
Does a Rural-Urban Gradient Affect Beetle Assemblages in an Arid Ecosystem?

 , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
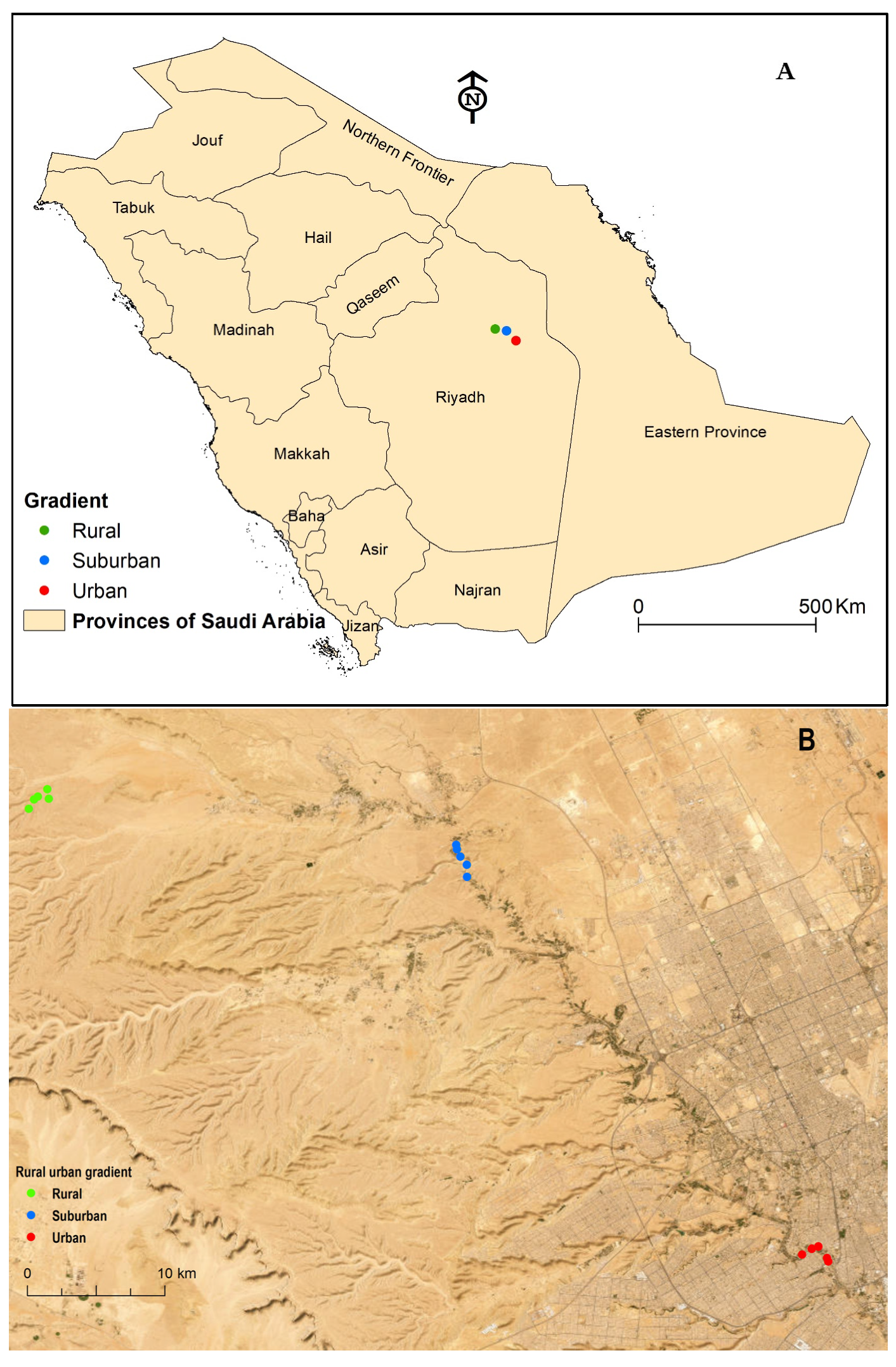
2.1. Study Area
2.2. Sampling Procedure
2.3. Data Analysis
3. Results
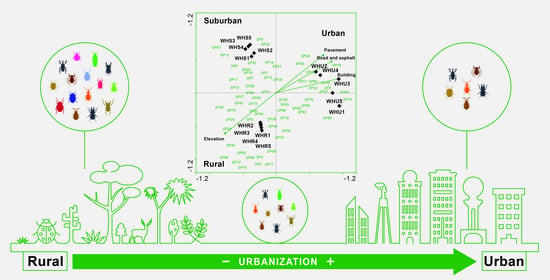
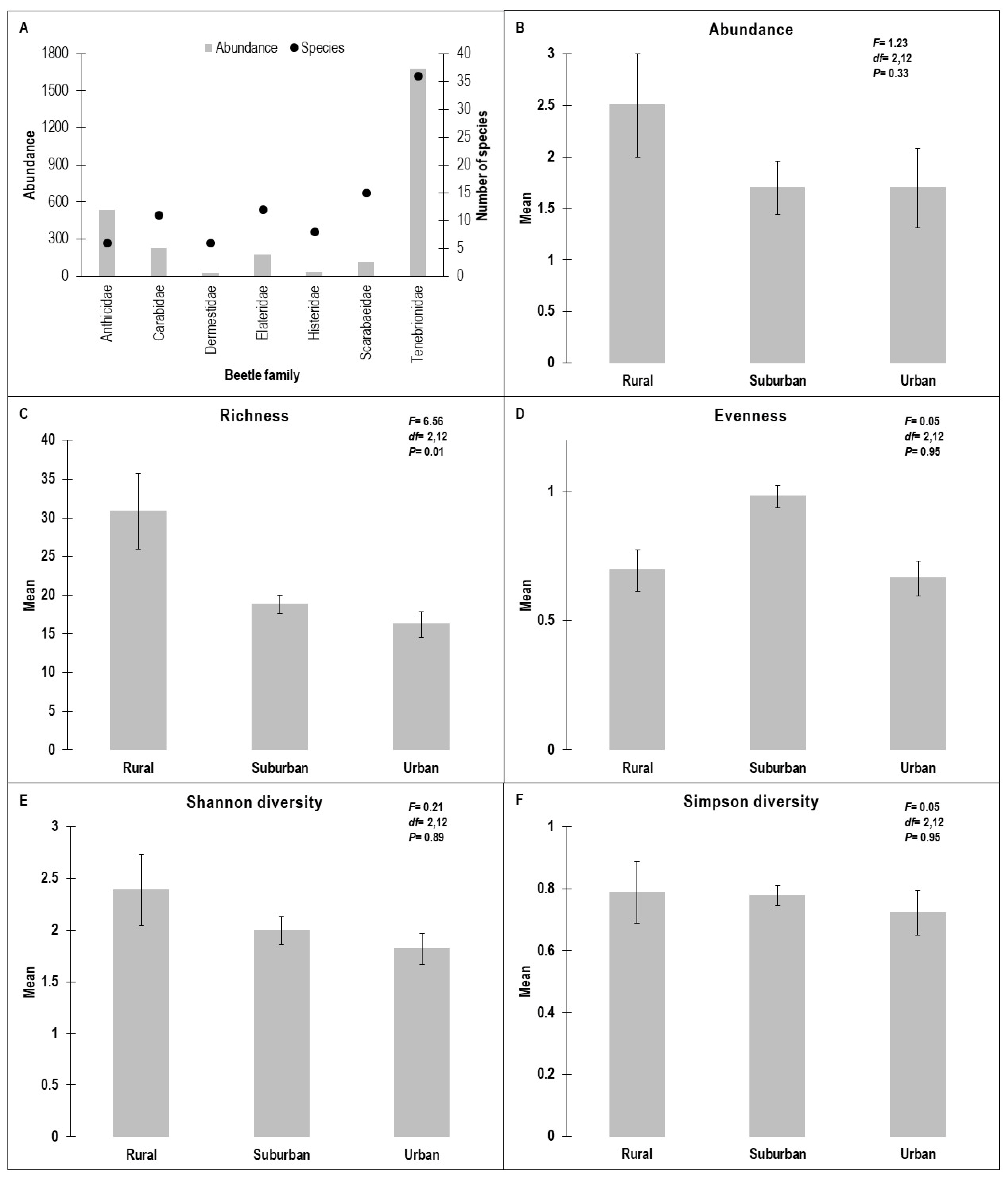
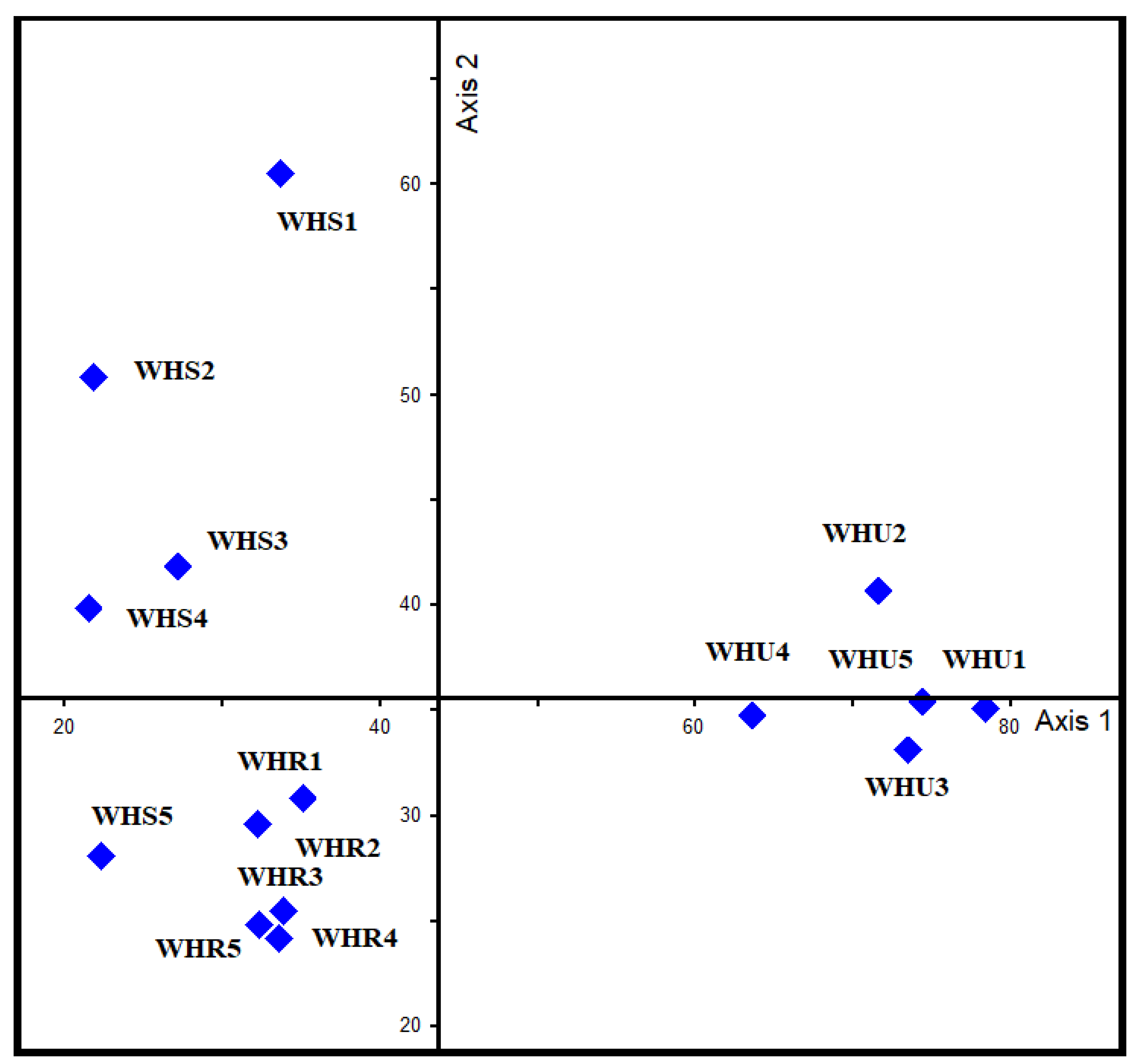
