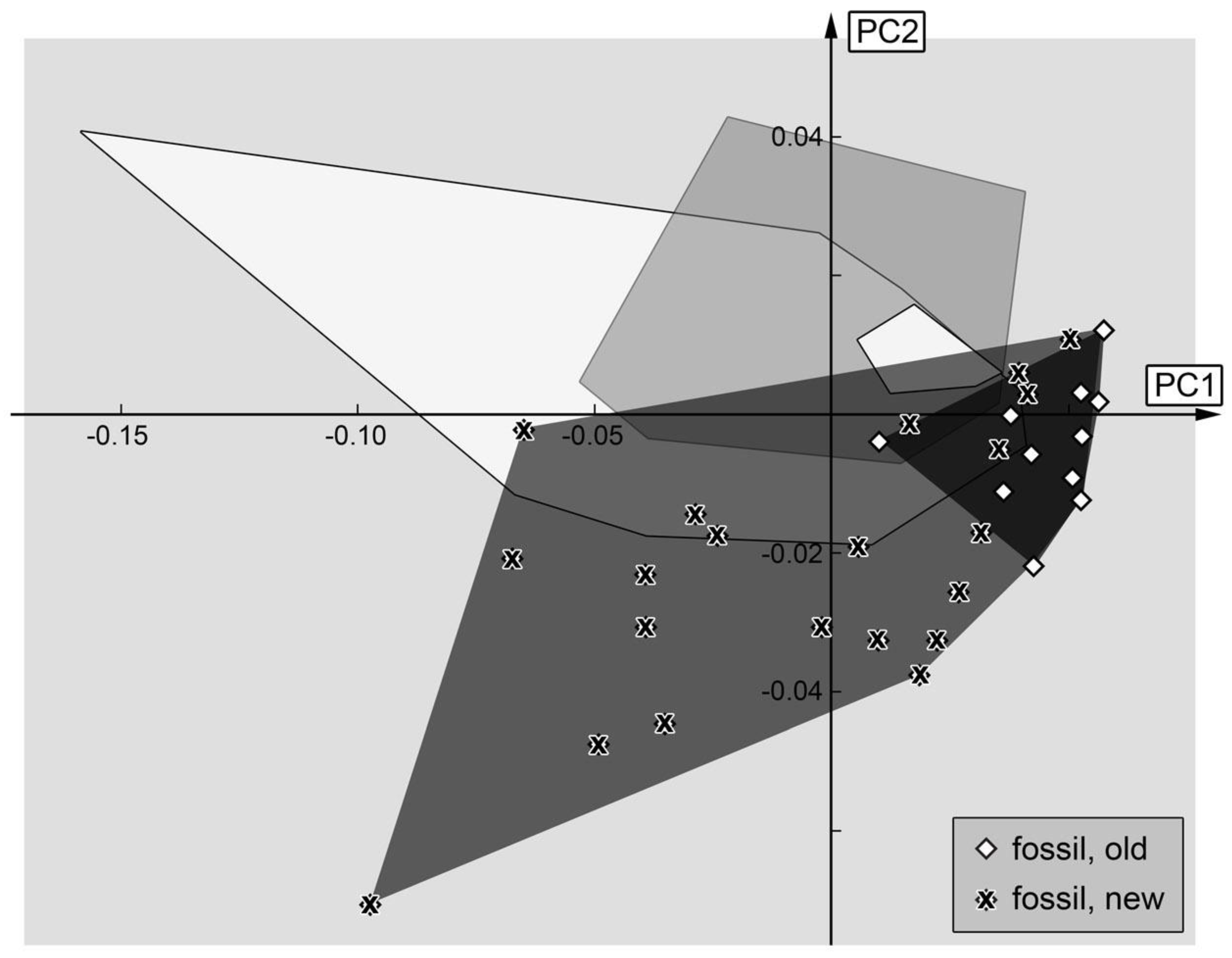
3.2.1. Material Examined
We examined 110 specimens: PED 0490, PED 0497, PED 0511, PED 0548, PED 0550, PED 0551, PED 0567-1, PED 0567-2, PED 0567-3, PED 0599-1, PED 0599-2, PED 0601, PED 0611-1, PED 0611-2, PED 0611-3, PED 0614, PED 0617, PED 0620, PED 0622, PED 0628, PED 0653, PED 0678, PED 0681, PED 0685, PED 0691, PED 0701, PED 0712, PED 0714, PED 0721, PED 0729, PED 0738, PED 0743, PED 0749, PED 0752, PED 0760, PED 0761-1, PED 0761-2, PED 0761-3, PED 0762, PED 0764, PED 0766, PED 0770, PED 0771, PED 0773-1, PED 0773-2, PED 0773-3, PED 0777, PED 0799, PED 0811-1, PED 0811-2, PED 0811-3, PED 0817, PED 0819-1, PED 0819-2, PED 0819-3, PED 0819-4, PED 0819-5, PED 0819-6, PED 0819-7, PED 0819-8, PED 0819-9, PED 0846, PED 0850, PED 0861, PED 0873-1, PED 0873-2, PED 0921, PED 0923, PED 0927, PED 0934, PED 0962, PED 0966, PED 0976-1, PED 0976-2, PED 0978, PED 0987, PED 0990, PED 1001, PED 1007-1, PED 1007-2, PED 1007-3, PED 1010, PED 1014, PED 1018, PED 1025, PED 1044, PED 1129, PED 1137, PED 1140, PED 1167, PED 1174, PED 1185, PED 1192, PED 1200-1, PED 1200-2, PED 1200-3, PED 1200-4, PED 1247, PED 1274, PED 1276, PED 1279, PED 1282, PED 1349, PED 1353, PED 1405, PED 1535, PED 1548-1, PED 1548-2, PED 1560, PED 1567.
3.2.3. Short Descriptions of Individual Specimens of Morphotype 1
