

Experimental Suppression of Red Imported Fire Ants (Solenopsis invicta) Has Little Impact on the Survival of Eggs to Third Instar of Spring-Generation Monarch Butterflies (Danaus plexippus) Due to Buffering Effects of Host-Plant Arthropods

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Monarch Survival and Host-Plant Arthropods
2.2. RIFA Density and Abundance among Controls
2.3. Experimental Manipulation of RIFA Abundance
2.4. Statistical Analyses
3. Results
3.1. Analyses of RIFA Measures and Monarch Survival among Controls
3.2. Effect of Experimental Manipulation of RIFA on Measures of RIFA Abundance
3.3. Effect of Experimental Manipulation of RIFA on Monarch Survival
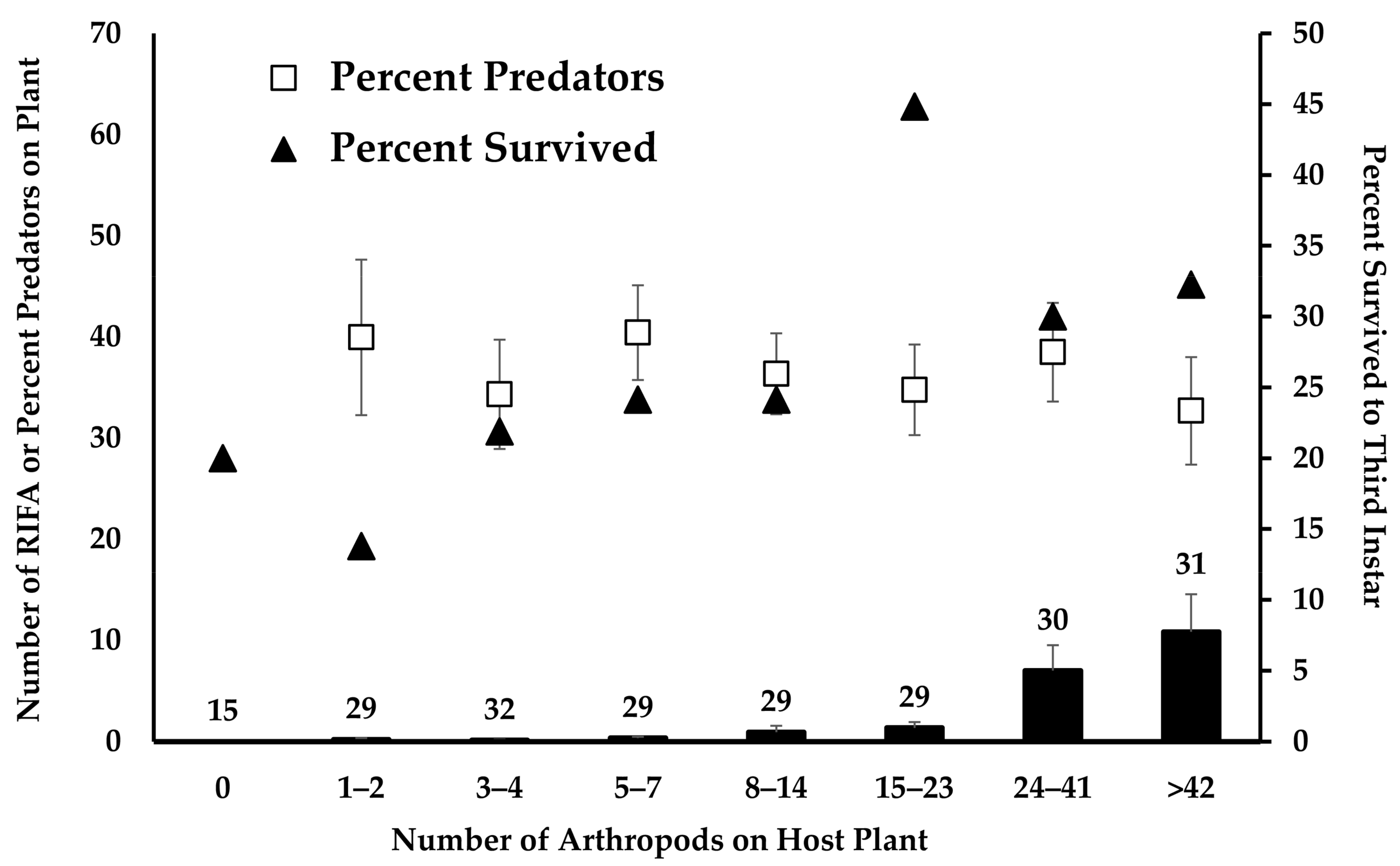
3.4. What Attracts RIFA onto Host Plants and How Does That Impact Monarch Survival?
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Common Name | Included Taxa | Abundance | Frequency | % Frequency |
|---|---|---|---|---|
| Aphids | Aphids (Aphididae) | 10,792 | 80 | 35.71 |
| Other Ants | Hymenoptera, Formicidae, Others | 907 | 37 | 16.52 |
| Little Black Ant | Hymenoptera, Formicidae, Monomorium minimum | 855 | 74 | 33.04 |
| Weevils | Coleoptera, Curculionidae | 471 | 67 | 29.91 |
| Mites | Arachnida, Acari, Mites | 268 | 62 | 27.68 |
| Other Non-Predatory Arthropods | Stick Insects (Phasmatodea), Crickets (Gryllidae), Click Beetles (Elateridae), Darkling Beetles (Tenebrionidae), Leaf-footed Bugs (Coreidae), Seed Bugs (Lygaeidae), Plant Bugs (Miridae), Unidentified Wasps (Apocrita), Millipedes (Diplopoda), Springtails (Collembola), Ticks (Acari), Shield Bugs (Pentatomoidea, non-predatory), Butterflies, Skippers, and Moths (Lepidoptera), Caddisflies (Trichoptera), Slugs and Snails (Mollusca), Mayflies (Ephemeroptera), Harvestmen (Opiliones), Bees (Hymenoptera, Apidae), Grasshoppers (Caelifera), Katydids (Tettigoniidae), Unidentified True Bugs (Hemiptera, Heteroptera) | 267 | 111 | 49.55 |
| Jumping Spiders | Araneae, Salticidae | 247 | 116 | 51.79 |
