
Characterizing and Quantifying Water Content in 14 Species of Bryophytes Present in Azorean Native Vegetation


Abstract
:1. Introduction
- How will absolute water content (AWC) vary among the selected species?
- Which pathway, ectohydric (external) or endohydric (internal), retains the most water? How are these pathways related to the morphological features of the species?
- How fast do bryophytes lose their water through direct evaporation and how much do they lose?
2. Materials and Methods
2.1. Study Sites
2.2. Study Species
2.3. Sampling Procedure
2.4. Processing Samples in the Laboratory
2.5. Data Analysis
2.5.1. Water Content
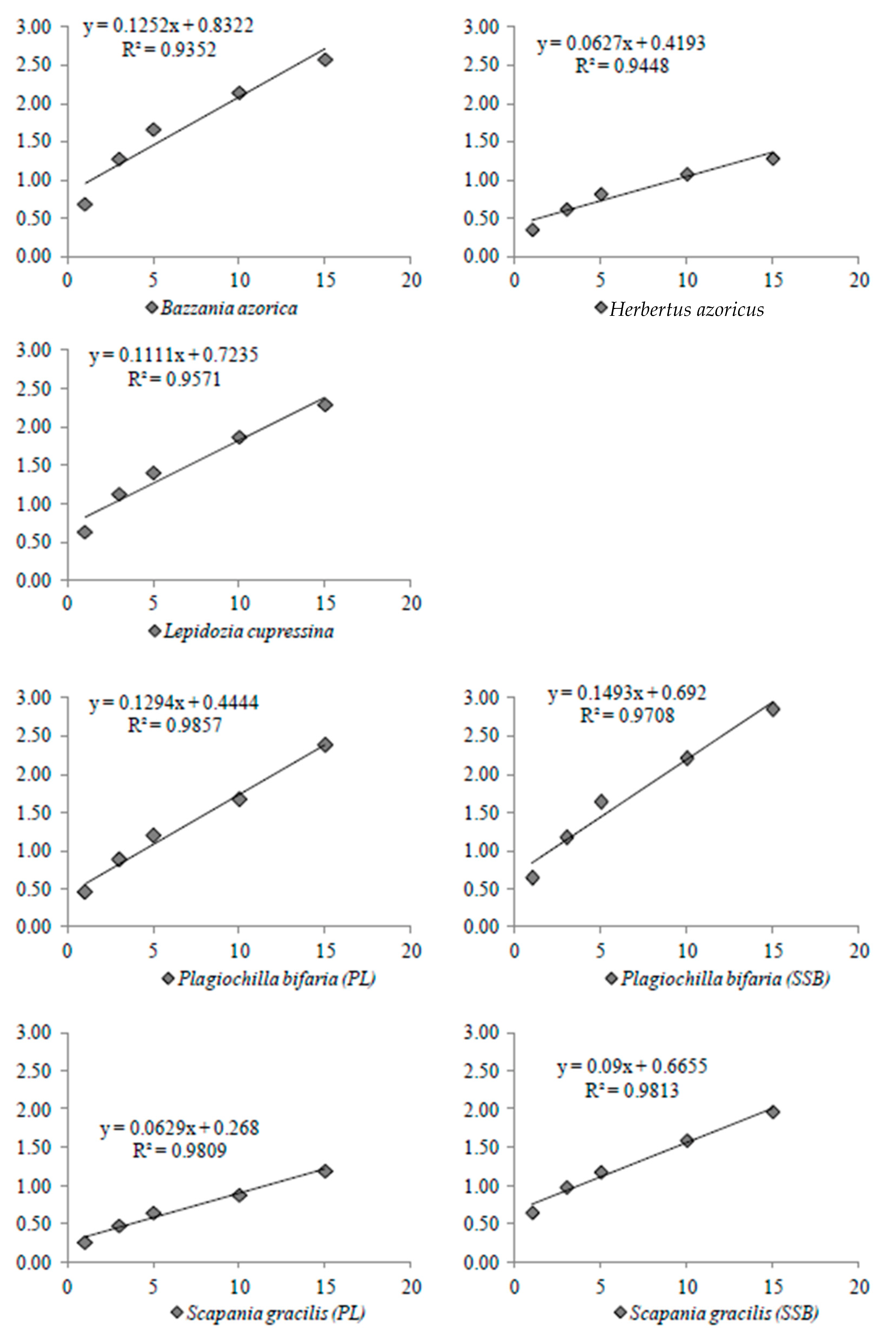
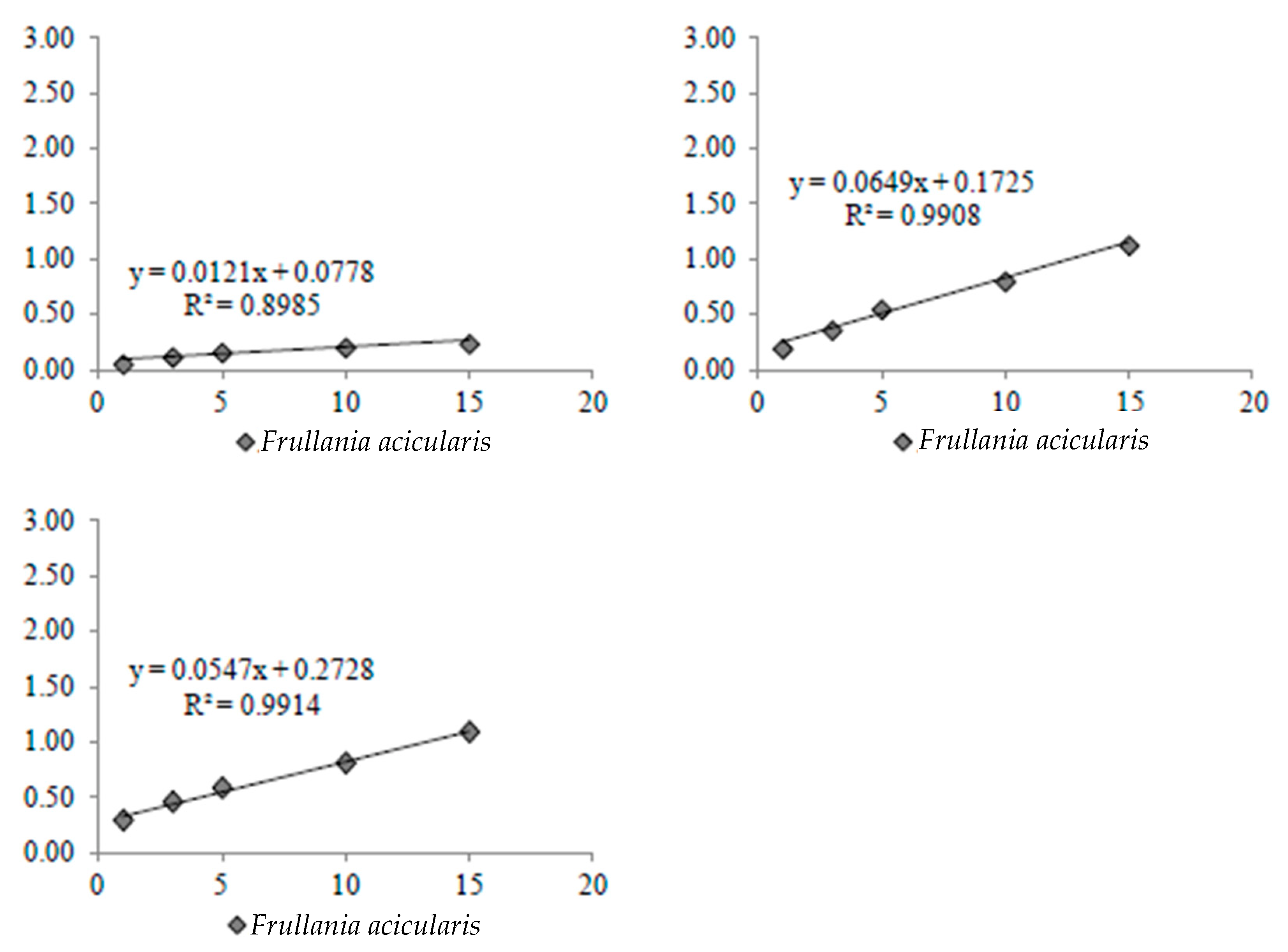
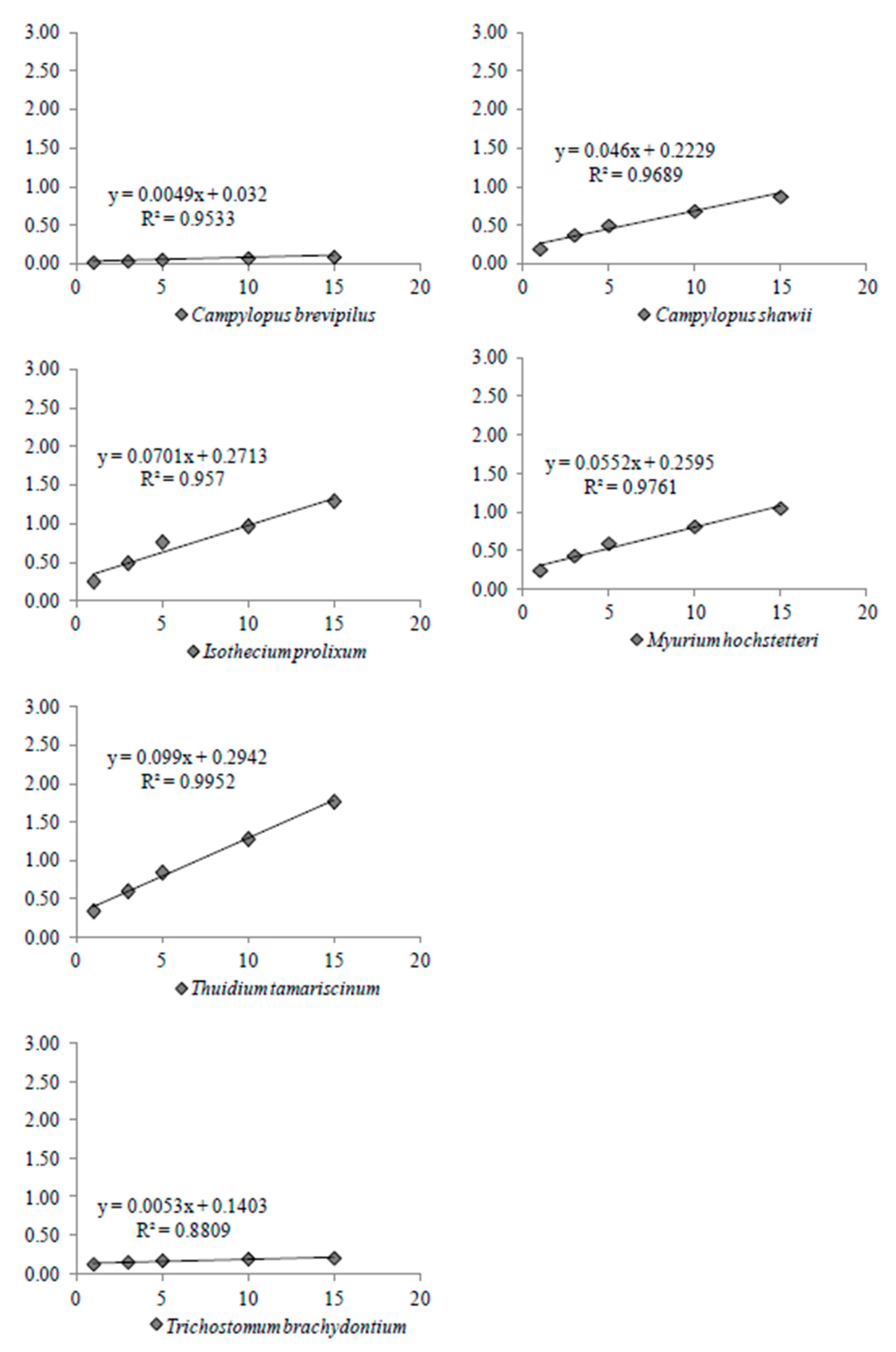
2.5.2. Direct Evaporation Rate
2.5.3. Statistical Analysis
3. Results
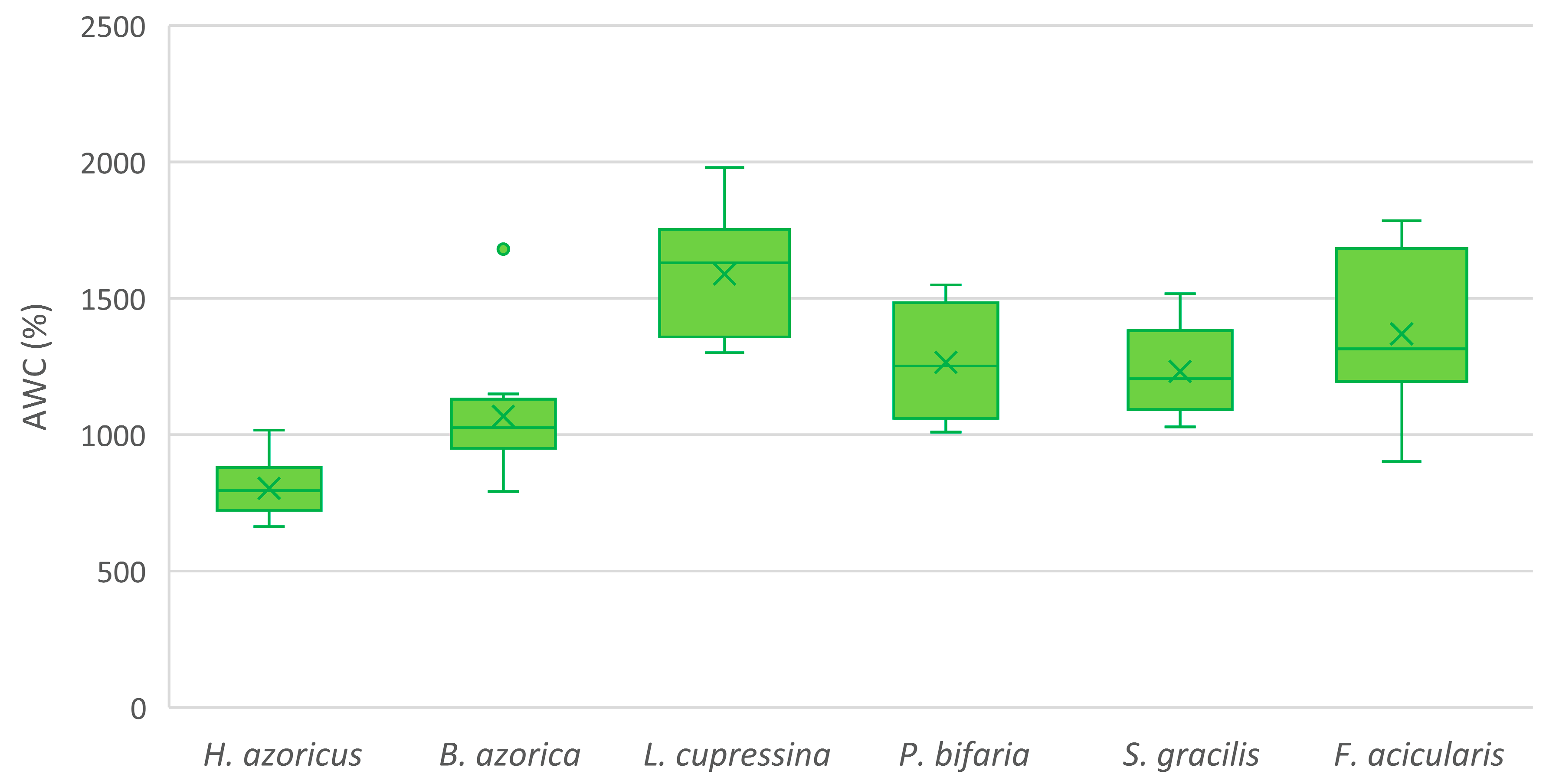