Each specimen of morphotype 1 is briefly described, including the condition of the amber piece and preservation quality of the larvae. Visible details of every specimen and dimensions of the body and head capsule are provided. Although every specimen should have all the mentioned details available, if the detail was not visible, for example due to being covered by dirt particles or being only accessible from one side, it is not mentioned.
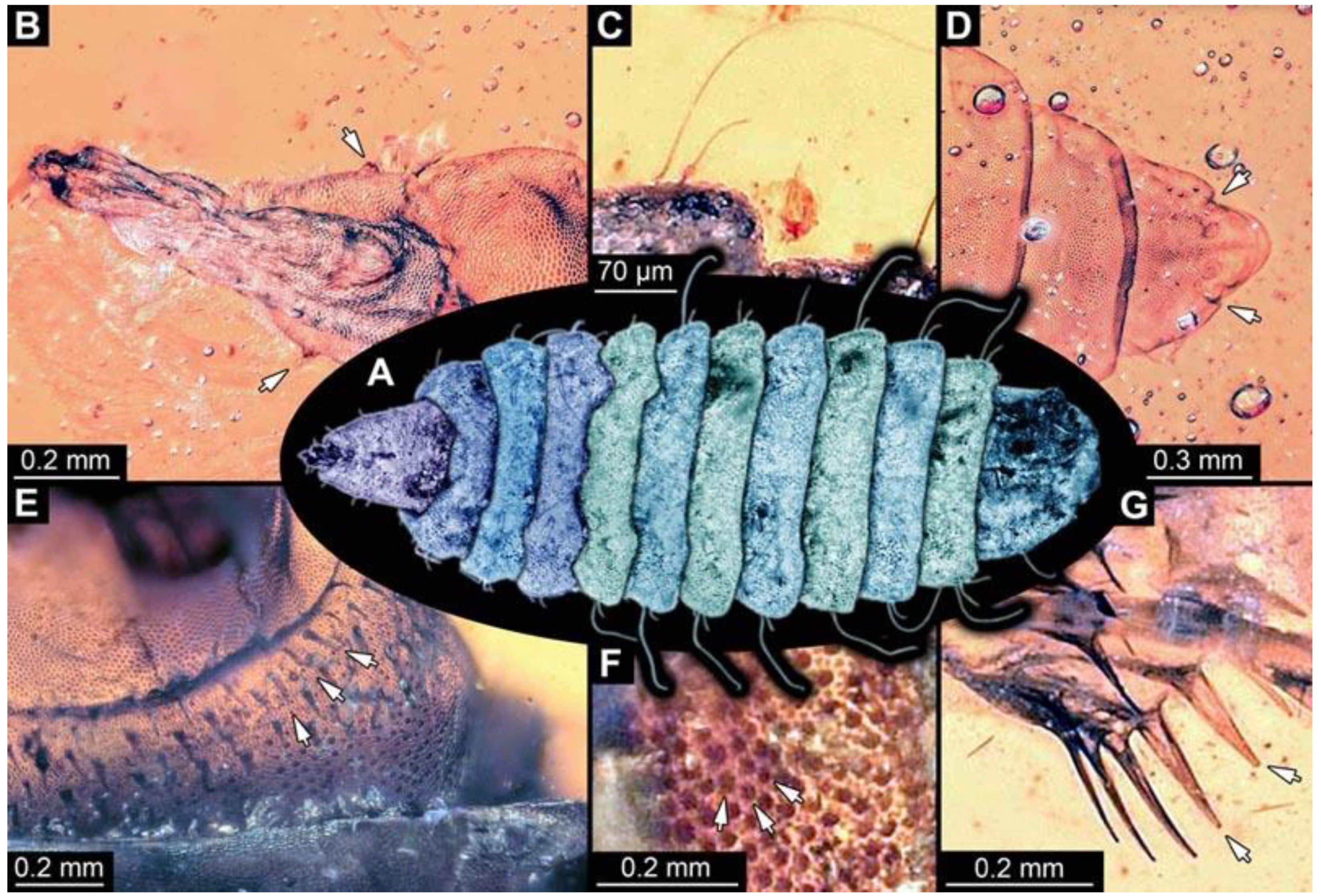
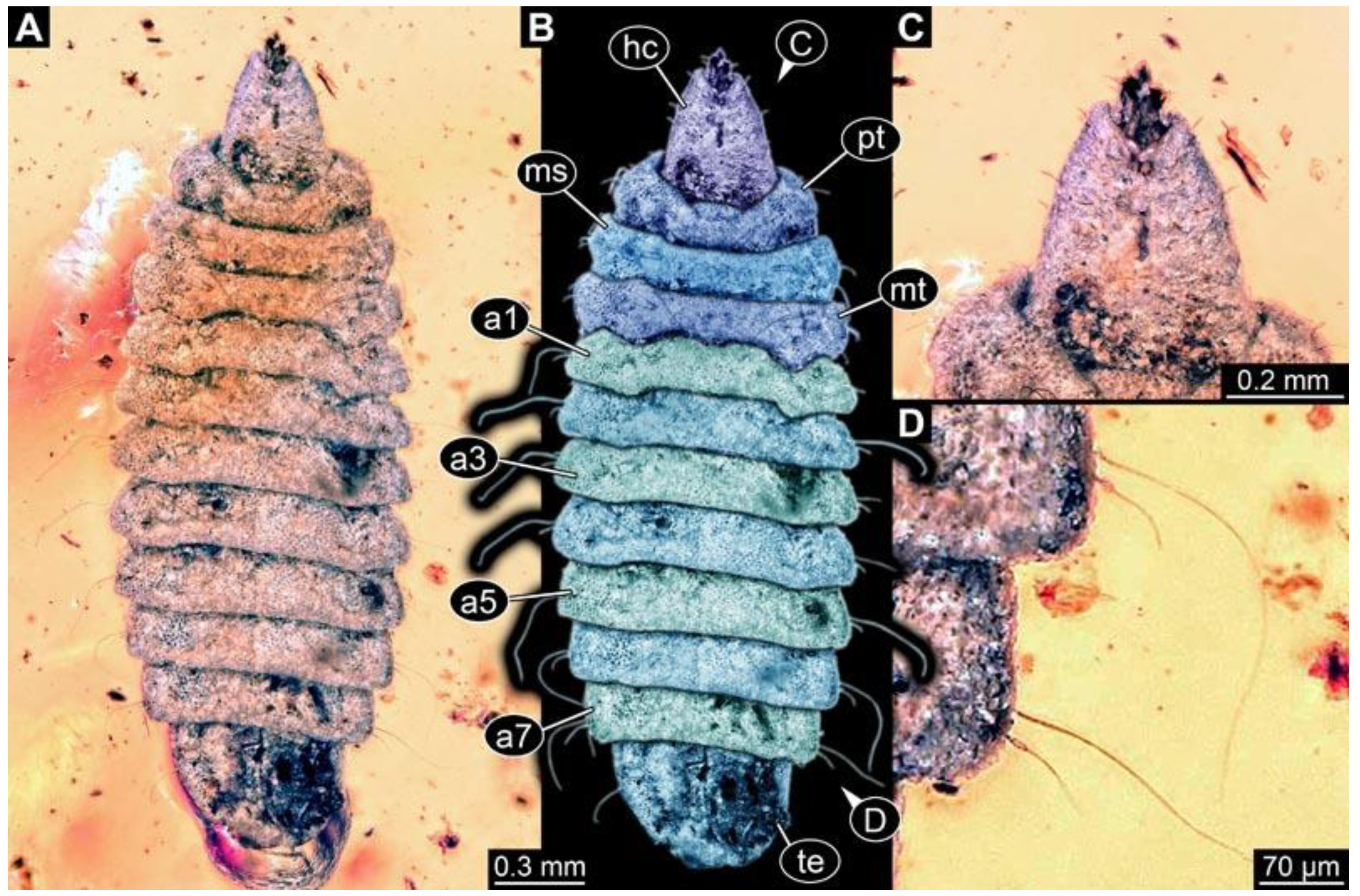
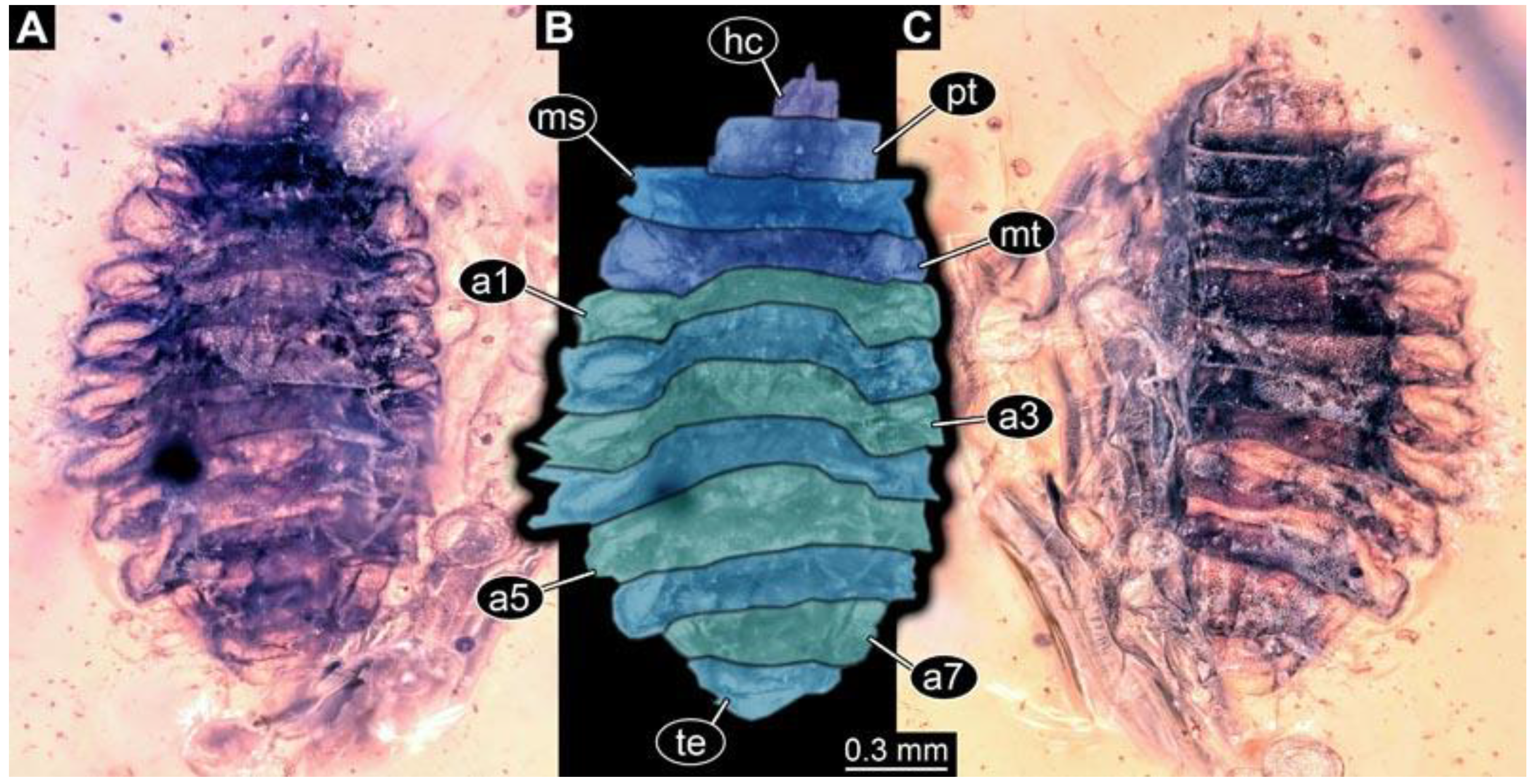
(1) PED 0617: Specimen strongly bent on anterior part (
Figure 2). Surface structure apparent. Head capsule well preserved; due to position, length could not be reliably measured. This also applies to total body length. Short spine-like setae on thorax segments well developed, longer ventral spine-like setae on abdomen segments well developed. Paired spiracles at trunk end well apparent. Dimensions: body width 1.58 mm, head capsule width 0.13 mm.
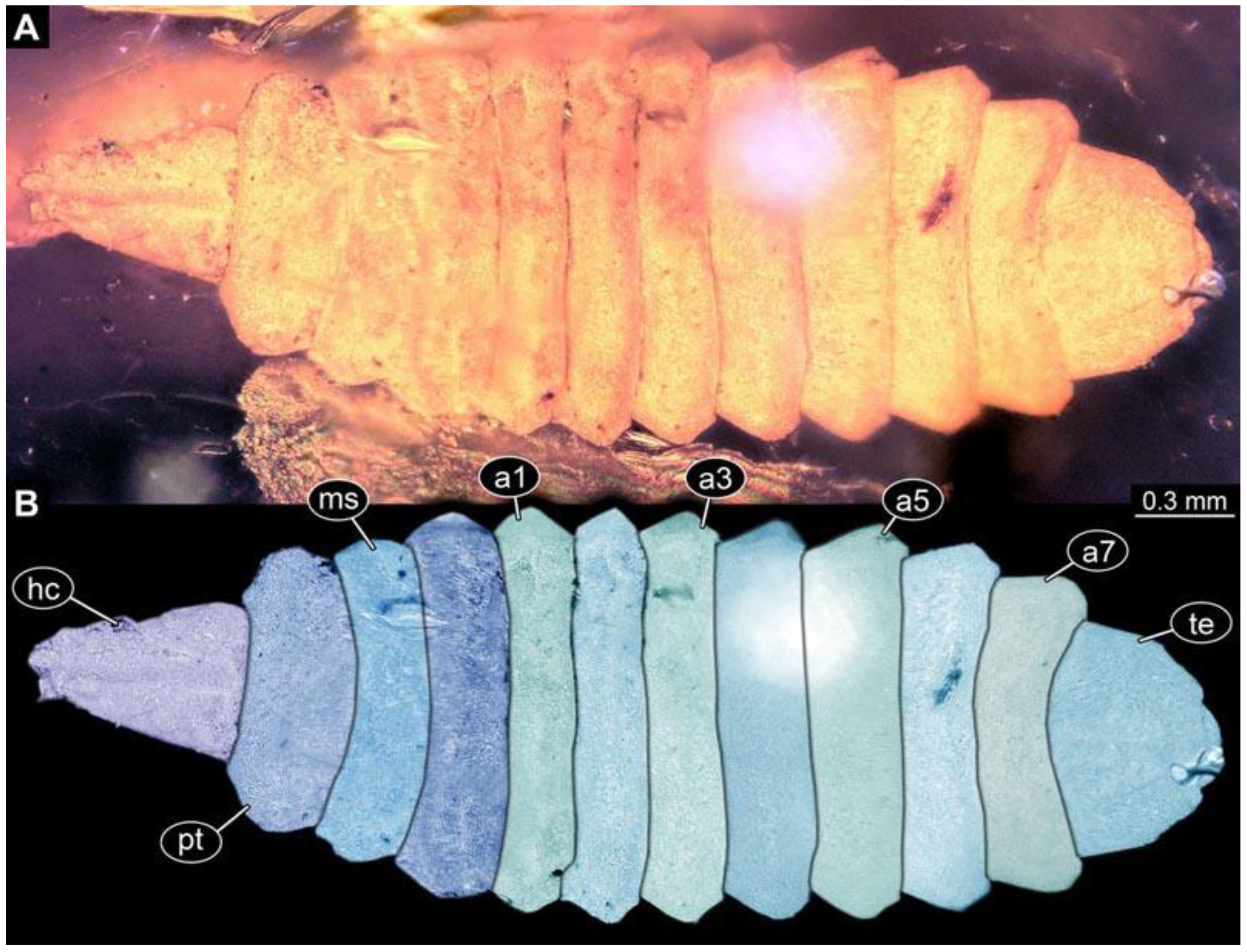
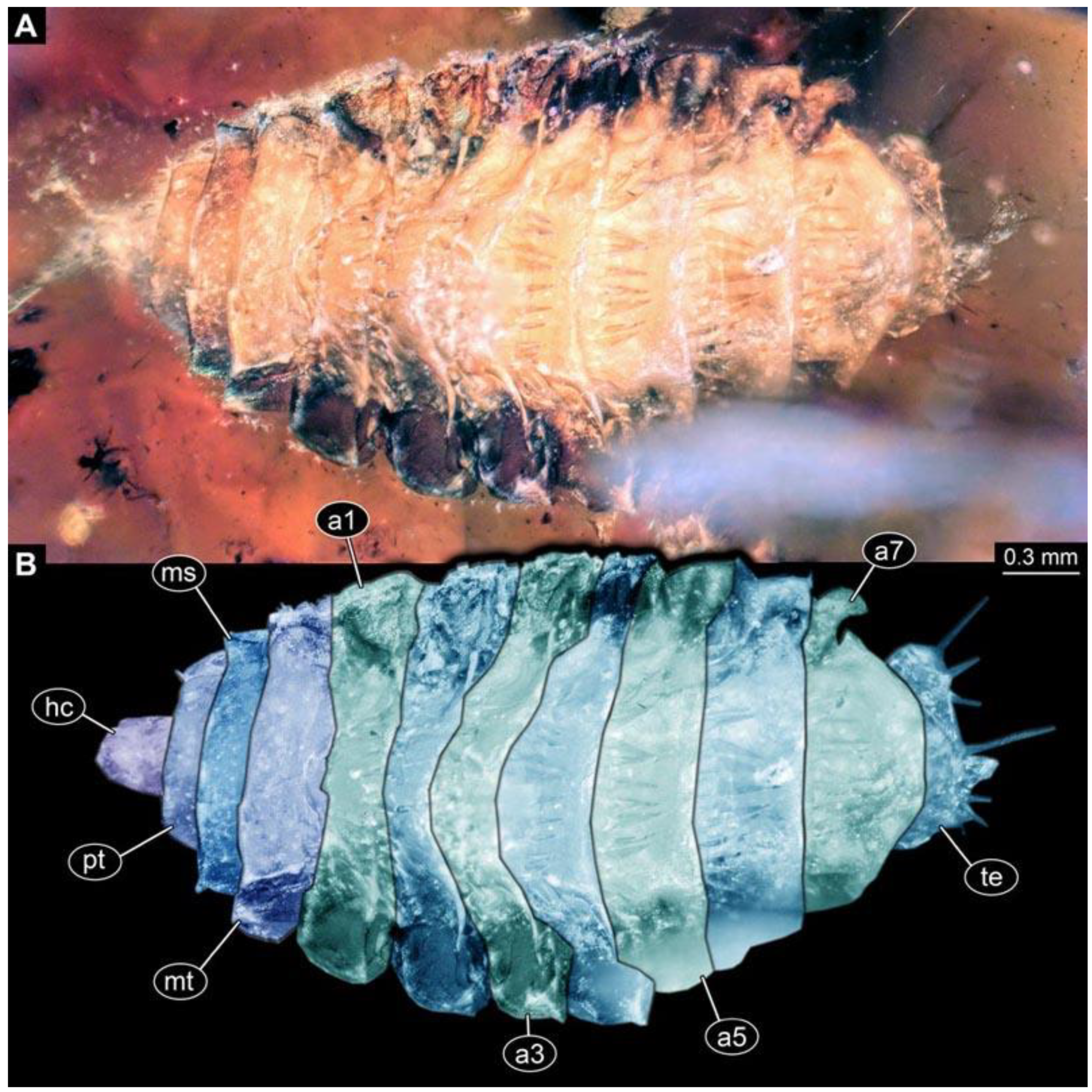
(2) PED 0685: Specimen appears slightly stretched (
Figure 3), partly concealed by blurry amber and particles. Long ventral spine-like setae on abdomen segments well developed. Surface structure apparent. Dimensions: body length 8.27 mm, body width 1.05 mm, head capsule length 0.79 mm, head capsule width 0.11 mm.
