


New and Noteworthy Taxa of the Genus Dactylorhiza Necker ex Nevski (Orchidaceae Juss.) in Kazakhstan Flora and Its Response to Global Warming

 , , , and
, , , and
Abstract
:1. Introduction
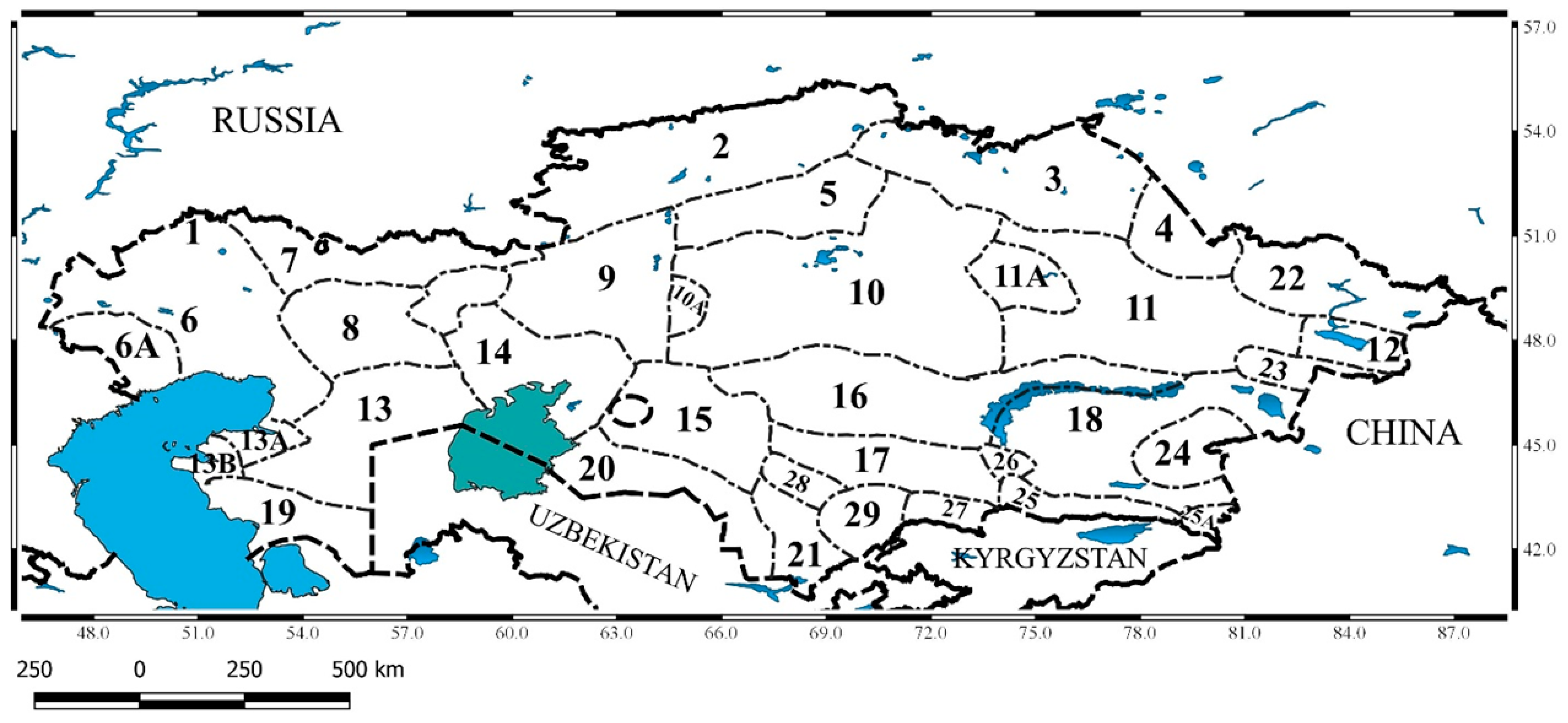
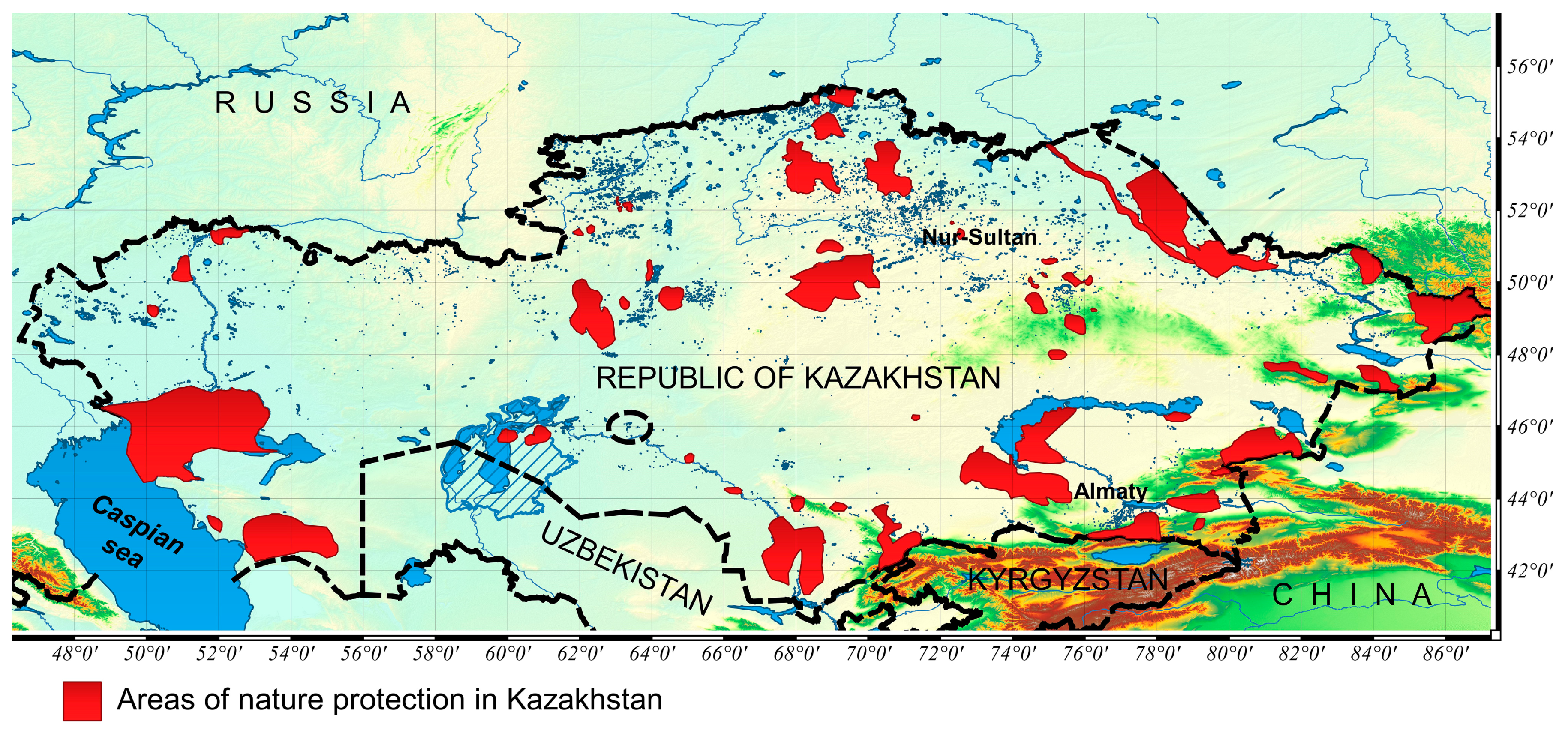
2. Materials and Methods
2.1. Taxonomic Treatment
2.2. Ecological Niche Modeling
2.3. Species Presence Data
2.4. Changes in the Distribution of Suitable Niches
3. Results
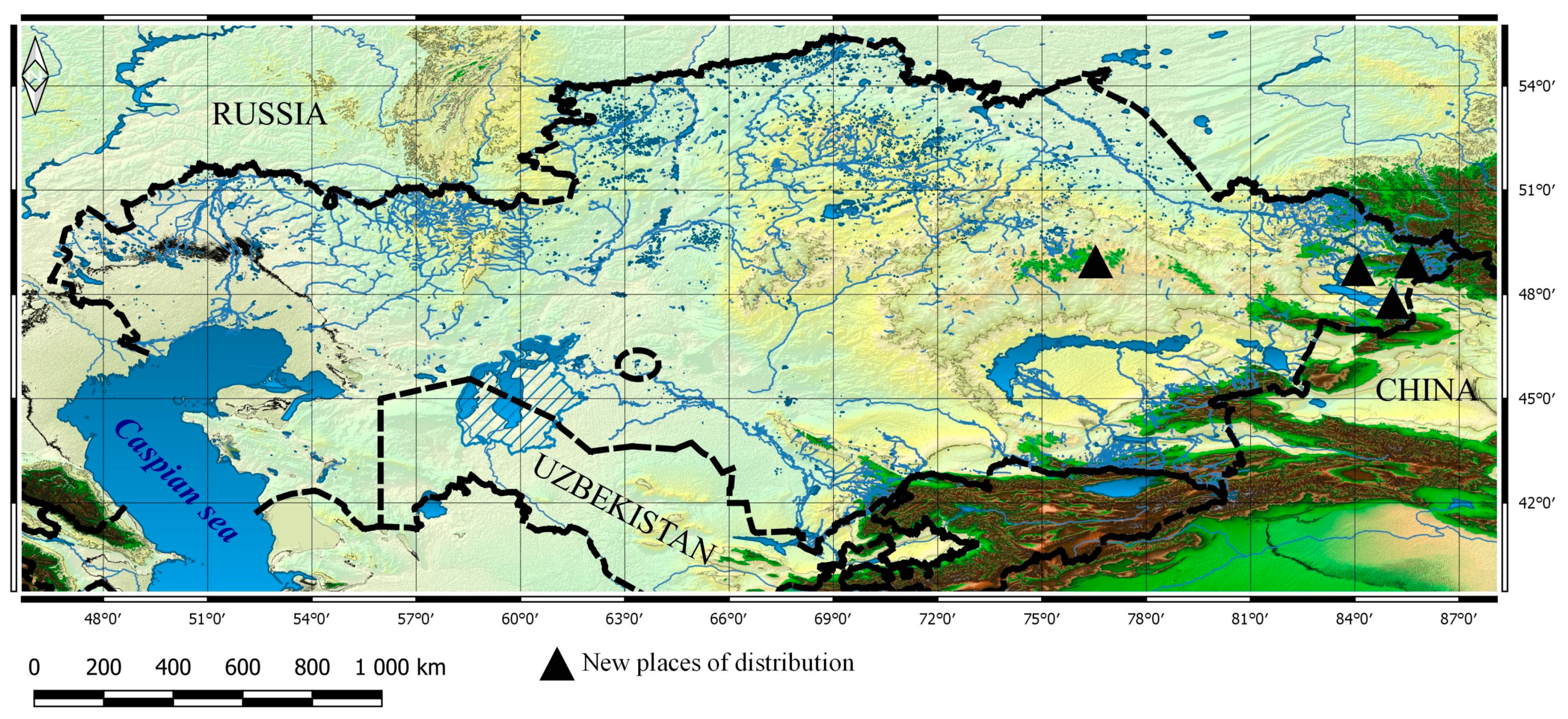
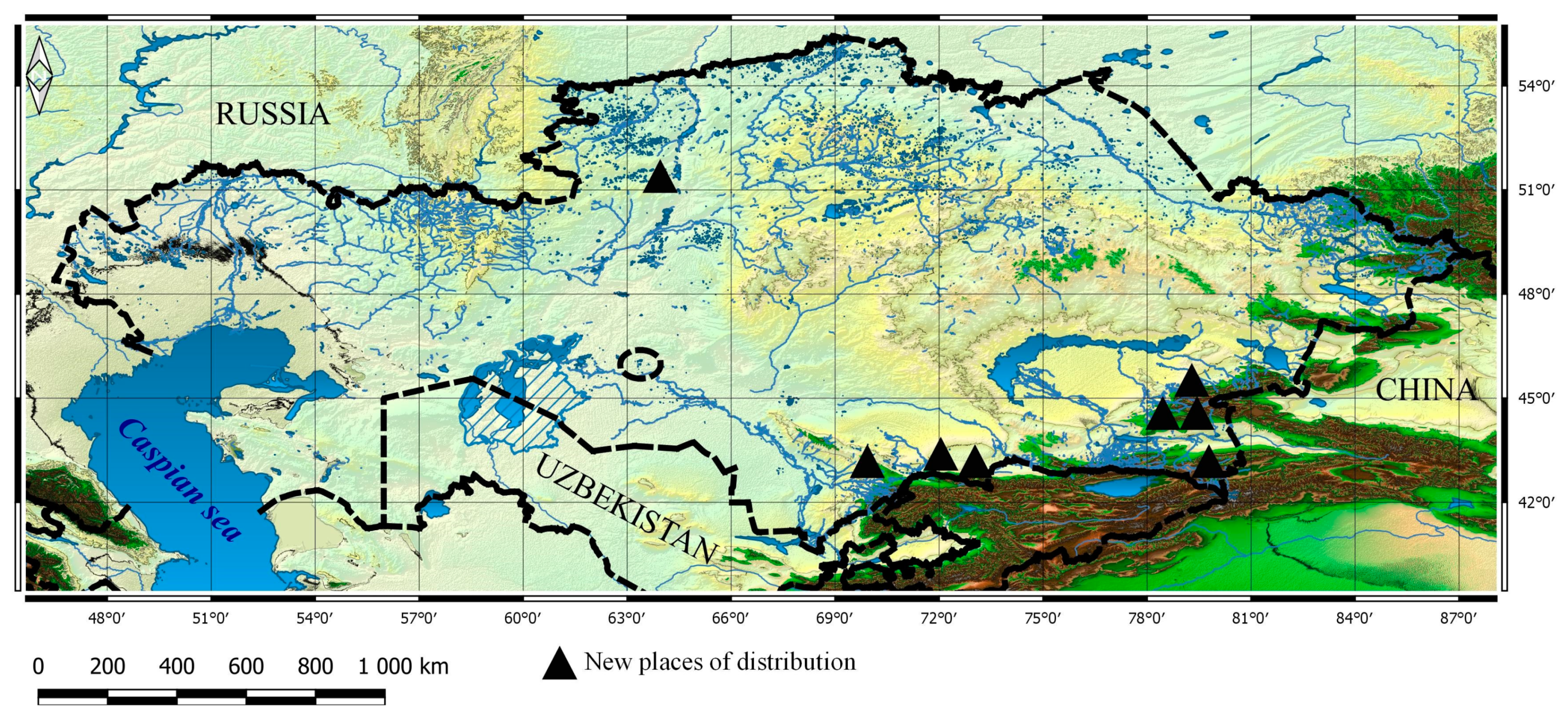
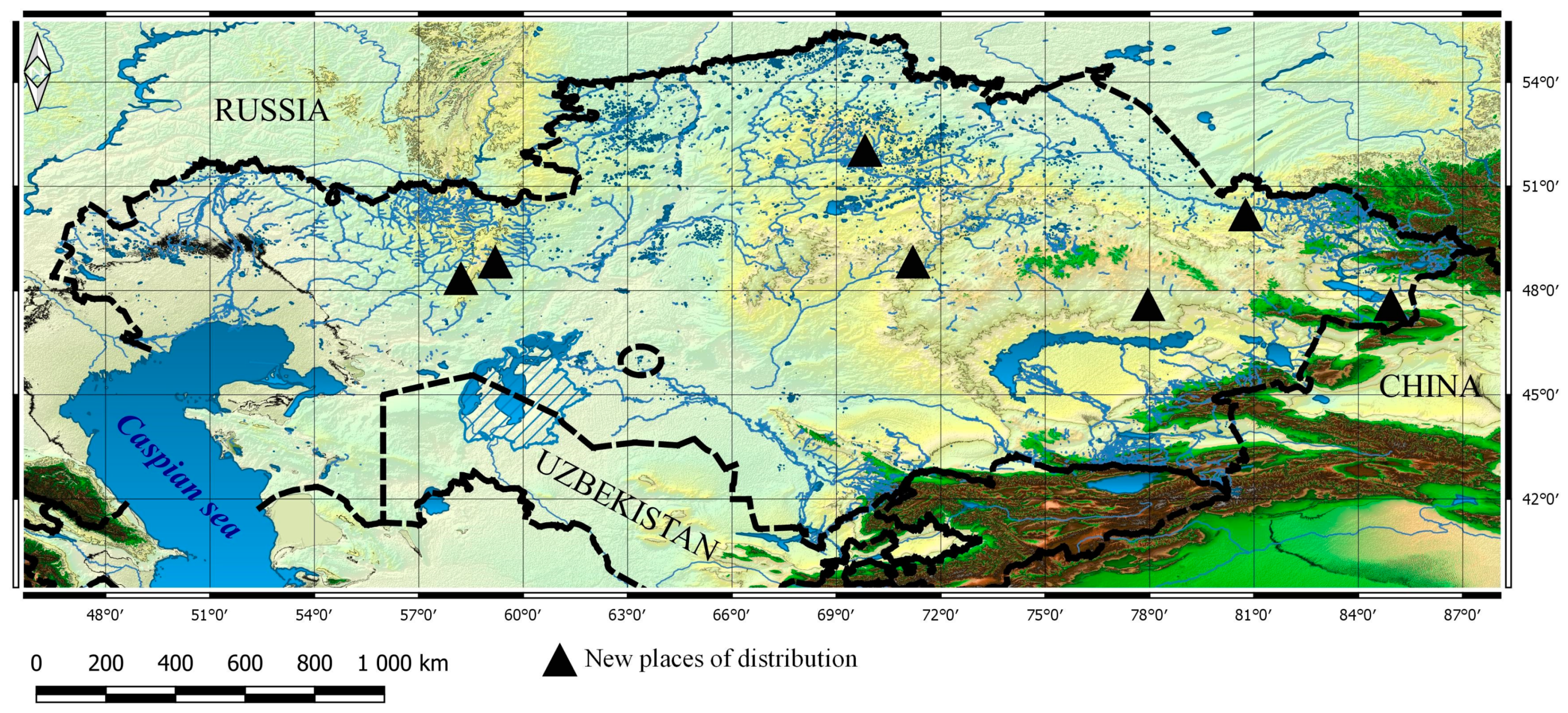
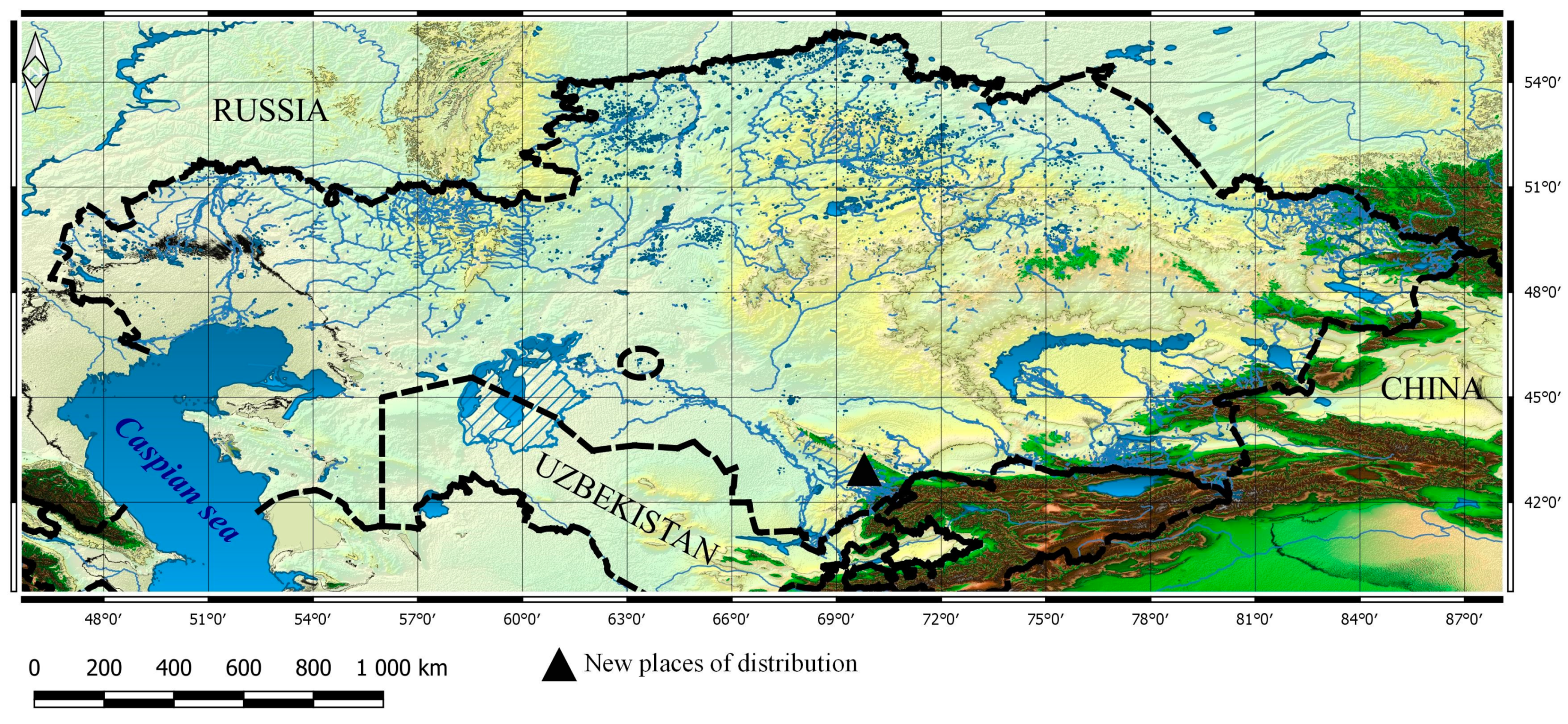
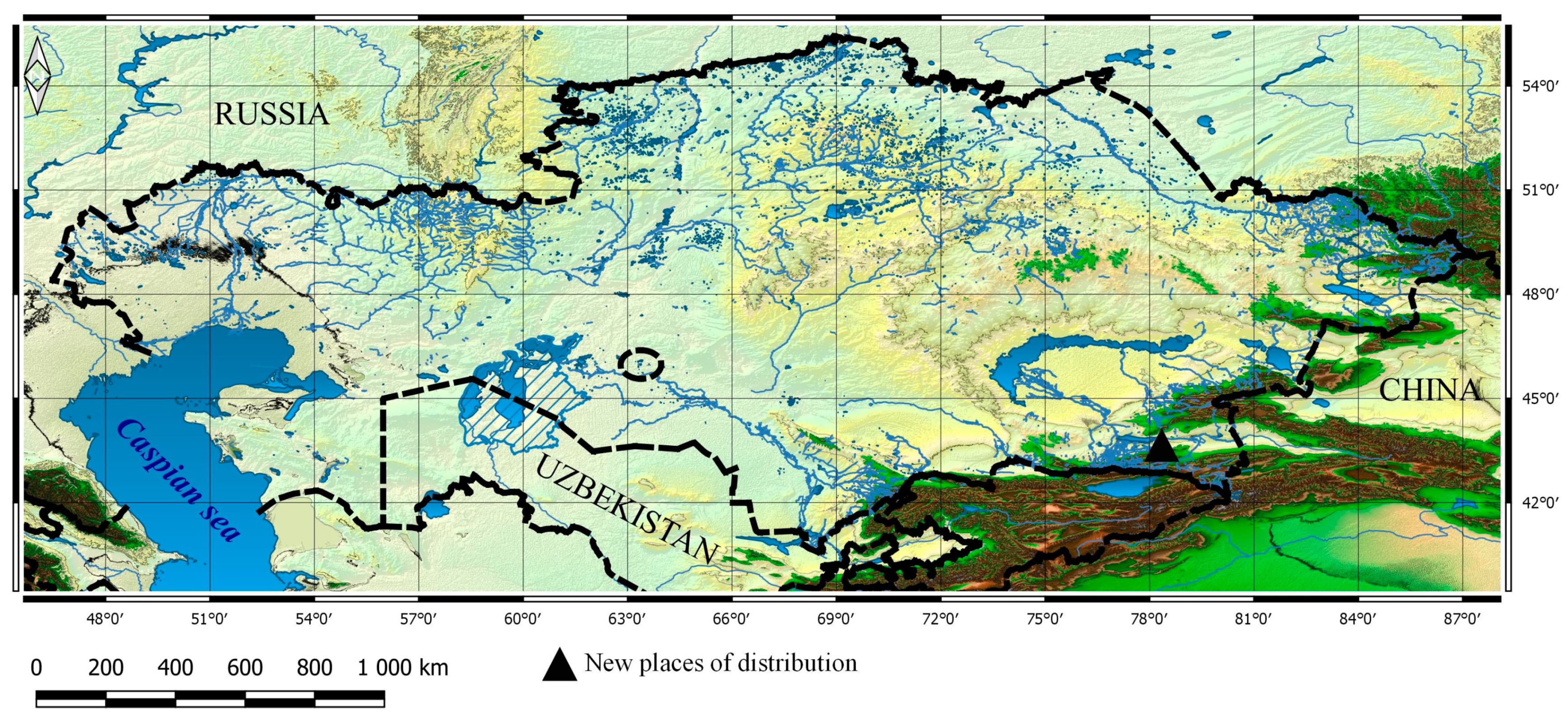
3.1. Taxa New for the Flora of Kazakhstan
3.1.1. Dactylorhiza fuchsii subsp. hebridensis (Wilmott) Soó, Nom. Nov. Gen. Dactylorhiza: 8 (1962). ≡ Orchis hebridensis Wilmott, J. Bot. 77: 192 (1939). ≡ Dactylorhiza hebridensis (Wilmott) Aver., Bot. J. 71: 92 (1986). ≡ Dactylorhiza maculata subsp. hebridensis (Wilmott) H.Baumann and Künkele, Mitt. Arbeitskreis Heimische Orchid. Baden-Württemberg 20: 631 (1988). ≡ Dactylorhiza fuchsii var. hebridensis (Wilmott) R.M. Bateman and Denholm, Watsonia 17: 340 (1989)
- = Orchis maculata L. var. meyeri Reichenb., Icon. Fl. Germ. 13–14: 67 (1851).
- = Orchis maculata var. cornubiensis Pugsley, J. Bot. 78: 178 (1940). ≡ Dactylorhiza fuchsii var. cornubiensis (Pugsley) Soó, Nom. Nov. Gen. Dactylorhiza: 8 (1962).