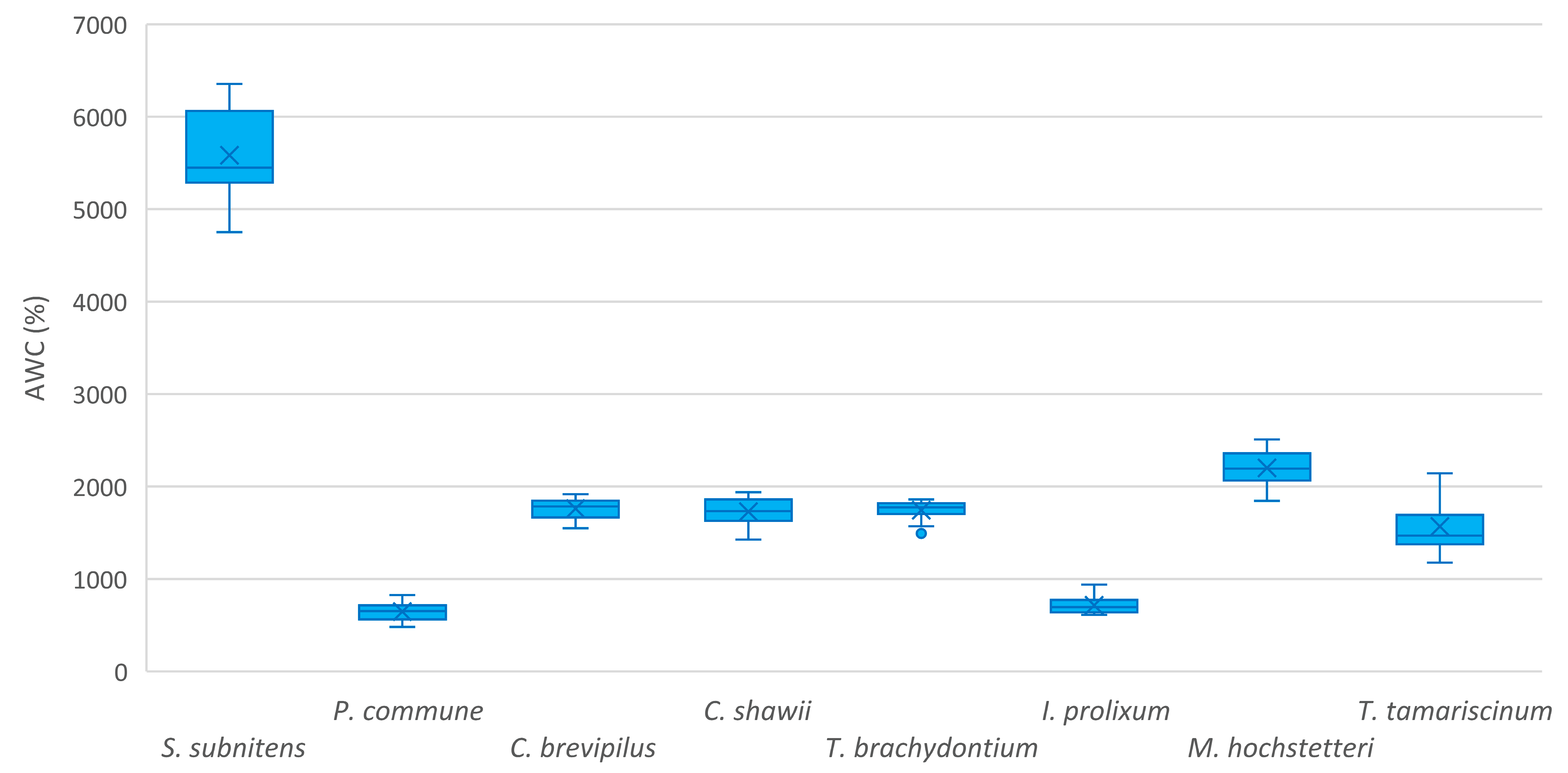
3.1. Absolute Water Content in Azorean Native Bryophytes
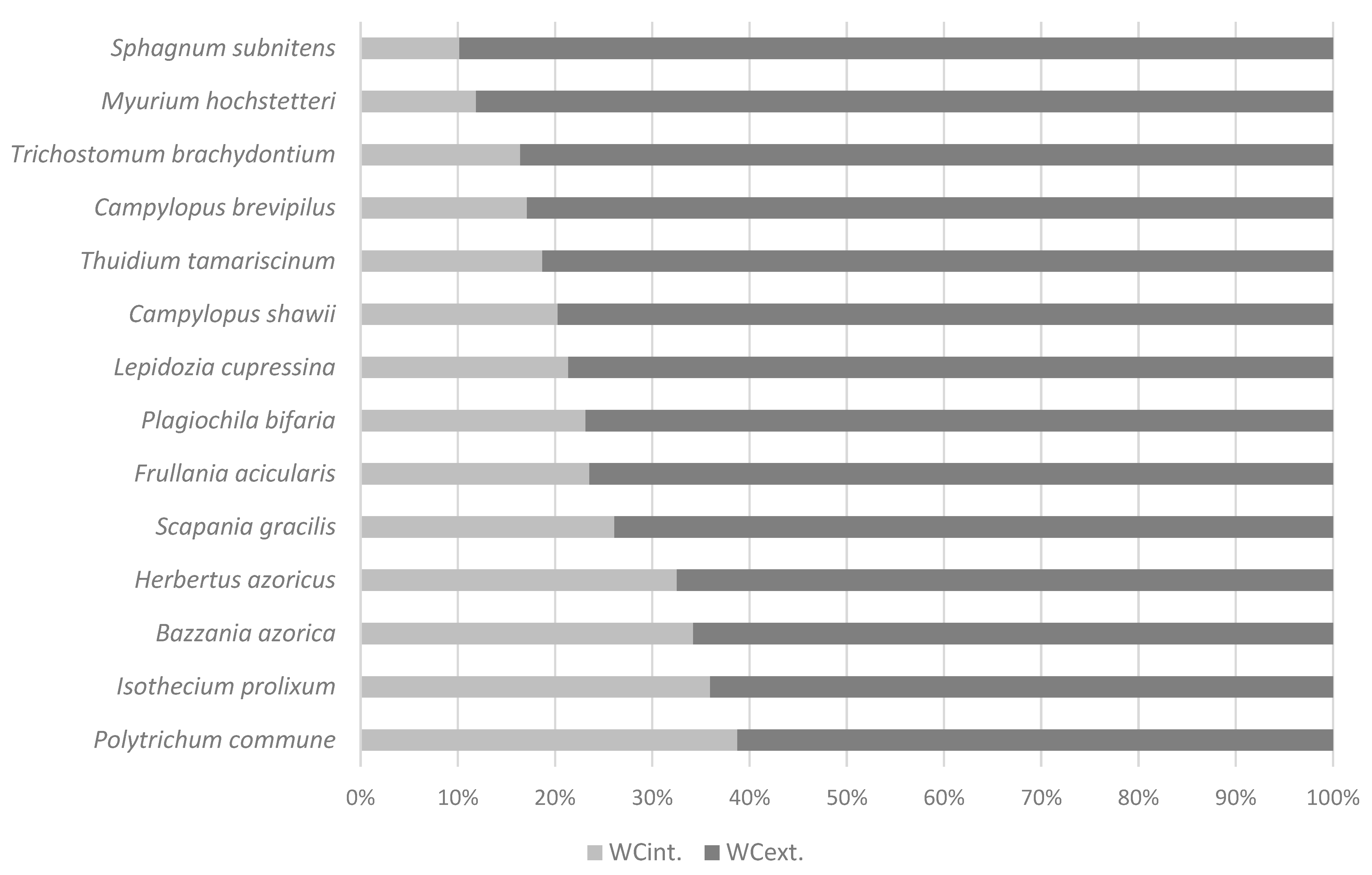
3.2. Internal and External Water Content in Azorean Bryophytes
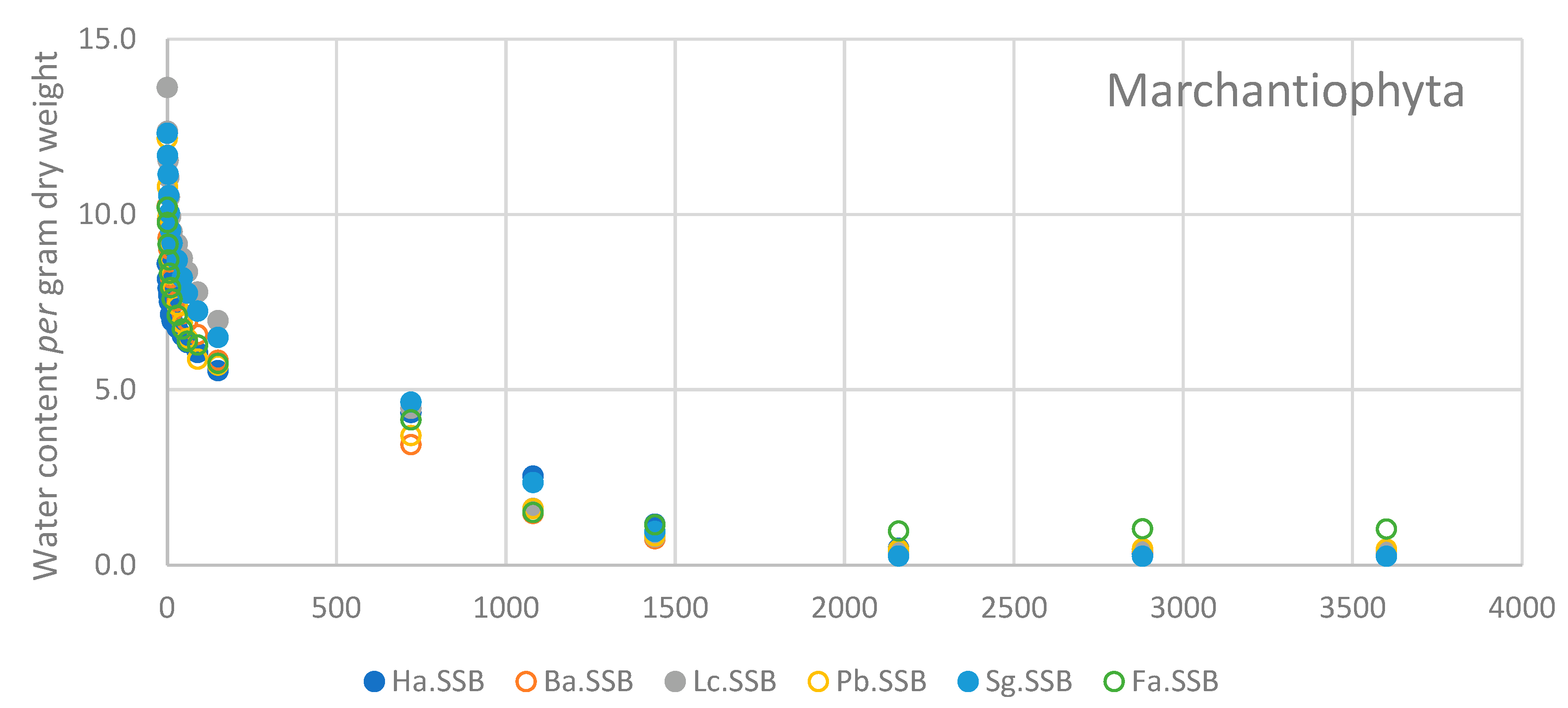
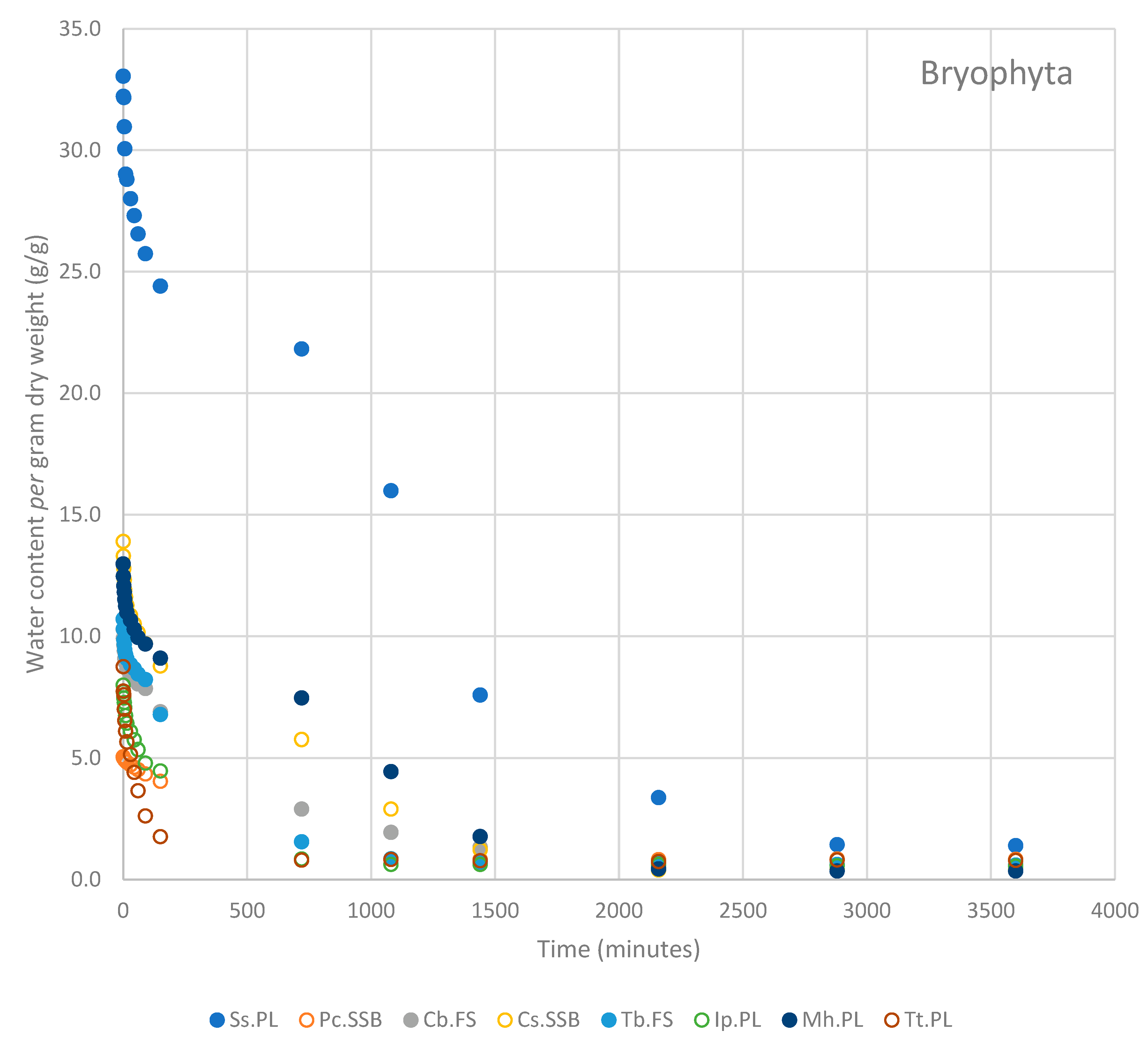
3.3. Rate of Water Loss
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
- A01—Herbertus azoricus (Steph.) P.W.Richards

