

Diversity of the Summer Phytoplankton of 43 Waterbodies in Bulgaria and Its Potential for Water Quality Assessment

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
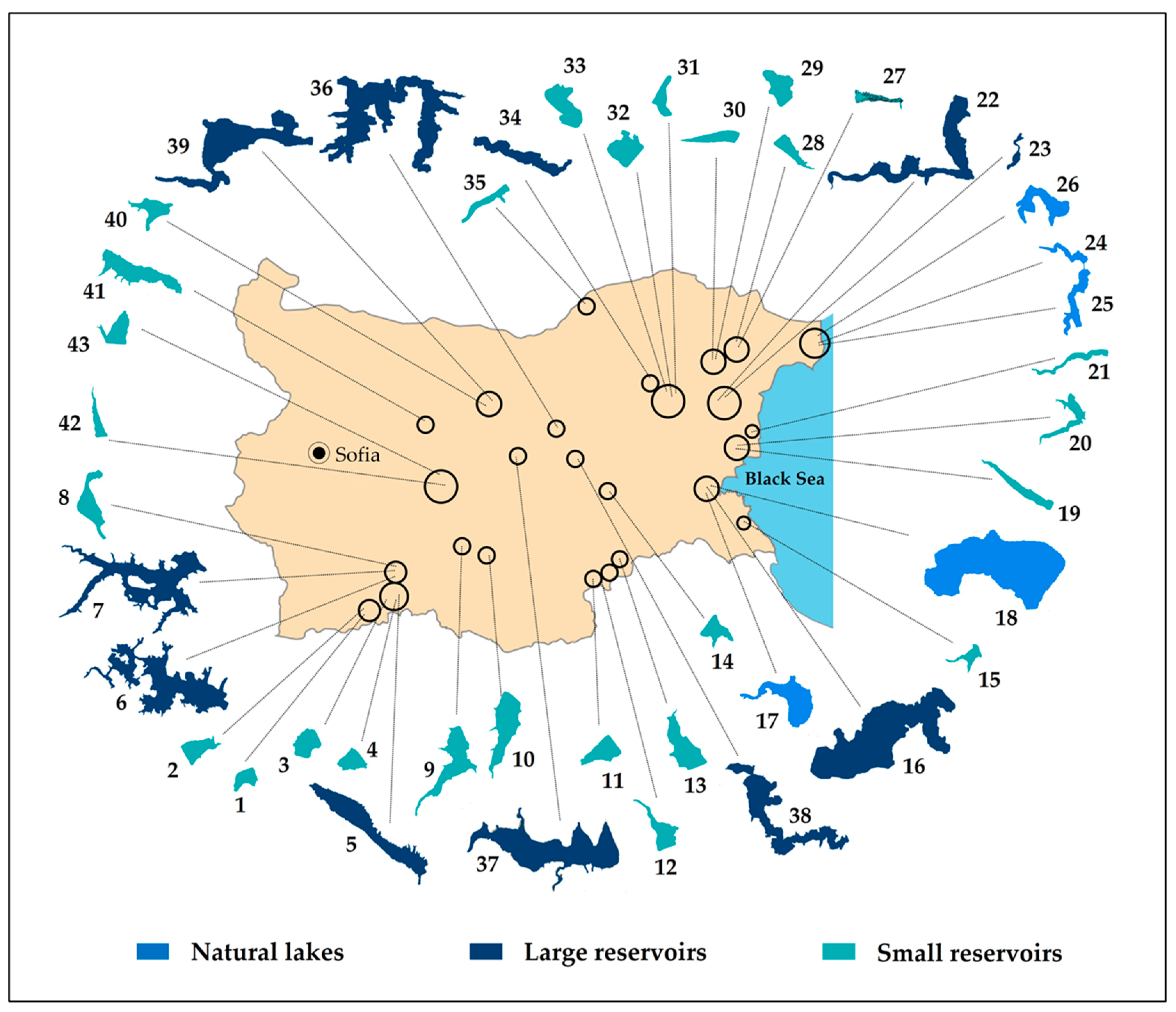
2.1. Sampling Sites
2.2. Algal Identification and Counting by Light Microscopy