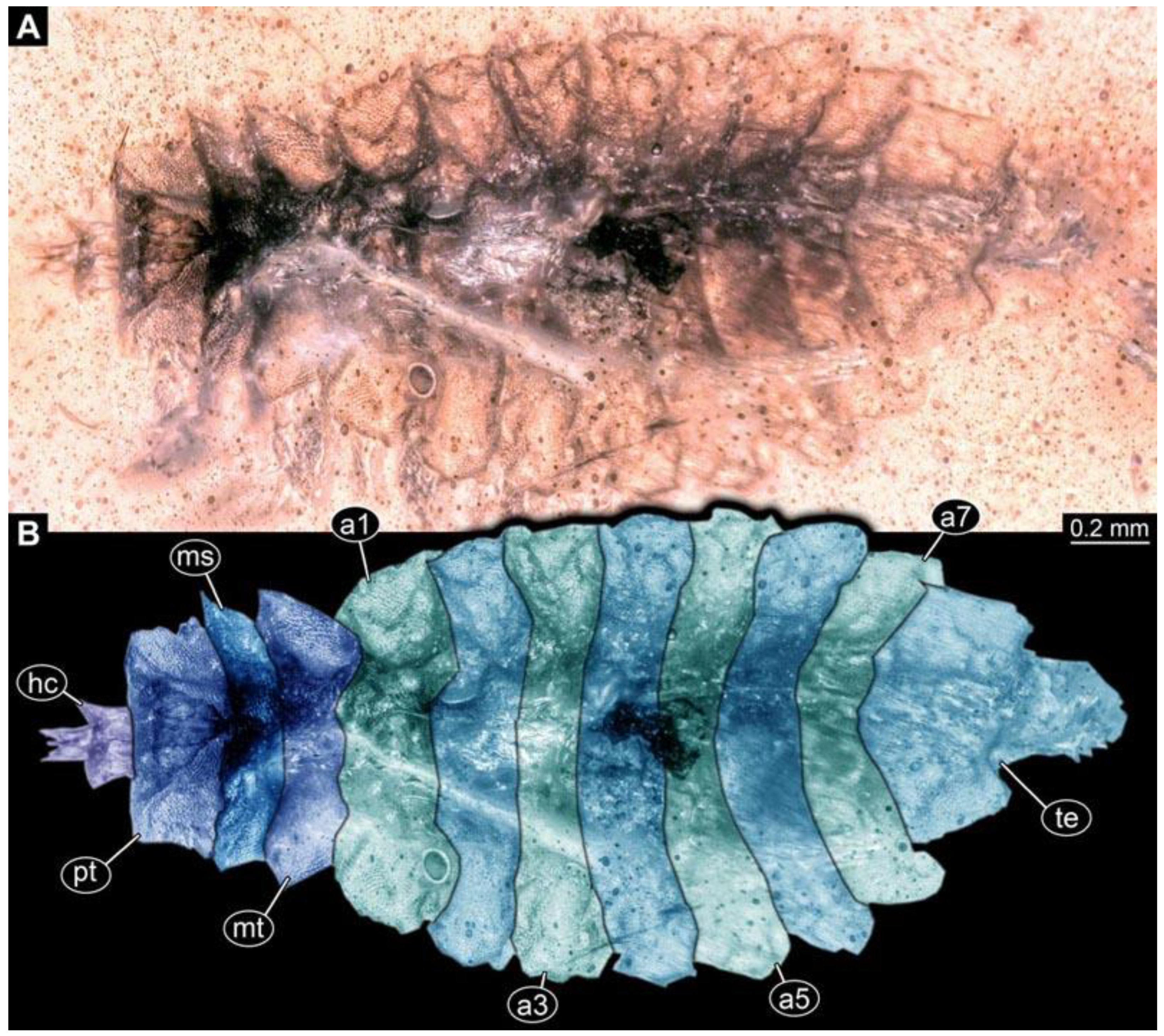
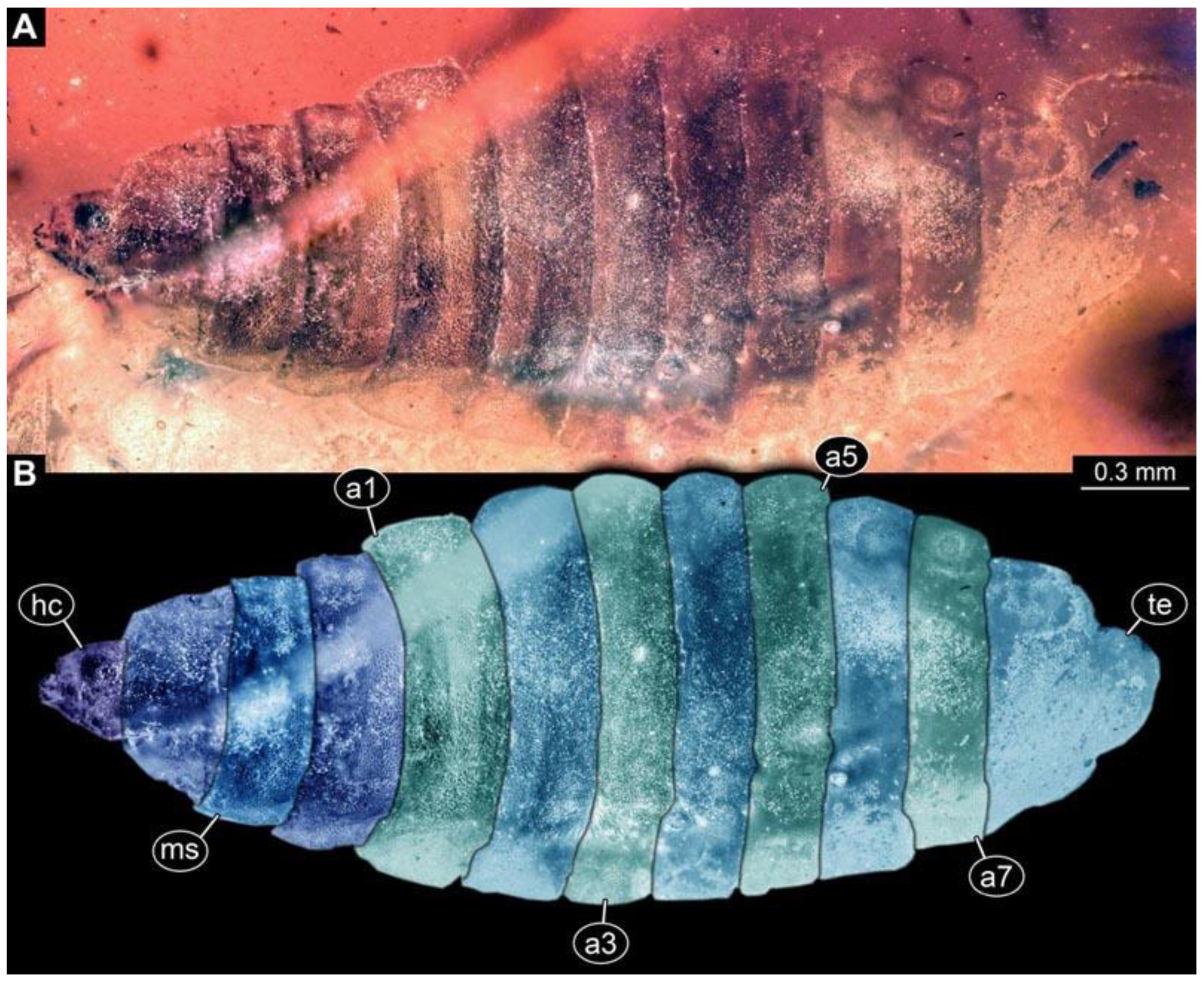
(3) PED 0701: Specimen torn apart ventrally in the more posterior region, also slightly bent (
Figure 4). Surface structure apparent. Head capsule and thorax well preserved. Short spine-like setae on thorax segments and longer ventral spine-like setae on abdomen segments well developed (
Figure 4C). Dimensions: body length 5.08 mm, body width 0.57 mm, head capsule length 0.67 mm, head capsule width 0.13 mm.
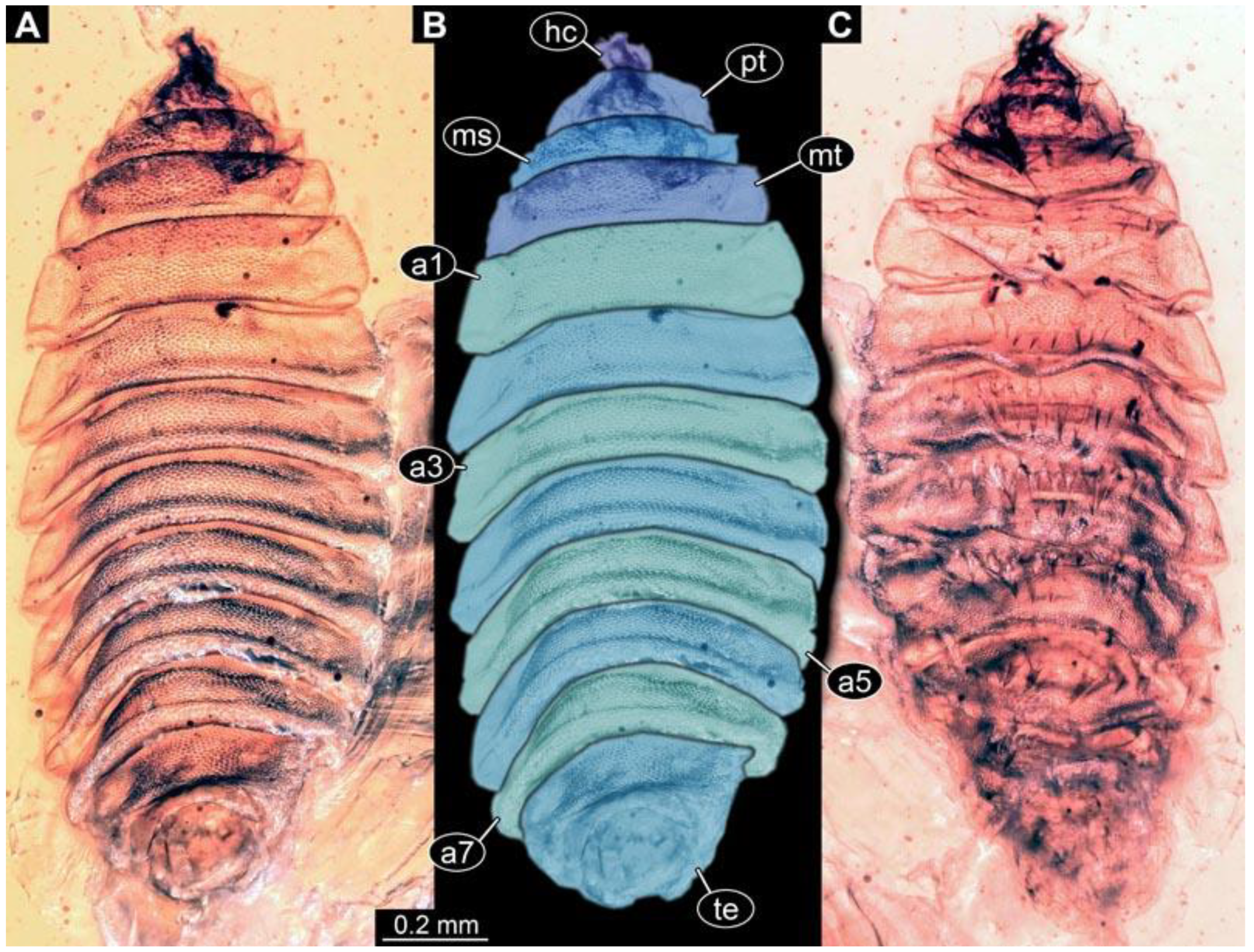
(4) PED 1200-4: Specimen relatively straight, slightly damaged on lateral side of abdomen segment 5 (
Figure 5). Surface structure apparent (
Figure 5D). Head capsule well preserved (
Figure 5C). Two rows of short, spine-like setae per segment of abdomen on ventral and dorsal sides (
Figure 5D). Paired spiracle visible on trunk end. Dimensions: body length 1.3 mm, body width 0.77 mm, head capsule length 0.28 mm, head capsule width 0.13 mm.
