1. Introduction
Maximizing genetic diversity in founding populations is critical for three reasons: it avoids inbreeding depression, it perpetuates the benefits of natural selection, and it helps conserve the species [
1,
2,
3]. Inbreeding depression, which occurs when there is low genetic diversity within a population and related individuals interbreed, has been shown to be deleterious to bison productivity and survivorship [
4]. Natural selection occurs when environmental pressures act on a population and impact survivorship or reproductive success. Individuals that survive these pressures pass on their genetic material to more offspring, resulting in a change to allele frequencies at loci under selection pressure [
5,
6]. The more genetic diversity within a population, the more likely some alleles will provide a selective benefit, allowing individuals to adapt to changing environmental conditions. Finally, generations of selection pressures (i.e., evolution) lead to highly adapted species. The creation of new, geographically distinct populations within such species can act as a genetic diversity bank that guards against the loss of such diversity [
7].
American bison are currently listed as near threatened on the IUCN Red List, due to their dependence on ongoing conservation programs and generally small population size [
8]. The plains bison subspecies once numbered in the tens of millions in North America [
9,
10]. By the late 1800s, they were reduced to less than 1000 animals, and were extirpated from the wild in Canada [
9]. The Government of Canada has a long history of plains bison recovery that dates back to the early 20th century when it purchased several hundred descendants of the last known wild plains bison except for Yellowstone National Park, shipped them by rail from northern Montana to Alberta, and began several conservation programs that led to the herds in Elk Island, Prince Albert, and Grasslands National Parks and over 30 conservation sites elsewhere across North America [
11].
Despite these efforts, plains bison are still recommended by the Committee on the Status of Endangered Wildlife in Canada (COSEWIC) to be considered a threatened species in Canada [