| Other Predatory Arthropods | Rove Beetles (Staphylinidae), Soldier Beetles (Cantharidae), Ground Beetles (Carabidae), Assassin Bugs (Reduviidae), Predatory Stink Bugs (Pentatomidae, Asopinae), Vespid Wasps (Vespidae), Scorpionflies (Mecoptera), Lacewings (Neuroptera), Hoverflies (Syrphidae), Ladybeetles (Coccinellidae), Wolf Spiders (Lycosidae), Grass Spiders (Agelenidae), Nursery Web Spiders (Pisuridae), Long-jawed Orb Weavers (Tetragnathidae), Lynx Spiders (Oxyopidae), Crab Spiders (Thomisidae), Unidentified Spiders | 227 | 123 | 54.91 |
| Other Leaf Beetles | Coleoptera: Flea Beetles (Chysomelidae, Alticini) and all other Leaf Beetles (Chysomelidae) | 171 | 65 | 29.02 |
| Dermestid Beetles | Coleoptera, Dermestidae | 139 | 28 | 12.50 |
| Flies | Midge Flies (Chironomidae), Fruit Flies (Drosophilidae), Mosquitoes (Culicidae), and Unknown Flies | 138 | 81 | 36.16 |
| Leafhoppers | Hemiptera, Cicadomorpha | 137 | 77 | 34.38 |
| Large Milkweed Bugs | Hemiptera, Lygaeidae, Oncopeltus fasciatus | 108 | 38 | 16.96 |
| Other Milkweed Herbivores | Small Milkweed Bugs (Lygaeus kalmia), Milkweed Longhorn Beetles (Tetraopes texanus), and Thrips (Thysanoptera) | 48 | 34 | 15.18 |
| All Other Beetles | Unidentified Beetles (Coleoptera) | 33 | 22 | 9.82 |
References
- Petition for Protection of the Monarch Butterfly under the Endangered Species Act. Xerces Society. Available online: https://xerces.org/publications/policy-statements/petition-for-esa-protection-monarchs (accessed on 14 February 2023).
- Nail, K. Monarch (Danaus Plexippus) Species Status Assessment Report, Version 2.1 September 2020. 2020, p. 126. Available online: https://www.fws.gov/media/monarch-butterfly-species-status-assessment-ssa-report (accessed on 14 February 2023).
- Walker, A.; Oberhauser, K.S.; Pelton, E.M.; Pleasants, J.M.; Thogmartin, W.E. Danaus Plexippus ssp. Plexippus. Available online: https://doi.org/10.2305/IUCN.UK.2022-1.RLTS.T194052138A200522253.en (accessed on 14 February 2023).