- A02— Bazzania azorica H.Buch and Perss.

- A03—Lepidozia cupressina (Sw.) Lindenb. subsp. cupressina

- A04—Plagiochila bifaria (Sw.) Lindenb.

- A05—Scapania gracilis Lindb.

- A06—Frullania acicularis Hentschel and von Konrat

- A07—Sphagnum subnitens Russow and Warnst.

- A08—Polytrichum commune Hedw.

- A09—Campylopus brevipilus Bruch and Schimp.

- A10—Campylopus shawii Wilson

- A11—Trichostomum brachydontium Bruch

- A12—Isothecium prolixum (Mitt.) Stech, Sim-Sim, Tangney and D.Quandt.

- A13—Myurium hochstetteri (Schimp.) Kindb.

- A14—Thuidium tamariscinum (Hedw.) Schimp.

Appendix B




Appendix C

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LOCATION | ACTION | OBSERVATIONS |
|---|---|---|
| In the field | Look for an appropriate area, with many large populations of the species you need to collect. | Assure yourself that you have the proper authorization to collect field samples.Assure yourself that the target species occurs in many different patches and that it is abundant in the area, before collecting. |
| Gather an appropriate number of replicates of the target bryophyte species (e.g., 25 cm2) to sealed polyethylene vials, previously marked and weighted. | Try to collect the purest populations, to diminish the cleaning time in the laboratory. Avoid collecting just the central part of the patch, to keep the integrity of the sample in the field, as much as possible. | |
| Collect a small Herbarium sample of each target-species to be able to confirm its identification and compare it with other reference material. | ||
| In the laboratory | Confirm the identity of each species using a stereo microscope and a light-microscope, and different floras and identification keys. | |
| Clean the fresh shoots/thalli under a stereo microscope to remove all non-target materials (e.g., other bryophyte species, vascular fragments, soil, and litter). | Try to preserve the structure of the patch as much as possible. Try to work fast in order to avoid undue evaporation. | |
| Weigh each sample on an electronic scale, in order to obtain the field weight. | This step is optional. | |
| Submerse each replicate in water to reach the full turgor (c. 12 consecutive hours). | ||
| Leave the hydrated samples over a structure of 4–6 wires, assembled 15–20 cm from the bench, and let the samples drip the excess water. | Samples should keep the field structure, as much as possible. | |
| Weigh the samples only when plants drip less than one drop per minute. This is the saturated weight (Ms), which accounts for all the water retained in the external capillarity spaces, as bryophytes can do in their colonies. | ||
| Gently blot the samples using absorbent paper to remove external water. No droplets of water should be seen on the shoots. | It is also possible to centrifuge the samples, however Santarius (1994) affirms that blotting generally gives more reliable estimates of full-turgor water content than centrifuging. | |
| Weigh the blotted samples to obtain the internal water content weight (Ma) | ||
| Oven-dry each sample for 48 h at 100 °C. | ||
| Weighed the oven-dried samples to determine specimens’ dry weight (Md). |
Appendix D
| Time (min) | Ha.SSB | Ba.SSB | Lc.SSB | Pb.SSB | Sg.SSB | Fa.SSB |
|---|---|---|---|---|---|---|
| 0 | ||||||
| 1 | 5.1 ± 2.04 | 4.1 ± 1.35 | 8.8 ± 3.42 | 11.3 ± 5.28 | 5.1 ± 1.04 | 4.2 ± 0.81 |
| 3 | 8.1 ± 2.89 | 8.9 ± 2.46 | 14.7 ± 4.67 | 18.2 ± 5.72 | 9.4 ± 1.72 | 10.2 ± 3.84 |
| 5 | 10.5 ± 3.14 | 12.0 ± 3.65 | 18.3 ± 5.05 | 24.5 ± 6.51 | 14.2 ± 2.78 | 14.6 ± 4.04 |
| 7 | 12.7 ± 3.74 | 15.7 ± 4.67 | 22.3 ± 6.00 | 28.2 ± 6.43 | 18.3 ± 3.24 | 18.3 ± 4.66 |
| 10 | 16.9 ± 3.98 | 20.1 ± 5.54 | 26.2 ± 6.76 | 31.8 ± 5.97 | 22.2 ± 3.36 | 22.3 ± 4.98 |
| 15 | 19.0 ± 4.14 | 22.8 ± 6.17 | 29.4 ± 6.67 | 35.1 ± 5.44 | 25.2 ± 3.72 | 25.5 ± 4.90 |
| 30 | 21.4 ± 4.45 | 26.6 ± 7.32 | 32.0 ± 6.46 | 39.1 ± 5.31 | 29.2 ± 3.42 | 30.0 ± 4.67 |
| 45 | 23.8 ± 4.51 | 30.2 ± 7.93 | 35.0 ± 6.41 | 43.3 ± 5.02 | 33.2 ± 3.38 | 33.9 ± 4.57 |
| 60 | 26.2 ± 4.97 | 32.4 ± 8.04 | 37.9 ± 6.12 | 46.9 ± 5.02 | 36.9 ± 2.98 | 37.3 ± 4.55 |
| 90 | 29.5 ± 5.73 | 36.0 ± 8.69 | 42.2 ± 5.49 | 51.7 ± 4.06 | 41.0 ± 2.70 | 38.4 ± 4.61 |
| 150 | 35.5 ± 6.60 | 43.1 ± 9.92 | 48.2 ± 5.07 | 53.1 ± 5.09 | 47.3 ± 2.27 | 43.6 ± 4.56 |
| 720 | 49.1 ± 11.64 | 66.9 ± 16.66 | 66.5 ± 5.84 | 69.4 ± 5.12 | 62.3 ± 6.06 | 59.4 ± 8.78 |
| 1080 | 70.2 ± 10.67 | 86.0 ± 14.40 | 88.0 ± 2.45 | 86.6 ± 3.80 | 81.1 ± 5.14 | 85.6 ± 6.91 |
| 1440 | 86.3 ± 5.82 | 92.8 ± 5.72 | 94.0 ± 0.82 | 93.2 ± 1.30 | 92.3 ± 1.77 | 89.2 ± 4.99 |
| 2160 | 94.3 ± 4.13 | 96.3 ± 1.51 | 97.1 ± 0.56 | 96.4 ± 1.48 | 97.9 ± 0.38 | 90.9 ± 4.40 |
| 2880 | 96.3 ± 1.73 | 96.4 ± 1.35 | 96.8 ± 0.63 | 96.2 ± 1.47 | 98.0 ± 0.35 | 90.3 ± 4.33 |
| 3600 | 96.7 ± 1.28 | 96.4 ± 1.31 | 97.1 ± 0.57 | 96.3 ± 1.49 | 98.0 ± 0.33 | 90.3 ± 4.42 |
| Time (min) | Ss.PL | Pc.SSB | Cb.FS | Cs.SSB | Tb.FS | Ip.PL | Mh.PL | Tt.PL |
|---|---|---|---|---|---|---|---|---|
| 0 | ||||||||
| 1 | 2.5 ± 1.52 | 0.8 ± 0.27 | 3.6 ± 0.48 | 4.2 ± 1.24 | 3.6 ± 2.09 | 3.4 ± 0.63 | 3.9 ± 1.14 | 11.8 ± 3.13 |
| 3 | 2.7 ± 1.60 | 1.5 ± 0.49 | 6.3 ± 1.14 | 8.0 ± 1.10 | 7.8 ± 3.33 | 6.7 ± 0.84 | 6.9 ± 1.62 | 13.6 ± 3.44 |
| 5 | 6.3 ± 0.52 | 2.1 ± 0.70 | 8.6 ± 1.27 | 11.3 ± 0.67 | 9.6 ± 2.80 | 9.0 ± 0.95 | 9.0 ± 2.32 | 20.7 ± 4.72 |
| 7 | 9.0 ± 0.93 | 2.7 ± 0.89 | 11.6 ± 1.83 | 14.3 ± 1.17 | 11.6 ± 2.83 | 11.6 ± 1.03 | 11.2 ± 2.64 | 26.1 ± 5.33 |
| 10 | 12.2 ± 1.37 | 3.5 ± 1.07 | 13.4 ± 1.04 | 16.4 ± 1.25 | 13.5 ± 2.86 | 15.3 ± 2.13 | 13.3 ± 2.79 | 31.3 ± 6.00 |
| 15 | 12.9 ± 1.08 | 4.7 ± 1.44 | 15.8 ± 1.71 | 18.8 ± 1.27 | 15.1 ± 3.16 | 19.3 ± 2.40 | 15.3 ± 2.89 | 36.4 ± 6.83 |
| 30 | 15.3 ± 1.13 | 6.9 ± 2.16 | 18.2 ± 1.62 | 21.8 ± 1.30 | 17.3 ± 3.66 | 23.6 ± 2.58 | 17.7 ± 2.87 | 42.4 ± 8.13 |
| 45 | 17.4 ± 1.12 | 9.0 ± 2.75 | 19.8 ± 1.42 | 24.3 ± 1.28 | 18.9 ± 3.68 | 27.8 ± 2.60 | 20.6 ± 2.89 | 51.0 ± 9.29 |