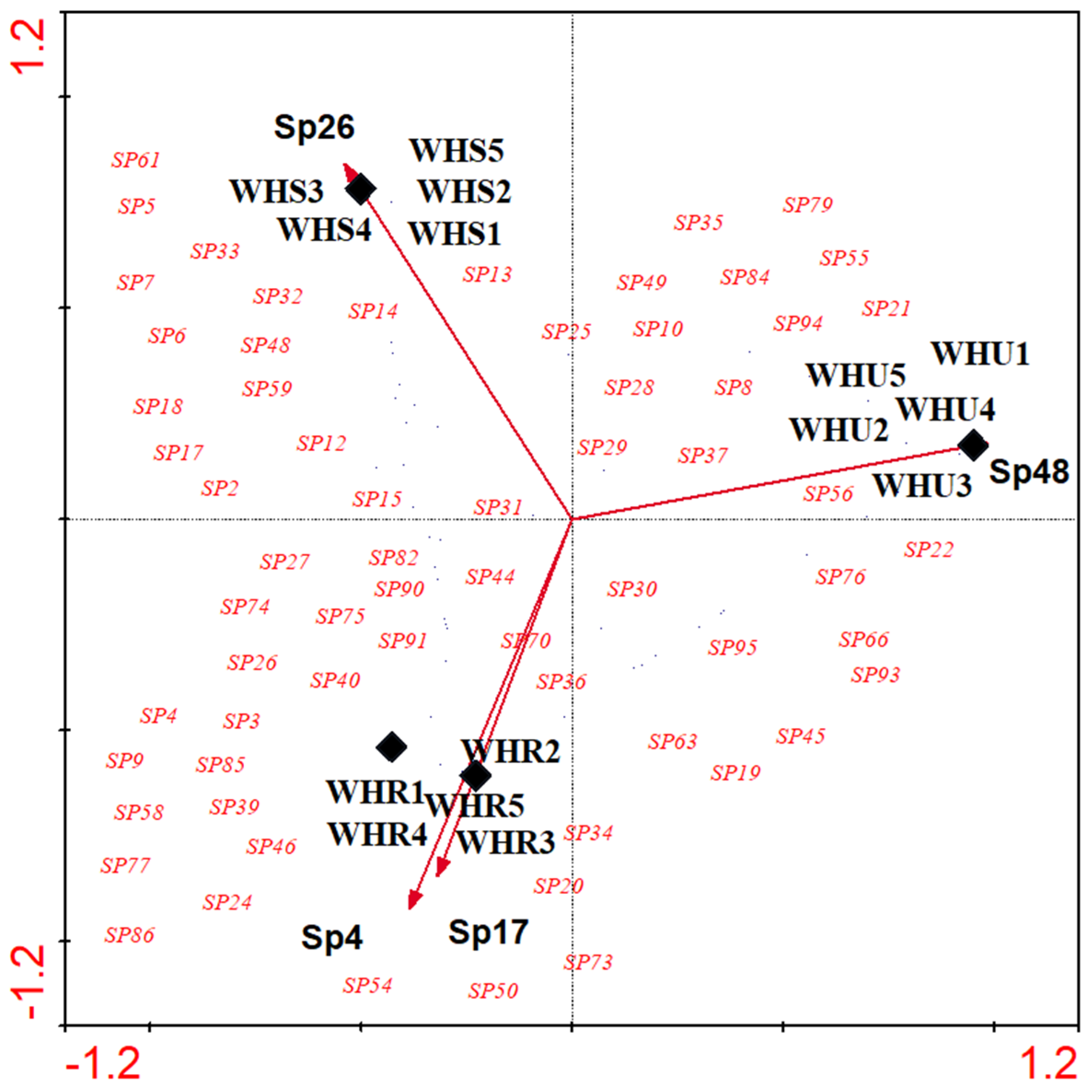
3.1. Beetle Diversity
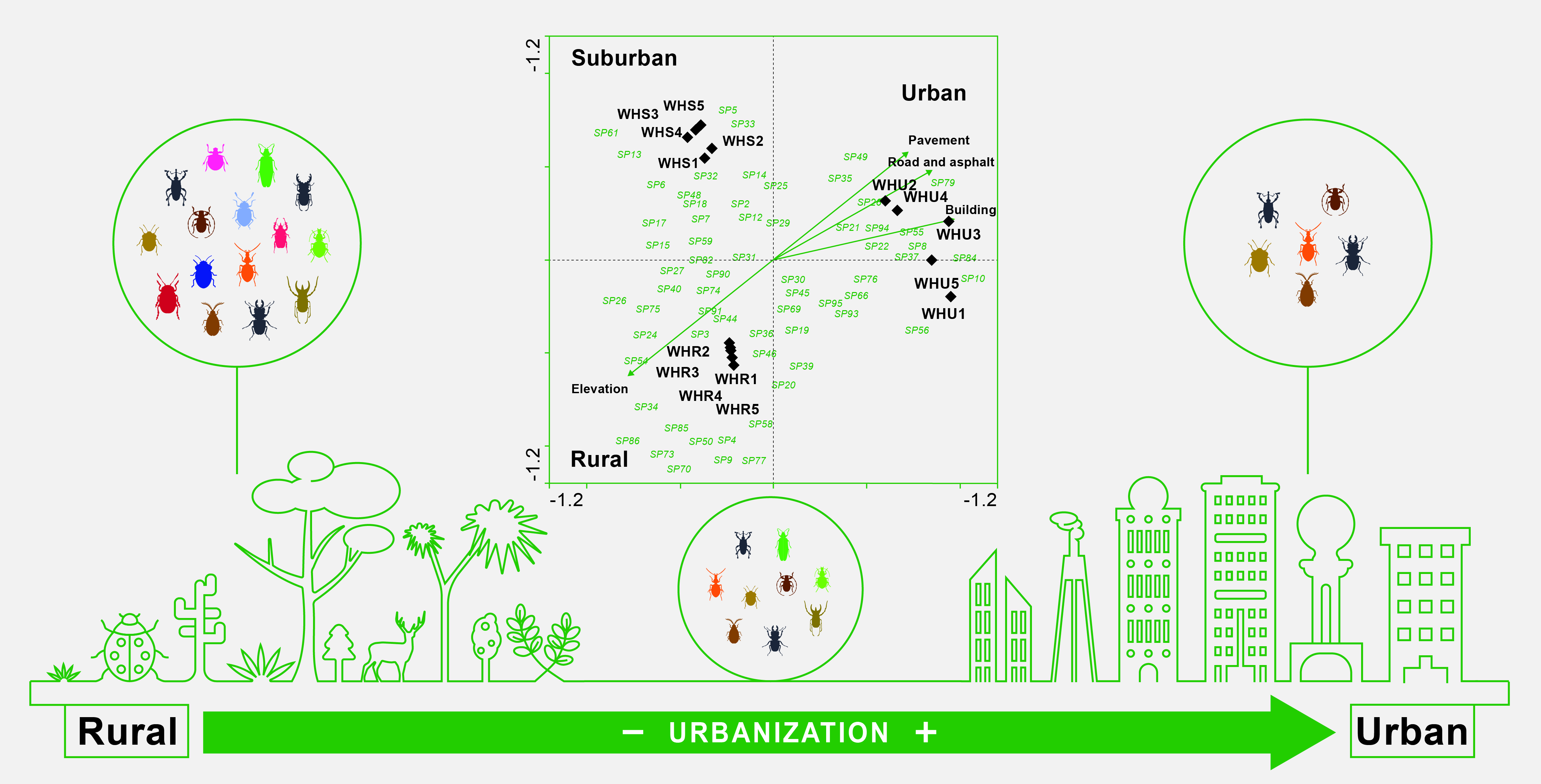
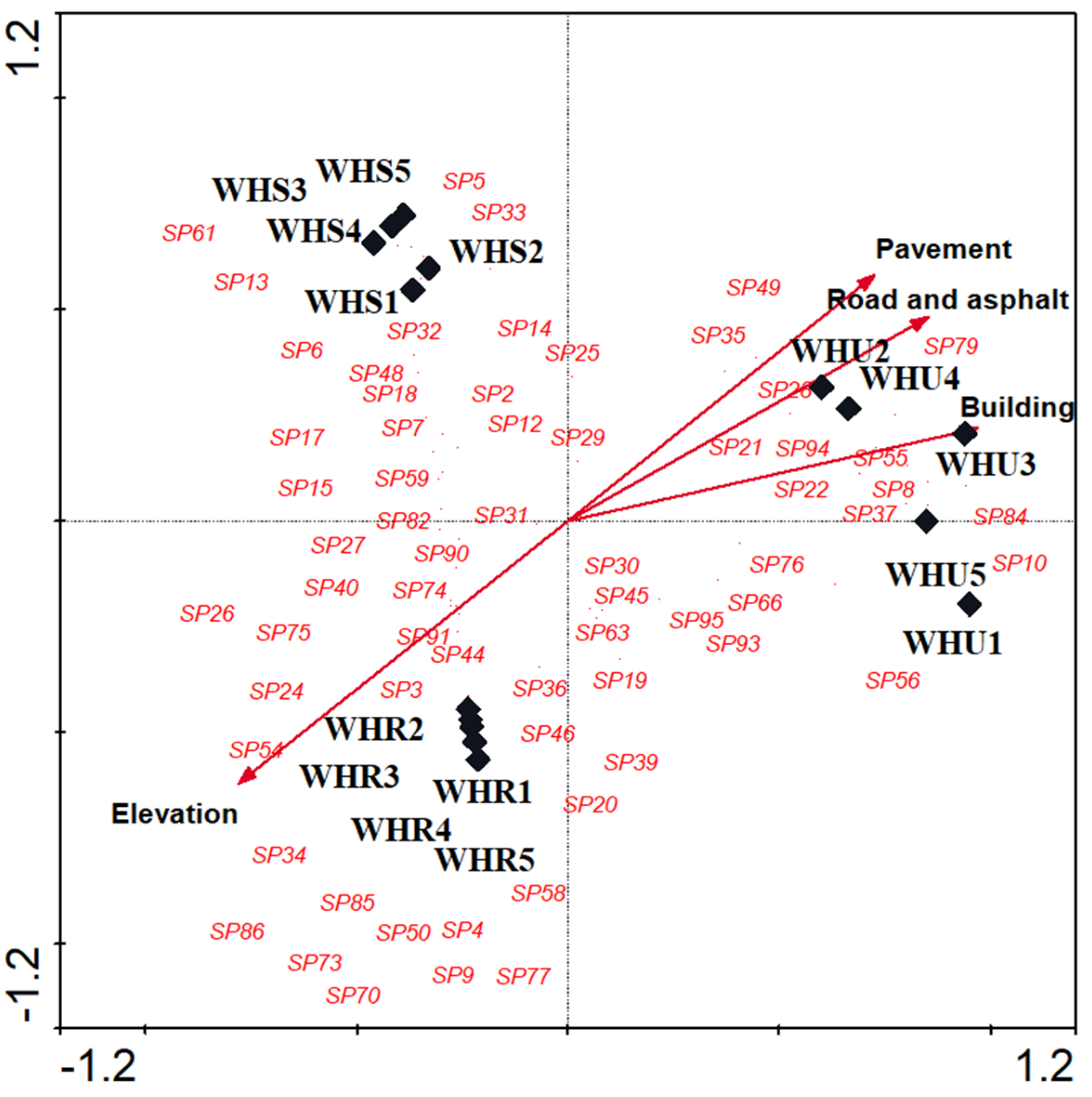
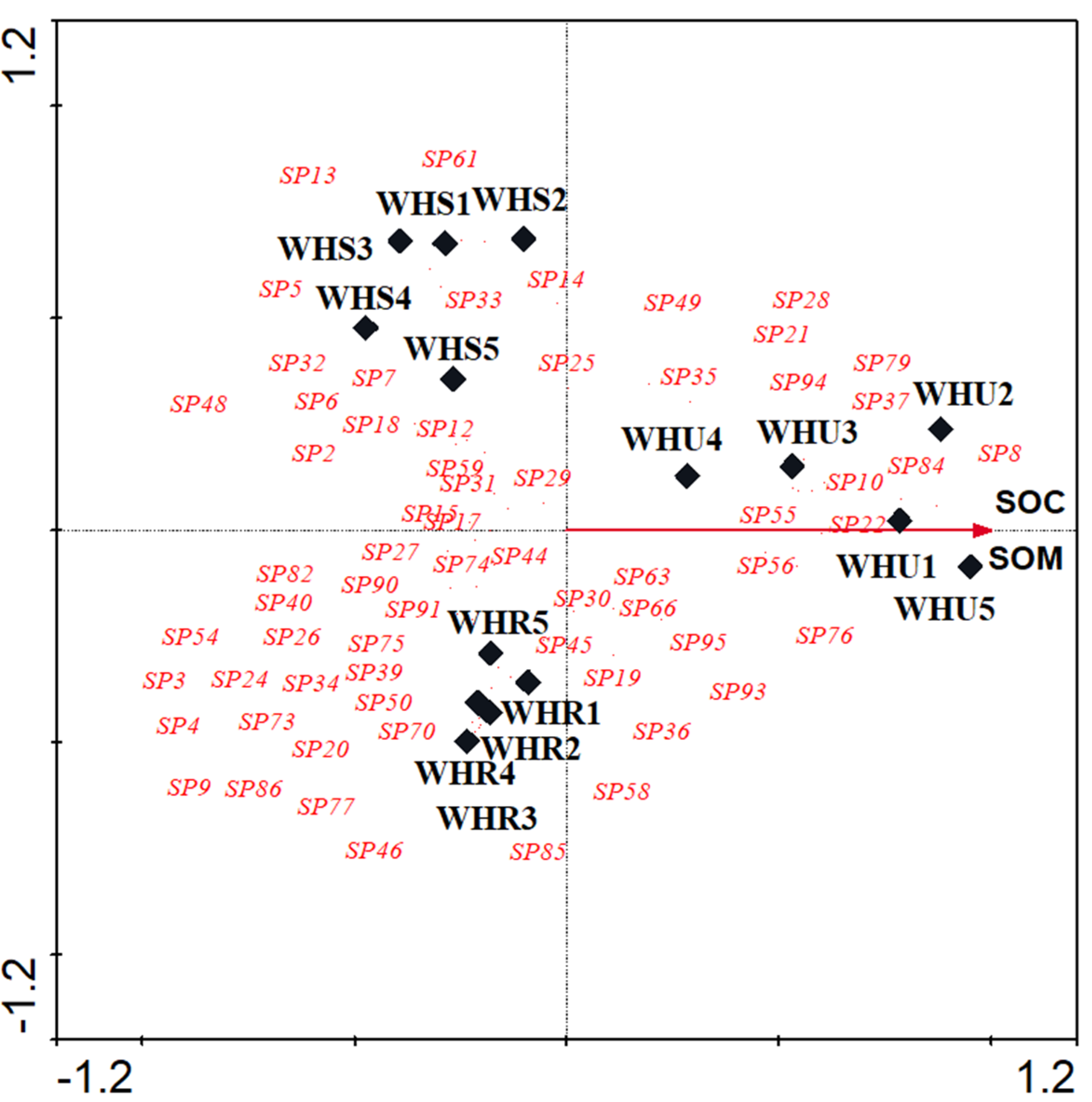
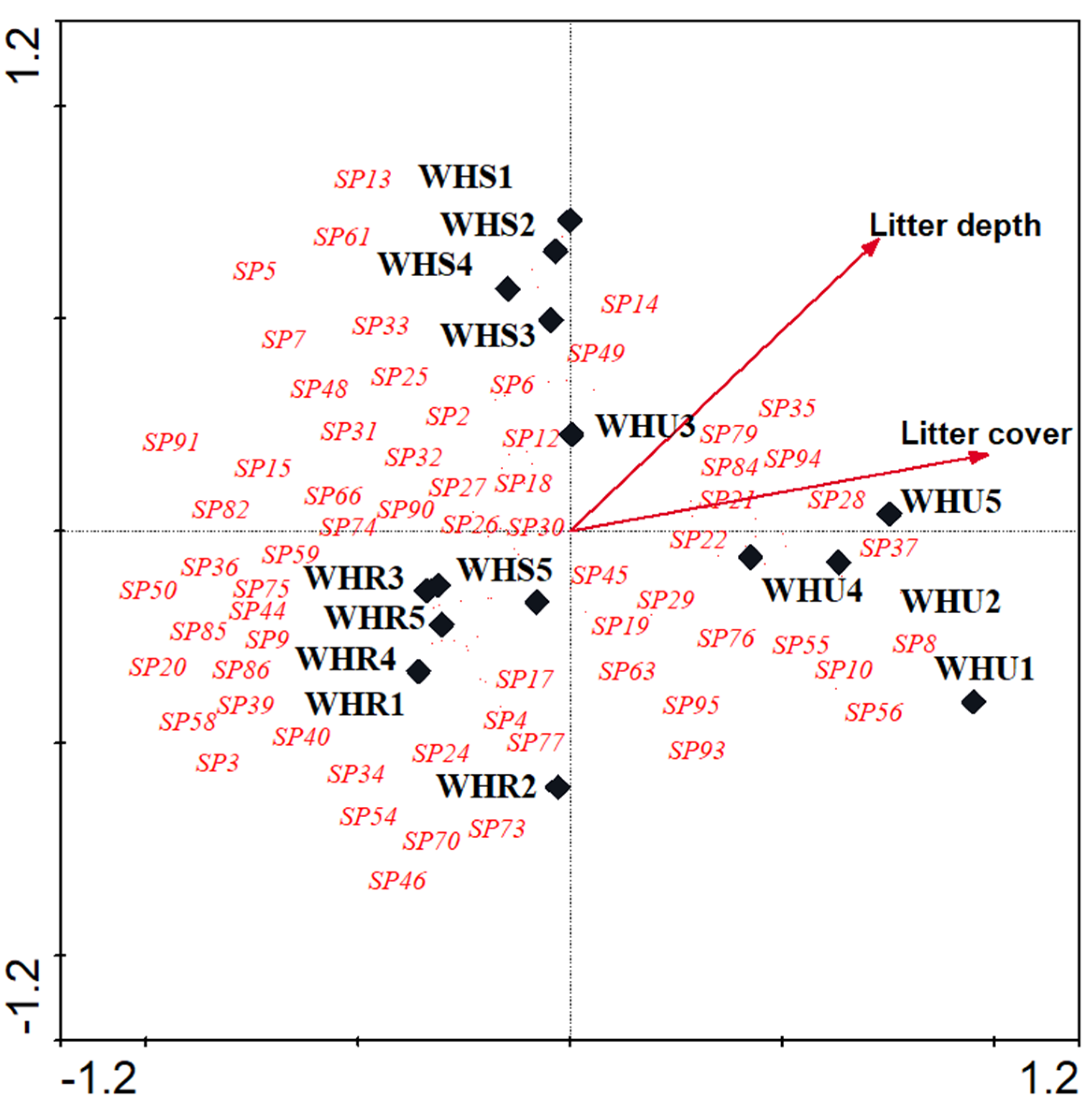
3.2. Species Composition
3.3. Beetle Indicator Species
4. Discussion
4.1. Beetle Diversity
4.2. Species Composition
4.3. Beetle Indicator Species
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Antrop, M. Changing patterns in the urbanized countryside of Western Europe. Landsc. Ecol. 2000, 15, 257–270. [Google Scholar] [CrossRef]
- Magura, T.; Lövei, G.L. Consequences of Urban Living: Urbanization and Ground Beetles. Curr. Landsc. Ecol. Rep. 2021, 6, 9–21. [Google Scholar] [CrossRef]
- Chaolin, G. Urbanization. In International Encyclopedia of Human Geography; Kobayashi, A., Ed.; Elsevier: Oxford, UK, 2020; Volume 14, pp. 141–153. [Google Scholar]
- United Nations. World Urbanization Prospects: The 2014 Revision: Highlights in P.D. Available online: https://population.un.org/wup/publications/files/wup2014-report.pdf (accessed on 11 April 2022).