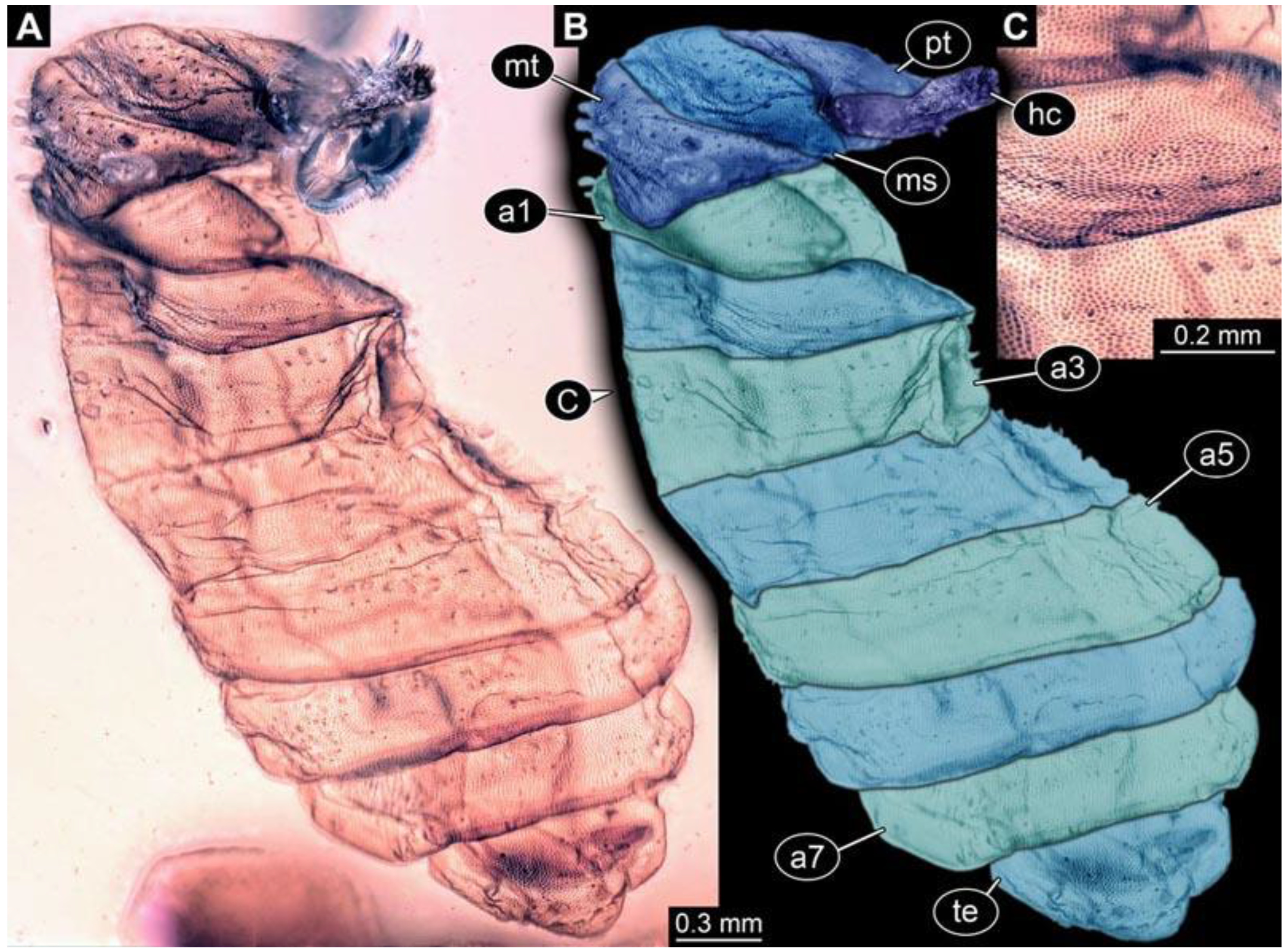
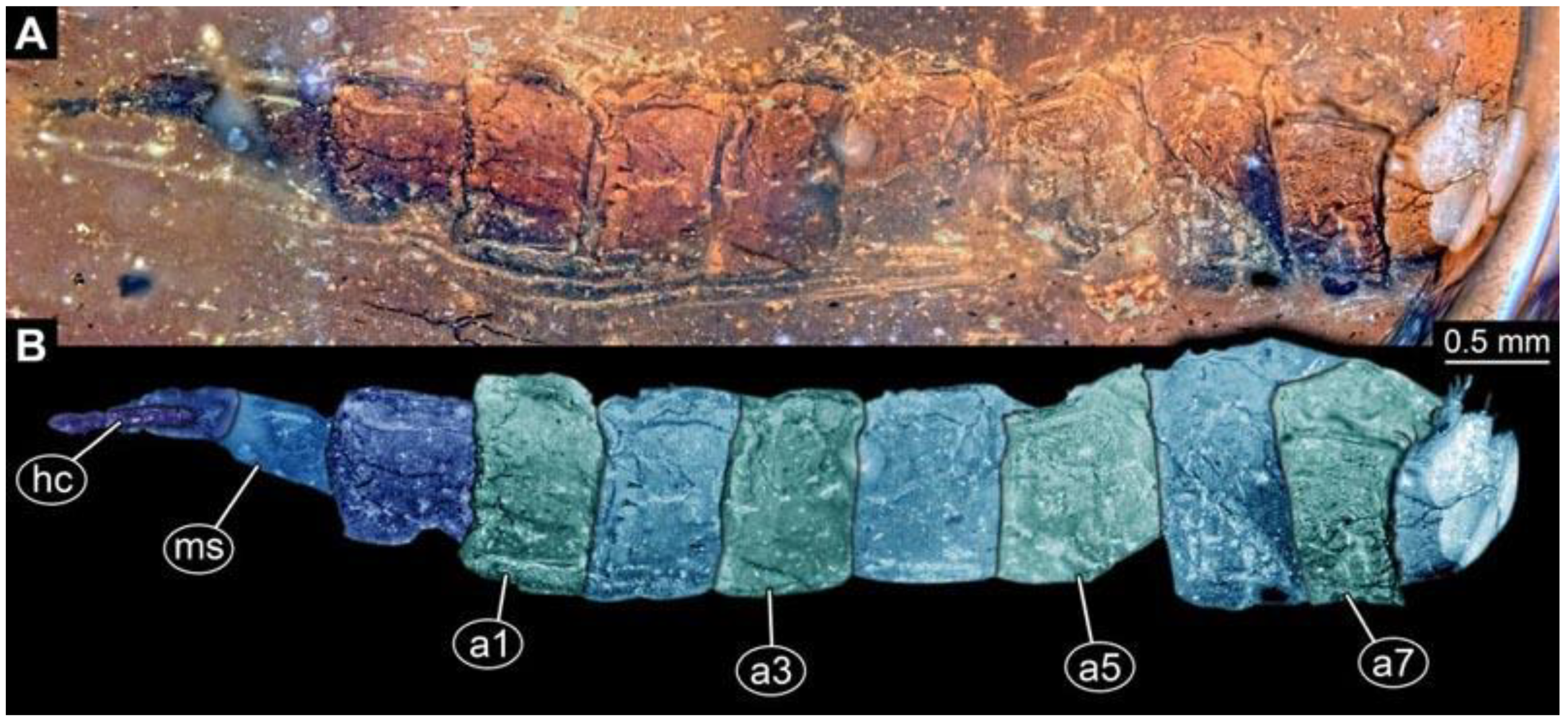
(5) PED 1276: Specimen visible in lateral view, most detail-rich specimen of morphotype 1 (
Figure 6). Surface structure apparent. Head capsule and trunk well preserved (
Figure 6F). One anterior spiracle visible on prothorax. Short spine-like setae on thorax segments and longer ventral spine-like setae on abdomen segments well developed (
Figure 6C–E). Ventral spines on abdomen segment 1 appear shorter; two rows instead of one. Spine-like setae visible on the trunk end. Dimensions: body length 5.58 mm, body width 1.29 mm, head capsule length 0.68 mm, head capsule width 0.25 mm.