2.3. Analysis of Phytoplankton Marker Pigments
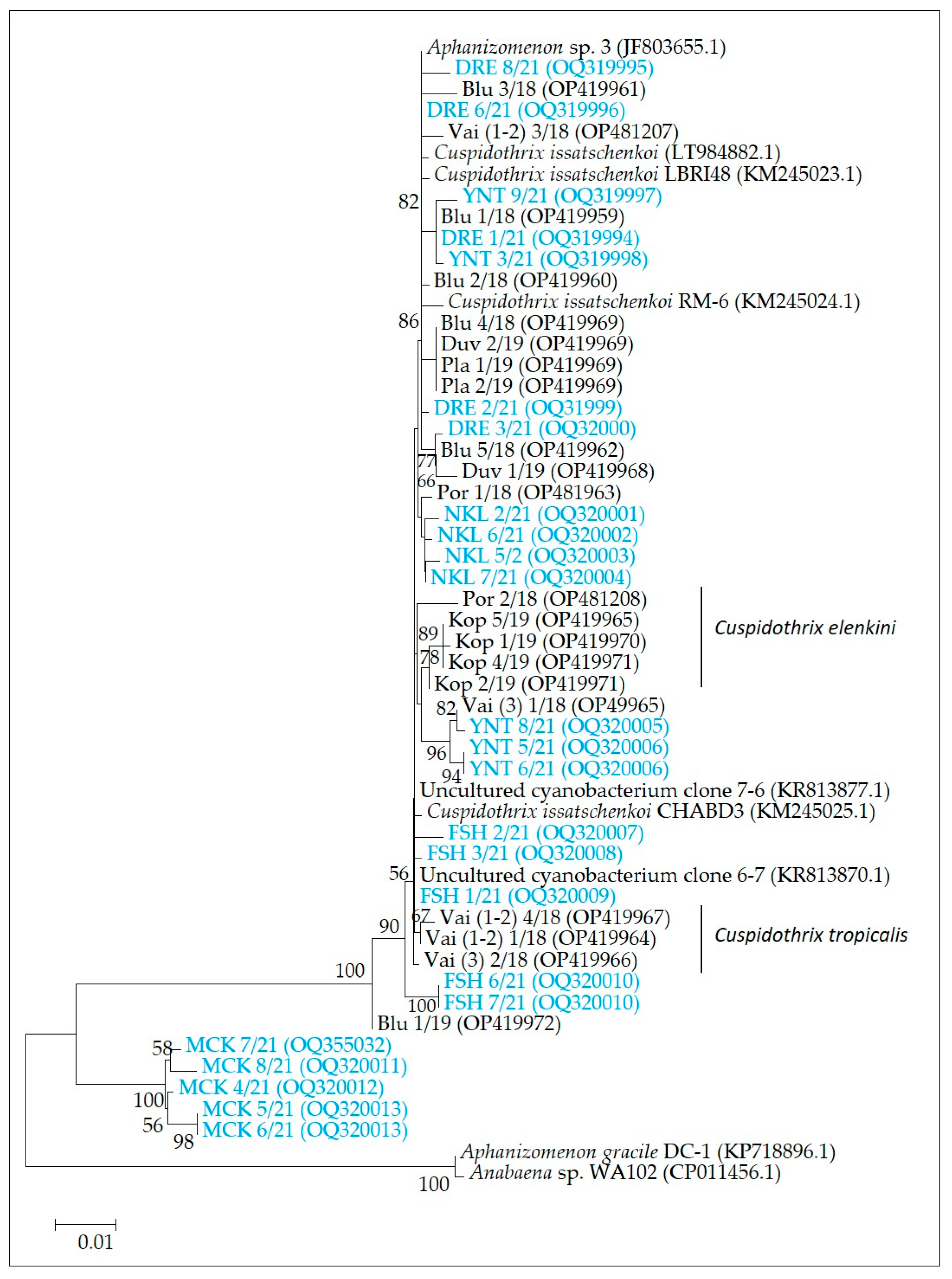
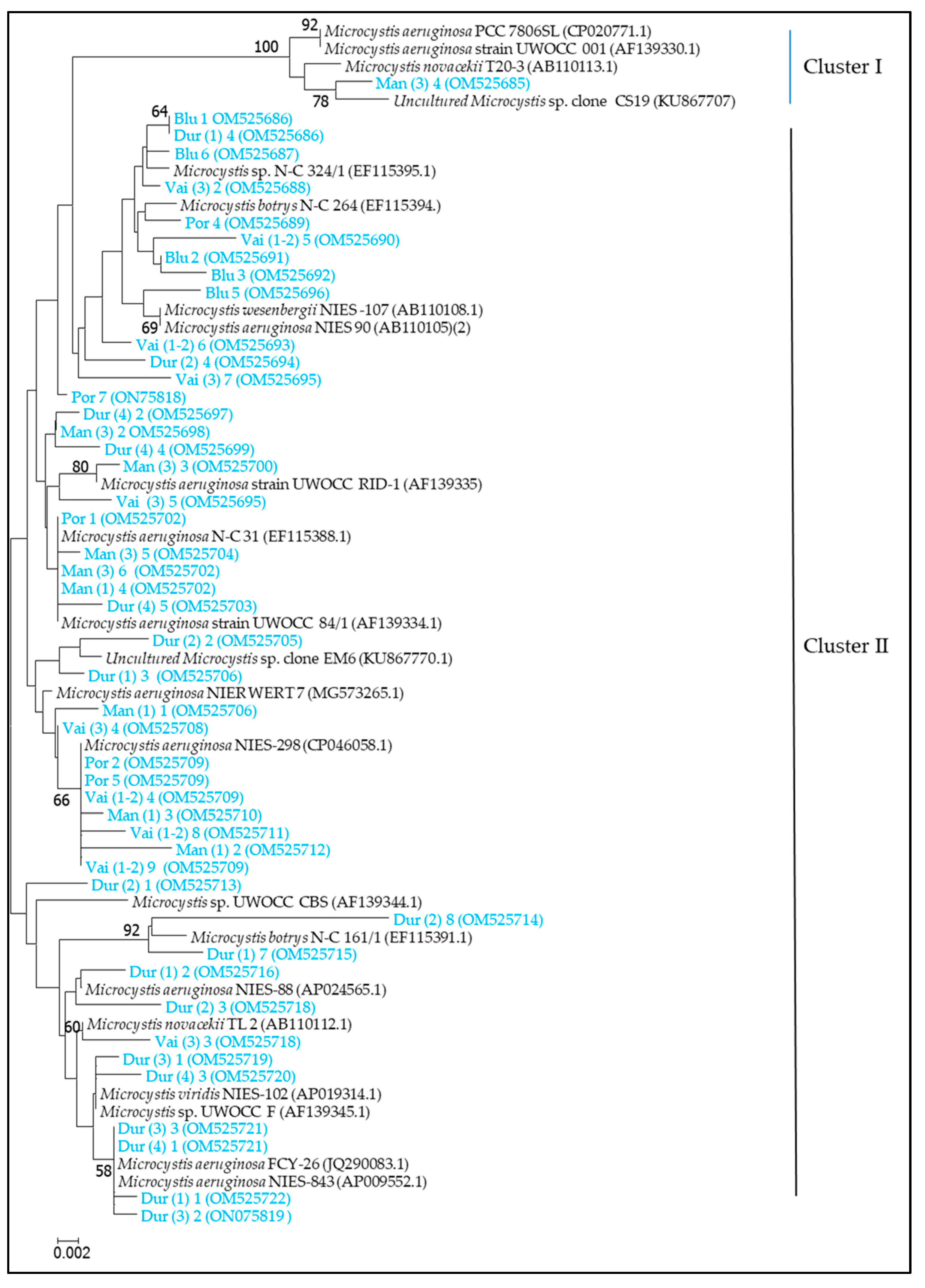
2.4. Molecular-Genetic Analysis