| 60 | 19.7 ± 1.36 | 11.1 ± 3.40 | 21.9 ± 1.11 | 26.9 ± 1.26 | 20.9 ± 3.51 | 32.7 ± 3.17 | 23.3 ± 2.91 | 59.9 ± 10.9 |
| 90 | 22.1 ± 1.74 | 14.8 ± 4.47 | 23.7 ± 0.90 | 30.2 ± 0.72 | 23.1 ± 3.74 | 39.6 ± 3.23 | 25.3 ± 3.06 | 72.1 ± 12.1 |
| 150 | 26.2 ± 2.14 | 21.3 ± 6.40 | 32.9 ± 2.35 | 36.9 ± 1.26 | 36.5 ± 3.42 | 43.8 ± 2.22 | 29.9 ± 3.05 | 82.3 ± 14.4 |
| 720 | 33.8 ± 6.49 | 82.2 ± 13.1 | 71.5 ± 7.38 | 59.0 ± 10.0 | 85.2 ± 3.41 | 89.4 ± 2.06 | 42.5 ± 4.65 | 90.7 ± 2.85 |
| 1080 | 51.6 ± 6.79 | 82.2 ± 12.9 | 80.9 ± 7.20 | 79.6 ± 8.69 | 91.8 ± 2.52 | 92.3 ± 1.59 | 65.9 ± 7.29 | 90.3 ± 2.83 |
| 1440 | 77.2 ± 5.23 | 82.7 ± 13.0 | 86.8 ± 6.14 | 91.3 ± 2.74 | 93.5 ± 1.50 | 92.3 ± 1.56 | 86.4 ± 2.93 | 91.0 ± 2.82 |
| 2160 | 90.0 ± 4.94 | 82.9 ± 13.0 | 94.3 ± 1.46 | 97.2 ± 1.02 | 93.9 ± 1.35 | 92.3 ± 1.55 | 96.6 ± 0.96 | 91.4 ± 2.81 |
| 2880 | 95.8 ± 2.65 | 82.3 ± 12.9 | 96.0 ± 0.97 | 96.9 ± 1.05 | 94.1 ± 1.28 | 93.0 ± 1.58 | 97.3 ± 0.79 | 90.6 ± 2.82 |
| 3600 | 95.9 ± 2.66 | 82.6 ± 13.1 | 96.1 ± 0.93 | 97.1 ± 1.03 | 94.4 ± 1.27 | 93.2 ± 1.58 | 97.2 ± 0.81 | 90.8 ± 2.85 |
References
- Vitt, D.H.; Crandall-Stotler, B.; Wood, A. Bryophytes: Survival in a dry world through tolerance and avoidance. In Plant Ecology and Evolution in Harsh Enviroments; Rajakaruna, N., Boyd, R.S., Harris, T.B., Eds.; Environmental Research Advances: New York, NY, USA, 2014; pp. 267–295. [Google Scholar]
- Bowen, E.J. The Mechanism of Water Conduction in the Musci considered in Relation to Habitat. II. Mosses growing in damp situations. Ann. Bot. 1933, 47, 635–661. [Google Scholar] [CrossRef]
- Proctor, M.C.F.; Tuba, Z. Poikilohydry and homoihydry: Antithesis or spectrum of possibilities? New Phytol. 2002, 156, 327–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kappen, L.; Valladares, F. Opportunistic Growth and Desiccation Tolerance: The Ecological Success of Poikilohydrous Autotrophs. Funct. Plant Ecol. 2007, 2, 7–66. [Google Scholar]
- Green, A.T.G.; Sancho, L.G.; Pintado, A. Ecophysiology of desiccation—Rehydration cycles in mosses and lichens. In Plant Desiccation Tolerance; Lüttge, U., Beck, E., Bartels, D., Eds.; Ecological Studies; Springer: Cham, Switzerland, 2011; pp. 89–120. [Google Scholar]
- Glime, J.M. Water Relations: Movement. In Bryophyte Ecology; Glime, J.M., Ed.; Physiological 7-2-1 Ecology; Ebook Sponsored by Michigan Technological University and the International Association of Bryologists; Last Updated 7 March 2017; Michigan Technological University: Houghton, MI, USA, 2017; Volume 1, Available online: www.bryoecol.mtu.edu (accessed on 30 December 2022).
- Tuba, Z.; Csintalan, Z.; Proctor, M.C.F. Photosynthetic responses of a moss, Tortula ruralis, ssp. ruralis, and the lichens Cladonia convoluta and C. furcata to water deficit and short periods of desiccation, and their ecophysiological significance: A baseline study at present-day CO2 concentrat. New Phytol. 1996, 133, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Proctor, M.C.F. Physiological ecology: Water relations, light and temperature responses, carbon balance. In Bryophyte Ecology; Smith, A.G.E., Ed.; Chapman and Hall: London, UK, 1982; pp. 333–381. [Google Scholar]
- Proctor, M.C.F. Physiological ecology. In Bryophyte Biology; Goffinet, B., Shaw, A.J., Eds.; Cambridge University Press: Cambridge, UK, 2009; pp. 237–268. [Google Scholar]
- Rundel, P.W. Water uptake by organs other than roots. In Physiological Plant Ecology II—Water Relations and Carbon Assimilation; Lange, O.L., Nobel, P.S., Osmond, C., Ziegler, C., Eds.; Encyclopedia of Plant Physiology: New York, NY, USA, 1982; pp. 111–128. [Google Scholar]
- Guerra, J.; Martinez-Sanchez, J.J.; Ros, R.M. On the degree of adaptation of the moss flora and vegetation in gypsiferous zones of the south-east Iberian Peninsula. J. Bryol. 1992, 17, 133–142. [Google Scholar] [CrossRef]
- Elumeeva, T.G.; Soudzilovskaia, N.A.; During, H.J.; Cornelissen, J.H.C. The importance of colony structure versus shoot morphology for the water balance of 22 subarctic bryophyte species. J. Veg. Sci. 2011, 22, 152–164. [Google Scholar] [CrossRef]
- Dilks, T.J.K.; Proctor, M.C.F. Photosynthesis Respiration and Water Content in Bryophytes. New Phytol. J. 1979, 82, 97–114. [Google Scholar] [CrossRef]
- Noakes, T.D.; Longton, R.E. Pathways of water movement in relation to structure and micromorphology in Antarctic mosses. Polarforschun 1988, 58, 125–138. [Google Scholar]
- Proctor, M.C.F. The bryophyte paradox: Tolerance of desiccation, evasion of drought. Plant Ecol. 2000, 151, 41–49. [Google Scholar] [CrossRef]
- Mauseth, J. Botany: An Introduction to Plant Biology, 6th ed.; Jones & Bartlett Learning: Burlington, MA, USA, 2016; p. 808. [Google Scholar]
- Proctor, M.C.F.; Nagy, Z.; Csintalan, Z.; Takács, Z. Water-content components in bryophytes: Analysis of pressure-volume curves. J. Exp. Bot. 1998, 49, 1845–1854. [Google Scholar] [CrossRef]
- Ingrouille, M. Adaptative growth forms: The limiting physical environment. In Diversity and Evolution of Land Plants; Chapman and Hall: Boca Raton, FL, USA, 1992. [Google Scholar]
- Longton, R.E. Growth and productivity of the moss Polytrichum alpestre Hoppe in Antarctic regions. In Antarctic Ecology; Holdgate, M.W., Ed.; Academic Press: London, UK, 1970; pp. 818–837. [Google Scholar]
- Longton, R.E. Studies on growth, reproduction and population ecology in relation to microclimate in the bipolar moss Polytrichum alpestre Hoppe. Bryologist 1979, 82, 325–367. [Google Scholar] [CrossRef]
- Rundel, P.W.; Lange, O.L. Water relations and photosynthetic response of a desert moss. Flora 1980, 169, 329–335. [Google Scholar] [CrossRef]
- Hietz, P. Ecology and ecophysiology of epiphytes in tropical montane cloud forests. In Tropical Montane Cloud Forests: Science for Conservation and Management; Bruijnzeel, L.A., Scatena, F.F.N., Hamilton, L.S., Eds.; Cambridge University Press: New York, NY, USA, 2010; pp. 67–71. [Google Scholar]