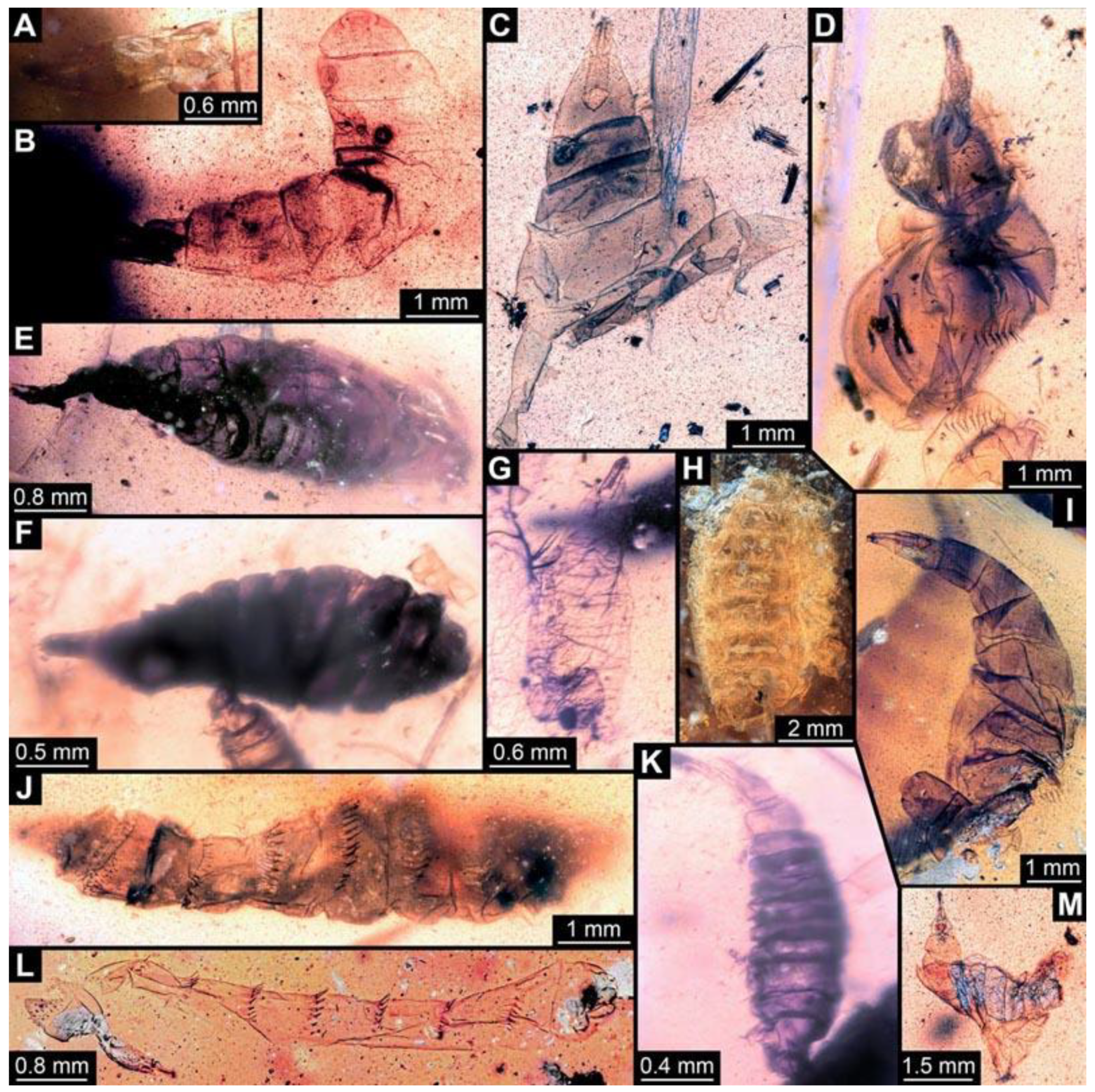
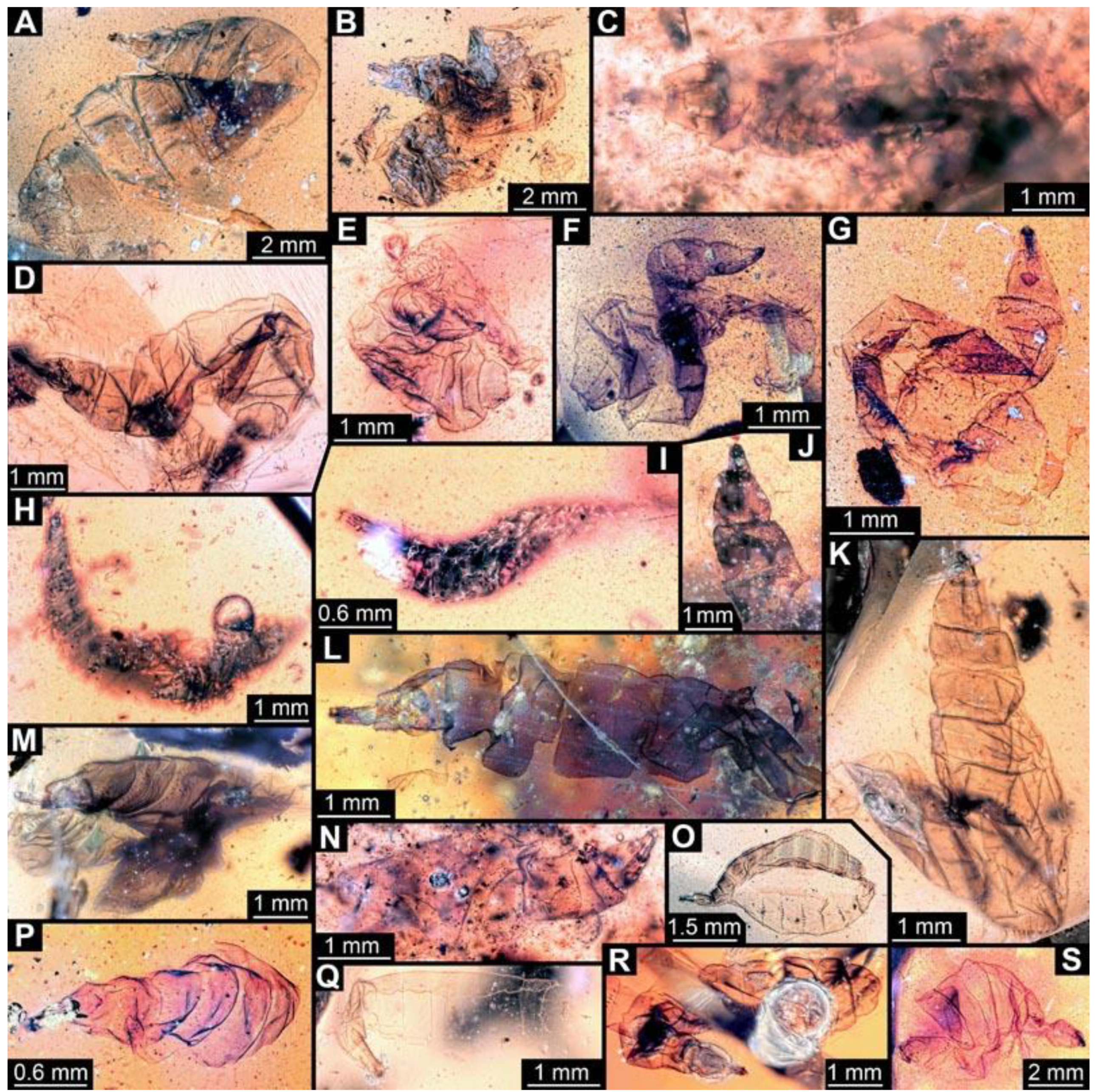
(6) PED 0511: Specimen oblique in the amber (
Figure 7A), numerous particles conceal details. Surface structure apparent. Head capsule and thorax dark pigmented. Ventral spine-like setae on abdomen segments well developed. Specimen bent in the middle of the trunk. No measurements possible due to position of the larva.
(7) PED 0628: Specimen bent in the anterior region, torn apart at the posterior end, slightly concealed by irregularities and particles in the amber (
Figure 7B). Surface structure apparent. Head capsule well preserved. Long ventral spine-like setae on abdomen segments well developed. Dimensions: body width 1.23 mm, head capsule length 0.84 mm, head capsule width 0.14 mm.
(8) PED 1349: Specimen oblique in the amber piece, only accessible in antero-dorsal view (
Figure 7E). Surface structure apparent. Head capsule well-preserved. No measurements could be recorded due to the orientation of the specimen.
(9) PED 0490: Specimen accessible only in dorsal view (
Figure 7F). Numerous particles conceal details, surface structure apparent. Head capsule well preserved. Short spine-like setae on thorax segments well developed. Specimen is crumpled at the posterior part; total body length could not be measured. Dimensions: body width 1.57 mm, head capsule length 1.4 mm, head capsule width 0.42 mm.
(10) PED 0548: Specimen accessible only in ventral view (
Figure 7G), slightly bent, surrounded by cracks/air inclusions that conceal details. Body and head capsule well preserved. Dimensions: body length 1.3 mm, body width 0.5 mm, head capsule length 0.27 mm, head capsule width 0.11 mm.
(11) PED 0714: Specimen torn apart in the trunk region, only head capsule, thorax and abdomen segments 1 and 2 visible and well-preserved. (
Figure 7H). Surface structure apparent. One anterior spiracle apparent in prothorax. Short spine-like setae on thorax segments and longer ventral spine-like setae on abdomen segments well developed. Total body length could not be measured. Dimensions: body width 1.39 mm, head capsule length 0.98 mm, head capsule width 0.15 mm.
(12) PED 0567-1: Specimen ripped apart at the posterior end (
Figure 7I), therefore total body length could not be reliably measured. Surface structure apparent. Head capsule well preserved. Dimensions: body width 1.2 mm, head capsule length 0.57 mm, head capsule width 0.15 mm.
(13) PED 0567-3: Specimen cut off at the edge of the amber, only prothorax, mesothorax, metathorax and abdomen segment 1 apparent (
Figure 7J). Head capsule well preserved. Short spine-like setae on thorax segments well developed. Dimensions: body width 1.7 mm, head capsule length 1.08 mm, head capsule width 0.32 mm.
(14) PED 0497: Specimen accessible in dorsal and ventral view, better visible in dorsal view (
Figure 8B). Numerous dirt particles in the amber, one large dark object conceals the head capsule, which is only visible with transmitted light and enhanced brightness (
Figure 8A). Short spine-like setae on thorax segments well developed. Paired posterior spiracles in trunk end. Trunk is bent by about 90° in the middle, so the total body length could not reliably be measured. Dimensions: body width 1.67 mm, head capsule length 1.8 mm, head capsule width 0.5 mm.
(15) PED 0599-1: Specimen torn off at abdomen segment 2. Head, thorax and abdomen segment 1 well apparent (
Figure 8C). Surface structure apparent. Head capsule well preserved. Short spine-like setae on thorax segments well developed. Total body width was measured on the widest point of the last intact segment. Dimensions: body width 2.33 mm, head capsule length 1.4 mm, head capsule width 0.24 mm.
(16) PED 0599-2: Specimen very crumpled (
Figure 8D), therefore no measurements of the body possible. Surface structure apparent. Head capsule well preserved. Long ventral spine-like setae on abdomen segments well developed. Dimensions: head capsule length 1.14 mm, head capsule width 0.46 mm.
(17) PED 0550: Specimen oblique in the amber (
Figure 8E). Head capsule is slightly stretched. Posterior part of body concealed by particles in the amber; therefore body length could not be reliably measured. Dimensions: body width 1.76 mm, head capsule length 1.51 mm, head capsule width 0.29 mm.
(18) PED 0611-1: Specimen very darkly pigmented (
Figure 8F), dirt particles and blurry amber conceal most details. Head capsule well preserved. Dimensions: body length 3 mm, body width 1 mm, head capsule length 0.42 mm, head capsule width 0.2 mm.
(19) PED 0611-3: Specimen is slightly bent, parts concealed by blurry amber (
Figure 8G). Head capsule and trunk well preserved. Long ventral spine-like setae on abdomen segments well developed. Dimensions: body length 2.18 mm, body width 0.56 mm, head capsule length 0.67 mm, head capsule width 0.09 mm.
(20) PED 0620: Specimen seemingly dorso-ventrally compressed (
Figure 8H), many cracks and air inclusions in the amber piece concealing details. Trunk end slightly folded inwards but still apparent. Head capsule bent by 90° to the trunk, but still visible and well-preserved. Dimensions: body length 5.78 mm, body width 2.63 mm, head capsule length 1.2 mm, head capsule width 0.13 mm.
(21) PED 0601: Specimen slightly bent, the posterior part is torn apart (
Figure 8I). Surface structure apparent. Head capsule well preserved. Ventral spine-like setae visible on torn posterior part. Total length of the body could not be reliably measured. Dimensions: body width 2.18 mm, head capsule length 1.2 mm, head capsule width 0.36 mm.
(22) PED 0551: Specimen accessible in dorsal and ventral view, better visible in ventral view (
Figure 8J), surrounding amber blurry in some areas. Head capsule well preserved, but partly concealed by irregularities in the amber. Short spine-like setae on thorax segments well developed, longer ventral spine-like setae on abdomen segments well developed. Dimensions: body length 8 mm, body width 1.78 mm, head capsule length 1.34 mm, head capsule width 0.34 mm.
(23) PED 0611-2: Specimen concealed by dirt particles and blurry amber, posterior part crumpled (
Figure 8K). Head capsule and trunk well preserved. Total body length could not be reliably measured. Dimensions: body width 0.74 mm, head capsule length 0.33 mm, head capsule width 0.1 mm.
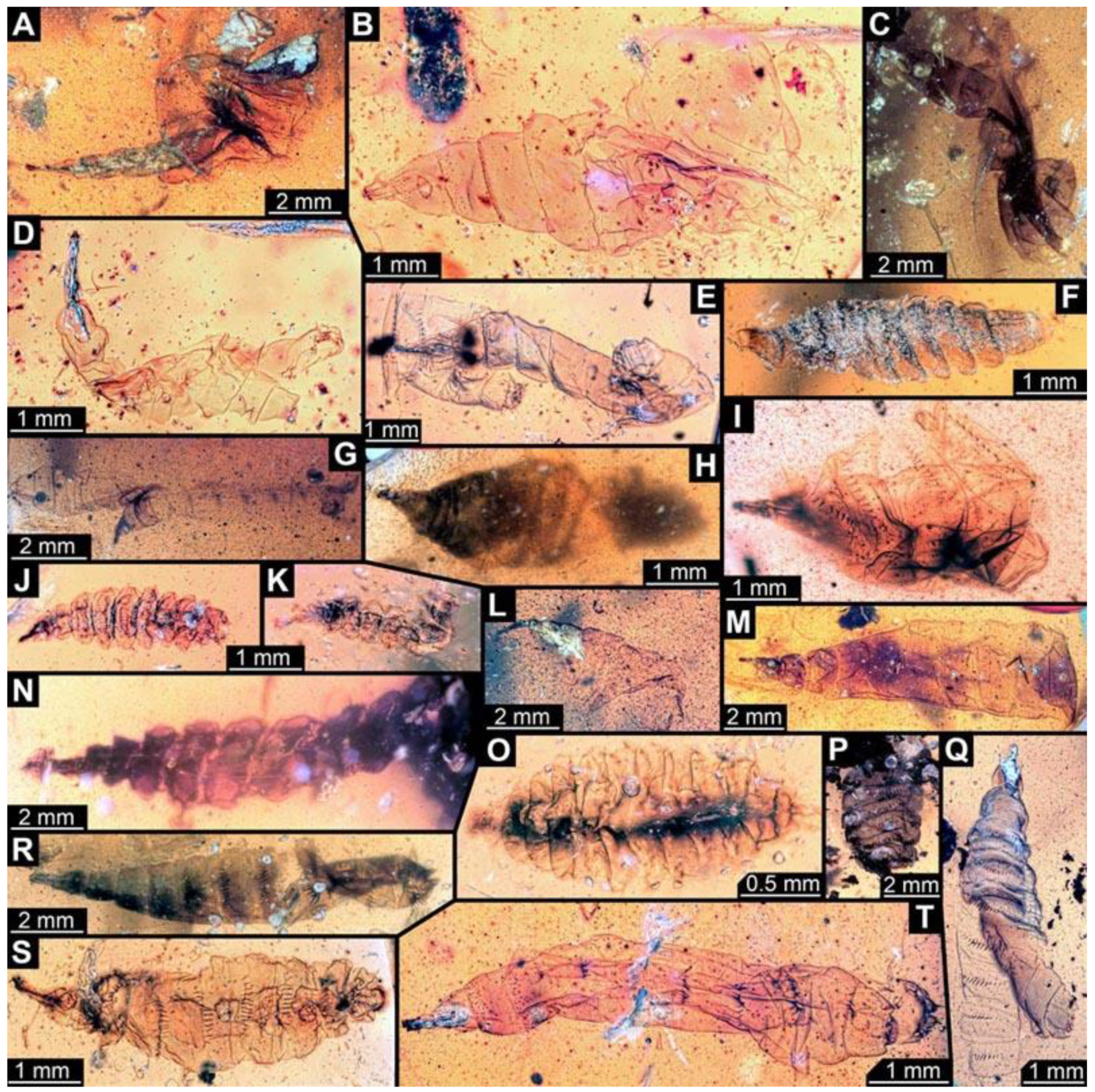
(24) PED 0721: Specimen appears stretched (
Figure 8L). Surface structure apparent. Head capsule, prothorax and parts of mesothorax torn off from the rest of the trunk. Head capsule well preserved. Short spine-like setae on thorax segments and longer ventral spine-like setae on abdomen segments well developed. Dimensions: body width 0.58 mm, head capsule length 0.86 mm, head capsule width 0.14 mm.