- Molecular-genetic analysis for the identification of anatoxin producers
- Molecular-genetic analysis for the identification of microcystin producers
2.5. Statistical Analysis
3. Results
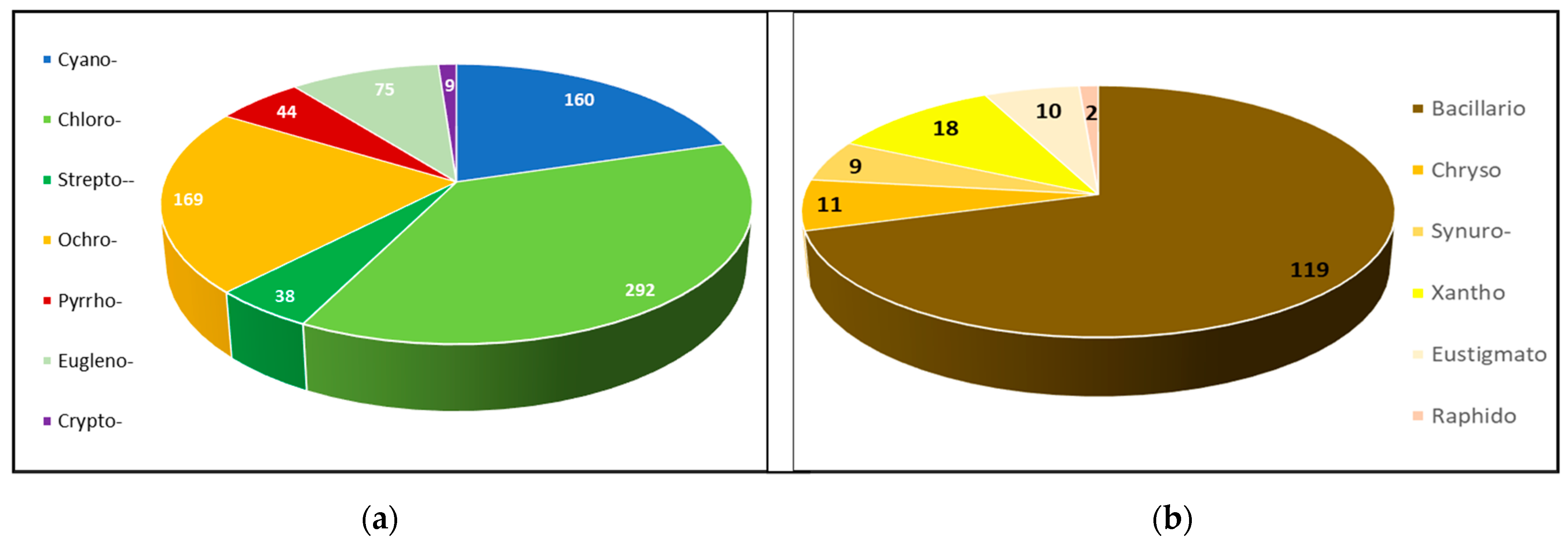
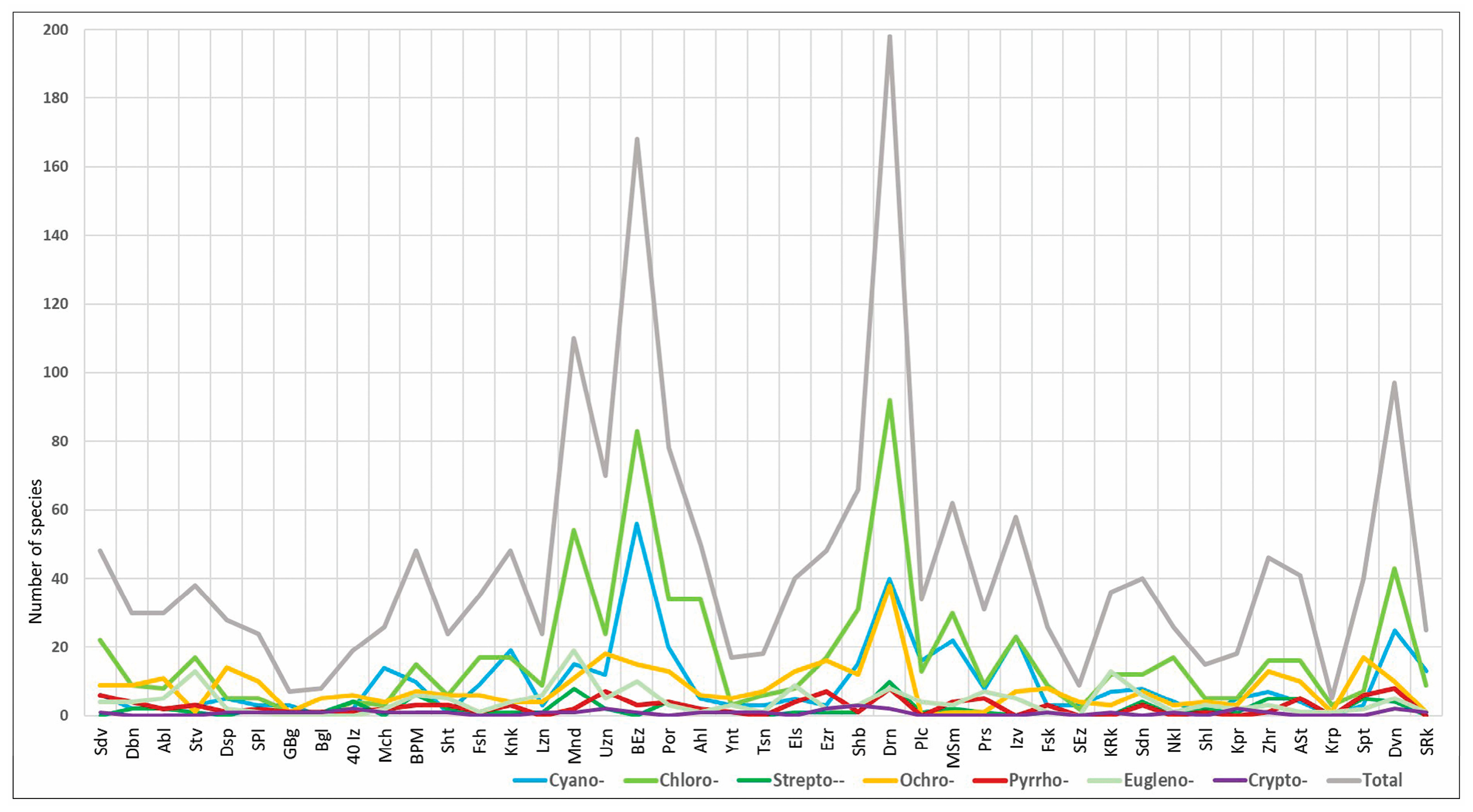
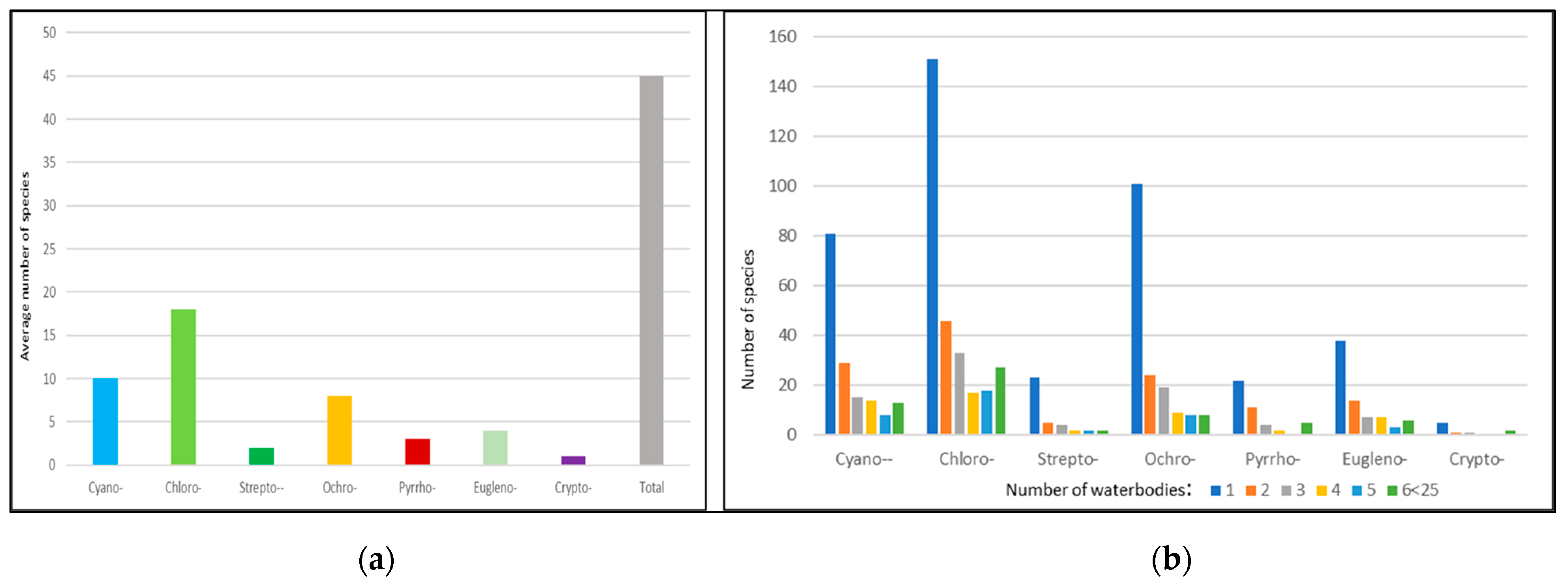