- Flockhart, D.T.T.; Wassenaar, L.I.; Martin, T.G.; Hobson, K.A.; Wunder, M.B.; Norris, D.R. Tracking Multi-Generational Colonization of the Breeding Grounds by Monarch Butterflies in Eastern North America. Proc. R. Soc. B 2013, 280, 20131087. [Google Scholar] [CrossRef] [PubMed]
- Brower, L.P.; Fink, L.S.; Walford, P. Fueling the Fall Migration of the Monarch Butterfly. Integr. Comp. Biol. 2006, 46, 1123–1142. [Google Scholar] [CrossRef] [PubMed]
- Malcolm, S.B.; Cockrell, B.J.; Brower, L.P. Spring Recolonization of Eastern North America by the Monarch Butterfly: Successive Brood or Single Sweep Migration. In Biology and Conservation of the Monarch Butterfly; Natural History Museum of Los Angeles County: Science Series; Natural History Museum of Los Angeles County: Los Angeles, CA, USA, 1993; pp. 253–267. ISBN 978-99940-0-921-3. [Google Scholar]
- Agrawal, A. Monarchs and Milkweed; Princeton University Press: Princeton, NJ, USA, 2017; p. 283. ISBN 978-0-691-16635-3. [Google Scholar]
- Pocius, V.M.; Cibotti, S.; Ray, S.; Ankoma-Darko, O.; McCartney, N.B.; Schilder, R.J.; Ali, J.G. Impacts of Larval Host Plant Species on Dispersal Traits and Free-Flight Energetics of Adult Butterflies. Commun. Biol. 2022, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Pocius, V.M.; Pleasants, J.M.; Debinski, D.M.; Bidne, K.G.; Hellmich, R.L.; Bradbury, S.P.; Blodgett, S.L. Monarch Butterflies Show Differential Utilization of Nine Midwestern Milkweed Species. Front. Ecol. Evol. 2018, 6, 169. [Google Scholar] [CrossRef]
- Calvert, W.H. Patterns in the Spatial and Temporal Use of Texas Milkweeds (Asclepiadaceae) by the Monarch Butterfly (Danaus Plexippus L.) during Fall, 1996. J. Lepid. Soc. 1999, 53, 37–44. [Google Scholar]
- Tracy, J.L.; Kantola, T.; Baum, K.A.; Coulson, R.N. Distribution and phenology of monarch butterfly larvae and their milkweed hosts in the South Central US. Biodivers. Conserv. 2022, 31, 1797–1827. [Google Scholar] [CrossRef]
- Scott, A.; Contreras, K.; Stevenson, M.; Hudman, K.L.; Kopachena, J.G. Survival of Eggs to Third Instar of Late-Summer and Fall-Breeding Monarch Butterflies (Danaus Plexippus) and Queen Butterflies (Danaus Gilippus) in North Texas. J. Insect Conserv. 2022. [Google Scholar] [CrossRef]
- Stevenson, M.; Hudman, K.L.; Scott, A.; Contreras, K.; Kopachena, J.G. High Survivorship of First-Generation Monarch Butterfly Eggs to Third Instar Associated with a Diverse Arthropod Community. Insects 2021, 12, 567. [Google Scholar] [CrossRef]
- Betz, R.F.; Rommel, W.R.; Dichtl, J.J. Insect Herbivores of 12 Milkweed (Asclepias) Species. In Fifteenth North American Prairie Conference; The Natural Areas Association: Bend, OR, USA, 1997; pp. 7–19. Available online: https://search.library.wisc.edu/digital/A2CKQ72QXTC6FV87/pages/AQESWKN565SM458Z (accessed on 14 February 2023).
- Fishbein, M.; Venable, D.L. Diversity and Temporal Change in the Effective Pollinators of Asclepias Tuberosa. Ecology 1996, 77, 1061–1073. [Google Scholar] [CrossRef]
- Liaw, S.-W. The Structure of the Gynostegium, Breeding System, and Pollination Ecology of Spider Milkweed, Asclepias Viridis Walter (Apocynaceae). ONPR 2016, 16, 10–44. [Google Scholar] [CrossRef]
- Agrawal, A.A.; Ali, J.G.; Rasmann, S.; Fishbein, M. Macroevolutionary trends in the defense of milkweeds against monarchs. In Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly; Cornell University Press: Ithaca, NY, USA, 2015; pp. 47–59. ISBN 978-0-8014-5315-1. [Google Scholar]
- Malcolm, S.B. Cardenolide-Mediated Interactions between Plants and Herbivores. In Herbivores: Their Interactions with Secondary Plant Metabolites; Elsevier: Amsterdam, The Netherlands, 1991; pp. 251–296. ISBN 978-0-12-597183-6. [Google Scholar]
- Agrawal, A.A.; Petschenka, G.; Bingham, R.A.; Weber, M.G.; Rasmann, S. Toxic Cardenolides: Chemical Ecology and Coevolution of Specialized Plant–Herbivore Interactions. New Phytol. 2012, 194, 28–45. [Google Scholar] [CrossRef]
- Petschenka, G.; Fei, C.S.; Araya, J.J.; Schröder, S.; Timmermann, B.N.; Agrawal, A.A. Relative Selectivity of Plant Cardenolides for Na+/K+-ATPases From the Monarch Butterfly and Non-Resistant Insects. Front. Plant Sci. 2018, 9, 1424. [Google Scholar] [CrossRef]