(25) PED 0691: Specimen crumpled and damaged (
Figure 8M). Surface structure visible. Head capsule well preserved. Paired anterior spiracles visible on prothorax. Short spine-like setae on thorax segments well developed. No measurements of the body were recorded. Dimensions: head capsule length 0.92 mm, head capsule width 0.17 mm.
(26) PED 0622: Specimen strongly bent between abdomen segments 2 and 3 (
Figure 9A). Surface structure apparent. Head capsule well preserved. Ventral spine-like setae on abdomen segments well developed. The total body length could not be reliably measured. Dimensions: body width 3.33 mm, head capsule length 1.37 mm, head capsule width 0.27 mm.
(27) PED 0653: Specimen strongly crumpled (
Figure 9B), air inclusions and particles in the amber conceal most details, no measurements of the body were recorded. Head capsule well preserved. Short spine-like setae on thorax segments and longer ventral spine-like setae on abdomen segments well developed. Dimensions: head capsule length 1.31 mm, head capsule width 0.33 mm.
(28) PED 0761-1: Mostly concealed by dirt particles, only head capsule, thorax and abdomen segment 1 visible (
Figure 9C). Surface structure apparent. Head capsule well preserved. Paired anterior spiracles visible on prothorax. Total body length could not be measured. Dimensions: body width 1.21 mm, head capsule length 0.98 mm, head capsule width 0.26 mm.
(29) PED 0729: Specimen very crumpled (
Figure 9D). Surface structure apparent. Head capsule well preserved. Short spine-like setae on thorax segments well developed. Measurements of the body could not be recorded. Dimensions: head capsule length 1.36 mm, head capsule width 0.17 mm.
(30) PED 0743: Specimen very crumpled (
Figure 9E). Surface structure apparent. Head capsule well preserved. Short spine-like setae on thorax segments well developed. Measurements of the body could not be recorded. Dimensions: head capsule length 0.75 mm, head capsule width 0.14 mm.
(31) PED 0749: Specimen very crumpled (
Figure 9F). Surface structure apparent. Head capsule well preserved. Short spine-like setae on thorax segments well developed. Some of the longer ventral spine-like setae on abdomen segments well developed. No measurements of the body could be recorded. Dimensions: head capsule length 1.01 mm, head capsule width 0.33 mm.
(32) PED 0752: Specimen very crumpled and torn apart (
Figure 9G). Surface structure apparent. Head capsule well preserved. Short spine-like setae on thorax segments well developed. Some of the longer ventral spine-like setae on abdomen segments visible. No measurements of the body could be recorded. Dimensions: head capsule length 1.43 mm, head capsule width 0.5 mm.
(33) PED 0771: Only head capsule and thorax visible, rest of trunk torn off (
Figure 9J). Surface structure apparent. Head capsule well preserved. Short spine-like setae on thorax segments well developed. The total body length could not be measured. Dimensions: body width 1.53 mm, head capsule length 1.45 mm, head capsule width 0.48 mm.
(34) PED 0770: Specimen strongly crumpled and torn apart (
Figure 9K). Surface structure apparent. Head capsule well preserved. Short spine-like setae on thorax segments well developed. Total body length could not be measured. Dimensions: body width 1.53 mm, head capsule length 1.09 mm, head capsule width 0.36 mm.
(35) PED 0761-2: Specimen strongly damaged, one half of the trunk torn off (
Figure 9L). Surface structure apparent. Head capsule well preserved. Short spine-like setae on thorax segments well developed. Total body length could not be measured. Dimensions: body width 1.61 mm, head capsule length 1.44 mm, head capsule width 0.26 mm.
(36) PED 0762: Specimen oblique in the amber piece, strongly crumpled (
Figure 9M). Surface structure apparent. Head capsule well preserved. Short spine-like setae on thorax segments well developed. No measurements of the body could be recorded. Dimensions: head capsule length 1.41 mm, head capsule width 0.23 mm.
(37) PED 0761-3: Mostly concealed by dirt particles, only head capsule, thorax and some abdomen segments visible (
Figure 9N). Surface structure apparent. Head capsule well preserved. One anterior spiracle apparent on prothorax. Short spine-like setae on thorax segments well developed. Dimensions: body width 0.87 mm, head capsule length 0.88 mm, head capsule width 0.25 mm.
(38) PED 0846: Specimen separated into ventral and dorsal halves (
Figure 9O). Surface structure apparent. Head capsule and prothorax well preserved. Short spine-like setae on thorax segments and longer ventral spine-like setae on abdomen segments well developed. Paired spiracles visible on trunk end. No measurements of the body could be recorded. Dimensions: head capsule length 0.77 mm, head capsule width 0.12 mm.
(39) PED 0766: Specimen torn in the posterior region (
Figure 9P). Surface structure apparent. Head capsule well preserved. Short spine-like setae on thorax segments well developed. No measurements of the body could be recorded. Dimensions: head capsule length 0.68 mm, head capsule width 0.17 mm.
(40) PED 0811-3: Head capsule and thorax of specimen bent, rest of body straight and relatively transparent (
Figure 9Q). Surface structure apparent. Head capsule well preserved. Short spine-like setae on thorax segments well developed. Total body length could not be reliably measured. Dimensions: body width 1.08 mm, head capsule length 0.79 mm, head capsule width 0.26 mm.
(41) PED 0817: Specimen bent in posterior region, separated into ventral and dorsal halves (
Figure 9R). Surface structure apparent. Head capsule well preserved. Short spine-like setae on thorax segments and longer ventral spine-like setae on abdomen segments well developed. Paired spiracles visible on trunk end. Total body length could not be reliably measured. Dimensions: body width 1.42 mm, head capsule length 0.89 mm, head capsule width 0.22 mm.
(42) PED 0923: Specimen crumpled and torn apart (
Figure 9S). Head capsule appears inflated. No measurements could be recorded. Short spine-like setae on thorax segments and longer ventral spine-like setae on abdomen segments well developed. Surface structure apparent.
(43) PED 0850: Specimen crumpled and torn apart, dirt particles conceal details (
Figure 10A). Head capsule slightly stretched. Long ventral spine-like setae on abdomen segments well developed. No measurements of the body could be recorded. Dimensions: head capsule length 1.29 mm, head capsule width 0.19 mm.
(44) PED 0873-1: Specimen crumpled and torn apart from abdomen segment 2 to trunk end (
Figure 10B). Surface structure apparent. Head capsule slightly damaged. Paired anterior spiracles visible on prothorax. Short spine-like setae on thorax segments and longer ventral spine-like setae visible on remains of abdomen segments. Total body length could not be measured. Dimensions: body width 1.42 mm, head capsule length 0.95 mm, head capsule width 0.25 mm.
(45) PED 1018: Specimen very crumpled, air inclusions cover some parts (
Figure 10C). Surface structure apparent on thorax segments. Head capsule well preserved, with bright pigmentation. Paired anterior spiracles are visible on prothorax. Short spine-like setae on thorax segments and longer ventral spine-like setae on abdomen segments well developed. Total body length could not be reliably measured. Dimensions: body width 1.33 mm, head capsule length 1.44 mm, head capsule width 0.31 mm.
(46) PED 0873-2: Specimen crumpled and torn apart from mesothorax to trunk end (
Figure 10D). Surface structure apparent. Head capsule stretched. Short spine-like setae on thorax segments and longer ventral spine-like setae on abdomen segments well developed. No measurements of body could be recorded. Dimensions: head capsule length 1.71 mm, head capsule width 0.27 mm.
(47) PED 0962: Specimen separated into ventral and dorsal halves (
Figure 10E). Surface structure apparent. Head capsule well preserved. Paired anterior spiracles visible on prothorax. Short spine-like setae on thorax segments and longer ventral spine-like setae on abdomen segments well developed. No measurements of the body could be recorded. Dimensions: head capsule length 0.69 mm, head capsule width 0.14 mm.
(48) PED 1137: Specimen only accessible in dorsal view, concealed by numerous bubbles (
Figure 10F). Surface structure apparent. Head capsule missing, but width could be approximated by opening of prothorax. Short spine-like setae on thorax segments well developed. Dimensions: body width 1.18 mm, head capsule width 0.21 mm.
(49) PED 1560: Specimen torn apart in ventral and dorsal halves from thorax to trunk end (
Figure 10G). Numerous dirt particles and air inclusions conceal details. Surface structure apparent. Head capsule well preserved. Short spine-like setae on thorax segments and longer ventral spine-like setae on abdomen segments well developed. Total body length could not be measured. Dimensions: body width 1.33 mm, head capsule length 0.83 mm, head capsule width 0.12 mm.
(50) PED 0987: Specimen twisted in the posterior trunk region, concealed by numerous dirt particles (
Figure 10H). Head capsule not completely visible because of state of the amber and darker pigmentation on the larva. Short spine-like setae on thorax segments well developed. Total body length and length of head capsule could not be reliably measured. Dimensions: body width 2.87 mm, head capsule width 0.28 mm.
(51) PED 1167: Specimen crumpled, numerous dirt particles in the amber conceal details (
Figure 10I). Surface structure apparent. Head capsule well preserved but concealed by blurry amber. Short spine-like setae on thorax segments and longer ventral spine-like setae on abdomen segments well developed. Total body length could not be measured. Dimensions: body width 1.35 mm, head capsule length 1.42 mm, head capsule width 0.26 mm.
(52) PED 0921: Specimen visible in ventral view, overall well preserved (
Figure 10J). Long ventral spine-like setae on abdomen segments well developed. Surface structure apparent. Dimensions: body length 2.95 mm, body width 0.88 mm, head capsule length 0.46 mm, head capsule width 0.08 mm.
(53) PED 0934: Specimen crumpled and torn apart in posterior region (
Figure 10K). Surface structure apparent. Head capsule not fully visible, concealed by dirt particles and bubbles. Total body length could not be measured. Dimensions: body width 0.61 mm, head capsule width 0.06 mm.
(54) PED 1001: Specimen torn apart in the posterior region, numerous particles conceal more details (
Figure 10L). Surface structure apparent on thorax segments. Head capsule slightly bent but overally well preserved. Thorax segments with bright pigmentation. Long ventral spine-like setae on abdomen segments well developed. Total body length could not be measured. Dimensions: body width 1.2 mm, head capsule length 1.12 mm, head capsule width 0.19 mm.