- Hajek, T.; Beckett, R.P. Effect of Water Content Components on Desiccation and Recovery in Sphagnum Mosses. Ann. Bot. 2008, 101, 165–173. [Google Scholar] [CrossRef] [Green Version]
- Schofield, W.B. Ecological significance of morphological characters in the moss gametophyte. Bryologist 1981, 84, 149–165. [Google Scholar] [CrossRef]
- Zotz, G.; Schweikert, A.; Jetz, W.; Westerman, H. Water relations and carbon gain are closely related to cushion size in the moss Grimmia pulvinata. New Phytol. 2000, 148, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Fowbert, J.A. An experimental study of growth in relation to morphology and shoot water content in maritime Antarctic mosses. New Phytol. 1996, 133, 363–373. [Google Scholar] [CrossRef]
- Robroek, B.J.M.; Schouten, M.G.C.; Limpens, J.; Berendse, F.; Poorter, H. Interactive effects of water table and precipitation on net CO 2 assimilation of three co-occurring Sphagnum mosses differing in distribution above the water table. Glob. Chang. Biol. 2009, 15, 680–691. [Google Scholar] [CrossRef]
- Veneklaas, E.J.; Zagt, R.J.; Leerdam, A.; Ek, R.; Broekhoven, A.J.; Genderen, M. Hydrological properties of the epiphyte mass of a montane tropical rain forest, Colombia. Vegetatio 1990, 89, 183–192. [Google Scholar] [CrossRef]
- Zhang, Y.; Peña-Arancibia, J.L.; McVicar, T.R.; Chiew, F.H.S.; Vaze, J.; Liu, C.; Lu, X.; Zheng, X.; Wang, Y.; Liu, Y.; et al. Multi-decadal trends in global terrestrial evapotranspiration and its components. Sci. Rep. 2016, 6, 19124. [Google Scholar] [CrossRef] [Green Version]
- Oliver, M.J. Desiccation tolerance in vegetative plant cells. Physiol. Plant. 1996, 97, 779–787. [Google Scholar] [CrossRef]
- Köhler, L.; Tobón, C.; Frumau, K.F.A.; Bruijnzeel, L.A. Biomass and water storage dynamics of epiphytes in old-growth and secondary montane cloud forest stands in Costa Rica. Plant Ecol. 2007, 193, 171–184. [Google Scholar] [CrossRef]
- DROTRH–IA. Plano Regional da Água. Relatório Técnico; Versão Para Consulta Pública: Ponta Delgada, Portugal, 2001. [Google Scholar]
- Cruz, J.V.; Pacheco, D.; Costa, S.; Melo, C.; Cymbron, R.; Nogueira, R.; Brito, A.G. Implementation of the Water Framework Directive in an Outermost EU Region: The Case of Azores Archipelago. Open Hydrol. J. 2012, 6, 1–14. [Google Scholar] [CrossRef]
- Rodrigues, F.C. Hidrogeologia da Ilha Terceira—Contributo Para o Seu Conhecimento. Ph.D Thesis, Universidade dos Açores, Angra do Heroísmo, Portugal, 1993. [Google Scholar]
- PGRH. Plano de Gestão da Região Hidrográfica dos Açores (RH9) 2016–2021; Direção Regional do Ambiente—RAA: Horta, Portugal, 2016. [Google Scholar]
- Harter, D.E.; Irl, S.D.; Seo, B.; Steinbauer, M.J.; Gillespie, R.G.; Triantis, K.A.; Fernández–Palacios, J.M.; Beierkuhnlein, C. Impacts of global climate change on the floras of oceanic islands–projections, implications and current knowledge. Perspect. Plant Ecol. Evol. Syst. 2015, 17, 160–183. [Google Scholar] [CrossRef]
- Patiño, J.; Mateo, R.G.; Zanatta, F.; Marquet, A.; Aranda, S.C.; Borges, P.A.V.; Dirkse, G.; Gabriel, R.; Gonzalez-Mancebo, J.M.; Guisan, A.; et al. Climate threat on the Macaronesian endemic bryophyte flora. Sci. Rep. 2016, 6, 29156. [Google Scholar] [CrossRef] [Green Version]
- Gabriel, R.; Sjögren, E.; Schumacker, R.; Sérgio, C.; Aranda, S.; Claro, D.; Homem, N.; Martins, B. List of bryophytes (Anthocerotophyta, Marchantiophyta, Bryophyta). In A List of the Terrestrial and Marine Biota from the Azores; Borges, P.A.V., Costa, A., Cunha, R., Gabriel, R., Gonçalves, V., Martins, A.F., Melo, I., Parente, M., Raposeiro, P., Rodrigues, P., et al., Eds.; Princípia: Cascais, Portugal, 2010; pp. 99–115. [Google Scholar]
- Silva, L.; Moura, M.; Schäfer, H.; Rumsey, F.; Dias, E.F. List of Vascular Plants (Tracheobionta). In A List of the Terrestrial and Marine Biota from the Azores; Borges, P.A.V., Costa, A., Cunha, R., Gabriel, R., Gonçalves, V., Martins, A.F., Melo, I., Parente, M., Raposeiro, P., Rodrigues, P., et al., Eds.; Princípia: Cascais, Portugal, 2010; pp. 117–146. [Google Scholar]
- Proctor, M.C.F. Water-relations parameters of some bryophytes evaluated by thermocouple psychrometry. J. Bryol. 1999, 21, 263–270. [Google Scholar] [CrossRef]
- Hildenbrand, A.; Weis, D.; Madureira, P.; Marques, F.O. Recent plate re-organization at the Azores Triple Junction: Evidence from combined geochemical and geochronological data on Faial, S. Jorge and Terceira volcanic islands. Lithos 2014, 210, 27–39. [Google Scholar] [CrossRef] [Green Version]
- Demografia. 2015. Available online: http://srea.azores.gov.pt/conteudos/Relatorios/lista_relatorios.aspx?idc=392&idsc=407&lang_id=1. (accessed on 16 October 2017).
- Forjaz, V.H. Atlas Básico dos Açores; Observatório Vulcanológico e Geotérmico dos Açores: Ponta Delgada, Portugal, 2004; p. 112. [Google Scholar]
- Decreto Legislativo Regional No. 11/2011/A de 20 de Abril da Assembleia Legislativa. Diário da República, 1.ª Série, No 78 (2011). Available online: http://www.azores.gov.pt/NR/rdonlyres/45B97553-5DFC-4856-841C-A61C5D98625D/530133/DLR_11_2011_A.pdf (accessed on 16 October 2017).
- Henriques, D.S.; Elias, R.B.; Coelho, M.C.; Hérnandez, R.H.; Pereira, F.; Gabriel, R. Long-Term monitoring across elevational gradients (III): Vascular plants on Terceira Island (Azores) transect. Arquipel. -Life Mar. Sci. 2017, 34, 1–20. [Google Scholar]
- Paton, J.A. The Liverwort Flora of the British Isles; Harley Books: Colchester, UK, 1999; p. 626. [Google Scholar]
- Smith, A.J.E. The Moss Flora of Britain and Ireland, 2nd ed.; Cambridge University Press: Cambridge, UK, 2004; p. 1012. [Google Scholar]
- Schumacker, R.; Váňa, J. Identification Keys to the Liverworts and Hornworts of Europe and Macaronesia (Distribution and Status), 2nd ed.; Sorus: Poznan, Poland, 2005; p. 171. [Google Scholar]
- Casas, C.; Brugués, M.; Cros, R.M.; Sérgio, C. Handbook of Mosses of the Iberian Peninsula and the Balearic Islands; Institut d’Estudis Catalans: Barcelona, Spain, 2006; p. 349. [Google Scholar]