- McDonnell, M.J.; Pickett, S.T.A.; Groffman, P.; Bohlen, P.; Pouyat, R.V.; Zipperer, W.C.; Parmelee, R.W.; Carreiro, M.M.; Medley, K. Ecosystem processes along an urban-to-rural gradient. Urban Ecosyst. 1997, 1, 21–36. [Google Scholar] [CrossRef]
- Piano, E.; De Wolf, K.; Bona, F.; Bonte, D.; Bowler, D.E.; Isaia, M.; Lens, L.; Merckx, T.; Mertens, D.; van Kerckvoorde, M.; et al. Urbanization drives community shifts towards thermophilic and dispersive species at local and landscape scales. Glob. Chang. Biol. 2017, 23, 2554–2564. [Google Scholar] [CrossRef] [Green Version]
- Kwon, T.-S.; Kim, Y.S.; Lee, S.W.; Park, Y.-S. Changes of soil arthropod communities in temperate forests over 10 years (1998–2007). J. Asia-Pacif. Entomol. 2016, 19, 181–189. [Google Scholar] [CrossRef]
- Parris, K.M. Ecology of Urban Environments, 1st ed.; Wiley-Blackwell: New York, NY, USA, 2016; p. 240. [Google Scholar]
- Parris, K.M.; Schneider, A. Impacts of Traffic Noise and Traffic Volume on Birds of Roadside Habitats. Ecol. Soc. 2009, 14, 29. [Google Scholar] [CrossRef] [Green Version]
- Rotholz, E.; Mandelik, Y. Roadside habitats: Effects on diversity and composition of plant, arthropod, and small mammal communities. Biodivers. Conserv. 2013, 22, 1017–1031. [Google Scholar] [CrossRef]
- Small, E.C.; Sadler, J.P.; Telfer, M.G. Carabid beetle assemblages on urban derelict sites in Birmingham, UK. J. Insect Conserv. 2002, 6, 233–246. [Google Scholar] [CrossRef]
- Tóthmérész, B.; Máthé, I.; Balázs, E.; Magura, T. Responses of carabid beetles to urbanization in Transylvania (Romania). Landsc. Urban Plan. 2011, 101, 330–337. [Google Scholar] [CrossRef] [Green Version]
- Piano, E.; Bona, F.; Isaia, M. Urbanization drivers differentially affect ground arthropod assemblages in the city of Turin (NW-Italy). Urban Ecosyst. 2020, 23, 617–629. [Google Scholar] [CrossRef]
- Al-Homaidan, A.A.; Al-Ghanayem, A.A.; Alkhalifa, A.H. Green algae as bioindicators of heavy metal pollution in Wadi Hanifah Stream, Riyadh, Saudi Arabia. Int. J. Water Resour. 2011, 1, 10–15. [Google Scholar]
- Asabere, S.B.; Zeppenfeld, T.; Nketia, K.A.; Sauer, D. Urbanization Leads to Increases in pH, Carbonate, and Soil Organic Matter Stocks of Arable Soils of Kumasi, Ghana (West Africa). Front. Environ. Sci. 2018, 6, 119. [Google Scholar] [CrossRef]
- Kark, S.; Iwaniuk, A.; Schalimtzek, A.; Banker, E. Living in the city: Can anyone become an ‘urban exploiter’? J. Biogeogr. 2007, 34, 638–651. [Google Scholar] [CrossRef]
- McKinney, M.L. Effects of urbanization on species richness: A review of plants and animals. Urban Ecosyst. 2008, 11, 161–176. [Google Scholar] [CrossRef]
- Knop, E. Biotic homogenization of three insect groups due to urbanization. Glob. Chang. Biol. 2016, 22, 228–236. [Google Scholar] [CrossRef]
- McKinney, M.L. Urbanization as a major cause of biotic homogenization. Biol. Conserv. 2006, 127, 247–260. [Google Scholar] [CrossRef]
- Pickett, S.T.A.; Cadenasso, M.L.; Grove, J.M.; Nilon, C.H.; Pouyat, R.V.; Zipperer, W.C.; Costanza, R. Urban Ecological Systems: Linking Terrestrial Ecological, Physical, and Socioeconomic Components of Metropolitan Areas. Annu. Rev. Ecol. Syst. 2001, 32, 127–157. [Google Scholar] [CrossRef] [Green Version]
- New, T.R. Insect Conservation and Urban Environments, 1st ed.; Springer: Cham, Switzerland, 2015; p. 244. [Google Scholar]
- Niemelä, J.; Kotze, D.J. Carabid beetle assemblages along urban to rural gradients: A review. Landsc. Urban Plan. 2009, 92, 65–71. [Google Scholar] [CrossRef]
- Bouchard, P.; Grebennikov, V.V.; Smith, A.B.T.; Douglas, H. Biodiversity of Coleoptera. In Insect Biodiversity: Science and Society; Foottit, R.G., Adler, P.H., Eds.; Wiley-Blackwell: West Sussex, UK, 2009; pp. 265–301. [Google Scholar]
- New, T.R. Beetles in Conservation, 1st ed.; Springer: Dordrecht, The Netherlands, 2010; p. 246. [Google Scholar]
- NCWCD. The National Strategy for Conservation of Biodiversity in the Kingdom of Saudi Arabia. Available online: http://www.cbd.int/doc/world/sa/sa-nbsap-01-en.pdf. (accessed on 11 April 2022).
- Aldryhim, Y.N.; Mills, C.W.; Aldawood, A.S. Ecological distribution and seasonality of darkling beetles (Coleoptera: Tenebrionidae) in the central region of Saudi Arabia. J. Arid Environ. 1992, 23, 415–422. [Google Scholar] [CrossRef]
- Al-Dawood, A.S.; Alahmed, A.M.; Kheir, S.M.; Hussein, S.M. Population dynamics of sandflies (Diptera: Psychodidae) in Hanifah valley, Riyadh, Saudi Arabia. Pak. J. Biol. Sci. 2004, 7, 464–467. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Dayem, M.S.; Sharaf, M.R.; Majer, J.D.; Al-Sadoon, M.K.; Aldawood, A.S.; Aldhafer, H.M.; Orabi, G.M. Ant diversity and composition patterns along the urbanization gradients in an arid city. J. Nat. Hist. 2021, 55, 2521–2547. [Google Scholar] [CrossRef]
- al-Asad, M.; Yavuz, Y. Wadi Hanifah Wetland. Available online: https://archive.archnet.org/sites/4458/publications/1206 (accessed on 23 December 2022).
- Samhouri, W. On Site Review Report: Wadi Hanifa Wetlands. Available online: http://archnet.org (accessed on 23 December 2022).