- Jones, P.L.; Petschenka, G.; Flacht, L.; Agrawal, A.A. Cardenolide Intake, Sequestration, and Excretion by the Monarch Butterfly along Gradients of Plant Toxicity and Larval Ontogeny. J. Chem. Ecol. 2019, 45, 264–277. [Google Scholar] [CrossRef]
- Isman, M.B.; Duffey, S.S.; Scudder, G.G.E. Cardenolide Content of Some Leaf- and Stem-Feeding Insects on Temperate North American Milkweeds (Asclepias Spp.). Can. J. Zool. 1977, 55, 1024–1028. [Google Scholar] [CrossRef]
- Malcolm, S.B. Milkweeds, Monarch Butterflies and the Ecological Significance of Cardenolides. Chemoecology 1994, 5–6, 101–117. [Google Scholar] [CrossRef]
- Petschenka, G.; Agrawal, A.A. Milkweed Butterfly Resistance to Plant Toxins Is Linked to Sequestration, Not Coping with a Toxic Diet. Proc. R. Soc. B 2015, 282, 20151865. [Google Scholar] [CrossRef]
- Calvert, W.H. Fire Ant Predation on Monarch Larvae (Nymphalidae: Danainae) in a Central Texas Prairie. J. Lepid. Soc. 1996, 50, 149–151. [Google Scholar]
- Lynch, S.P.; Martin, R.A. Milkweed Host Plant Utilization and Cardenolide Sequestration by Monarch Butterflies in Louisiana and Texas. In Biology and Conservation of the Monarch Butterfly; Natural History Museum of Los Angeles County: Science Series; Natural History Museum of Los Angeles County: Los Angeles, CA, USA, 1993; pp. 107–123. ISBN 978-9994009213. [Google Scholar]
- Calvert, W.H. The Effect of Fire Ants on Monarchs Breeding in Texas. In The Monarch Butterfly: Biology and Conservation; Cornell University Press: Ithaca, NY, USA, 2004; pp. 47–53. ISBN 978-0-8014-4188-2. [Google Scholar]
- De Anda, A.; Oberhauser, K.S. Invertebrate Natural Enemies and Stage-Specific Mortality Rates of Monarch Eggs and Larvae. In Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly; Cornell University Press: Ithaca, NY, USA, 2015; pp. 60–70. ISBN 978-0-8014-5315-1. [Google Scholar]
- Oberhauser, K.S.; Anderson, M.; Anderson, S.; Caldwell, W.; De Anda, A.; Hunter, M.; Kaiser, M.C.; Solensky, M.J.; De Anda, A.; Hunter, M.D. Lacewings, Wasps, and Flies—Oh My: Insect Enemies Take a Bite out of Monarchs. In Monarchs in a Changing World: Biology and Conservation of an Iconic Insect; Cornell University Press: Ithaca, NY, USA, 2015; pp. 71–82. ISBN 978-0-8014-5315-1. [Google Scholar]
- Prysby, M.D. Natural Enemies and Survival of Monarch Eggs and Larvae. In The Monarch Butterfly: Biology and Conservation; Cornell University Press: Ithaca, NY, USA, 2004; pp. 27–37. ISBN 978-0-8014-4188-2. [Google Scholar]
- McCoshum, S.M.; Andreoli, S.L.; Stenoien, C.M.; Oberhauser, K.S.; Baum, K.A. Species Distribution Models for Natural Enemies of Monarch Butterfly (Danaus Plexippus) Larvae and Pupae: Distribution Patterns and Implications for Conservation. J. Insect Conserv. 2016, 20, 223–237. [Google Scholar] [CrossRef]
- Pocius, V.M.; Majewska, A.A.; Freedman, M.G. The Role of Experiments in Monarch Butterfly Conservation: A Review of Recent Studies and Approaches. Ann. Entomol. Soc. Am. 2022, 115, 10–24. [Google Scholar] [CrossRef]
- Ascunce, M.S.; Yang, C.-C.; Oakey, J.; Calcaterra, L.; Wu, W.-J.; Shih, C.-J.; Goudet, J.; Ross, K.G.; Shoemaker, D. Global Invasion History of the Fire Ant Solenopsis invicta. Science 2011, 331, 1066–1068. [Google Scholar] [CrossRef]
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species: A Selection from the Global Invasive Species Database; Invasive Species Specialist Group: Auckland, New Zealand, 2000; Volume 12, Available online: https://www.academia.edu/download/33655728/100_world_worst_invasive_alien_species_English.pdf (accessed on 14 February 2023).
- Chen, S.; Ding, F.; Hao, M.; Jiang, D. Mapping the Potential Global Distribution of Red Imported Fire Ant (Solenopsis Invicta Buren) Based on a Machine Learning Method. Sustainability 2020, 12, 10182. [Google Scholar] [CrossRef]
- Vinson, S.B.; Sorensen, A.A. Imported Fire Ants: Life History and Impac; Texas Department of Agriculture: Austin, TX, USA, 1986; p. 28. [Google Scholar]
- Cokendolpher, J.C.; Phillips, S.A. Rate of Spread of the Red Imported Fire Ant, Solenopsis Invicta (Hymenoptera: Formicidae), in Texas. Southwest. Nat. 1989, 34, 443–449. [Google Scholar] [CrossRef]
- USDA; APHIS. Imported Fire Ants (IFA) Interactive Map. Available online: https://www.aphis.usda.gov/aphis/maps/plant-health/ifa-quarantine-mapping (accessed on 7 February 2023).