- Casas, C.; Brugués, M.; Cros, R.M.; Sérgio, C.; Infante, M. Handbook of Liverworts of the Iberian Peninsula and the Balearic Islands; Institut d’Estudis Catalans: Barcelona, Spain, 2009; p. 177. [Google Scholar]
- Atherton, D.M.; Bosanquet, S.D.S.; Lawley, M. Mosses and Liverworts of Britain and Ireland: A Field Guide; British Bryological Society: Plymouth, MA, USA, 2010; p. 848. [Google Scholar]
- Hodgetts, N.G.; Söderström, L.; Blockeel, T.L.; Caspari, S.; Ignatov, M.S.; Konstantinova, N.A.; Lockhart, N.; Papp, B.; Schröck, C.; Sim-Sim, M.; et al. An annotated checklist of bryophytes of Europe, Macaronesia and Cyprus. J. Bryol. 2020, 42, 1–116. [Google Scholar] [CrossRef]
- Watkins, J.E., Jr.; Mack, M.C.; Sinclair, T.R.; Mulkey, S.S. Ecological and evolutionary consequences of desiccation tolerance in tropical fern gametophytes. New Phytol. 2007, 176, 708–717. [Google Scholar] [CrossRef]
- Santarius, K. Apoplasmic water fractions and osmotic water potentials at full turgidity of some Bryidae. Planta 1994, 193, 32–37. [Google Scholar] [CrossRef]
- Turner, N.C. Techniques and experimental approaches for the measurement of plant water status. Plant Soil 1981, 58, 339–366. [Google Scholar] [CrossRef]
- Glime, J.M. Water Relations: Plant Strategies. In Bryophyte Ecology; Glime, J.M., Ed.; Physiological 7-3-1 Ecology; Ebook Sponsored by Michigan Technological University and the International Association of Bryologists; Last Updated 7 March 2017; Michigan Technological University: Houghton, MI, USA, 2017; Volume 1, Available online: www.bryoecol.mtu.edu (accessed on 30 December 2022).
- Jauhiainen, J.; Silvola, J.; Vasander, H. Effects of increased carbon dioxide and nitrogen supply on mosses. In Bryology for the Twenty-First Century; Bates, J.W., Ashton, N.W., Duckett, J.G., Eds.; British Bryological Society: London, UK, 1998; pp. 343–360. [Google Scholar]
- Gabriel, R. Ecophysiology of Azorean Forest Bryophytes. Ph.D. Thesis, Imperial College of Science, Technology and Medicine, University of London, London, UK, 2000. [Google Scholar]
- Smith, R.I.L. Aspects of Cryptogam Water Relations at a Continental Antarctic Site. Polarforschung 1988, 58, 139–153. [Google Scholar]
- Núñez-Olivera, E.; Martínez-Abaigar, J.; Tomás, R.; Beaucourt, N.; Arróniz-Crespo, M. Influence of temperature on the effects of artificially enhanced UV-B radiation on aquatic bryophytes under laboratory conditions. Photosynthetica 2004, 42, 201–212. [Google Scholar] [CrossRef]
- Vanderpoorten, A.; Goffinet, B. Introduction to Bryophytes, 1st ed.; Cambridge University Press: Cambridge, UK, 2009; p. 313. [Google Scholar]
- Rydin, H.; Gunnarsson, U.; Sundberg, S. The role of Sphagnum in peatland development and persistence. In Boreal Peatland Ecosystems (Ecological Studies); Vitt, D.H., Wieder, R.K., Eds.; Springer: Berlin, Germany, 2006; Volume 188, pp. 49–65. [Google Scholar]
- Tuba, Z.; Slack, N.G.; Stark, L.R. Bryophyte Ecology and Climate Change; Cambridge University Press: New York, NY, USA, 2011; p. 528. [Google Scholar]
- Hayward, P.M.; Clymo, R.S. Profiles of Water Content and Pore Size in Sphagnum and Peat, and their Relation to Peat Bog Ecology. Proc. R. Soc. B Biol. Sci. 1982, 215, 299–325. [Google Scholar]
- Glime, J.M. Water Relations: Physiological Adaptations. In Bryophyte Ecology; Glime, J.M., Ed.; 7-5-1 Physiological Ecology; Ebook Sponsored by Michigan Technological University and the International Association of Bryologists; Last Updated 7 March 2017; Michigan Technological University: Houghton, MI, USA, 2017; Volume 1, Available online: www.bryoecol.mtu.edu (accessed on 30 December 2022).
- Schipperges, B.; Rydin, H. Response of photosynthesis of Sphagnum species from contrasting microhabitats to tissue water content and repeated desiccation. New Phytol. 1998, 140, 677–684. [Google Scholar] [CrossRef]
- Noailles, M.C. Les frontières des plantes vasculaires. Bull. La Société Bot. Fr. 1987, 134, 53–61. [Google Scholar] [CrossRef] [Green Version]
- Hébant, C. The Conducting Tissues of Bryophytes; J. Cramer: Vaduz, Liechtenstein, 1977. [Google Scholar]
- Bayfield, N.G. Notes on water relations of Polytrichum commune Hedw. J. Bryol. 1973, 7, 607–617. [Google Scholar] [CrossRef]
- Sollows, M.C.; Frego, K.A.; Norfolk, C. Recovery of Bazzania trilobata following desiccation. Bryologist 2001, 104, 421–429. [Google Scholar] [CrossRef]
- Ah-Peng, C.; Cardoso, A.W.; Flores, O.; West, A.; Wilding, N.; Strasberg, D.; Hedderson, T.A.J. The role of epiphytic bryophytes in interception, storage, and the regulated release of atmospheric moisture in a tropical montane cloud forest. J. Hydrol. 2017, 548, 665–673. [Google Scholar] [CrossRef]
- Cleavitt, N.L.; Klima, K.L.; Fahey, T.J. The role of water availability in seasonal growth of Bazzania trilobata on boulders in an eastern hemlock stand. Bryologist 2007, 110, 295–308. [Google Scholar] [CrossRef]
- Cary, G.; Lindenmayer, D.; Dovers, S. Australia Burning: Fire Ecology. Policy and Management Issues; CSIRO Publishing: Clayton, Australia, 2003; p. 269. [Google Scholar]
- Song, L.; Zhang, Y.J.; Chen, X.; Li, S.; Lu, H.Z.; Wu, C.S.; Tan, Z.H.; Shi, X.M. Water relations and gas exchange of fan bryophytes and their adaptations to microhabitats in an Asian subtropical montane cloud forest. J. Plant Res. 2015, 128, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Heijmans, M.M.P.D.; Arp, W.J.; Chapin, F.S. Controls on moss evaporation in a boreal black spruce forest. Glob. Biogeochem. Cycles 2004, 18, 1–8. [Google Scholar] [CrossRef]
- Magdefrau, K. Life forms of Bryophytes. Bryophyt. Ecol. 1982, 2, 45–58. [Google Scholar]
- Rixen, C.; Mulder, C.P.H. Improved water retention links high species richness with increased productivity in arctic tundra moss communities. Oecologia 2005, 146, 287–299. [Google Scholar] [CrossRef]
- Frahm, J.P. Manual of Tropical Bryology. Trop. Bryol. 2003, 23, 1–200. [Google Scholar]