12]. Aware of the unique opportunity to establish only the sixth free roaming population of plains bison in the world, Parks Canada endeavoured to reintroduce 16 bison to suitable habitat in the backcountry of Banff National Park (abbreviated NP hereafter) in 2017 [
13,
14]. Free-ranging bison herds have been absent from this area for approximately 150 years. Prior to this reintroduction, there were only five conservation herds of plains bison in Canada, with a total of fewer than 1500 mature individuals [
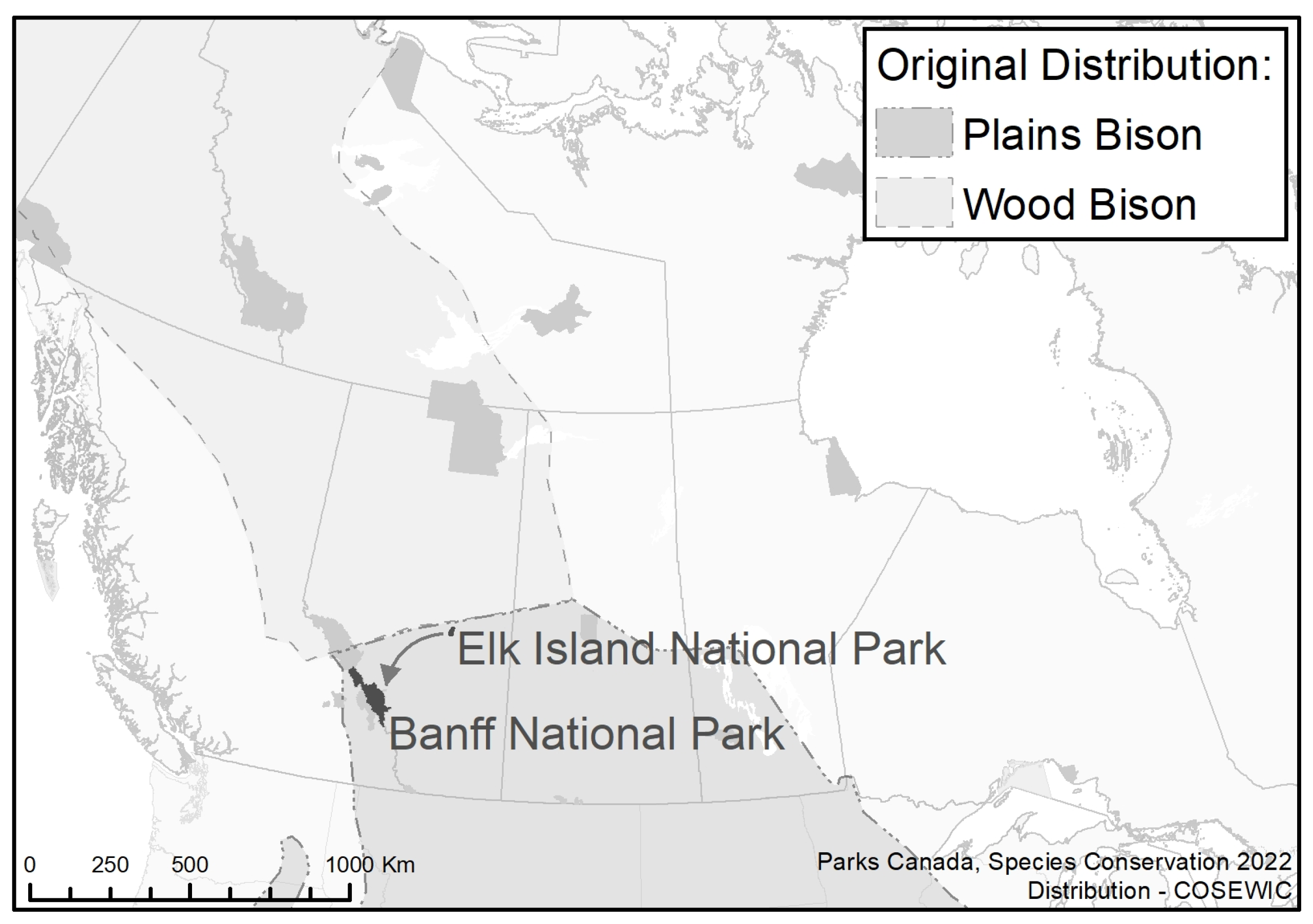
12]. Animals were sourced from Elk Island NP, one of the most genetically diverse bison populations in North America (
Figure 1) [
15]. Translocation logistics, which included a final helicopter lift of animals into Banff NP’s backcountry, limited the size of the founding group to 16 animals.
Small founding populations are prone to the loss of genetic diversity [
16], so efforts were made to ensure the founding herd in Banff NP was as genetically diverse as possible. This was done by (1) measuring variability for some candidate founders at a suite of microsatellite loci, (2) choosing founders that were as unrelated to one another as possible, (3) considering candidates with unique alleles, and (4) ensuring founding females were already impregnated with fetuses sired by nonfounding males. The first two strategies work better than random selections in maintaining genetic diversity in bison populations [
17,
18], whereas the fourth strategy ensures genetic contributions from more males without having to physically move them. While maximizing genetic diversity was considered critically important to the success of the project, especially when considering the relatively small number of founders, the health and safety of the bison during translocation was of utmost importance. When it was observed that a small number of the proposed founders were behaviourally incompatible and risking the safety of the rest of the herd, they were replaced with other, less genetically diverse, animals. This study determined the effectiveness of these techniques in ensuring that the Banff NP population founders were genetically diverse, and assessed the genetic impact of having to replace some individuals for unforeseen reasons just before translocation.