- Wojcik, D.P.; Allen, C.R.; Brenner, R.J.; Forys, E.A.; Jouvenaz, D.P.; Lutz, R.S. Red Imported Fire Ants: Impact on Biodiversity; Nebraska Cooperative Fish & Wildlife Research Unit—Staff Publications. University of Nebraska-Lincoln: Lincoln, NB, USA, 2001; p. 23. [Google Scholar]
- Holway, D.A.; Lach, L.; Suarez, A.V.; Tsutsui, N.D.; Case, T.J. The Causes and Consequences of Ant Invasions. Annu. Rev. Ecol. Syst. 2002, 33, 181–233. [Google Scholar] [CrossRef]
- Kopachena, J.G.; Buckley, A.J.; Potts, G.A. Effects of the Red Imported Fire Ant (Solenopsis invicta) on Reproductive Success of Barn Swallows (Hirundo rustica) in Northeast Texas. Southwest. Nat. 2000, 45, 477–482. [Google Scholar] [CrossRef]
- Allen, C.R.; Epperson, D.M.; Garmestani, A.S. Red Imported Fire Ant Impacts on Wildlife: A Decade of Research. Am. Midl. Nat. 2004, 152, 88–103. [Google Scholar] [CrossRef]
- Porter, S.D.; Savignano, D.A. Invasion of Polygyne Fire Ants Decimates Native Ants and Disrupts Arthropod Community. Ecology 1990, 71, 2095–2106. [Google Scholar] [CrossRef]
- Morrison, L.W. Long-Term Impacts of an Arthropod-Community Invasion by the Imported Fire Ant, Solenopsis Invicta. Ecology 2002, 83, 2337–2345. [Google Scholar] [CrossRef]
- Epperson, D.M.; Allen, C.R.; Hogan, K.F. Red Imported Fire Ants Reduce Invertebrate Abundance, Richness, and Diversity in Gopher Tortoise Burrows. Diversity 2020, 13, 7. [Google Scholar] [CrossRef]
- Epperson, D.M.; Allen, C.R. Red Imported Fire Ant Impacts on Upland Arthropods in Southern Mississippi. Am. Midl. Nat. 2010, 163, 54–63. [Google Scholar] [CrossRef]
- Lei, W.; XU, Y.; Ling, Z.; LU, Y. Impact of the Red Imported Fire Ant Solenopsis Invicta Buren on Biodiversity in South China: A Review. J. Integr. Agric. 2019, 18, 788–796. [Google Scholar] [CrossRef]
- King, J.R.; Tschinkel, W.R. Experimental Evidence That the Introduced Fire Ant, Solenopsis Invicta, Does Not Competitively Suppress Co-Occurring Ants in a Disturbed Habitat. J. Anim. Ecol. 2006, 1370–1378. [Google Scholar] [CrossRef]
- Schmid, R.B.; Lundgren, J.G. Red Imported Fire Ant, Solenopsis Invicta (Burden) (Hymenoptera: Formicidae), Abundance and Arthropod Community Diversity Affected by Pasture Management. Fla. Entomol. 2022, 105, 53–57. [Google Scholar] [CrossRef]
- Tschinkel, W.R. The Organization of Foraging in the Fire Ant, Solenopsis Invicta. J. Insect Sci. 2011, 11, 26. [Google Scholar] [CrossRef]
- Traniello, J.F. Foraging Strategies of Ants. Annu. Rev. Entomol. 1989, 34, 191–210. [Google Scholar] [CrossRef]
- Vogt, J.T.; Grantham, R.A.; Corbett, E.; Rice, S.A.; Wright, R.E. Dietary Habits of Solenopsis Invicta (Hymenoptera: Formicidae) in Four Oklahoma Habitats. Environ. Entomol. 2002, 31, 47–53. [Google Scholar] [CrossRef]
- Cassill, D.L.; Tschinkel, W.R. Regulation of Diet in the Fire Ant, Solenopsis Invicta. J. Insect Behav. 1999, 12, 307–328. [Google Scholar] [CrossRef]
- Stein, M.B.; Thorvilson, H.G.; Johnson, J.W. Seasonal Changes in Bait Preference by Red Imported Fire Ant, Solenopsis Invicta (Hymenoptera: Formicidae). Fla. Entomol. 1990, 73, 117–123. [Google Scholar] [CrossRef]
- Ali, A.D.; Reagan, T.E. Comparison of Baits for Monitoring Foraging Activity of the Red Imported Fire Ant (Hymenoptera: Formicidae). J. Econ. Entomol. 1986, 79, 1404–1405. [Google Scholar] [CrossRef]
- Porter, S.D.; Tschinkel, W.R. Foraging in Solenopsis invicta (Hymenoptera: Formicidae): Effects of Weather and Season. Environ. Entomol. 1987, 16, 802–808. [Google Scholar] [CrossRef]
- Wilder, S.M.; Holway, D.A.; Suarez, A.V.; LeBrun, E.G.; Eubanks, M.D. Intercontinental Differences in Resource Use Reveal the Importance of Mutualisms in Fire Ant Invasions. Proc. Natl. Acad. Sci. USA 2011, 108, 20639–20644. [Google Scholar] [CrossRef]
- Roeder, K.A.; Penuela Useche, V.; Levey, D.J.; Resasco, J. Testing Effects of Invasive Fire Ants and Disturbance on Ant Communities of the Longleaf Pine Ecosystem. Ecol. Entomol. 2021, 46, 964–972. [Google Scholar] [CrossRef]