(55) PED 1140: Specimen relatively straight, visible in dorsal view, the largest of all specimens (
Figure 10M). Surface structure apparent. Head capsule well preserved. Short spine-like setae on thorax segments and longer ventral spine-like setae on abdomen segments well developed. Dimensions: body length 12.57 mm, body width 3.07 mm, head capsule length 1.24 mm, head capsule width 0.24 mm.
(56) PED 0990: Specimen poorly recognizable due to numerous particles and bubbles in the amber piece (
Figure 10N). Head capsule mostly covered by bubbles, length of head capsule could not be measured. Posterior part of trunk completely covered by large particles. Total body length could not be measured. Dimensions: body width 2.57 mm, head capsule width 0.45 mm.
(57) PED 1279: Specimen crumpled, numerous particles conceal parts of the head capsule and most of the thorax (
Figure 10P). Surface structure apparent. Head capsule strongly bent, not fully visible, length could not be measured. Long ventral spine-like setae on abdomen segments well developed. Dimensions: body width 3.05 mm, head capsule width 0.33 mm.
(58) PED 1174: Specimen separated into ventral and dorsal halves at posterior part of abdomen (
Figure 10Q). Head capsule missing. Metathorax and abdomen segments 1–3 with bright pigmentation. Surface structure apparent. Short spine-like setae on thorax segments and longer ventral spine-like setae on abdomen segments well developed. Dimensions: body length 6.01 mm, body width 1.25 mm.
(59) PED 1014: Specimen damaged and twisted in posterior part of abdomen, numerous particles and bubbles conceal more details (
Figure 10R). Surface structure apparent. Head capsule well preserved. Short spine-like setae on thorax segments and longer ventral spine-like setae on abdomen segments well developed. Total body length could not be reliably measured. Dimensions: body width 2.05 mm, head capsule length 1.55 mm, head capsule width 0.36 mm.
(60) PED 1025: Specimen only accessible in ventral view, numerous particles conceal some details (
Figure 10S). Surface structure apparent. Head capsule partially concealed by dirt particles. Short spine-like setae on thorax segments and longer ventral spine-like setae on abdomen segments well developed. Paired spiracles visible on trunk end. Dimensions: body length 5.15 mm, body width 1.57 mm, head capsule length 0.87 mm, head capsule width 0.18 mm.
(61) PED 1044: Specimen crumpled, trunk end split (
Figure 10T). Paired anterior spiracles visible on prothorax. Surface structure apparent. Head capsule well preserved. Short spine-like setae on thorax segments and longer ventral spine-like setae on abdomen segments well developed. Paired spiracles visible on trunk end. Dimensions: body length 6.32 mm, body width 1.21 mm, head capsule length 0.81 mm, head capsule width 0.18 mm.
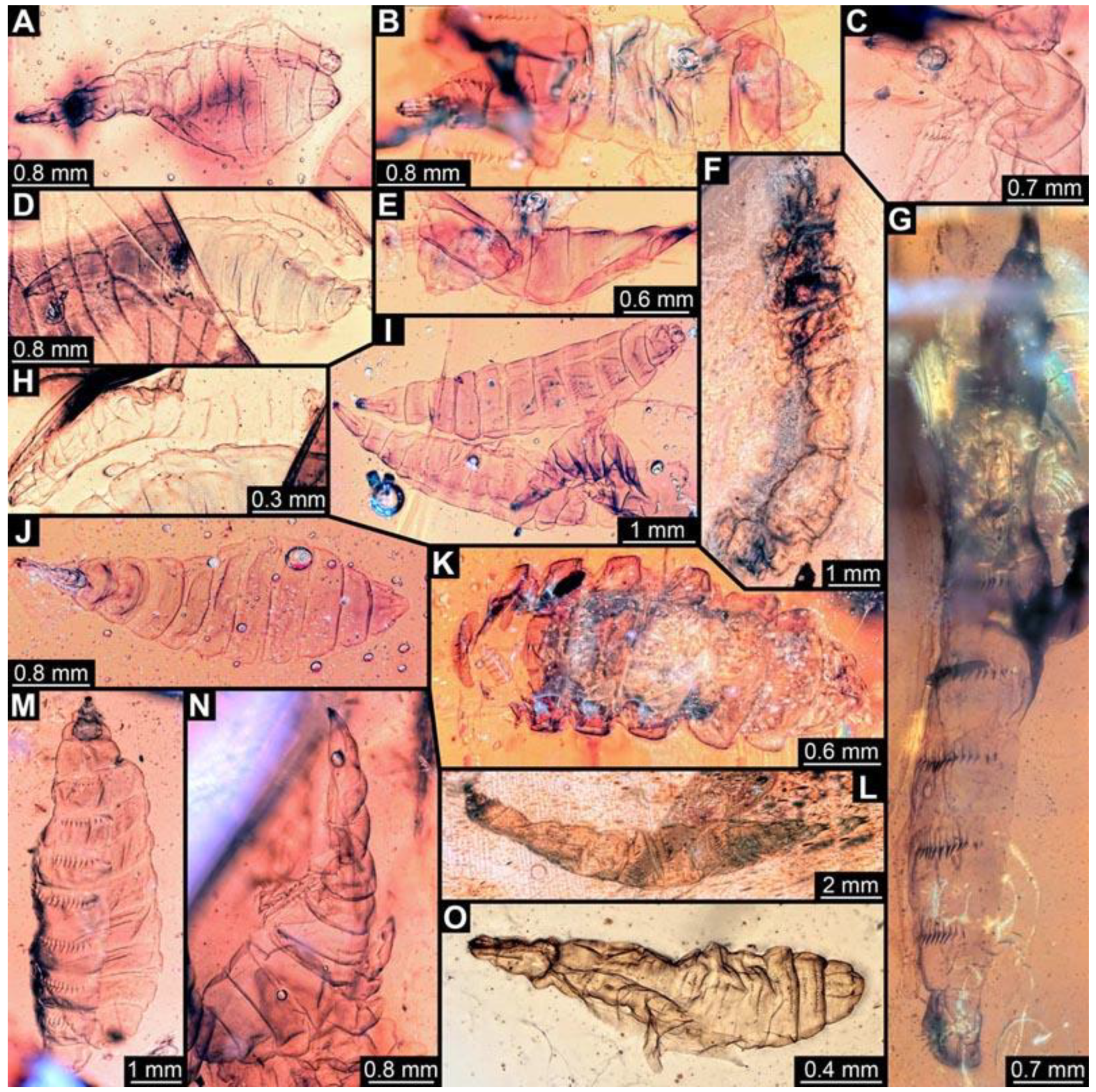
(62) PED 0819-1: Specimen slightly inflated, separated into ventral and dorsal halves (
Figure 11A). Surface structure apparent. Head capsule well preserved. Paired anterior spiracles visible on prothorax. Paired posterior spiracles visible on the trunk end. Ventral spine-like setae on abdomen segments well developed. Body width could not be reliably measured. Dimensions: body length 3.36 mm, head capsule length 0.69 mm, head capsule width 0.19 mm.
(63) PED 0819-4: Specimen partly concealed by other specimens in the amber piece (
Figure 11B). Surface structure apparent. Head capsule well preserved. Paired spiracles on trunk end apparent. Dimensions: body length 3.88 mm, body width 1.13 mm, head capsule length 0.61 mm, head capsule width 0.17 mm.
(64) PED 1200-1: Specimen stretched and bent (
Figure 11C). Surface structure apparent. Head capsule well preserved, but partially covered by particles and bubbles. Short spine-like setae on thorax segments well developed. Total body length could not be measured. Dimensions: body width 0.95 mm, head capsule length 0.64 mm, head capsule width 0.13 mm.
(65) PED 0819-2: Specimen slightly bent (
Figure 11D). Surface structure apparent. Head capsule well preserved. Short spine-like setae on thorax segments well developed. Dimensions: body length 3.51 mm, body width 0.99 mm, head capsule length 0.44 mm, head capsule width 0.19 mm.
(66) PED 0819-5: Specimen partly concealed by other specimens in the amber piece, also posterior part torn off (
Figure 11E). Surface structure apparent. Head capsule well preserved. Long ventral spine-like setae on abdomen segments well developed. Total body length could not be measured. Dimensions: body width 0.71 mm, head capsule length 0.72 mm, head capsule width 0.24 mm.
(67) PED 1274: Specimen crumpled and partly concealed by dirt in the amber (
Figure 11F). Most details not accessible, therefore not measurable.
(68) PED 1247: Specimen stretched, bubbles conceal parts of thorax (
Figure 11G). Surface structure apparent. Head capsule well preserved, but concealed by blurry amber, length of head capsule could not be measured. Short spine-like setae on thorax segments and longer ventral spine-like setae on abdomen segments well developed. Spine-like setae are visible on trunk end. Dimensions: body length 8.3 mm, body width 0.99 mm, head capsule width 0.28 mm.
(69) PED 0819-3: Specimen slightly bent, slightly stretched, oblique in the amber piece (
Figure 11H). Head capsule well preserved. Ventral spine-like setae on abdomen segments well developed, appear rather short. Total body length could not be reliably measured. Dimensions: head capsule length 0.36 mm, head capsule width 0.23 mm.
(70) PED 0819-6: Specimen crumpled and torn apart in trunk region, overlapping with two specimens (0819-7 and 0819-8;
Figure 11I). Surface structure apparent. Head capsule well preserved, but concealed by other specimen. Total body length could not be measured. Dimensions: body width 0.75 mm, head capsule length 0.56 mm, head capsule width 0.25 mm.
(71) PED 0819-7: Specimen overlapping with other specimens in the amber piece (
Figure 11I). Surface structure apparent. Head capsule well preserved. One anterior spiracle apparent on prothorax. Short spine-like setae on thorax segments and longer ventral spine-like setae on abdomen segments well developed. Paired spiracles visible in the trunk end. Dimensions: body length 4.6 mm, body width 1 mm, head capsule length 0.55 mm, head capsule width 0.2 mm.
(72) PED 0819-8: Specimen overlapping with other specimens in the amber piece (
Figure 11I). Surface structure apparent. Head capsule well preserved. Paired anterior spiracles are visible on prothorax. Short spine-like setae on thorax segments and longer ventral spine-like setae on abdomen segments well developed. Dimensions: body length 3.75 mm, body width 0.88 mm, head capsule length 0.55 mm, head capsule width 0.17 mm.
(73) PED 0819-9: Specimen accessible in dorsal and ventral view, better visible in dorsal view, some parts concealed by bubbles (
Figure 11J). Surface structure apparent. Head capsule well preserved. Paired anterior spiracles are visible on prothorax. Paired spiracles visible on trunk end. Dimensions: body length 4.1 mm, body width 1.29 mm, head capsule length 0.69 mm, head capsule width 0.15 mm.
(74) PED 1282: Specimen slightly twisted and bent (
Figure 11L). Numerous particles conceal most details. Surface structure visible in some areas of the abdomen. Head capsule well preserved. Long ventral spine-like setae on abdomen segments well developed. Total body length could not be measured. Dimensions: body width 1.02 mm, head capsule length 0.74 mm, head capsule width 0.2 mm.
(75) PED 1185: Specimen separated into ventral and dorsal halves from thorax to trunk end (
Figure 11M). Surface structure apparent. Head capsule well preserved. Short spine-like setae on thorax segments and longer ventral spine-like setae on abdomen segments well developed. Dimensions: body length 7.38 mm, body width 1.55 mm, head capsule length 0.83 mm, head capsule width 0.19 mm.
(76) PED 1200-2: Specimen slightly bent (
Figure 11N). Surface structure apparent. Head capsule well preserved. Short spine-like setae on thorax segments well developed. Some small ventral spine-like setae are visible on abdomen segment 6. Dimensions: body length 5.03 mm, body width 1.42 mm, head capsule length 0.72 mm, head capsule width 0.13 mm.
(77) PED 1567: Specimen damaged on ventral side, slightly twisted (
Figure 11O). Surface structure barely visible. Head capsule well preserved. Two rows of short spine-like setae visible on dorsal side of abdomen segments, one row of short spine-like setae visible on ventral parts of abdomen segments. Paired spiracles visible on trunk end. Dimensions: body length 1.96 mm, body width 0.5 mm, head capsule length 0.29 mm, head capsule width 0.13 mm.