2. Materials and Methods
Elk Island NP is a fenced herd, and large predators are virtually absent from the region. As a consequence, regular removals are required to ensure that the population does not exceed the carrying capacity of its habitat. In January 2017, approximately 70 bison from the ~400 source herd were baited into the handling facility at Elk Island NP and identified for removal. Candidate founders were initially identified based on three criteria: age (2 or 3 years old), sex (predominantly female), and pregnancy status (fetuses in second trimester). Age was estimated by examining tooth irruption and wear [
19], while pregnancy status was determined through manual rectal palpation by qualified wildlife veterinarians. Twenty-five of the 70 animals met these criteria, and were kept in a separate corral at Elk Island NP until the 16 founders could be chosen and moved to Banff NP. The pool of potential candidates consisted of 15 females and 10 males, from which 12 females and 4 males were recommended as founders for the Banff NP herd. This sex ratio of 3:1 was recommended by bison ranchers to minimize aggression between males. A preponderance of females was also considered advantageous for rapid population growth, which should result in less genetic drift. A previous study has shown that bison younger than five are generally incapable of breeding in populations where more mature males are present [
20]. Due to the young age of the prospective male founders, and the size of the Elk Island NP population, it was considered unlikely any of the male founders were sires for the pregnant females. This was confirmed through parentage analysis after the calves were born.
Tail hair samples were collected from each candidate founder, along with 33 other individuals from Elk Island NP to estimate the genetic diversity in the larger source population, for a total of 58 individuals. The same 10 microsatellite loci that were used in previous bison genetic studies in Canada (with the removal of BM4513 [
15,
21,
22]) were analyzed, allowing for direct comparison of the results among studies. These loci were BM143, BM2830, BMC1222, BM1225, BOVFSH, ETH121, RT9, RT24, RT27, and RT29. DNA was extracted by using a Qiagen DNeasy* Blood and Tissue extraction kit (Qiagen, Valencia, SCV, USA). Genotyping was performed at the Molecular Biology Service Unit at the University of Alberta.
All genetic diversity analyses (mean number of alleles, observed and expected heterozygosity, and probability of identity) of the resultant genotypes were performed by using Genalex 6.5 [
23,
24]. Individual mean allele frequency (MAF) is the mean frequency of each allele that makes up an individual’s genotype. Smaller values are representative of individuals that possess alleles that are less common, and who would therefore make a larger contribution to the overall genetic diversity of the founder population. Candidate founders were ranked according to their mean allele frequency (MAF), calculated as in [
18], and the individuals with the lowest MAF scores were considered the most valuable founders. The presence of unique alleles was also considered when choosing founders. Potential founders were surveyed for unique (not present in any other potential founders) or rare (present in one other potential founder) alleles at 10 microsatellite loci. Finally, relatedness measures LR [
25] and QG [
26] were calculated to ensure that close relatives were not included as founders. As QG ranges from −1 to 1, and LR ranges from −0.5 to 0.5, LR measurements were multiplied by two in order to standardize these values. Full-siblings and parent/offspring pairs are expected to be related at the 0.5 level, whereas half-siblings are expected to be related at the 0.25 level. Average unrelated individuals are expected to have pairwise values of 0. Robertson and Hill’s F
IS [
27] was calculated to estimate the level of inbreeding in the Elk Island NP herd and the Banff NP founders by using Genepop [
28,
29].
Candidate founders also had to unexpectedly be screened for behavioral incompatibilities after they were selected for genetic diversity. This occurred once they were penned together and conspecific harassment, which led to extreme and prolonged stress for a few, subdominant individuals, led to the aggressors being removed and replaced. This was deemed necessary due to concerns for how they would fare in the upcoming translocation, in which they would be forced to share 8 m2 transport containers with as many as three other animals for up to 24 h. We selected animals for removal and replacement based on mean stress level scores, which four observers assigned after three hour-long observation periods spread over the initial 24-h period after each unique group of bison was first brought together. Values assigned to animals ranged between 1 (relaxed) and 10 (fleeing or panting) and were averaged between sessions and observers. Animals with a score of 8 or higher were switched out with the next best genetic candidates and the whole process was repeated until scores of 5 or lower were achieved for all animals. This not only led to a different group of founders than was recommended for optimal genetic diversity but, due to a limited number of young, pregnant female candidates, a slightly different sex ratio.