- = Dactylorhiza fuchsii f. alpina Landwehr, Orchideeën 37: 78 (1975). ≡ Dactylorhiza fuchsii var. alpina (Landwehr) R.M. Bateman and Denholm, Watsonia 17: 341 (1989).
3.1.2. Dactylorhiza × kerneri (Soó) Soó, Nom. Nov. Gen. Dactylorhiza: 9 (1962). ≡ Orchis × kerneri Soó Notizbl. Bot. Gart. Berlin-Dahlem 9: 904 (1926), as ‘kerneriorum’
- = Orchis × ambigua A. Kern., Verh. K. K. Zool.-Bot. Ges. Wien 15: 205 (1865), nom. illeg.
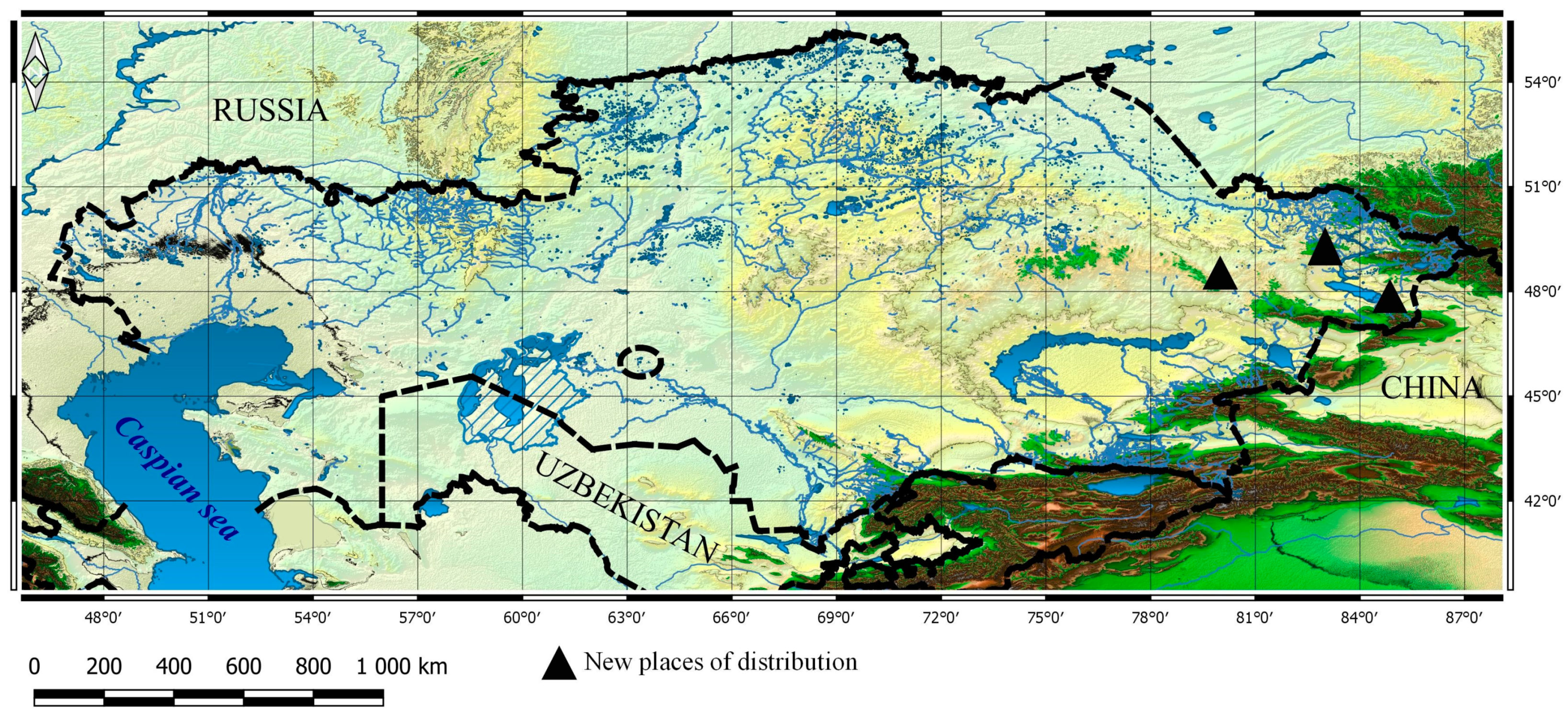
3.2. New Taxa for Particular Botanical and Geographical Areas
3.2.1. Dactylorhiza incarnata (L.) Soó, Nom. Nov. Gen., Dactylorhiza: 3 (1962). ≡ Orchis latifolia L., 941, excl. var. (1753); Kuznetsov and Pavlov, Fl. Kaz. 2: 271 (1958). ≡ Orchis incarnata L., Fl. Suec., ed. 2: 312 (1755). ≡ Orchis mixta var. incarnata (L.) Retz., Fl. Scand. Prodr.: 167 (1779). ≡ Orchis latifolia var. incarnata (L.) Coss. and Germ., Fl. Descr. Anal. Paris, ed. 2: 684 (1861). ≡ Orchis latifolia subsp. incarnata (L.) Hook.f., Student. Fl. Brit. Isl.: 353 (1870). ≡ Dactylorchis incarnata (L.) Verm., Stud. Dactylorch.: 65 (1947)
3.2.2. Dactylorhiza salina (Turcz. ex Lindl.) Soó, Nom. Nov. Gen. Dactylorhiza: 4 (1962). ≡ Orchis salina Turcz. ex Lindl., Gen. Sp. Orchid. Pl.: 259 (1835); Kuznetsov and Pavlov, Fl. Kaz. 2: 273 (1958). ≡ Orchis orientalis subsp. salina (Turcz. ex Lindl.) Klinge, Trudy Imp. S.