- Ter Braak, C.J. Canonical community ordination. Part I: Basic theory and linear methods. Ecoscience 1994, 1, 127–140. [Google Scholar] [CrossRef]
- Ter Braak, C.J. Canonical correspondence analysis: A new eigenvector technique for multivariate direct gradient analysis. Ecology 1986, 67, 1167–1179. [Google Scholar] [CrossRef] [Green Version]
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Ramalho, C.E.; Hobbs, R.J. Time for a change: Dynamic urban ecology. Trends Ecol. Evol. 2012, 27, 179–188. [Google Scholar] [CrossRef]
- Alhamid, A.A.; Alfayzi, S.A.; Hamadto, M.A. A sustainable water resources management plan for Wadi Hanifa in Saudi Arabia. J. King Saud Univ. Sci. 2007, 19, 209–221. [Google Scholar] [CrossRef]
- Al-Obaid, S.; Samraoui, B.; Thomas, J.; El-Serehy, H.A.; Alfarhan, A.H.; Schneider, W.; O’connell, M. An overview of wetlands of Saudi Arabia: Values, threats, and perspectives. Ambio 2017, 46, 98–108. [Google Scholar] [CrossRef] [Green Version]
- Faragalla, A.; Adam, E. Pitfall trapping of tenebrionid and carabid beetles (Coleoptera) in different habitats of the Central Region of Saudi Arabia. J. Appl. Entomol. 1985, 99, 466–471. [Google Scholar] [CrossRef]
- Abdel-Dayem, M.S.; Orabi, G.M.; Semida, F.M. Assessing the potential role of beetles as bioindicators in south Sinai, Egypt. In Proceedings of the Second International Conference of Entomological Society of Egypt, Cairo, Egypt, 8–11 December 2007. [Google Scholar]
- Piñero, F.; Tinaut, A.; Aguirre-Segura, A.; Miñano, J.; Lencina, J.; Ortiz-Sánchez, F.; Pérez-López, F. Terrestrial arthropod fauna of arid areas of SE Spain: Diversity, biogeography, and conservation. J. Arid Environ. 2011, 75, 1321–1332. [Google Scholar] [CrossRef]
- El Surtasi, E.I.; Semida, F.M.; Abdel-Dayem, M.S.; El Bokl, M.M. The threat of urbanization on beetle diversity in New Damietta City, Egypt. Nat. Sci. Sleep. 2012, 10, 15–23. [Google Scholar]
- Aldhafer, H.M.; Abdel-Dayem, M.S.; Aldryhim, Y.N.; Fadl, H.H.; El-Torkey, A.M.; Elgharbawy, A.A.; Setyaningrum, H. Diversity and composition of ground-dwelling beetle assemblages (Insecta: Coleoptera) in Rawdhat Khorim National Park, Kingdom of Saudi Arabia. J. Arid Environ. 2016, 127, 187–191. [Google Scholar] [CrossRef]
- Magura, T.; Tóthmérész, B.; Molnár, T. A species-level comparison of occurrence patterns in carabids along an urbanisation gradient. Landsc. Urban Plan. 2008, 86, 134–140. [Google Scholar] [CrossRef] [Green Version]
- Bennett, A.B.; Gratton, C. Measuring natural pest suppression at different spatial scales affects the importance of local variables. Environ. Entomol. 2012, 41, 1077–1085. [Google Scholar] [CrossRef] [Green Version]
- Ismail, O.K.; Bartholomew, A. Increased preference of darkling beetles Akis subtricostata Redtenbacher, 1850 and Trachyderma philistina Reiche and Saulcy, 1857 (Coleoptera, Tenebrionidae) for vegetation with increasing temperature. Bull. Iraq Nat. Hist. Mus. 2020, 16, 113–124. [Google Scholar] [CrossRef]
- Pickett, S.T.; Cadenasso, M.L.; Grove, J.M.; Boone, C.G.; Groffman, P.M.; Irwin, E.; Kaushal, S.S.; Marshall, V.; McGrath, B.P.; Nilon, C.H. Urban ecological systems: Scientific foundations and a decade of progress. J. Environ. Manage. 2011, 92, 331–362. [Google Scholar] [CrossRef]
- Qiu, J.; Yang, X.; Cao, B.; Chen, Z.; Li, Y. Effects of urbanization on regional extreme-temperature changes in China, 1960–2016. Sustainability 2020, 12, 6560. [Google Scholar] [CrossRef]
- Foster, C.W.; Kelly, C.; Rainey, J.J.; Holloway, G.J. Effects of urbanisation and landscape heterogeneity mediated by feeding guild and body size in a community of coprophilous beetles. Urban Ecosyst. 2020, 23, 1063–1077. [Google Scholar] [CrossRef]
- Park, J.-W.; Lee, C.M. Response of Ground Beetle (Coleoptera: Carabidae) Communities to Effect of Urbanization in Southern Osaka: An Analytical Approach Using GIS. Sustainability 2021, 13, 7134. [Google Scholar] [CrossRef]
- Magura, T.; Horváth, R.; Tóthmérész, B. Effects of urbanization on ground-dwelling spiders in forest patches, in Hungary. Landsc. Ecol. 2010, 25, 621–629. [Google Scholar] [CrossRef] [Green Version]
- Gaublomme, E.; Hendrickx, F.; Dhuyvetter, H.; Desender, K. The effects of forest patch size and matrix type on changes in carabid beetle assemblages in an urbanized landscape. Biol. Conserv. 2008, 141, 2585–2596. [Google Scholar] [CrossRef]
- Philpott, S.M.; Cotton, J.; Bichier, P.; Friedrich, R.L.; Moorhead, L.C.; Uno, S.; Valdez, M. Local and landscape drivers of arthropod abundance, richness, and trophic composition in urban habitats. Urban Ecosyst. 2014, 17, 513–532. [Google Scholar] [CrossRef] [Green Version]
- Vergnes, A.; Pellissier, V.; Lemperiere, G.; Rollard, C.; Clergeau, P. Urban densification causes the decline of ground-dwelling arthropods. Biodivers. Conserv. 2014, 23, 1859–1877. [Google Scholar] [CrossRef]
- Egerer, M.H.; Arel, C.; Otoshi, M.D.; Quistberg, R.D.; Bichier, P.; Philpott, S.M. Urban arthropods respond variably to changes in landscape context and spatial scale. J. Urban Ecol. 2017, 3, jux001. [Google Scholar] [CrossRef] [Green Version]
- Chiu, M.-C.; Chou, T.-Y.; Kuo, M.-H. Seasonal patterns of stream macroinvertebrate communities in response to anthropogenic stressors in monsoonal Taiwan. J. Asia-Pacif. Entomol. 2018, 21, 423–429. [Google Scholar] [CrossRef]
- Eversham, B.C.; Roy, D.B.; Telfer, M.G. Urban, Industrial and Other Manmade Sites as Analogues of Natural Habitats for Carabidae; Finnish Zoological and Botanical Publishing Board: Helsinki, Finland, 1996; pp. 149–156. [Google Scholar]
- Magura, T.; Tóthmérész, B.; Molnár, T. Changes in carabid beetle assemblages along an urbanisation gradient in the city of Debrecen, Hungary. Landsc. Ecol. 2004, 19, 747–759. [Google Scholar] [CrossRef]
- Zolotarev, M.P.; Belskaya, E.A. Ground-dwelling invertebrates in a large industrial city: Differentiation of recreation and urbanization effects. Contemp. Probl. Ecol. 2015, 8, 83–90. [Google Scholar] [CrossRef]
- Philpott, S.M.; Albuquerque, S.; Bichier, P.; Cohen, H.; Egerer, M.H.; Kirk, C.; Will, K.W. Local and Landscape Drivers of Carabid Activity, Species Richness, and Traits in Urban Gardens in Coastal California. Insects 2019, 10, 112. [Google Scholar] [CrossRef] [Green Version]