The rapidity with which genetic diversity is lost from a population is primarily impacted by the effective size of the population (Ne) [
30]. The effective population size is reduced by large fluctuations in population size, differential reproductive success within the population, and sex ratio bias [
31]. Both the proposed and actual founding populations would have been subjected to the same fluctuations in population size and differential reproductive success prior to the inception of the Banff NP population, but the difference in sex ratio between these scenarios could impact effective size. N
e was calculated by using Wright’s formula [
30]: N
e = (4 N
m × N
f)/(N
m + N
f), where N
m is the number of males and N
f is the number of females.
Each of the ten females that did pass both the genetic and behavioral selection criteria produced calves a few months after being translocated in 2017. DNA samples from these calves were collected in spring 2018 via chemical immobilization while they were still in the soft release pasture so the following analyses could be performed: (1) maternity of the calves, (2) relationships among calves, and (3) their impact on the genetic diversity of the Banff NP herd. Genetic diversity in the calves was characterized with the 10 microsatellite loci described above to determine how they contributed to the overall genetic diversity of the herd. In order to determine maternity for these calves, a suite of 50 previously described autosomal microsatellites were generated by the Veterinary Genetics Laboratory at the University of California, Davis [
32,
33,
34]. Loci included in this analysis were AGLA232, BL1036, BM1225, BM1706, BM17132, BM1824, BM1862, BM2113, BM2830, BM4107, BM4311, BM4440, BM47, BM711, BM720, BM757, BMC4214, BMS1001, BMS1074, BMS1117, BMS1172, BMS1315, BMS1355, BMS1675, BMS1716, BMS1747, BMS1857, BMS1862, BMS2258, BMS2639, BMS410, BMS510, BMS527, BMS528, BMS601, BMS812, BMS941, HUJ246, IL4, ILSTS102, INRA037, INRA133, INRA194, RM372, SPS115, THLA122, TGLA44, TGLA53, and URB011. Maternity assignments were relatively straightforward because 100% of the potential dams were sampled and female bison only rarely produce twins, so every female could be assigned a single calf. Because of the small sample size, we used exclusion to assign each calf to its dam. Exclusion and pairwise relatedness values were compared to determine if the sires of the calves were present in the Banff NP population. This was considered unlikely given the young age of the male Banff NP founders at the time when the calves were conceived, as well as the large number of potential sires (~100) in the Elk Island National Park source population.
3. Results
3.1. Selection of Banff Founders
Mean allele frequency values for the potential founders are ranked in
Table 1 (males) and
Table 2 (females). MAF ranged from 0.2748 to 0.3881 for the 10 potential founder males and 0.2908 to 0.4081 for the 16 potential founder females.
In total, 15 individuals had unique or rare alleles (
Table 1 and
Table 2). Four individuals, (424, 433, 453, and 473), contained two unique or rare alleles.
A goal of the project was to avoid pairs of individuals related at the full sibling/parent–offspring level or higher (0.5). Full siblings are probably rare in large herds of bison like the source one at Elk Island NP, as calves are born as singletons each year, and would therefore only occur if the female successfully mated with the same male in two different years. Because potential founders for the Banff NP population are 2 or 3 years old, their dams would have had to produce a calf two years in a row with the same sire in order for two of the potential founders to be full siblings. All potential founders were only recently mature enough to reproduce, so parent–offspring relationships were also not possible. Only two pairs of potential founders had relatedness values of >0.5 for at least one of the relatedness measures (
Table 1 and
Table 2). The 421–436 pairing had values of (QG = 0.605, LR = 0.334) and the 420–425 pairing had values of (QG = 0.528, LR = 0.459).
Taking into account mean allele frequency, rare and unique alleles,, and pairwise relatedness, a list of ideal proposed founders was developed to maximize the genetic diversity in the new Banff NP population while avoiding closely related individual pairings (males: 431, 432, 473, and 474; females 414, 415, 424, 426, 428, 433, 435, 436, 437, 443, 449, and 453; see
Table 1 and
Table 2). Only one proposed founding male did not have a unique or rare allele. Three males not proposed as founders had a single unique or rare allele, but their MAF values were higher than the proposed founders, and were not proposed as founders. None of the females that were not proposed as founders contained unique or rare alleles. No pairs of individuals with relatedness values >0.5 were proposed to be included as founders. Female 436 was proposed to be included, but not the individual she shared a high level of relatedness with (421).