-Peterburgsk. Bot. Sada 17(1): 185 (1898), nom. superfl. ≡ Dactylorchis salina (Turcz. ex Lindl.) Verm., Stud. Dactylorch.: 66 (1947). ≡ Dactylorhiza cruenta subsp. salina (Turcz. ex Lindl.) E. Nelson, Monogr. and Ikonogr. Orchid.-Gattung. Serapias, Aceras, Loroglossum, Barlia: 70 (1976)
3.2.3. Dactylorhiza umbrosa (Kar. and Kir.) Nevski, Tr. Bot. Inst. Ac. Sci. USSR, Ser. 1,4: 332 (1937). ≡ Orchis umbrosa Kar. and Kir., Bull. Soc. Imp. Naturalistes Moscou 15: 504 (1842); Kuznetsov and Pavlov, Fl. Kaz. 2: 272 (1958). ≡ Dactylorchis umbrosa (Kar. and Kir.) Wendelbo, Nytt Mag. Bot. 1: 24 (1952)
- = Orchis orientalis subsp. turcestanica Klinge, Trudy Imp. S.-Peterburgsk. Bot. Sada 17(1): 183 (1898). ≡ Orchis turkestanica (Klinge) Klinge ex B.Fedtsch., Russk. Bot. Zhurn. 1908: 191 (1908). ≡ Dactylorhiza incarnata subsp. turcestanica (Klinge) H. Sund., Europ. Medit. Orchid., ed. 3: 40 (1980).
3.2.4. Dactylorhiza magna (Czerniak.) Iconn., Nov. Syst. Visch. Rast. 9:303 (1972). ≡ Orchis magna Czerniak., Fl. Uzbekistan. 1: 546 (1941); Kuznetsov and Pavlov, Fl. Kaz. 2: 272 (1958). ≡ Dactylorhiza umbrosa subsp. magna (Czerniak.) Soó, Ann. Univ. Sci. Budapest. Rolando Eötvös, Sect. Biol. 11: 68 (1969)
- = Dactylorhiza baldshuanica Cherniak., Bot. Zhurn. (Moscow and Leningrad) 68: 534 (1983).