- Castro, A.V.; Porrini, D.P.; Lupo, S.; Cicchino, A.C. Minimal stories in Southeast Buenos Aires grasslands: Carabid beetle biodiversity throughout an urban-rural gradient. Urban Ecosyst. 2020, 23, 331–343. [Google Scholar] [CrossRef]
- Pywell, R.F.; James, K.L.; Herbert, I.; Meek, W.R.; Carvell, C.; Bell, D.; Sparks, T.H. Determinants of overwintering habitat quality for beetles and spiders on arable farmland. Biol. Conserv. 2005, 123, 79–90. [Google Scholar] [CrossRef]
- Deichsel, R. Species change in an urban setting—Ground and rove beetles (Coleoptera: Carabidae and Staphylinidae) in Berlin. Urban Ecosyst. 2006, 9, 161–178. [Google Scholar] [CrossRef]
- Elek, Z.; Lövei, G.L. Patterns in ground beetle (Coleoptera: Carabidae) assemblages along an urbanisation gradient in Denmark. Acta Oecol. 2007, 32, 104–111. [Google Scholar] [CrossRef]
- Do, Y.; Lineman, M. Effects of agricultural abandonment on carabid beetles in paddy fields. Balt. J. Coleopterol. 2012, 12, 65–75. [Google Scholar]
- Fusco, N.A.; Zhao, A.; Munshi-South, J. Urban forests sustain diverse carrion beetle assemblages in the New York City metropolitan area. Peer J. 2017, 5, e3088. [Google Scholar] [CrossRef] [Green Version]
- Work, T.T.; Shorthouse, D.P.; Spence, J.R.; Volney, W.J.A.; Langor, D. Stand composition and structure of the boreal mixedwood and epigaeic arthropods of the Ecosystem Management Emulating Natural Disturbance (EMEND) landbase in northwestern Alberta. Can. J. For. Res. 2004, 34, 417–430. [Google Scholar] [CrossRef]
- Orabi, G.M.; Semida, F.M.; Abdel-Dayem, M.S.; Sharaf, M.R.; Zalat, S.M. Diversity patterns of ants along an elevation gradient at St. Catherine Protectorate, South Sinai, Egypt. Zool. Middle East 2011, 54, 101–112. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.-L.; Li, F.-R.; Liu, C.-A.; Liu, Q.-J. Influences of shrub vegetation on distribution and diversity of a ground beetle community in a Gobi desert ecosystem. Biodivers. Conserv. 2012, 21, 2601–2619. [Google Scholar] [CrossRef]
- Kosewska, A.; Nietupski, M.; Damszel, M. Role of urban forests as a source of diversity of carabids (Coleoptera: Carabidae) in urbanised areas. Balt. J. Coleopterol. 2013, 13, 27–39. [Google Scholar]
- Belskaya, E.; Zolotarev, M.; Zinovyev, E. Carabidae assemblages in pine forests with different recreation regimes within and outside a megalopolis. Urban Ecosyst. 2020, 23, 27–38. [Google Scholar] [CrossRef]
- Niemelä, J.; Haila, Y.; Halme, E.; Lahti, T.; Pajunen, T.; Punttila, P. The distribution of carabid beetles in fragments of old coniferous taiga and adjacent managed forest. Ann. Zool. Fenn. 1988, 25, 107–119. [Google Scholar]
- Nietupski, M. Assemblages of epigeic Carabidae [Col.] in a peatbog nature reserve situated in an urban area. Pol. J. Nat. Sci. 2008, 23, 611–623. [Google Scholar] [CrossRef]
- Seibold, S.; Bässler, C.; Baldrian, P.; Reinhard, L.; Thorn, S.; Ulyshen, M.D.; Weiß, I.; Müller, J. Dead-wood addition promotes non-saproxylic epigeal arthropods but effects are mediated by canopy openness. Biol. Conserv. 2016, 204, 181–188. [Google Scholar] [CrossRef]
- Ouchtati, N.; Brandmayr, P.; Saouache, Y. A comparative analysis of the community of carabid beetles associated with two native xerophytic shrub species (Atriplex halimus L. and Artemisia herba alba L.) in the semi-arid zone of Algeria. Afr. Entomol. 2020, 28, 164–174. [Google Scholar] [CrossRef]
- Ariza, G.M.; Jácome, J.; Esquivel, H.E.; Kotze, D.J. Early successional dynamics of ground beetles (Coleoptera, Carabidae) in the tropical dry forest ecosystem in Colombia. ZooKeys 2021, 1044, 877–906. [Google Scholar] [CrossRef]
- Langor, D.; Pohl, G.; Hammond, H. A coarse-filter approach to conserving arthropod biodiversity in Canadian forests. Arthropods Can. For. Newsl. 2006, 2, 9–13. [Google Scholar]
- Bergeron, C.; Spence, J.; Volney, J. Landscape patterns of species-level association between ground-beetles and overstory trees in boreal forests of western Canada (Coleoptera, Carabidae). ZooKeys 2011, 147, 577–600. [Google Scholar] [CrossRef]
- Sanderson, R.A.; Rushton, S.P.; Cherrill, A.; Byrne, J.P. Soil, Vegetation and Space: An Analysis of Their Effects on the Invertebrate Communities of a Moorland in North-East England. J. Appl. Ecol. 1995, 32, 506–518. [Google Scholar] [CrossRef]
- Ings, T.; Hartley, S. The effect of habitat structure on carabid communities during the regeneration of a native Scottish forest. For. Ecol. Manag. 1999, 119, 123–136. [Google Scholar] [CrossRef]
- Niemelä, J.K.; Spence, J.R. Distribution of forest dwelling carabids (Coleoptera): Spatial scale and the concept of communities. Ecography 1994, 17, 166–175. [Google Scholar] [CrossRef]
- Koivula, M.; Punttila, P.; Haila, Y.; Niemelä, J. Leaf Litter and the Small-Scale Distribution of Carabid Beetles (Coleoptera, Carabidae) in the Boreal Forest. Ecography 1999, 22, 424–435. [Google Scholar] [CrossRef] [Green Version]
- Hopp, P.W.; Ottermanns, R.; Caron, E.; Meyer, S.; ROß-NICKOLL, M. Recovery of litter inhabiting beetle assemblages during forest regeneration in the Atlantic forest of Southern Brazil. Insect. Syst. Divers. 2010, 3, 103–113. [Google Scholar] [CrossRef]
- Guillemain, M.; Loreau, M.; Daufresne, T. Relationships beetween the regional distribution of carabid beetles (Coleoptera, Carabidae) and the abundance of their potential prey. Acta Oecol. 1997, 18, 465–483. [Google Scholar] [CrossRef]
- Molnár, T.; Magura, T.; Tóthmérész, B.; Elek, Z. Ground beetles (Carabidae) and edge effect in oak-hornbeam forest and grassland transects. Eur. J. Soil Biol. 2001, 37, 297–300. [Google Scholar] [CrossRef]
- Fuller, R.J.; Oliver, T.H.; Leather, S.R. Forest management effects on carabid beetle communities in coniferous and broadleaved forests: Implications for conservation. Insect. Syst. Divers. 2008, 1, 242–252. [Google Scholar] [CrossRef]
- Lal, R. Soil Carbon Sequestration Impacts on Global Climate Change and Food Security. Science 2004, 304, 1623–1627. [Google Scholar] [CrossRef] [Green Version]
- Lal, R.; Augustin, B.J. Carbon Sequestration in Urban Ecosystems; Springer Science & Business Media: Cham, The Netherlands, 2012. [Google Scholar]