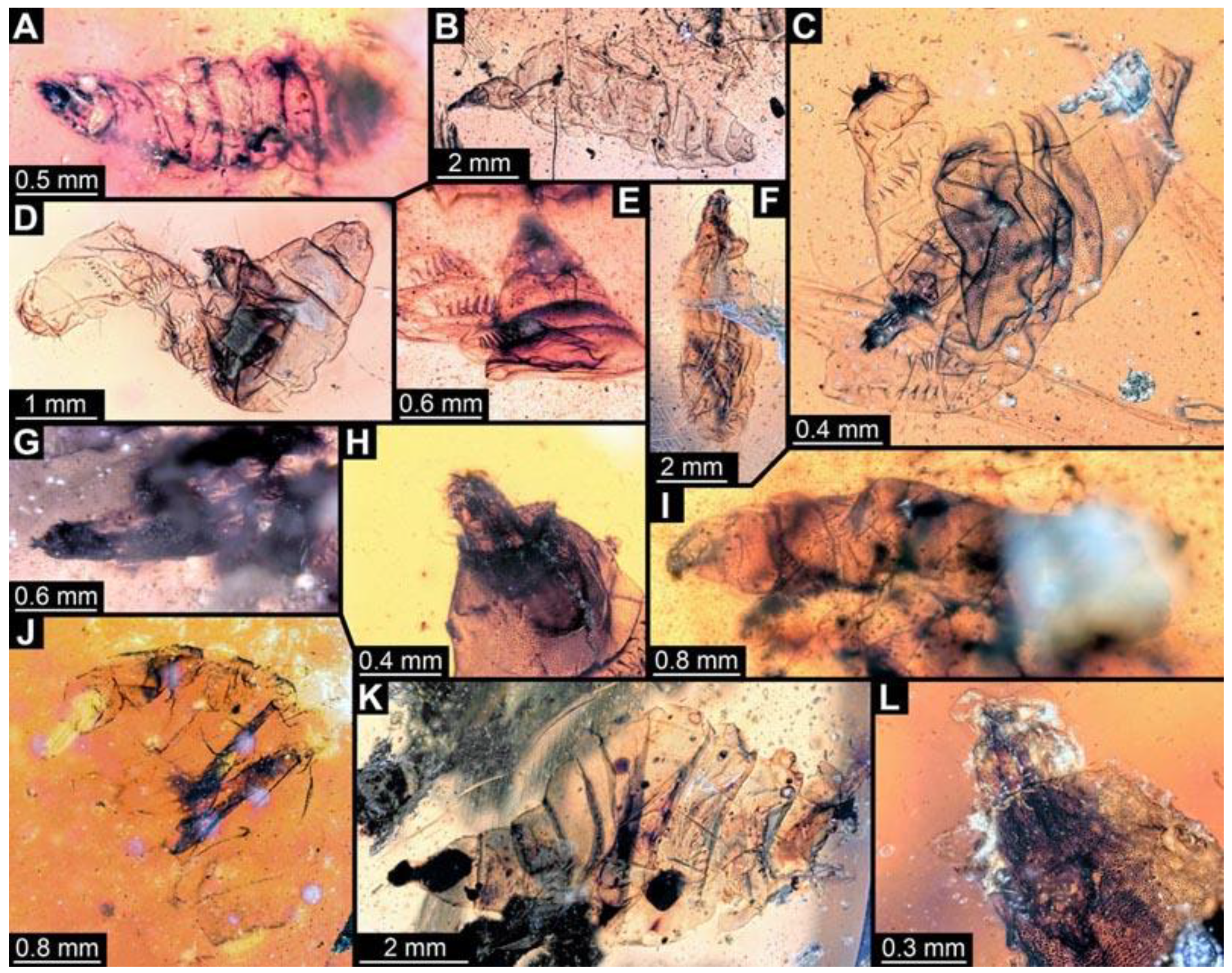
(78) PED 0567-2: Specimen cut off at the edge of the amber, only prothorax and head capsule apparent (
Figure 12A). Dimensions: head capsule length 1.04 mm, head capsule width 0.5 mm.
(79) PED 0681: Specimen torn apart and partly scattered in the amber piece (
Figure 12B). Surface structure apparent. Head capsule well preserved, still attached to prothorax and mesothorax, this anterior region separated from the rest of the trunk. One anterior spiracle apparent on prothorax. Short spine-like setae on thorax segments well developed. No measurements of the body could be recorded. Dimensions: head capsule length 1.12 mm, head capsule width 0.41 mm.
(80) PED 0773-1: Multiple crumpled specimens overlapping in this amber piece, only head capsule of specimen apparent (
Figure 12C). Dimensions: head capsule length 0.42 mm, head capsule width 0.1 mm.
(81) PED 0773-2: Multiple crumpled specimens overlapping in this amber piece, only head capsule and prothorax of specimen apparent (
Figure 12D). Surface structure apparent on prothorax. Dimensions: head capsule length 0.69 mm, head capsule width 0.15 mm.
(82) PED 0773-3: Multiple crumpled specimens overlapping in this amber piece, only head capsule and thorax of specimen apparent (
Figure 12E). Surface structure apparent on thorax. Dimensions: head capsule length 0.6 mm, head capsule width 0.14 mm.
(83) PED 0811-1: Specimen very crumpled and torn apart (
Figure 12F). Only the head capsule and the thorax of the specimen well preserved. Surface structure apparent on thorax. Short spine-like setae on thorax segments well developed. Dimensions: head capsule length 1.62 mm, head capsule width 0.52 mm.
(84) PED 0764: Specimen very crumpled and torn apart (
Figure 12G). Surface structure apparent. Head capsule well preserved. Short spine-like setae on thorax segments well developed. No measurements of the body could be recorded. Dimensions: head capsule length 0.87 mm, head capsule width 0.19 mm.
(85) PED 0811-2: Specimen very crumpled and torn apart (
Figure 12H). Only the head capsule and the prothorax of the specimen well preserved. Surface structure apparent on prothorax. Short spine-like setae on the prothorax well developed. No measurements on body could be recorded. Dimensions: head capsule length 1.46 mm, head capsule width 0.26 mm.
(86) PED 0978: Specimen stretched, crumpled, bent at thorax (
Figure 12I). Surface structure apparent. Head capsule stretched and bent, length of the head capsule could not be reliably measured. Short spine-like setae on thorax segments and longer ventral spine-like setae on abdomen segments well developed. No measurements of the body could be recorded. Dimensions: head capsule width 0.21 mm.
(87) PED 1200-3: Specimen mostly concealed by other specimens, only head capsule and thorax well visible (
Figure 12J). Surface structure apparent. Head capsule well preserved. Short spine-like setae on thorax segments well developed. No measurements of the body could be recorded. Dimensions: head capsule length 0.82 mm, head capsule width 0.25 mm.
(88) PED 1353: Specimen separated into ventral and dorsal halves from thorax to trunk end (
Figure 12K). Numerous particles and bubbles conceal details. Surface structure apparent. Head capsule well preserved, but not completely visible due to dark pigmented thorax segments, length of the head capsule could not be measured. Short spine-like setae on thorax segments and longer ventral spine-like setae on abdomen segments well developed. Dimensions: body width 1.96 mm, head capsule width 0.18 mm.
(89) PED 1535: Specimen only accessible in lateral view, torn apart posteriorly, particles conceal some details (
Figure 12L). Surface structure apparent. Head capsule well preserved, but partly concealed by blurry amber. Long ventral spine-like setae on abdomen segments well developed. Dimensions: body length 3.37 mm, body width 0.46 mm, head capsule length 0.4 mm, head capsule width 0.14 mm.
(90) PED 0614: Specimen oblique within the amber piece, bent in multiple directions (
Figure 12M), numerous cracks and air inclusions conceal most details. Surface structure barely visible on some parts of the larva. No measurements could be recorded.
(91) PED 1548-1: Specimen torn apart posteriorly, numerous particles and bubbles conceal details (
Figure 12N). Surface structure apparent. Head capsule well preserved, partially with bright pigmentation. Long ventral spine-like setae on abdomen segments well developed. Total body length could not be measured. Dimensions: body width 0.61 mm, head capsule length 0.71 mm, head capsule width 0.11 mm.
(92) PED 1548-2: Specimen torn apart posteriorly, numerous particles and bubbles conceal details (
Figure 12O). Surface structure barely visible. Head capsule well preserved, partially with bright pigmentation. Paired spiracles barely visible on trunk end. Total body length could not be measured. Dimensions: body width 0.49 mm, head capsule length 0.33 mm, head capsule width 0.13 mm.
(93) PED 0799: Specimen very crumpled and torn apart (
Figure 12P). Numerous particles conceal most details. Surface structure apparent. Head capsule barely visible and crumpled. Short spine-like setae on thorax segments and longer ventral spine-like setae on abdomen segments well developed. No measurements could be recorded.
(94) PED 0678: Specimen strongly crumpled and damaged (
Figure 12Q), large cracks and bubbles conceal most parts of the larva. Trunk end and abdomen segments 6 and 7 are the only visible, well-preserved parts. Some ventral spine-like setae are visible through the larva. Surface structure visible on the well-preserved segments. No measurements could be recorded.
(95) PED 0738: Specimen barely visible, very crumpled, numerous particles in the amber conceal details of the body (
Figure 12R). Head capsule slightly inflated, with bright pigmentation. No measurements could be recorded.
(96) PED 0760: Specimen torn apart and crumpled (
Figure 12S). Surface structure visible on remnants of the larva. Head capsule mostly covered by cracks and air inclusions. No measurements could be recorded.
(97) PED 0712: Specimen mostly concealed by large cracks and bubbles (
Figure 12T). Mostly thorax visible. Surface structure apparent. Short spine-like setae on thorax segments well developed. No measurements could be recorded.
(98) PED 0927: Specimen crumpled, stretched and bent in multiple directions (
Figure 12U). Concealed by numerous dirt particles and blurry amber. Head capsule appears stretched. No measurements could be recorded.
(99) PED 0777: Specimen twisted, bent, damaged (
Figure 12V). Numerous particles conceal details. Head capsule stretched. No measurements could be recorded.
(100) PED 0861: Specimen damaged (
Figure 13B). Surface structure apparent. Only head capsule, prothorax and mesothorax are well preserved. Short spine-like setae on thorax segments and longer ventral spine-like setae on abdomen segments well developed. No measurements could be recorded.
(101) PED 1007-3: Specimen crumpled and torn apart (
Figure 13C). Surface structure apparent. Head capsule partially covered by bubbles, but well preserved. Long ventral spine-like setae on abdomen segments well developed. Spine-like setae are visible on trunk end. No measurements could be recorded.
(102) PED 0966: Specimen torn apart and crumpled (
Figure 13D). Surface structure apparent. Head capsule oblique, could not be measured. Long ventral spine-like setae on abdomen segments well developed. Spine-like setae visible on trunk end. Paired spiracles visible on trunk end. No measurements could be recorded.
(103) PED 0976-1: Specimen crumpled and torn apart from abdomen segment 2 to trunk end; oblique in the amber piece (
Figure 13E). Numerous particles conceal most parts of specimen. Surface structure apparent. Ventral spine-like setae visible on abdomen. No measurements could be recorded.
(104) PED 1010: Specimen very crumpled (
Figure 13F). Surface structure apparent. Head capsule missing. Short spine-like setae on thorax segments and longer ventral spine-like setae on abdomen segments well developed.
(105) PED 0976-2: Specimen crumpled and torn apart, most parts concealed by particles (
Figure 13G). No measurements could be recorded.
(106) PED 1007-1: Specimen torn apart (
Figure 13H). Surface structure apparent. Head capsule and thorax visible and well-preserved. Short spine-like setae on thorax segments well developed. No measurements could be recorded.
(107) PED 1129: Specimen twisted and mostly concealed by blurry amber (
Figure 13I). Surface structure barely visible. Head capsule missing. Paired spiracles barely visible on trunk end. No measurements could be recorded.
(108) PED 1192: Specimen crumpled, bent and torn apart (
Figure 13J). Details concealed by numerous particles and dark colour of the amber. No measurements could be recorded.
(109) PED 1405: Specimen crumpled, torn apart at trunk end, accessible in ventral view (
Figure 13K). Large particles conceal major parts of the specimen. Surface structure apparent. Head capsule and part of prothorax very dark. Long ventral spine-like setae on abdomen segments well developed. No measurements could be recorded.
(110) PED 1007-2: Only head capsule and prothorax of specimen visible, partially covered by bubbles (
Figure 13L). Surface structure apparent on prothorax. Paired anterior spiracles visible in prothorax. No measurements could be recorded.


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





