3.2.5. Dactylorhiza majalis subsp. baltica (Klinge) H.Sund., Europ. Medit. Orchid., ed. 3: 40 (1980). ≡ Orchis latifolia subsp. baltica Klinge, Trudy Imp. S.-Peterburgsk. Bot. Sada 17(1): 153, 170 (1898); Kuznetsov and Pavlov, Fl. Kaz.2: 271 (1958). ≡ Orchis latifolia f. baltica (Klinge) M.Schulze, Mitth. Thüring. Bot. Vereins, n.f., 17: 51 (1902). ≡ Orchis baltica (Klinge) A.Fuchs, Ber. Naturwiss. Vereins Schwaben Neuburg 42: 42 (1919). ≡ Dactylorchis baltica (Klinge) Verm., Stud. Dactylorch.: 67 (1947). ≡ Dactylorhiza latifolia subsp. baltica (Klinge) Soó, Nom. Nov. Gen. Dactylorhiza: 5 (1962). ≡ Dactylorhiza baltica (Klinge) N.I. Orlova, Konsp. Fl. Pskov. Obl., Leningrad: 57 (1970)
- = Orchis longifolia Neuman, Bot. Not. 1909: 241 (1909). ≡ Dactylorchis longifolia (Neuman) Verm., Stud. Dactylorch.: 67 (1947). ≡ Dactylorhiza latifolia var. longifolia (Neuman) Soó, Nom. Nov. Gen. Dactylorhiza: 5 (1962). ≡ Dactylorhiza longifolia (Neuman) Aver., Bot. Zhurn. (Moscow and Leningrad) 69: 875 (1984).
3.3. Ecological Niche Modeling
3.3.1. ENM Analysis Evaluation and Bioclimatic Variables Contribution
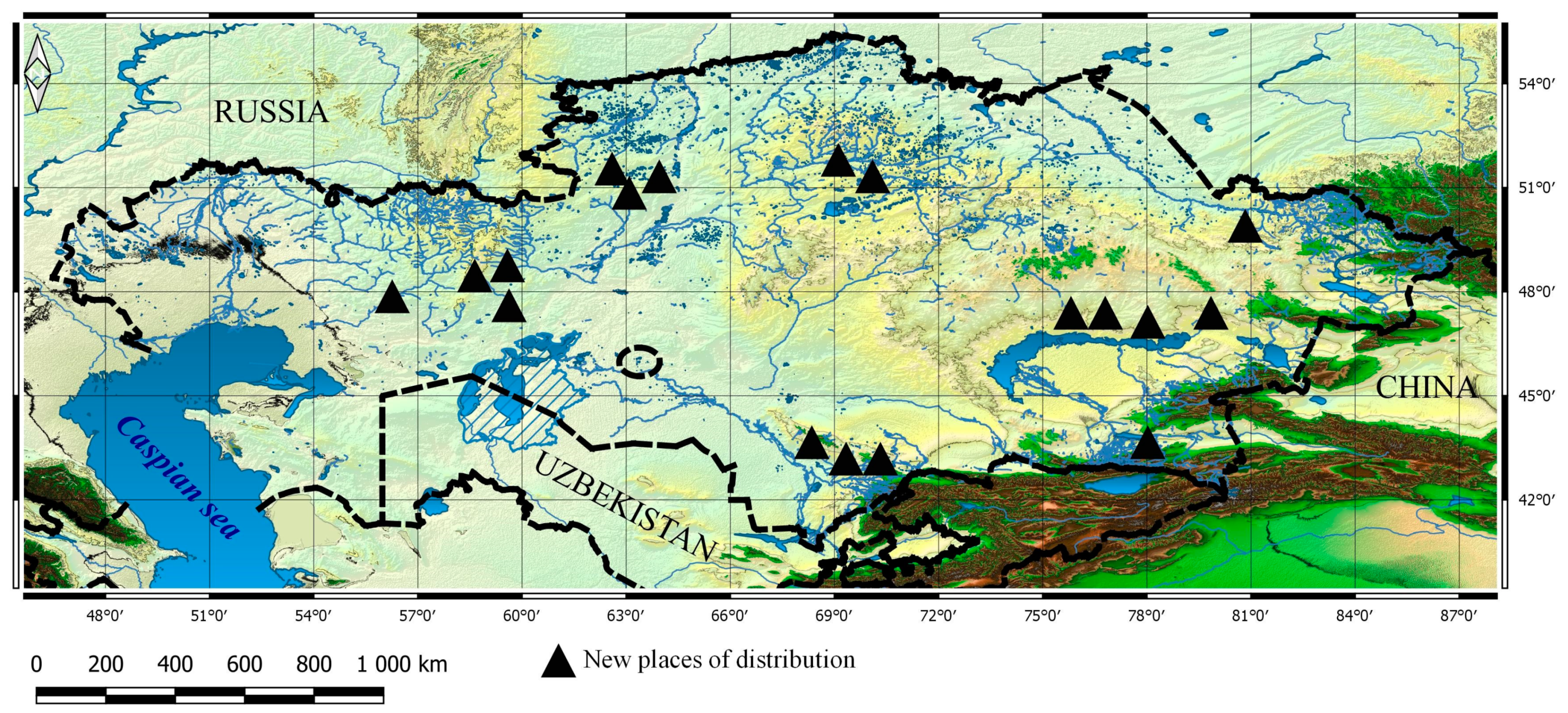
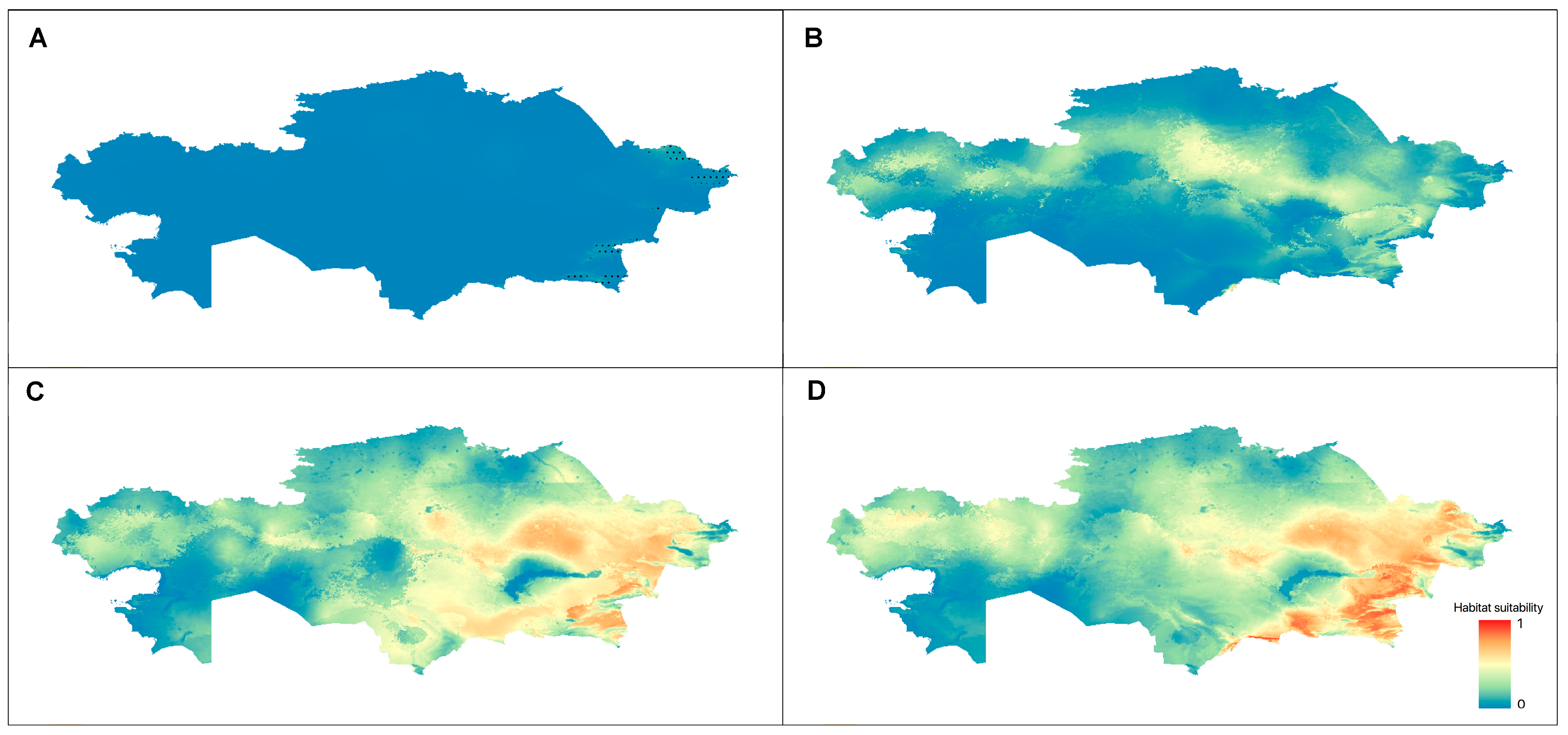
3.3.2. Current Distribution and the Ecological Niche Models for Taxa
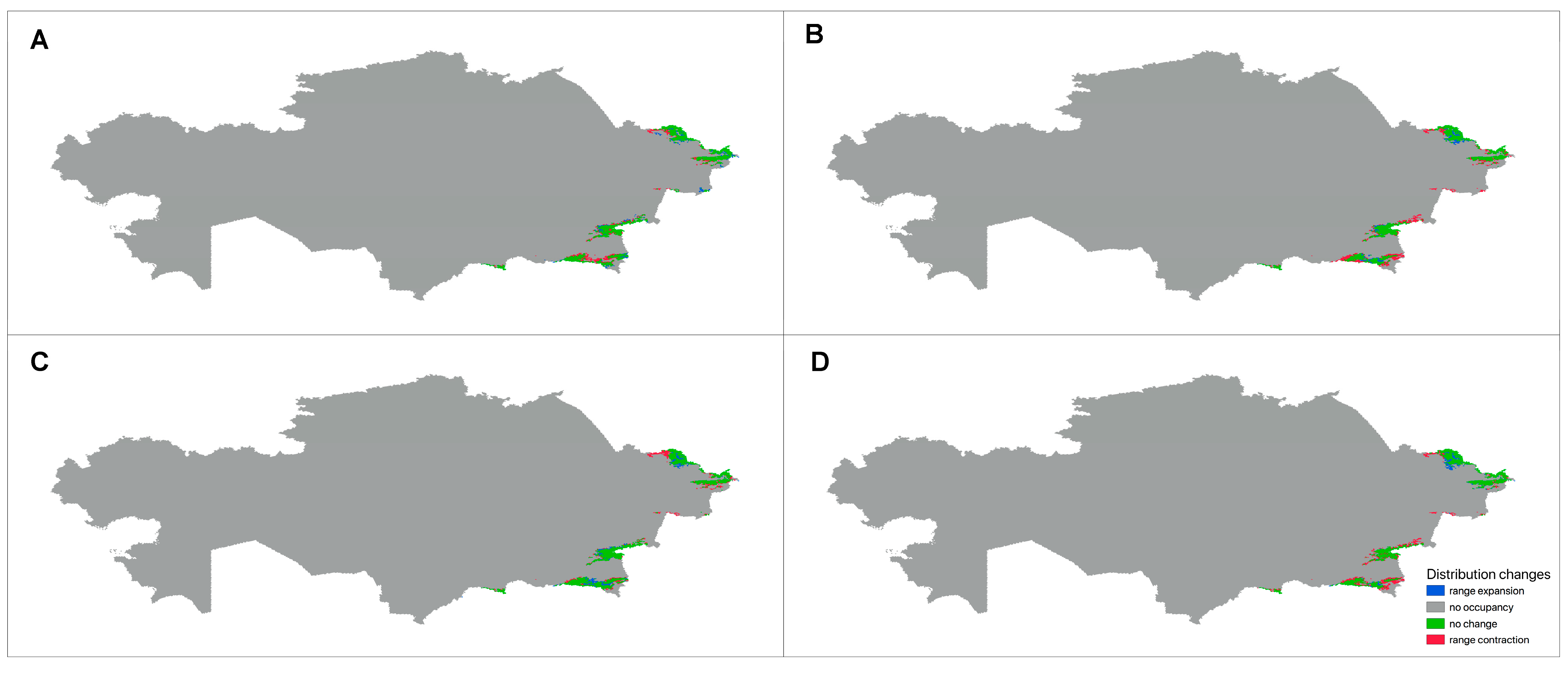
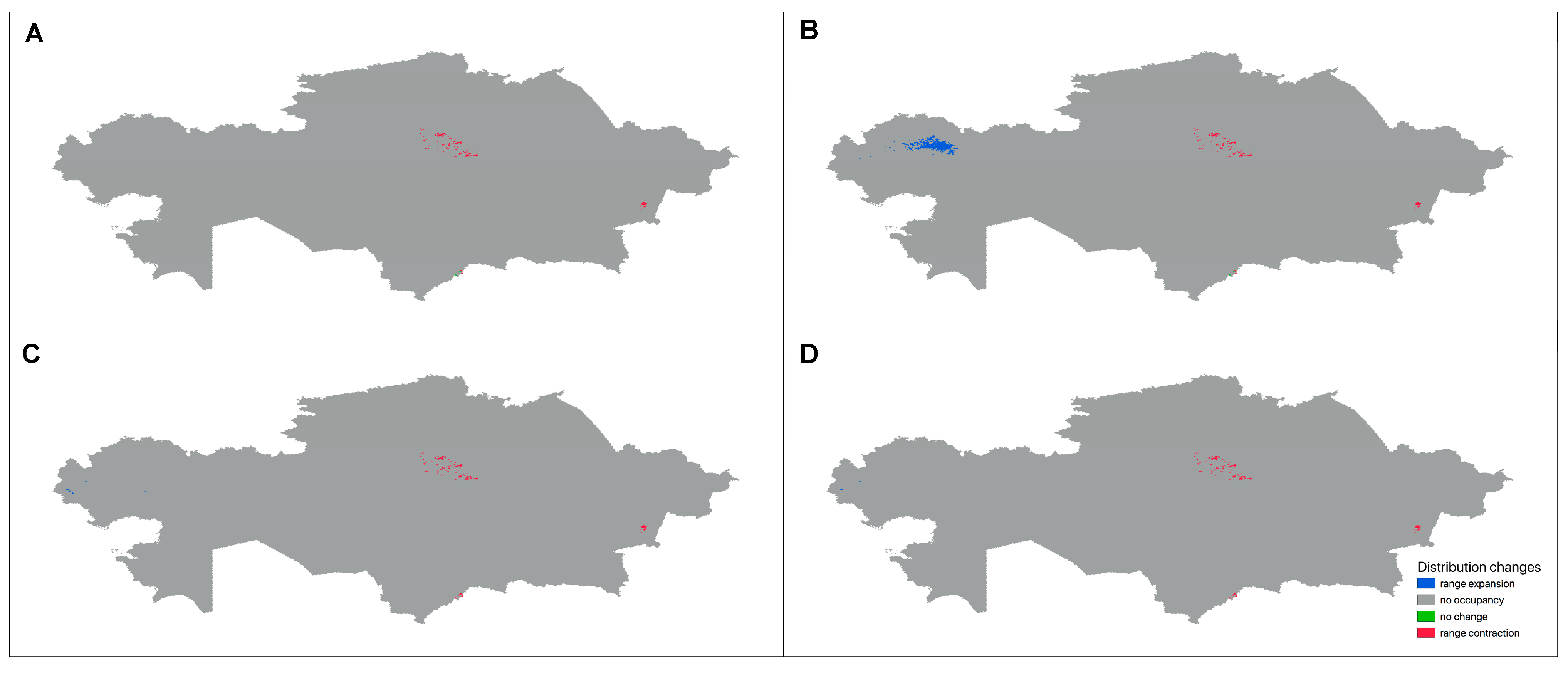
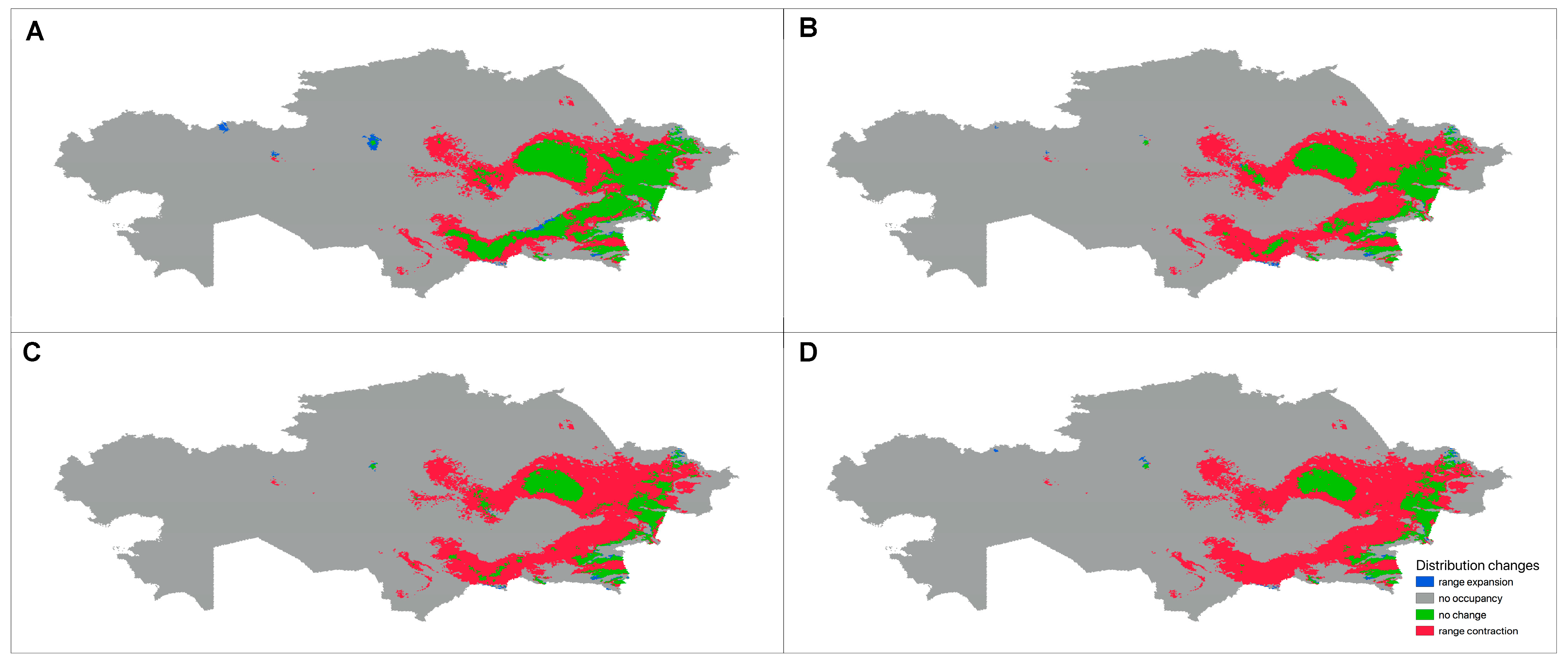
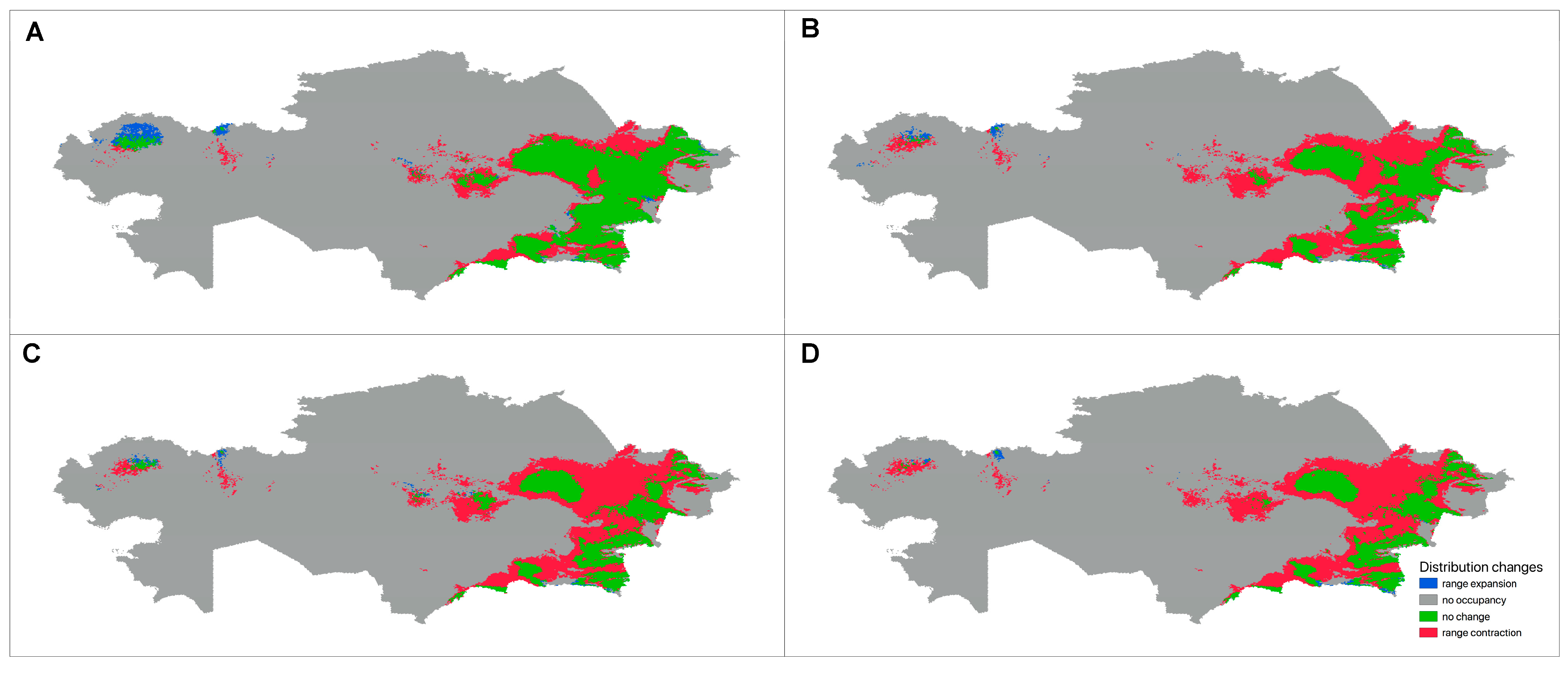
3.3.3. Future Changes in the Distribution
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Necker, N.J. Elementa Botanica 3, Neowedae ad Rhenum; Societatis Typographicae Neowedensis: Paris, Frane, 1790. [Google Scholar]
- Nevski, S.A. Materials for the flora of the Kugitang Mountains and its foothills. Tr. Bot. Inst. Akad. Nauk. SSSR Seriya 1 Flora I Sist. Vyss. Rastenii 1937, 4, 199–346. [Google Scholar]