- Hemerik, L.; Brussaard, L. Diversity of soil macro-invertebrates in grasslands under restoration succession. Eur. J. Soil Biol. 2002, 38, 145–150. [Google Scholar] [CrossRef]
- Magdoff, F.; Weil, R.R. Soil Organic Matter in Sustainable Agriculture, 1st ed.; CRC Press: Boca Raton, FL, USA, 2004; p. 412. [Google Scholar]
- Brock, C.; Oberholzer, H.R.; Franko, U. Soil organic matter balance as a practical tool for environmental impact assessment and management support in arable farming. Eur. J. Soil Sci. 2017, 68, 951–952. [Google Scholar] [CrossRef] [Green Version]
- Lundholm, J.T.; Marlin, A. Habitat origins and microhabitat preferences of urban plant species. Urban Ecosyst. 2006, 9, 139–159. [Google Scholar] [CrossRef]
- Sadler, J.; Small, E.C.; Fiszpan, H.A.; Telfer, M.; Niemela, J. Investigating environmental variation and landscape characteristics of an urban-rural gradient using woodland carabid assemblages. J. Biogeogr. 2006, 33, 1126–1138. [Google Scholar] [CrossRef]
- Gagne, S.A.; Fahrig, L. Do birds and beetles show similar responses to urbanization? Ecol. Appl. 2011, 21, 2297–2312. [Google Scholar] [CrossRef] [Green Version]
- Hartley, J.D.; Koivula, J.M.; Spence, R.J.; Pelletier, R.; Ball, E.G. Effects of urbanization on ground beetle assemblages (Coleoptera, Carabidae) of grassland habitats in western Canada. Ecography 2007, 30, 673–684. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Variable | Rural | Suburban | Urban |
|---|---|---|---|
| Elevation range (m) | 800–820 | 690–710 | 570–590 |
| Average Buildings (area in km2) | 0 | 0.0218 | 0.2564 |
| Average Road & Asphalt (length in km) | 0 | 3.728 | 11.06 |
| Average Pavement (length in km) | 0 | 1.094 | 2.242 |
| Average pH | 8.618 | 8.562 | 8.416 |
| Average EC (µS/cm) | 159.10 | 493.42 | 1196.30 |
| Average Soil Organic Carbon (SOC) | 0.383 | 0.311 | 0.883 |
| Average Soil Organic Matter (SOM) | 0.659 | 0.534 | 1.521 |
| Average clay | 24 | 21.5 | 23.5 |
| Average silt | 14.5 | 10.5 | 14.5 |
| Average sand | 61.5 | 68 | 62 |
| Texture | Sandy Clay Loam | Sandy Clay Loam | Sandy Clay Loam |
| Average bar ground percentage | 34 | 26 | 20 |
| Average plant cover percentage | 66 | 74 | 79 |
| Average litter cover percentage | 1.36 | 0.5 | 11 |
| Average litter depth | 0.31 | 0.05 | 0.01 |
| Average log percentage | 7.94 | 32.54 | 28.08 |
| Eigenvalue | F | p | Weighted Correlation Matrix | ||||
|---|---|---|---|---|---|---|---|
| Axis 1 | Axis 2 | Total | Axis 1 | Axis 2 | |||
| Urbanization level | 0.638 | 0.356 | 2.169 | ||||
| Buildings | 5.18 | 0.0020 | 0.9589 | 0.2135 | |||
| Pavements | 1.21 | 0.270 | 0.7182 | 0.5657 | |||
| Roads and asphalt | 0.62 | 0.884 | 0.8437 | 0.4683 | |||
| Elevation | 3.75 | 0.0020 | −0.7707 | −0.6028 | |||
| Vegetation cover | 0.459 | 0.314 | 2.169 | ||||
| Litter cover | 3.45 | 0.0020 | 0.8676 | 0.1654 | |||
| Litter depth | 2.74 | 0.0020 | 0.6417 | 0.6300 | |||
| Flora | 0.649 | 0.366 | 2.195 | ||||
| Alhagi graecorum | 3.327 | 0.01 | −0.5362 | 0.8231 | |||
| Heliotropium currasavicum | 5.33 | 0.0020 | 0.9770 | 0.1765 | |||
| Launaea capitata | 3.80 | 0.0020 | −0.3841 | −0.9034 | |||
| Lycium shawii | 1.88 | 0.0260 | −0.3173 | −0.8271 | |||
| Soil chemical properties | 0.568 | 0.385 | 2.195 | ||||
| Soil Organic Carbon (SOC) | 4.53 | 0.0020 | 0.9420 | 0.0000 | |||
| Soil Organic Matter (SOM) | 0.67 | 0.80 | 0.9415 | −0.0007 | |||
| Family | Species | Gradient | IV | p |
|---|---|---|---|---|
| Tenebrionidae | Adesmia stoeckleini Koch, 1940 | 0 | 100 | 0.001 |
| Tenebrionidae | Blaps kollari kollari Seidlitz, 1896 | 0 | 66.7 | 0.01 |
| Tenebrionidae | Scleropatroides sp. | 0 | 66.7 | 0.02 |
| Tenebrionidae | Oxycara saudarabica Kaszab, 1979 | 0 | 72.3 | 0.007 |
| Carabidae | Mesolestes quadriguttatus (Mateu, 1979) | 0 | 66.7 | 0.02 |
| Carabidae | Eremolestes sulcatus (Chaudoir, 1876) | 0 | 66.7 | 0.02 |
| Tenebrionidae | Akis spinosa (Linnaeus, 1764) | 0 | 66.7 | 0.016 |
| Tenebrionidae | Pimelia thomasi thomasi Blair, 1931 | 1 | 75 | 0.01 |
| Tenebrionidae | Gonocephalum soricinum (Reiche & Saulcy, 1857) | 1 | 73.3 | 0.01 |
| Tenebrionidae | Tentyrina deserta deserta Kaszab, 1981 | 1 | 54.5 | 0.045 |
| Scarabaeidae | Rhyssemus saudi Pittion, 1984 | 1 | 63.8 | 0.035 |
| Tenebrionidae | Zophosis punctata Brullé, 1832 | 1 | 72.5 | 0.007 |
| Dermestidae | Anthrenus malkini Mroczkowski, 1980 | 1 | 50 | 0.05 |
| Tenebrionidae | Gonocephalum besnardi Kaszab, 1982 | 2 | 60 | 0.02 |
| Tenebrionidae | Sclerum orientalis (Fabricius, 1775) | 2 | 80 | 0.006 |
| Anthicidae | Anthelephila caeruleipennis (La Ferté-Sénectère, 1847) | 2 | 85.7 | 0.003 |
| Anthicidae | Anthicus crinitus LaFerté-Sénectère, 1849 | 2 | 66.7 | 0.011 |
| Scarabaeidae | Maladera insanabilis (Brenske, 1894) | 2 | 61 | 0.03 |
| Anthicidae | Endomia lefebvrei (LaFerté-Sénectère, 1849) | 2 | 100 | 0.001 |
| Scarabaeidae | Pentodon algerinus (Füessly, 1778) | 2 | 60 | 0.03 |
| Carabidae | Microlestes infuscatus (Motschulsky, 1859) | 2 | 60 | 0.03 |
| Tenebrionidae | Gonocephalum prolixum (Erichson, 1843) | 2 | 60 | 0.02 |
| Tenebrionidae | Gonocephalum rusticum (Olivier, 1811) | 2 | 70 | 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdel-Dayem, M.S.; Sharaf, M.R.; Majer, J.D.; Al-Sadoon, M.K.; Soliman, A.M.; Aldawood, A.S.; Aldhafer, H.M.; Orabi, G.M. Does a Rural-Urban Gradient Affect Beetle Assemblages in an Arid Ecosystem? Diversity 2023, 15, 303. https://doi.org/10.3390/d15020303
Abdel-Dayem MS, Sharaf MR, Majer JD, Al-Sadoon MK, Soliman AM, Aldawood AS, Aldhafer HM, Orabi GM. Does a Rural-Urban Gradient Affect Beetle Assemblages in an Arid Ecosystem? Diversity. 2023; 15(2):303. https://doi.org/10.3390/d15020303
Chicago/Turabian StyleAbdel-Dayem, Mahmoud S., Mostafa R. Sharaf, Jonathan D. Majer, Mohammed K. Al-Sadoon, Ahmed M. Soliman, Abdulrahman S. Aldawood, Hathal M. Aldhafer, and Gamal M. Orabi. 2023. "Does a Rural-Urban Gradient Affect Beetle Assemblages in an Arid Ecosystem?" Diversity 15, no. 2: 303. https://doi.org/10.3390/d15020303
APA StyleAbdel-Dayem, M. S., Sharaf, M. R., Majer, J. D., Al-Sadoon, M. K., Soliman, A. M., Aldawood, A. S., Aldhafer, H. M., & Orabi, G. M. (2023). Does a Rural-Urban Gradient Affect Beetle Assemblages in an Arid Ecosystem? Diversity, 15(2), 303. https://doi.org/10.3390/d15020303