However, once isolated and confined in a 1 ha translocation pen at Elk Island NP, aggressive social interactions among the ideal 12 female and four male founders necessitated changes to group composition for the health and safety of the animals during upcoming translocation (see stress scores in
Table 1 and
Table 2). Changes were made iteratively: first, females 453, 414, and 435 were removed and replaced with females 421, 479, and 425. This led to new aggression focused on 437 and 449 and elevated stress scores (
Table 2). They too were removed and, with no eligible two-year-old pregnant females remaining in the larger pool of 25 candidates, were replaced with males 457 and 445. Once they were added, however, high stress scores for male 445 led to him being switched for male 420 whereby relative calm finally presided over the herd. (
Table 1 and
Table 2). The largest difference between the two pools is the sex ratio. Four bulls were originally targeted as founders, but six were actually translocated. These changes also resulted in the inclusion of the two pairs of individuals that had been identified as having high levels of relatedness (
Table 1 and
Table 2).
These last-minute changes to the founding animals ultimately reduced the genetic diversity of the Banff NP founding population. Thirteen unique/rare alleles were represented in the actual founder pool, and 16 were in the proposed founder pool (
Table 1,
Table 2 and
Table S1). The actual founder pool does not contain three of the top five ranked mean allele frequency female candidates (
Table 2). The number of alleles and observed heterozygosity were similar between the proposed and actual founders, with the actual founders having slightly lower levels; however, expected heterozygosity and probability of identity are substantially lower in the actual founders (
Table 3).
With a few exceptions, allele frequencies are similar between Elk Island NP, the proposed founders, and the actual founders (
Supplementary Table S1). The most common allele at locus BMC1225 was 240, but the most common allele in the proposed and actual founders is 270. Four alleles present at low frequencies in the Elk Island NP population (allele 273 at BMC1222, allele 268 at BMC1225, allele 198 at Eth121, and allele 298 at BOVFSH) are absent from both the potential and actual founders. Additionally, allele 316 at BOVFSH is absent from the proposed founders and allele 299 at BOVFSH is absent from the actual founders. Allele 213 from RT24 is also absent in the actual founders.
Ultimately, however, it appears that the reduction in genetic diversity from the proposed to the actual founders is minimal (
Table 3). Mean number of alleles observed at each locus dropped from 5.3 to 5.2, and observed heterozygosity dropped from 0.621 (SE ± 0.073) to 0.620 (SE ± 0.073), well within the standard error. However, expected heterozygosity dropped from 0.663 (SE ± 0.054) to 0.650 (SE ± 0.058), which is lower, even allowing for standard error. Both of these values are greater than that observed in the Elk Island NP population (0.646 ± 0.068) likely due to preferentially choosing individuals with rare alleles as founders. Probability of identity, which measures the probability that two individuals in a population will have the same genotype, also decreased. However, the effective size of the founders increased. The proposed founders had an N
e of 12, whereas the change in sex ratio in the actual founders increased the N
e to 15.
3.2. Calf Parentage
Ten calves (seven female and three male) were born to the founders in 2017. Maternity was successfully assigned by using exclusion in each case. Each mother–offspring pair had 100% concordance across all 50 loci. As expected, all of the pairwise relatedness values between the male founders and the calves were below 0.5, the level at which one would expect parents and offspring to be related. Exclusion also eliminated all male founders as sires at a minimum of three loci.
Because the sires of the 2017 calves were not part of the Banff NP founder population, the birth of these calves should have increased the genetic diversity present in the Banff NP herd by increasing the number of ancestors who had genetic input into the herd. To assess this, we compared the mean number of alleles, observed heterozygosity, expected heterozygosity, and probability of identity of the founders, and the founders plus their calves, to the larger source population in Elk Island (
Table 3).