- Nevski, S.A. Fam. 36, The Orchids-Orchidaceae Lindl; Komarov, V.L., Ed.; Publishers of Academy of Sciences: Cambridge, MA, USA, 1935; pp. 589–730. [Google Scholar]
- Taraška, V.; Batoušek, P.; Duchoslav, M.; Temsch, E.M.; Weiss-Schneeweiss, H.; Trávníček, B. Morphological variability, cytotype diversity, and cytogeography of populations traditionally called Dactylorhiza fuchsii in Central Europe. Plant Syst. Evol. 2021, 307, 1–21. [Google Scholar] [CrossRef]
- D’Emerico, S.; Cozzolino, S.; Pellegrino, G.; Pignone, D.; Scrugli, A. Karyotype structure, supernumerary chromosomes and heterochromatin distribution suggest a pathway of karyotype evolution in Dactylorhiza (Orchidaceae). Bot. J. Linn. Soc. 2002, 138, 85–91. [Google Scholar] [CrossRef] [Green Version]
- Averyanov, L. A Review of the Genus Dactylorhiza. In Orchid Biology—Reviews and Perspectives; Arditti, J., Ed.; V. Timber Press Inc.: Portland, OR, USA, 1990; pp. 159–206. [Google Scholar]
- Pedersen, H. Species concept and guidelines for infraspecific taxonomic ranking in Dactylorhiza (Orchidaceae). Nord. J. Bot. 1998, 18, 289–310. [Google Scholar] [CrossRef]
- Pillon, Y.; Fay, M.F.; Hedrén, M.; Bateman, R.M.; Devey, D.S.; Shipunov, A.B.; van der Bank, M.; Chase, M.W. Evolution and temporal diversification of western European polyploid species complexes in Dactylorhiza (Orchidaceae). Taxon 2007, 56, 1185–1208. [Google Scholar] [CrossRef] [Green Version]
- Brandrud, M.K.; Baar, J.; Lorenzo, M.T.; Athanasiadis, A.; Bateman, R.M.; Chase, M.W.; Hedrén, M.; Paun, O. Phylogenomic Relationships of Diploids and the Origins of Allotetraploids in Dactylorhiza (Orchidaceae). Syst. Biol. 2020, 69, 91–109. [Google Scholar] [CrossRef]
- Tropicos.org. Missouri Botanical Garden. Available online: https://tropicos.org (accessed on 4 May 2021).