- Zalucki, M.P.; Kitching, R.L. Temporal and Spatial Variation of Mortality in Field Populations of Danaus Plexippus L. and D. Chrysippus L. Larvae (Lepidoptera: Nymphalidae). Oecologia 1982, 53, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Zalucki, M.P.; Brower, L.P. Survival of First Instar Larvae Of Danaus plexippus (Lepidoptera: Danainae) in Relation to Cardiac Glycoside and Latex Content Of Asclepias humistrata (Asclepiadaceae). Chemoecology 1992, 3, 81–93. [Google Scholar] [CrossRef]
- Borkin, S.S. Notes on Shifting Distribution Patterns and Survival of Immature Danaus plexippus (Lepidoptera: Danaidae) on the Food Plant Asclepias Syriaca. Great Lakes Entomol. 1982, 15, 199–206. [Google Scholar]
- Rawlins, J.E.; Lederhouse, R.C. Developmental Influences of Thermal Behavior on Monarch Caterpillars (Danaus plexippus): An Adaptation for Migration (Lepidoptera: Nymphalidae: Danainae). J. Kans. Entomol. Soc. 1981, 54, 387–408. [Google Scholar]
- Fisher, K.E.; Hellmich, R.L.; Bradbury, S.P. Estimates of Common Milkweed (Asclepias syriaca) Utilization by Monarch Larvae (Danaus plexippus) and the Significance of Larval Movement. J. Insect Conserv. 2020, 24, 297–307. [Google Scholar] [CrossRef]
- Fisher, K.E.; Bradbury, S.P. Plant Abandonment Behavior and Fitness of Monarch Larvae (Danaus plexippus) Is Not Influenced by an Intraspecific Competitor. J. Insect Conserv. 2022, 26, 851–856. [Google Scholar] [CrossRef]
- Zalucki, M.P.; Brower, L.P.; Malcolm, S.B. Oviposition by Danaus plexippus in Relation to Cardenolide Content of Three Asclepias Species in the Southeastern USA. Ecol. Entomol. 1990, 15, 231–240. [Google Scholar] [CrossRef]
- Drees, B.M.; Calixto, A.A.; Nester, P.R. Integrated Pest Management Concepts for Red Imported Fire Ants Solenopsis invicta (Hymenoptera: Formicidae). Insect Sci. 2013, 20, 429–438. [Google Scholar] [CrossRef]
- Eubanks, M.D.; Blackwell, S.A.; Parrish, C.J.; Delamar, Z.D.; Hull-Sanders, H. Intraguild Predation of Beneficial Arthropods by Red Imported Fire Ants in Cotton. Environ. Entomol. 2002, 31, 1168–1174. [Google Scholar] [CrossRef]
- Morrow, M.E.; Chester, R.E.; Lehnen, S.E.; Drees, B.M.; Toepfer, J.E. Indirect Effects of Red Imported Fire Ants on Attwater’s Prairie-Chicken Brood Survival. J. Wildl. Manag. 2015, 79, 898–906. [Google Scholar] [CrossRef]
- Extoxnet Pip-Cyfluthrin. Available online: http://cues.cfans.umn.edu/old/blackvw/insecticides/cyfluthrin.htm (accessed on 14 February 2023).
- Murtaugh, P.A. In Defense of P Values. Ecology 2014, 95, 611–617. [Google Scholar] [CrossRef]
- Wagenmakers, E.-J.; Farrell, S. AIC Model Selection Using Akaike Weights. Psychon. Bull. Rev. 2004, 11, 192–196. [Google Scholar] [CrossRef]
- Li, Y.; Liu, J.S. Robust Variable and Interaction Selection for Logistic Regression and General Index Models. J. Am. Stat. Assoc. 2019, 114, 271–286. [Google Scholar] [CrossRef]
- Greenland, S.; Mansournia, M.A.; Altman, D.G. Sparse Data Bias: A Problem Hiding in Plain Sight. BMJ Med. 2016, 352, 1–6. [Google Scholar] [CrossRef]
- Macom, T.E.; Porter, S.D. Comparison of Polygyne and Monogyne Red Imported Fire Ant (Hymenoptera: Formicidae) Population Densities. Ann. Entomol. Soc. Am. 1996, 89, 535–543. [Google Scholar] [CrossRef]
- Porter, S.D.; Williams, D.F.; Patterson, R.S.; Fowler, H.G. Intercontinental Differences in the Abundance of Solenopsis Fire Ants (Hymenoptera: Formicidae): Escape from Natural Enemies? Environ. Entomol. 1997, 26, 373–384. [Google Scholar] [CrossRef]
- Ricks, B.L.; Vinson, B.S. Feeding Acceptability of Certain Insects and Various Water-Soluble Compounds to Two Varieties of the Imported Fire Ant. J. Econ. Entomol. 1970, 63, 145–148. [Google Scholar] [CrossRef]