For the mean number of alleles, observed heterozygosity, and probability of identity, the source herd in Elk Island NP is the most variable. The founding population has higher expected heterozygosity than the source herd, despite being a subset of it, likely due to selecting individuals with rare alleles as part of the founding population. If rare alleles are at a higher frequency in the founding population than in the source Elk Island NP population, expected heterozygosity will be higher.
As expected, the genetic diversity in the Banff NP population increased after the calves born in 2017 are included, using all measures of genetic diversity. This is likely the result of the sires of the calves not being members of the Banff NP population, so their genetic diversity is not already represented. However, even with the inclusion of the calves, the recently founded population in Banff NP has a lower expected heterozygosity and probability of identity than the proposed founders, that were chosen without consideration of behavioural incompatibilities.
The most common allele at BMC1225 in the actual founders plus the calves is 240, which is also the most common allele in the Elk Island NP population but not in the founders before the addition of the calves. Allele 298 at BOVFSH is present in the calves, but not the actual founders, so the use of pregnant females as founders ensured the presence of this allele in the Banff NP population.
FIS, or the potential level of inbreeding, is lower in the actual founders than the proposed founders. This suggests that MAF may not be the ideal tool to use if the goal is to reduce inbreeding in founding populations. FIS is lower when calves are included, and lowest for the Elk Island NP population, likely due to the increase in population size for these scenarios.
4. Discussion
Genetic diversity is key to avoiding inbreeding depression in small populations and providing variability upon which natural selection can act. One of the goals while establishing the Banff NP plains bison population was to attempt to capture as much of the diversity present in Elk Island NP, the source population, as possible. To this end, MAF, the presence of unique alleles, and relatedness were used to identify four males and 12 pregnant females as founders for the Banff NP population. However, although genetic diversity was an important consideration in choosing founders, we also needed to focus on behavioral dynamics within the herd, which led to several last-minute changes. As a result, the ideal individuals from a genetic perspective were not all able to be used as founders. Although this reduced the genetic diversity of the actual Banff NP founders, it does not appear that this reduction was substantial. The use of founder females that were impregnated before they were selected from the larger Elk Island NP population also increased the diversity of the Banff herd after the birth of these calves.
One of the more notable differences between the proposed and actual founding pools for the Banff National Park herd is the change in the sex ratio from 4:12 (males: females) to 6:10. There could be long-term effects of this change on the genetic diversity of the Banff NP herd. As each of the females was pregnant, the reduction in female founders by two means two fewer calves were born into the Banff NP population in the first year. Because the sires for these calves were not members of the founding population, the reduction in pregnant females used to found the herd that are carrying genetic contributions from unsampled males means that there will be a consequent reduction in number of effective founders for the population. As there are fewer females in the population, the growth rate will also be slightly lower, which can impact the maintenance of genetic diversity over time, as populations are more likely to lose diversity while they are small. Another consideration is the reproductive success of four vs. six sires in the Banff NP founder population. Genetic diversity in subsequent generations increases with the number of reproductively successful sires in the previous generation. If all six sires among the actual founders are successful, the potential for inbreeding in subsequent generations will be reduced, and the diversity will be higher than if only four sires were present. This is reflected in the increase in the effective size of the herd from 12 for the proposed founders to 15 for the actual founders. A final consideration is the potential loss of males from the population. If only four male founders had been used, a single mortality event would reduce the number of potential sires by 25%, thereby reducing the effective size of the population. In fact, two males emigrated out of the park and had to be removed from the population within a year of the release, which has reduced the potential sires for subsequent generations. Work is underway to determine if these males were reproductively successful before leaving the park. Due to the unexpected early loss of these males, it is actually fortuitous that the number of founding males was increased at the last minute due to behavioral incompatibilities among the proposed founders.