- IPNI—The International Plant Names Index. Available online: http://www.ipni.org (accessed on 4 May 2021).
- Pillon, Y.; Fay, M.F.; Shipunov, A.B.; Chase, M.W. Species diversity versus phylogenetic diversity: A practical study in the tax-onomically difficult genus Dactylorhiza (Orchidaceae). Biol. Conserv. 2006, 4, 129. [Google Scholar] [CrossRef]
- Hedrén, M.; Birkedal, S.; de Boer, H.; Ghorbani, A.; Gravendeel, B.; Hansson, S.; Svensson, Å.; Zarre, S. Asymmetric contributions of seed and pollen to gene dispersal in the marsh orchid Dactylorhiza umbrosa in Asia Minor. Mol. Ecol. 2021, 30, 1791–1805. [Google Scholar] [CrossRef] [PubMed]
- Devos, N.; Raspe, O.; Jacquemart, A.L.; Tyteca, D. On the monophyly of Dactylorhiza Necker ex Nevski (Orchidaceae): Is Coelo-glossum viride (L.) Hartman a Dactylorhiza? Bot. J. Linn. Soc. 2006, 152, 261–269. [Google Scholar] [CrossRef] [Green Version]
- Shipunov, A.B.; Fay, M.F.; Pillon, Y.; Bateman, R.M.; Chase, M.W. Dactylorhiza (Orchidaceae) in European Russia: Combined molecular and morphological analysis. Am. J. Bot. 2004, 91, 1419–1426. [Google Scholar] [CrossRef]
- Shipunov, A.B.; Bateman, R.M. Geometric morphometrics as a tool for understanding Dactylorhiza (Orchidaceae) diversity in European Russia. Biol. J. Linn. Soc. 2005, 85, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Hedrén, M.; Fay, M.F.; Chase, M.W. Amplified fragment length polymorphisms (AFLP) reveal details of polyploid evolution in Dactylorhiza (Orchidaceae). Am. J. Bot. 2001, 88, 1868–1880. [Google Scholar] [CrossRef]
- Abdulina, S.A. Spisok Sosudistyh Rastenij Kazahstana; Kamelina, R.V., Ed.; Academy of Sciences of the Republic of Kazakhstan: Almaty, Kazakhstan, 1998; p. 187. [Google Scholar]
- The Red Book of Kazakhstan; Astana LTD: Astana, Kazakhstan, 2014; Volume 2, p. 452.
- Akzhigitova, N.I.; Brekle, Z.V.; Volkova, E.A.; Vinkler, G.; Vukhrer, V.; Ogar, Y.Y.; Rachkovskaya, Y.I.; Safronova, I.N.; Khramstov, V.N.; Makulbekova, G.B.; et al. Botanicheskaya Geografiya Kazakhstana i Srednei Azii (v Predelakh Pustynnoi Oblasti); Boston-Spectrum: St. Petersburg, Russia, 2003. [Google Scholar]
- Kamp, J.; Koshkin, M.A.; Bragina, T.M.; Katzner, T.E.; Milner-Gulland, E.J.; Schreiber, D.; Sheldon, R.; Shmalenko, A.; Smelansky, I.; Terraube, J.; et al. Persistent and novel threats to the biodiversity of Kazakhstan’s steppes and semi-deserts. Biodivers. Conserv. 2016, 25, 2521–2541. [Google Scholar] [CrossRef]
- Hu, Y.; Han, Y.; Zhang, Y. Land desertification and its influencing factors in Kazakhstan. J. Arid. Environ. 2020, 180, 104203. [Google Scholar] [CrossRef]
- UNECE. Environmental Performance Review; United Nations: Geneva, Switzerland, 2019. [Google Scholar]
- BIFOR. Biodiversity Assessment for Kazakhstan; Chemonics International INC.: Washington, DC, USA, 2001. [Google Scholar]
- Wani, I.; Kumar, V.; Verma, S.; Jan, A.T.; Rather, I. Dactylorhiza hatagirea (D. Don) Soo: A Critically Endangered Perennial Orchid from the North-West Himalayas. Plants 2020, 9, 1644. [Google Scholar] [CrossRef] [PubMed]
- Kelly, A.E.; Goulden, M.L. Rapid shifts in plant distribution with recent climate change. Proc. Natl. Acad. Sci. USA 2008, 105, 11823–11826. [Google Scholar] [CrossRef] [Green Version]
- Dainese, M.; Aikio, S.; Hulme, P.E.; Bertolli, A.; Prosser, F.; Marini, L. Human disturbance and upward expansion of plants in a warming climate. Nat. Clim. Chang. 2017, 7, 577–580. [Google Scholar] [CrossRef]
- Lamprecht, A.; Semenchuk, P.R.; Steinbauer, K.; Winkler, M.; Pauli, H. Climate change leads to accelerated transformation of high-elevation vegetation in the central Alps. New Phytol. 2018, 220, 447–459. [Google Scholar] [CrossRef]
- Geppert, C.; Perazza, G.; Wilson, R.J.; Bertolli, A.; Prosser, F.; Melchiori, G.; Marini, L. Consistent population declines but idi-osyncratic range shifts in Alpine orchids under global change. Nat. Commun. 2020, 11, 5835. [Google Scholar] [CrossRef]
- Kolanowska, M.; Rewicz, A.; Baranow, P. Ecological niche modeling of the pantropical orchid Polystachya concreta (Orchidaceae) and its response to climate change. Sci. Rep. 2020, 10, 1–17. [Google Scholar] [CrossRef]
- Kolanowska, M.; Jakubska-Busse, A. Is the lady’s-slipper orchid (Cypripedium calceolus) likely to shortly become extinct in Europe?—Insights based on ecological niche modelling. PLoS ONE 2020, 15, e0228420. [Google Scholar] [CrossRef]
- Shrestha, B.; Tsiftsis, S.; Chapagain, D.J.; Khadka, C.; Bhattarai, P.; Kayastha Shrestha, N.; Kolanowska, M.; Kindlmann, P. Suitability of Habitats in Nepal for Dactylorhiza hatagirea Now and under Predicted Future Changes in Cli-mate. Plants 2021, 10, 467. [Google Scholar] [CrossRef] [PubMed]
- Kolanowska, M.; Rewicz, A.; Nowak, S. Significant habitat loss of the black vanilla orchid (Nigritella nigra s.l., Orchidaceae) and shifts in its pollinators availability as results of global warming. Glob. Ecol. Conserv. 2021, 27, e01560. [Google Scholar] [CrossRef]
- Kolanowska, M.; Kras, M.; Lipińska, M.; Mystkowska, K.; Szlachetko, D.L.; Naczk, A.M. Global warming not so harmful for all plants—response of holomycotrophic orchid species for the future climate change. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konowalik, K.; Kolanowska, M. Climatic niche shift and possible future spread of the invasive South African Orchid Disa bracteata in Australia and adjacent areas. PeerJ 2018, 6, e6107. [Google Scholar] [CrossRef] [Green Version]
- Thiers, B. Continuously Updated Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff. New York Botanical Garden’s Virtual Herbarium. Available online: http://sweetgum.nybg.org/science/ih/ (accessed on 28 December 2021).
- Seregin, A.P. (Ed.) Moscow Digital Herbarium: Electronic Resource; Moscow State University: Moscow, Russia, 2020; Available online: https://plant.depo.msu.ru/ (accessed on 10 January 2020).