- Lanza, J.; Vargo, E.L.; Pulim, S.; Chang, Y.Z. Preferences of the Fire Ants Solenopsis invicta and S. geminata (Hymenoptera: Formicidae) for Amino Acid and Sugar Components of Extrafloral Nectars. Environ. Entomol. 1993, 22, 411–417. [Google Scholar] [CrossRef]
- Vander Meer, R.K.; Lofgren, C.S.; Seawright, J.A. Specificity of the Red Imported Fire Ant (Hymenoptera: Formicidae) Phagostimulant Response to Carbohydrates. Fla. Entomol. 1995, 78, 144. [Google Scholar] [CrossRef]
- Tennant, L.E.; Porter, S.D. Comparison of Diets of Two Fire Ant Species (Hymenoptera: Formicidae): Solid and Liquid Components. J. Entomol. Sci. 1991, 26, 450–465. [Google Scholar] [CrossRef]
- Samuelson, G.A. Pollen Consumption and Digestion by Leaf Beetles. In Novel Aspects of the Biology of Chrysomelidae; Jolivet, P.H., Cox, M.L., Petitpierre, E., Eds.; Series Entomologica; Springer: Dordrecht, The Netherlands, 1994; pp. 179–183. ISBN 978-94-011-1781-4. [Google Scholar]
- Rayor, L.S. Effects of Monarch Larval Host Plant Chemistry and Body Size on Polistes Wasp Predation. In The Monarch Butterfly: Biology and Conservation; Cornell University Press: Ithaca, NY, USA, 2004; pp. 39–46. ISBN 978-0-8014-4188-2. [Google Scholar]
- Mooney, K.A.; Jones, P.; A. Agrawal, A. Coexisting Congeners: Demography, Competition, and Interactions with Cardenolides for Two Milkweed-Feeding Aphids. Oikos 2008, 117, 450–458. [CrossRef]
- Züst, T.; Agrawal, A.A. Plant Chemical Defense Indirectly Mediates Aphid Performance via Interactions with Tending Ants. Ecology 2017, 98, 601–607. [Google Scholar] [CrossRef] [PubMed]
- Mailleux, A.-C.; Deneubourg, J.-L.; Detrain, C. How Do Ants Assess Food Volume? Anim. Behav. 2000, 59, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Hertzog, L.R.; Meyer, S.T.; Weisser, W.W.; Ebeling, A. Experimental Manipulation of Grassland Plant Diversity Induces Complex Shifts in Aboveground Arthropod Diversity. PLoS ONE 2016, 11, e0148768. [Google Scholar] [CrossRef]
- Welti, E.; Helzer, C.; Joern, A. Impacts of Plant Diversity on Arthropod Communities and Plant–Herbivore Network Architecture. Ecosphere 2017, 8, e01983. [Google Scholar] [CrossRef]
- Forister, M.L.; Pelton, E.M.; Black, S.H. Declines in Insect Abundance and Diversity: We Know Enough to Act Now. Conserv. Sci. Pract. 2019, 1, e80. [Google Scholar] [CrossRef]
- Wagner, D.L. Insect Declines in the Anthropocene. Annu. Rev. Entomol. 2020, 65, 457–480. [Google Scholar] [CrossRef]
- Van Klink, R.; Bowler, D.E.; Gongalsky, K.B.; Swengel, A.B.; Gentile, A.; Chase, J.M. Meta-Analysis Reveals Declines in Terrestrial but Increases in Freshwater Insect Abundances. Science 2020, 368, 417–420. [Google Scholar] [CrossRef]
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 Percent Decline over 27 Years in Total Flying Insect Biomass in Protected Areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef]


| Model | AICc | dAICc | wi | Likelihood Ratio X2 | Model Probability |
|---|---|---|---|---|---|
| Intercept, Number of Ramets | 350.592 | 0.000 | 0.487 | 6.2681 | 0.0123 |
| Intercept, Number of Ramets, Number of RIFA Mounds | 351.585 | 0.993 | 0.297 | 7.3027 | 0.0260 |
| Intercept, Number of Ramets, Number of RIFA Mounds, Distance to Nearest RIFA Mound | 352.843 | 2.251 | 0.158 | 8.0805 | 0.0444 |
| Intercept Only | 354.842 | 4.250 | 0.058 | - | - |
| Summary of best-fit model. Concordance of this model was 49.9 percent. | |||||
| Parameter | DF | Estimate | Standard Error | Wald Chi-Square | p |
| Intercept | 1 | −2.3582 | 0.2461 | 91.8391 | <0.0001 |
| Number of Ramets | 1 | 0.1980 | 0.0760 | 6.7878 | 0.0092 |
| Parameter | Suppressed Treatment Mean ± SE (n) | Control Mean ± SE (n) | Enhanced Treatment Mean ± SE (n) | Kruskal–Wallis Test X2 (df = 2) | p |
|---|---|---|---|---|---|
| Distance to Nearest Mound (cm) | 1920.72 ± 102.54 (233) | 208.30 ± 9.79 (448) | 222.35 ± 11.51 (205) | 434.2832 | <0.0001 |
| Number of Mounds ≤ 4 m from Host Plant | 0.21 ± 0.03 (233) | 2.87 ± 0.08 (448) | 2.41 ± 0.10 (205) | 448.9712 | <0.0001 |