Using most measures of genetic diversity, the Banff NP founders are less genetically variable than the Elk Island NP population, and less genetically variable than the original founders proposed for the herd. This decrease was minimized when the calves, sired by bulls not translocated from the source herd, were born a few months after their mothers were translocated in 2017. Nonetheless, the mean number of alleles, observed heterozygosity, and probability of identity are still less than observed in Elk Island NP, likely due to the small number of founders and the relatively high genetic diversity present at Elk Island NP. Expected heterozygosity is higher in the recently founded population than in the source population at Elk Island NP, likely due to the use of MAF to choose founders, as it focuses on including individuals with rare alleles. Surprisingly, Fis is lower in the actual founders than in the proposed founders. This suggests that the process for choosing founders outlined here does not minimize the potential level of inbreeding. Ultimately, the Banff NP population is still among the most variable plains bison herds in North America when compared to other sampled locations [
15]. Periodic assessments of genetic diversity in the Banff NP herd is recommended to monitor against the loss of genetic diversity in this recently founded population.
Banff NP has a smaller founding population size than most other recently founded bison populations for which we have genetic information, but the focus on genetic diversity in the founding animals allowed us to capture amounts of diversity similar to or greater than those seen in herds with a much larger number of founders (
Table 4). For example, despite the use of 57 randomly selected founders, the expected heterozygosity in the Hook Lake Wood Bison Recovery Project was 5% lower than its source herd in the Hook Lake Region [
35]. Grasslands NP was the most successful project for capturing genetic diversity in its source herd. It was also recently established from Elk Island NP, with the transfer of 71 animals in 2005. The large number of founders for the population captured most of the diversity of the source herd, with 97% of the alleles and 98% of the mean heterozygosity present in the founded herd [
34]. Some other examples also exist, but the loss of diversity from the founding event is conflated with the loss due to genetic drift since the inception of the founded herd. For example, the Pink Mountain population was founded from 48 Elk Island National Park bison in 1971. When sampled in the late 1990s, it only contained 92% of the genetic diversity in its source herd and 87% of the mean number of alleles [
15]. The Sturgeon River bison herd was founded from Elk Island NP in 1969 from between 11 and 22 founders, and has an even lower mean number of alleles [
22]. The wood bison population at Elk Island NP (note: this is located in a separate part of Elk Island NP, and is not related to the plains bison used as founders for the Banff NP population) and the Mackenzie herd were both founded from the Wood Buffalo National Park population in the 1960s with 11 and 16 founders respectively. Both are substantially less diverse than their source population [
15]. Another founding event fairly successful at capturing genetic diversity was the transfer of 29 bison from Elk Island NP (wood) to Hay Zama in 1984. When samples were collected in the early 2010s, expected heterozygosity was actually higher in the Hay Zama population than its source herd at Elk Island NP [
21]. However, only 90% of the Elk Island NP alleles were present in Hay Zama.
There are two important factors to consider from a genetic diversity perspective when founding new populations. The first is to capture as much genetic diversity as possible from the source population. It appears that by considering genetic diversity and relationships in choosing founders for the Banff NP population, and by ensuring that female founders had conceived prior to being separated from the larger Elk Island NP population, the Banff population is almost as genetically variable as its source population at Elk Island NP.
The second challenge is minimizing the loss of diversity through time from the recently founded population, especially while it is small. Genetic diversity can be quickly lost from small populations if individuals are lost before they produce offspring, or through differential reproductive success [
36], as has been documented for males in another recently established bison herd [
35]. In polygynous species like bison, there can be concern that some founding males produce few, if any, offspring, in which case their genetic diversity is not passed on to future generations and the effective size of the population is reduced. As all founding females were pregnant, they have already successfully contributed at least one offspring to the Banff NP population, which increases confidence that their genetic diversity will be represented in subsequent generations. However, we do not yet know the genetic contributions each of the six founding males have each made to the 50+ calves born into the population subsequent to 2017. This is the subject of ongoing research and will help flag potential concerns about inbreeding and loss of diversity in the nascent Banff NP population, which is growing rapidly (~90 animals in 2022), and is in good health with low (<1%) annual mortality.







{kind=link}