- Pavlov, N.V. Flora of Kazakhstan; Academy of Sciences of the Kazakh SSR: Alma-Ata, Kazakhstan, 1956; Volume 1, p. 353. [Google Scholar]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Booth, T.H.; Nix, H.A.; Busby, J.R.; Hutchinson, M.F. BIOCLIM: The first species distribution modelling package, its early ap-plications and relevance to most current MAXENT studies. Divers. Distrib. 2014, 20, 1–9. [Google Scholar] [CrossRef]
- Barve, N.; Barve, V.; Jiménez-Valverde, A.; Lira-Noriega, A.; Maher, S.P.; Townsend Peterson, A.; Soberon, J.; Villalobos, F. The crucial role of the accessible area in ecological niche modeling and species distribution modeling. Ecol. Model. 2011, 222, 1810–1819. [Google Scholar] [CrossRef]
- Voldoire, A.; Saint-Martin, D.; Sénési, S.; Decharme, B.; Alias, A.; Chevallier, M.; Colin, J.; Guérémy, J.-F.; Michou, M.; Moine, M.-P.; et al. Evaluation of CMIP6 DECK experiments with CNRM-CM6-1. J. Adv. Mod. Earth Syst. 2019, 11, 2177–2213. [Google Scholar] [CrossRef] [Green Version]
- Eyring, V.; Bony, S.; Meehl, G.A.; Senior, C.A.; Stevens, B.; Stouffer, R.J.; Taylor, K.E. Overview of the Coupled Model Inter-comparison Project Phase 6 (CMIP6) experimental design and organization. Geosci. Model Dev. 2016, 9, 1937–1958. [Google Scholar] [CrossRef] [Green Version]
- Meinshausen, M.; Nicholls, Z.R.J.; Lewis, J.; Gidden, M.J.; Vogel, E.; Freund, M.; Beyerle, U.; Gessner, C.; Nauels, A.; Bauer, N.; et al. The shared socio-economic pathway (SSP) greenhouse gas concentrations and their extensions to 2500. Geosci. Model Dev. 2020, 13, 3571–3605. [Google Scholar] [CrossRef]
- Petrie, R.; Denvil, S.; Ames, S.; Levavasseur, G.; Fiore, S.; Allen, C.; Antonio, F.; Berger, K.; Bretonnière, P.-A.; Cinquini, L.; et al. Coordinating an operational data distribution network for CMIP6 data. Geosci. Model Dev. 2021, 14, 629–644. [Google Scholar] [CrossRef]
- Riahi, K.; Van Vuuren, D.P.; Kriegler, E.; Edmonds, J.; O’Neill, B.C.; Fujimori, S.; Bauer, N.; Calvin, K.; Dellink, R.; Fricko, O.; et al. The Shared Socioeconomic Pathways and their energy, land use, and greenhouse gas emissions implications: An overview. Glob. Environ. Chang. 2017, 42, 153–168. [Google Scholar] [CrossRef] [Green Version]
- Rogelj, J.; Popp, A.; Calvin, K.V.; Luderer, G.; Emmerling, J.; Gernaat, D.; Fujimori, S.; Strefler, J.; Hasegawa, T.; Marangoni, G.; et al. Scenarios towards limiting global mean temperature increase below 1.5 °C. Nat. Clim. Chang. 2018, 8, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Mason, S.J.; Graham, N.E. Areas beneath the relative operating characteristics (ROC) and relative operating levels (ROL) curves: Statistical significance and interpretation. Q. J. R. Meteorol. Soc. 2002, 128, 2145–2166. [Google Scholar] [CrossRef]
- Evangelista, P.H.; Kumar, S.; Stohlgren, T.J.; Jarnevich, C.S.; Crall, A.W.; Norman, J.B., III; Barnett, D.T. Modelling invasion for a habitat generalist and a specialist plant species. Divers. Distrib. 2008, 14, 808–817. [Google Scholar] [CrossRef]
- ESRI. ArcGIS Desktop: Release 10; Environmental Systems Research Institute: Redlands, CA, USA, 2021. [Google Scholar]
- QGIS Development Team. QGIS Geographic Information System. Open Source Geospatial Foundation Project. Available online: http://qgis.osgeo.org (accessed on 13 January 2020).
- GBIF. GBIF Home Page. Available online: https://www.gbif.org (accessed on 13 January 2020).
- Reddy, S.; Dávalos, L.M. Geographical sampling bias and its implications for conservation priorities in Africa. J. Biogeogr. 2003, 30, 1719–1727. [Google Scholar] [CrossRef]
- Reese, G.C.; Wilson, K.R.; Hoeting, J.A.; Flather, C.H. Factors Affecting Species Distribution Predictions: A Simulation Modeling Experiment. Ecol. Appl. 2005, 15, 554–564. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Dudík, M.; Elith, J.; Graham, C.H.; Lehmann, A.; Leathwick, J.; Ferrier, S. Sample selection bias and presence-only distribution models: Implications for background and pseudo-absence data. Ecol. Appl. 2009, 19, 181–197. [Google Scholar] [CrossRef] [Green Version]
- Hijmans, R.J. Cross-validation of species distribution models: Removing spatial sorting bias and calibration with a null model. Ecology 2012, 93, 679–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boria, R.A.; Olson, L.E.; Goodman, S.M.; Anderson, R.P. Spatial filtering to reduce sampling bias can improve the performance of ecological niche models. Ecol. Model. 2014, 275, 73–77. [Google Scholar] [CrossRef]
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Peterson, A.T. Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar. J. Biogeogr. 2007, 34, 102–117. [Google Scholar] [CrossRef]
- Wisz, M.S.; Hijmans, R.J.; Li, J.; Peterson, A.T.; Graham, C.H.; Guisan, A.; NCEAS Predicting Species Distributions Working Group. Effects of sample size on the performance of species distribution models. Divers. Distrib. 2008, 14, 763–773. [Google Scholar] [CrossRef]
- Brown, J.L. SDMtoolbox: A python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods Ecol. Evol. 2014, 5, 694–700. [Google Scholar] [CrossRef]
- Averyanov, L. Taxonomic and nomenclature variations in the genus Dactylorhiza (Orchidaceae). Bot. Zhurn. 1984, 71, 92–93. [Google Scholar]
- Pavlov, N.V. Flora of Kazakhstan; Academy of Sciences of the Kazakh SSR: Alma-Ata, Kazakhstan, 1958; Volume 2, p. 292. [Google Scholar]
- Averyanov, L.V. Synopsis of the genus Dactylorhiza Neck. ex Nevski (Orchidaceae). Novosti Sist. Vyssh. Rast. 1991, 28, 33–42. [Google Scholar]
- Stepanova, E.F. Vegetation and Flora of the Tarbagatai Ridge; Academy of Sciences of Kazakhstan: Alma-Ata, Kazakhstan, 1962; p. 433. [Google Scholar]
- Kadenova, A.B.; Kamkin, V.A.; Erzhanov, N.T.; Kamkina, E.V. Flora I Rastitel’nost’ Bajanaul’skogo Gosudarstvennogo Nacional’nogo Prirodnogo Parka: Monografija; Kereku: Pavlodar, Kazakhstan, 2008; p. 383. [Google Scholar]
- Karmysheva, N.H. Flora and Vegetation of the Western Spurs of Talas Alatau; Academy of Sciences of Kazakhstan: Alma-Ata, Kazakhstan, 1982; p. 160. [Google Scholar]
- Bajtenov, M.S. Alpine Flora of the Northern Tien Shan; Academy of Sciences of Kazakhstan: Alma-Ata, Kazakhstan, 1965; p. 232. [Google Scholar]
- Karmysheva, N.H. Flora I Rastitel’nost’ Zapovednika Aksu-Dzhabagly (Talasskij Alatau). Flora and Vegetation of the Reserve Ak-su-Dzhabagly (Talas Alatau); Academy of Sciences of Kazakhstan: Alma-Ata, Kazakhstan, 1973; p. 173. [Google Scholar]
- Nikitina, E.V. Flora and Vegetation of Pastures and Hayfields of the Kyrgyz Ala-Too ridge; Academy of Sciences of Kirghiz SSR: Frunze, Kyrgyzstan, 1962; p. 285. [Google Scholar]
- Goloskokov, V.P. Flora and Vegetation of the Highlands of Ile Alatau; Academy of Sciences of Kazakhstan: Alma-Ata, Kazakhstan, 1949; p. 202. [Google Scholar]
- Karamysheva, Z.V.; Rachkovskaja, E.I. Botanical Geography of the Steppe of Central Kazakhstan; Nauka, Komarov Botanical Institute: Sankt-Peterburg, Russia, 1973; p. 279. [Google Scholar]
- Ishmuratova, M.J.; Myrzaly, G.Z.; Ivlev, V.I.; Matveev, A.N. Flora of Ulytau Mountains (Central Kazakhstan); Academy of Sciences of Kazakhstan: Karagandy, Kazakhstan, 2016; p. 127. [Google Scholar]
- Grubov, V.I.; Egorova, T.V. (Eds.) Plants of Central Asia. In The Materials of the Botanical Institute; V.L. Komarov Academy of Sciences of the USSR. Liliace-ae-Orchidaceae; Nauka, Komarov Botanical Institute: Sankt-Peterburg, Russia, 1977; Volume 7, p. 138. [Google Scholar]
- Nelina, N.V.; Kudabaeva, G.M.; Veselova, P.V.; Bilibaeva, B.K. The Species Composition of the Flora of the Kaindy and Sugaty Gorges of the Western Part of the Kyrgyz Alatau. In XVI Mezhdunarodnaja Nauchno-Prakticheskaja Konferencija; Problemy Botaniki Juzhnoj Sibiri i Mongolii; ISSN National Centre for the Russian Federation: Moscow, Russia, 2017; Volume 16, pp. 42–45. [Google Scholar]
- Ju, V. Perezhogin Ujazvimye jelementy flory Kostanajskoj oblasti. In Biologicheskoe Raznoobrazie Aziatskih Stepej, Materialy III Mezhdunar.Nauchn konf; Braginoj, T.M., Ed.; KGPI: Kostanaj, Kazakhstan, 2017. [Google Scholar]
- Muhtubaeva, S.K.; Nelina, N.V.; Sitpaeva, G.M.; Kudabaeva, P.V.; Veselova, B.K.; Bilibaeva, A. Zhumadilova Redkie, jendemichnye, reliktovye i ischezajushhie vidy rastenij severnogo Tjan’-Shanja (Kungej i Kirgizskij Alatau). NAS RK 2017, 6, 103–110. [Google Scholar]
- Ivashhenko, A.A. Spisok flory Sajram-Ugamskogo Gosudarstvennogo nacional’nogo prirodnogo parka (Kazakhstan). Botanicheskie issledovanija Sibiri i Kazahstana. Vyp 2020, 26, 52–63. [Google Scholar]
- Sultangazina, G.Z.; Hrustaleva, I.A.; Kuprijanov, A.N. Redkie rastenija nacional’nogo prirodnogo parka. Vestn. KazNU 2013, 3, 264–270. [Google Scholar]
- Sultangazina, G.Z.; Hrustaleva, I.A.; Kuprijanov, A.N.; Adekenov, S.M. Flora Nacional’nogo Parka; SO RAN: Novosibirsk, Russia, 2014; p. 238. [Google Scholar]
- Sultangazina, G.Z.; Kuprijanov, A.N. Floristicheskie nahodki na territorii nacional’nogo parka. Vestnik KemGU 2012, 1, 23–26. [Google Scholar]