| Total Volume of Mounds ≤ 4 m from Host Plant (cm3) | 4817.11 ± 1674.09 (233) | 23,485.36 ± 1562.52 (448) | 26,182.02 ± 2994.29 (205) | 390.6882 | <0.0001 |
| Number of RIFA on Host Plant | 0.04 ± 0.02 (160) | 2.83 ± 0.67 (224) | 60.97 ± 3.63 (114) | 342.8847 | <0.0001 |
| Survival (%) | Chi-Square (1 df) | p | Odds Ratio | 95% Confidence Interval | ||
|---|---|---|---|---|---|---|
| Control | Suppressed | |||||
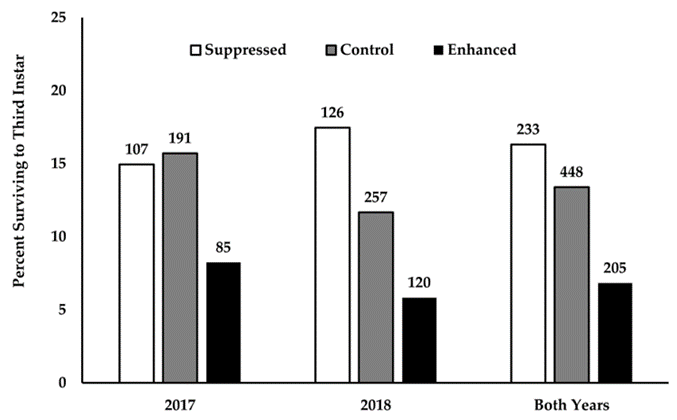
| 2017 | 15.71 (191) | 14.95 (107) | 0.0298 | 0.8629 | 0.9436 | 0.4882–1.8237 |
| 2018 | 11.67 (257) | 17.46 (126) | 2.4133 | 0.1203 | 1.6006 | 0.9763–2.9081 |
| Both Years | 13.39 (448) | 16.31 (233) | 1.0581 | 0.3075 | 1.2602 | 0.8106–1.9592 |
| Model | AICc | dAICc | wi | Likelihood Ratio X2 | Model Probability |
|---|---|---|---|---|---|
| Jumping Spiders, Weevils, Predators other than Ants and Jumping Spiders, Leaf Beetles, Ants other than RIFA or Little Black Ants, Aphids | 205.2060 | 0.0000 | 0.6249 | 85.4613 | <0.0001 |
| Jumping Spiders, Weevils, Predators other than Ants and Jumping Spiders, Leaf Beetles, Ants other than RIFA or Little Black Ants | 206.3443 | 1.1383 | 0.3537 | 82.3591 | <0.0001 |
| Jumping Spiders, Weevils, Predators other than Ants and Jumping Spiders, Leaf Beetles | 211.9911 | 6.7851 | 0.0210 | 74.7674 | <0.0001 |
| Jumping Spiders, Weevils, Predators other than Ants and Jumping Spiders | 219.9316 | 14.7256 | 0.0004 | 64.8995 | <0.0001 |
| Jumping Spiders, Weevils | 230.3222 | 25.1162 | 0.0000 | 52.6019 | <0.0001 |
| Jumping Spiders | 251.1491 | 45.9431 | 0.0000 | 29.8874 | <0.0001 |
| Intercept Only | 279.1675 | 73.9615 | 0.0000 | - | - |
| Summary of the best-fit logistic regression model. Concordance of this model was 83.6%. | |||||
| Parameter | DF | Estimate | Standard Error | Wald Chi-Square | p-value |
| Intercept | 1 | −2.3358 | 0.2879 | 65.8196 | <0.0001 |
| Aphids | 1 | 0.0016 | 0.0011 | 2.0962 | 0.1477 |
| Ants other than RIFA or Little Black Ants | 1 | 0.1870 | 0.1281 | 2.1317 | 0.1443 |
| Weevils | 1 | 0.1276 | 0.0402 | 10.0552 | 0.0015 |
| Predators other than Ants and Jumping Spiders | 1 | 0.3617 | 0.1598 | 5.1245 | 0.0236 |
| Jumping Spiders | 1 | 0.3089 | 0.1368 | 5.0988 | 0.0239 |
| Leaf Beetles | 1 | 0.3223 | 0.1124 | 8.2211 | 0.0041 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hudman, K.L.; Stevenson, M.; Contreras, K.; Scott, A.; Kopachena, J.G. Experimental Suppression of Red Imported Fire Ants (Solenopsis invicta) Has Little Impact on the Survival of Eggs to Third Instar of Spring-Generation Monarch Butterflies (Danaus plexippus) Due to Buffering Effects of Host-Plant Arthropods. Diversity 2023, 15, 331. https://doi.org/10.3390/d15030331
Hudman KL, Stevenson M, Contreras K, Scott A, Kopachena JG. Experimental Suppression of Red Imported Fire Ants (Solenopsis invicta) Has Little Impact on the Survival of Eggs to Third Instar of Spring-Generation Monarch Butterflies (Danaus plexippus) Due to Buffering Effects of Host-Plant Arthropods. Diversity. 2023; 15(3):331. https://doi.org/10.3390/d15030331
Chicago/Turabian StyleHudman, Kalynn L., Misty Stevenson, Kelsey Contreras, Alyx Scott, and Jeffrey G. Kopachena. 2023. "Experimental Suppression of Red Imported Fire Ants (Solenopsis invicta) Has Little Impact on the Survival of Eggs to Third Instar of Spring-Generation Monarch Butterflies (Danaus plexippus) Due to Buffering Effects of Host-Plant Arthropods" Diversity 15, no. 3: 331. https://doi.org/10.3390/d15030331
APA StyleHudman, K. L., Stevenson, M., Contreras, K., Scott, A., & Kopachena, J. G. (2023). Experimental Suppression of Red Imported Fire Ants (Solenopsis invicta) Has Little Impact on the Survival of Eggs to Third Instar of Spring-Generation Monarch Butterflies (Danaus plexippus) Due to Buffering Effects of Host-Plant Arthropods. Diversity, 15(3), 331. https://doi.org/10.3390/d15030331