| Div. | Class | Order | Family | Species | Colonization Status | Sampling Sites | Life Forms |
|---|---|---|---|---|---|---|---|
| Marchantiophyta | |||||||
| Jungermanniopsida | |||||||
| Jungermanniales | Herbertaceae | Herbertus azoricus (Steph.) P.W.Richards | END | SB | turfs | ||
| Lepidoziaceae | Bazzania azorica H.Buch and Perss. | END | SB | wefts | |||
| Lepidozia cupressina (Sw.) Lindenb. Subsp. cupressina | NAT | SB | wefts | ||||
| Plagiochilaceae | Plagiochila bifaria (Sw.) Lindenb. | NAT | PL, SB | turfs | |||
| Scapaniaceae | Scapania gracilis Lindb. | NAT | PL, SB | wefts | |||
| Porellales | Frullaniaceae | Frullania acicularis Hentschel and von Konrat | NAT | FS, PL, SB | mats | ||
| Bryophyta | |||||||
| Sphagnopsida | |||||||
| Sphagnales | Sphagnaceae | Sphagnum subnitens Russow and Warnst. | NAT | PL | turfs | ||
| Polytrichopsida | SB | ||||||
| Polytrichales | Polytrichaceae | Polytrichum commune Hedw. | NAT | turfs | |||
| Bryopsida | |||||||
| Dicranales | Leucobryaceae | Campylopus brevipilus Bruch and Schimp. | NAT | FS | cushions | ||
| Campylopus shawii Wilson | NAT | SB | turfs | ||||
| Pottiaceae | Trichostomum brachydontium Bruch | NAT | FS | turfs | |||
| Hypnales | Lembophyllaceae | Isothecium prolixum (Mitt.) M.Stech, Sim-Sim, Tangney and D.Quandt | MAC | PL | wefts | ||
| Myuriaceae | Myurium hochstetteri (Schimp.) Kindb. | NAT | PL | mats | |||
| Thuidiaceae | Thuidium tamariscinum (Hedw.) Schimp. | NAT | PL | wefts | |||
| Div. | Class | Species | AWC ± StD (%) | wCint. ± StD (%) | wCext. ± StD (%) |
|---|---|---|---|---|---|
| Marchantiophyta | |||||
| Jungermanniopsida | |||||
| Herbertus azoricus (Steph.) p.W.Richards | 804 ± 103 | 262 ± 20 | 2542 ± 099 | ||
| Bazzania azorica H.Buch and Perss. | 1067 ± 217 | 365 ± 65 | 702 ± 166 | ||
| Lepidozia cupressina (Sw.) Lindenb. | 1589 ± 215 | 340 ± 13 | 1250 ± 214 | ||
| Plagiochila bifaria (Sw.) Lindenb. | 1265 ± 200 | 301 ± 21 | 999 ± 232 | ||
| Scapania gracilis Lindb. | 1232 ± 163 | 322 ± 17 | 910 ± 164 | ||
| Frullania acicularis Hentschel and von Konrat | 1370 ± 293 | 322 ± 35 | 1048 ± 276 | ||
| Bryophyta | |||||
| Sphagnopsida | |||||
| Sphagnum subnitens Russow and Warnst. | 5584 ± 471 | 567 ± 47 | 5017 ± 455 | ||
| Polytrichopsida | |||||
| Polytrichum commune Hedw. | 646 ± 104 | 250 ± 13 | 395 ± 98 | ||
| Bryopsida | |||||
| Campylopus brevipilus Bruch and Schimp. | 1761 ± 113 | 301 ± 11 | 1460 ± 109 | ||
| Campylopus shawii Wilson | 1727 ± 154 | 356 ± 12 | 1400 ± 213 | ||
| Trichostomum brachydontium Bruch | 1743 ± 111 | 286 ± 08 | 1457 ± 109 | ||
| Isothecium prolixum (Mitt.) M.Stech, Sim-Sim, Tangney and D.Quandt | 714 ± 95 | 282 ± 19 | 503 ± 253 | ||
| Myurium hochstetteri (Schimp.) Kindb. | 2201 ± 199 | 261 ± 12 | 1939 ± 197 | ||
| Thuidium tamariscinum (Hedw.) Schimp. | 1568 ± 313 | 293 ± 28 | 1275 ± 299 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coelho, M.C.M.; Gabriel, R.; Ah-Peng, C. Characterizing and Quantifying Water Content in 14 Species of Bryophytes Present in Azorean Native Vegetation. Diversity 2023, 15, 295. https://doi.org/10.3390/d15020295
Coelho MCM, Gabriel R, Ah-Peng C. Characterizing and Quantifying Water Content in 14 Species of Bryophytes Present in Azorean Native Vegetation. Diversity. 2023; 15(2):295. https://doi.org/10.3390/d15020295
Chicago/Turabian StyleCoelho, Márcia C. M., Rosalina Gabriel, and Claudine Ah-Peng. 2023. "Characterizing and Quantifying Water Content in 14 Species of Bryophytes Present in Azorean Native Vegetation" Diversity 15, no. 2: 295. https://doi.org/10.3390/d15020295
APA StyleCoelho, M. C. M., Gabriel, R., & Ah-Peng, C. (2023). Characterizing and Quantifying Water Content in 14 Species of Bryophytes Present in Azorean Native Vegetation. Diversity, 15(2), 295. https://doi.org/10.3390/d15020295