- Besnard, G.; Gaudeul, M.; Lavergne, S.; Muller, S.; Rouhan, G.; Sukhorukov, A.P.; Vanderpoorten, A.; Jabbour, F. Herbarium-based science in the twenty-first century. Bot. Lett. 2018, 165, 323–327. [Google Scholar] [CrossRef] [Green Version]
- Willis, C.G.; Ellwood, E.R.; Primack, R.B.; Davis, C.C.; Pearson, K.D.; Gallinat, A.S.; Yost, J.M.; Nelson, G.; Mazer, S.J.; Rossington, N.L.; et al. Old Plants, New Tricks: Phenological Research Using Herbarium Specimens. Trends Ecol. Evol. 2017, 32, 531–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nualart, N.; Ibáñez, N.; Soriano, I.; López-Pujol, J. Assessing the relevance of herbarium collections as tools for conservation bi-ology. Bot. Rev. 2017, 83, 303–325. [Google Scholar] [CrossRef]
- Heberling, J.M.; Prather, L.A.; Tonsor, S.J. The Changing Uses of Herbarium Data in an Era of Global Change: An Overview Using Automated Content Analysis. Bioscience 2019, 69, 812–822. [Google Scholar] [CrossRef]
- Kim, K.C.; Byrne, L.B. Biodiversity loss and the taxonomic bottleneck: Emerging biodiversity science. Ecol. Res. 2006, 21, 794–810. [Google Scholar] [CrossRef] [Green Version]
- Schlick-Steiner, B.C.; Steiner, F.M.; Seifert, B.; Stauffer, C.; Christian, E.; Crozier, R.H.; Shewan, J.M.; McMeekin, T.A.; Palleroni, N.J.; Doudoroff, M.; et al. Integrative Taxonomy: A Multisource Approach to Exploring Biodiversity. Annu. Rev. Entomol. 2010, 55, 421–438. [Google Scholar] [CrossRef]
- Efimov, P.; Philippov, E.G.; Krivenko, D. Allopolyploid speciation in Siberian Dactylorhiza (Orchidaceae, Orchidoideae). Phytotaxa 2016, 258, 101–120. [Google Scholar] [CrossRef]
- Bolshanik, P.V.; Vladimirovich, B.P.; Kusnezova, S.B.; Borisovna, K.S.; Karaseva, D.D.; Dmitrievna, K.D. Natural features of the orchid refugium on the territory of the Natural Park”Samarovsky Chugas”. Environ. Dyn. Glob. Clim. Change 2021, 12, 15–26. [Google Scholar] [CrossRef]
- Shipunov, A.B.; Fay, M.F.; Chase, M.W. Evolution of Dactylorhiza baltica (Orchidaceae) in European Russia: Evidence from molecular markers and morphology. Bot. J. Linn. Soc. 2005, 147, 257–274. [Google Scholar] [CrossRef] [Green Version]
- Efimov, P.G. The study of ISSR polymorphism of Dactylorhiza baltica, D. fuchsii and D. incarnata (Orchidaceae) from the North-West European Russia. Bot. Zhurnal 2012, 97, 751–761. [Google Scholar]
- Pedersen, H.A.; Hedrén, M. On the distinction of Dactylorhiza baltica and D. pardalina (Orchidaceae) and the systematic affinities of geographically intermediate populations. Nord. J. Bot. 2010, 28, 1–12. [Google Scholar] [CrossRef]
- Heijmans, M.M.; Mauquoy, D.; van Geel, B.; Berendse, F. Long-term effects of climate change on vegetation and carbon dynamics in peat bogs. J. Veg. Sci. 2008, 19, 307–320. [Google Scholar] [CrossRef] [Green Version]
- Weltzin, J.F.; Pastor, J.; Harth, C.; Bridgham, S.D.; Updegraff, K.; Chapin, C.T. Response of bog and fen plant communities to warming and water-table manipulations. Ecology 2000, 81, 3464–3478. [Google Scholar] [CrossRef]
- Naczk, A.M.; Kolanowska, M. Glacial Refugia and Future Habitat Coverage of Selected Dactylorhiza Representatives (Orchi-daceae). PLoS ONE 2015, 10, e0143478. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, M.; Schatz, B.; Picó, F.X.; Passalacqua, N.G.; Fay, M.F.; Carey, P.D.; Jeltsch, F. Phylogeography and genetic structure of the orchid Himantoglossum hircinum (L.) Spreng. Across its European central–marginal gradient. J. Biogeogr. 2009, 36, 2353–2365. [Google Scholar]
- Pfeifer, M.; Passalacqua, N.G.; Bartram, S.; Schatz, B.; Croce, A.; Carey, P.D.; Kraudelt, H.; Jeltsch, F. Conservation priorities differ at opposing species borders of a European orchid. Biol. Conserv. 2010, 143, 2207–2220. [Google Scholar] [CrossRef]
- Wani, I.A.; Verma, S.; Mushtaq, S.; Alsahli, A.A.; Alyemeni, M.N.; Tariq, M.; Pant, S. Ecological analysis and environmental niche modelling of Dactylorhiza hatagirea (D. Don) Soo: A conservation approach for critically endangered medicinal orchid. Saudi J. Biol. Sci. 2021, 28, 2109–2122. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Description |
|---|---|
| BIO2 | Mean Diurnal Range (Mean of monthly (max temp–min temp)) |
| BIO3 | Isothermality (BIO2/BIO7) (×100) |
| BIO4 | Temperature Seasonality (standard deviation ×100) |
| BIO5 | Max Temperature of Warmest Month |
| BIO8 | Mean Temperature of Wettest Quarter |
| BIO9 | Mean Temperature of Driest Quarter |
| BIO12 | Annual Precipitation |
| BIO14 | Precipitation of Driest Month |
| BIO15 | Precipitation Seasonality (Coefficient of Variation) |
| BIO19 | Precipitation of Coldest Quarter |
| Taxon | Present | spp126 | spp245 | spp370 | spp585 |
|---|---|---|---|---|---|
| D. incarnata | 0.939 (0.005) | 0.940 (0.004) | 0.940 (0.004) | 0.940 (0.004) | 0.938 (0.005) |
| D. salina | 0.956 (0.010) | 0.951 (0.011) | 0.954 (0.010) | 0.955 (0.010) | 0.953 (0.010) |
| D. umbrosa | 0.956 (0.016) | 0.953 (0.015) | 0.954 (0.015) | 0.956 (0.015) | 0.954 (0.014) |
| D. majalis subsp. baltica | 0.984 (0.004) | 0.983 (0.005) | 0.984 (0.004) | 0.984 (0.004) | 0.983 (0.005) |
| Taxon | Present | spp126 | spp245 | spp370 | spp585 |
|---|---|---|---|---|---|
| D. incarnata | Bio14 (53.1) | Bio14 (54.1) | Bio14 (53.2) | Bio14 (52.5) | Bio14 (52.2) |
| Bio2 (11.5) | Bio2 (11.0) | Bio2 (11.3) | Bio2 (11.3) | Bio2 (12.2) | |
| Bio3 (6.4) | Bio3 (7.3) | Bio3 (6.5) | Bio3 (7.1) | Bio3 (7.7) | |
| D. salina | Bio2 (31.4) | Bio2 (28.0) | Bio2 (30.2) | Bio2 (29.9) | Bio2 (30.5) |
| Bio3 (17.3) | Bio14 (18.0) | Bio3 (16.9) | Bio14 (17.0) | Bio14 (17.5) | |
| Bio14 (16.4) | Bio3 (17.1) | Bio14 (16.7) | Bio3 (16.4) | Bio3 (16.7) | |
| D. umbrosa | Bio2 (24.4) | Bio2 (24.0) | Bio2 (23.7) | Bio2 (22.9) | Bio2 (24.1) |
| Bio14 (19.0) | Bio14 (18.7) | Bio14 (19.1) | Bio14 (19.5) | Bio14 (19.1) | |
| Bio3 (18.8) | Bio3 (18.5) | Bio3 (18.1) | Bio3 (18.7) | Bio3 (18.5) | |
| D. majalis subsp. baltica | Bio14 (52.7) | Bio14 (52.1) | Bio14 (53.1) | Bio14 (51.5) | Bio14 (51.4) |
| Bio3 (19.6) | Bio3 (21.4) | Bio3 (18.5) | Bio3 (19.2) | Bio3 (18.9) | |
| Bio5 (7.2) | Bio5 (6.3) | Bio5 (6.4) | Bio5 (7.7) | Bio5 (7.1) |
| Taxon | Change | spp126 | % | spp245 | % | spp370 | % | spp585 | % |
|---|---|---|---|---|---|---|---|---|---|
| D. incarnata | range expansion | 22.23 | 0.64 | 9202.87 | 265.38 | 200.06 | 5.77 | 66.69 | 1.92 |
| no occupancy | 2,725,095.23 | 99.87 | 2,715,914.59 | 99.54 | 2,724,917.40 | 99.87 | 2,725,050.78 | 99.87 | |
| no change | 133.37 | 3.85 | 111.15 | 3.21 | 0.00 | 0.00 | 0.00 | 0.00 | |
| range contraction | 3334.37 | 96.15 | 3356.60 | 96.79 | 3467.75 | 100.00 | 3467.75 | 100.00 | |
| D. salina | range expansion | 11,114.58 | 2.08 | 3956.79 | 0.74 | 4223.54 | 0.79 | 4134.62 | 0.77 |
| no occupancy | 2,183,259.40 | 80.01 | 2,190,417.19 | 80.28 | 2,190,150.44 | 80.27 | 2,190,239.36 | 80.27 | |
| no change | 260,525.78 | 48.77 | 140,932.89 | 26.38 | 106,655.52 | 19.97 | 96,763.54 | 18.11 | |
| range contraction | 273,685.45 | 51.23 | 393,278.34 | 73.62 | 427,555.71 | 80.03 | 437,447.68 | 81.89 | |
| D. umbrosa | range expansion | 23,718.52 | 4.58 | 7491.23 | 1.45 | 6979.96 | 1.35 | 5490.60 | 1.06 |
| no occupancy | 2,187,171.74 | 80.16 | 2,203,399.02 | 80.75 | 2,203,910.29 | 80.77 | 2,205,399.65 | 80.83 | |
| no change | 346,174.74 | 66.87 | 22,2047.10 | 42.89 | 175,276.94 | 33.86 | 163,740.01 | 31.63 | |
| range contraction | 171,520.22 | 33.13 | 295,647.86 | 57.11 | 342,418.02 | 66.14 | 353,954.95 | 68.37 | |
| D. majalis subsp. baltica | range expansion | 4245.77 | 10.08 | 3356.60 | 7.97 | 4868.19 | 11.56 | 3156.54 | 7.50 |
| no occupancy | 2,682,237.41 | 98.30 | 2,683,126.58 | 98.33 | 2,681,614.99 | 98.28 | 2,683,326.64 | 98.34 | |
| no change | 33,410.43 | 79.36 | 29,164.66 | 69.27 | 34,588.58 | 82.15 | 30,231.66 | 71.81 | |
| range contraction | 8691.60 | 20.64 | 12,937.37 | 30.73 | 7513.46 | 17.85 | 11,870.37 | 28.19 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sumbembayev, A.A.; Nowak, S.; Burzacka-Hinz, A.; Kosiróg-Ceynowa, A.; Szlachetko, D.L. New and Noteworthy Taxa of the Genus Dactylorhiza Necker ex Nevski (Orchidaceae Juss.) in Kazakhstan Flora and Its Response to Global Warming. Diversity 2023, 15, 369. https://doi.org/10.3390/d15030369
Sumbembayev AA, Nowak S, Burzacka-Hinz A, Kosiróg-Ceynowa A, Szlachetko DL. New and Noteworthy Taxa of the Genus Dactylorhiza Necker ex Nevski (Orchidaceae Juss.) in Kazakhstan Flora and Its Response to Global Warming. Diversity. 2023; 15(3):369. https://doi.org/10.3390/d15030369
Chicago/Turabian StyleSumbembayev, Aidar A., Sławomir Nowak, Aleksandra Burzacka-Hinz, Adrianna Kosiróg-Ceynowa, and Dariusz L. Szlachetko. 2023. "New and Noteworthy Taxa of the Genus Dactylorhiza Necker ex Nevski (Orchidaceae Juss.) in Kazakhstan Flora and Its Response to Global Warming" Diversity 15, no. 3: 369. https://doi.org/10.3390/d15030369
APA StyleSumbembayev, A. A., Nowak, S., Burzacka-Hinz, A., Kosiróg-Ceynowa, A., & Szlachetko, D. L. (2023). New and Noteworthy Taxa of the Genus Dactylorhiza Necker ex Nevski (Orchidaceae Juss.) in Kazakhstan Flora and Its Response to Global Warming. Diversity, 15(3), 369. https://doi.org/10.3390/d15030369