3.1. Total Biodiversity of the Phytoplankton
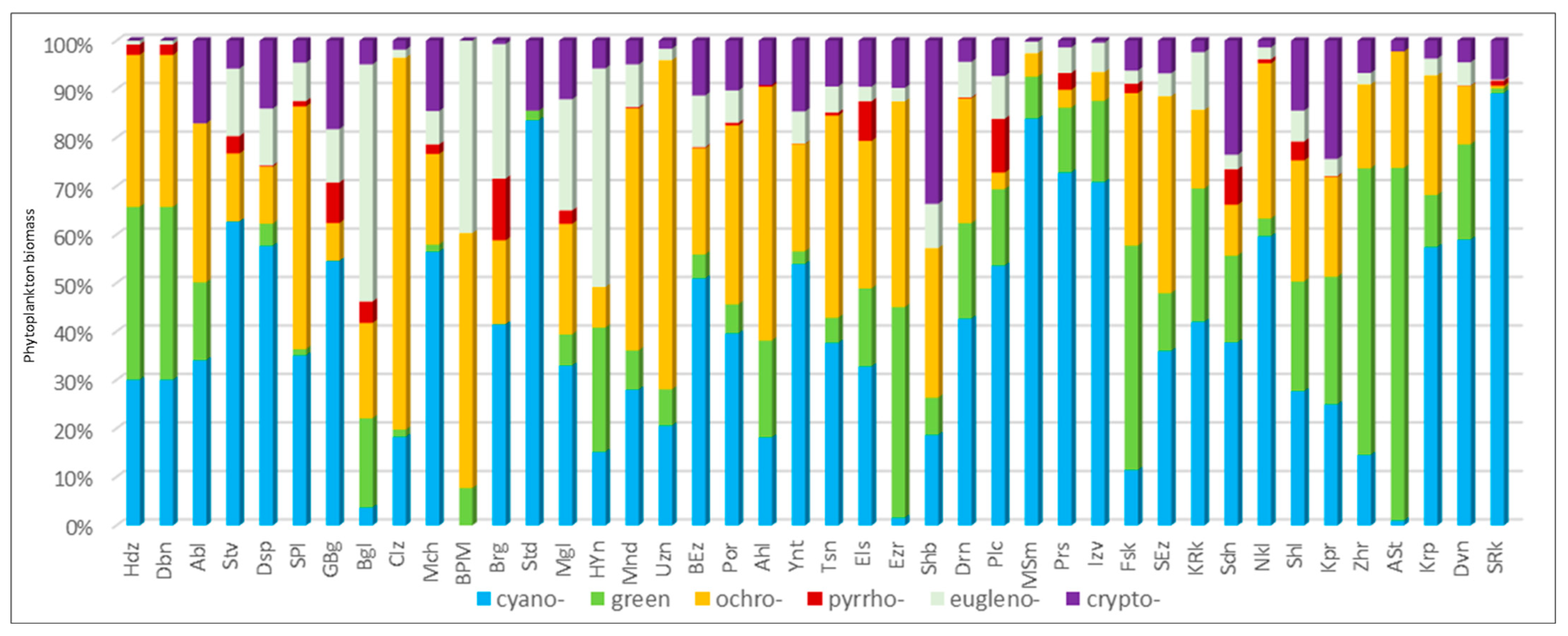
3.2. Phytoplankton Structure according to the Marker Pigment Composition
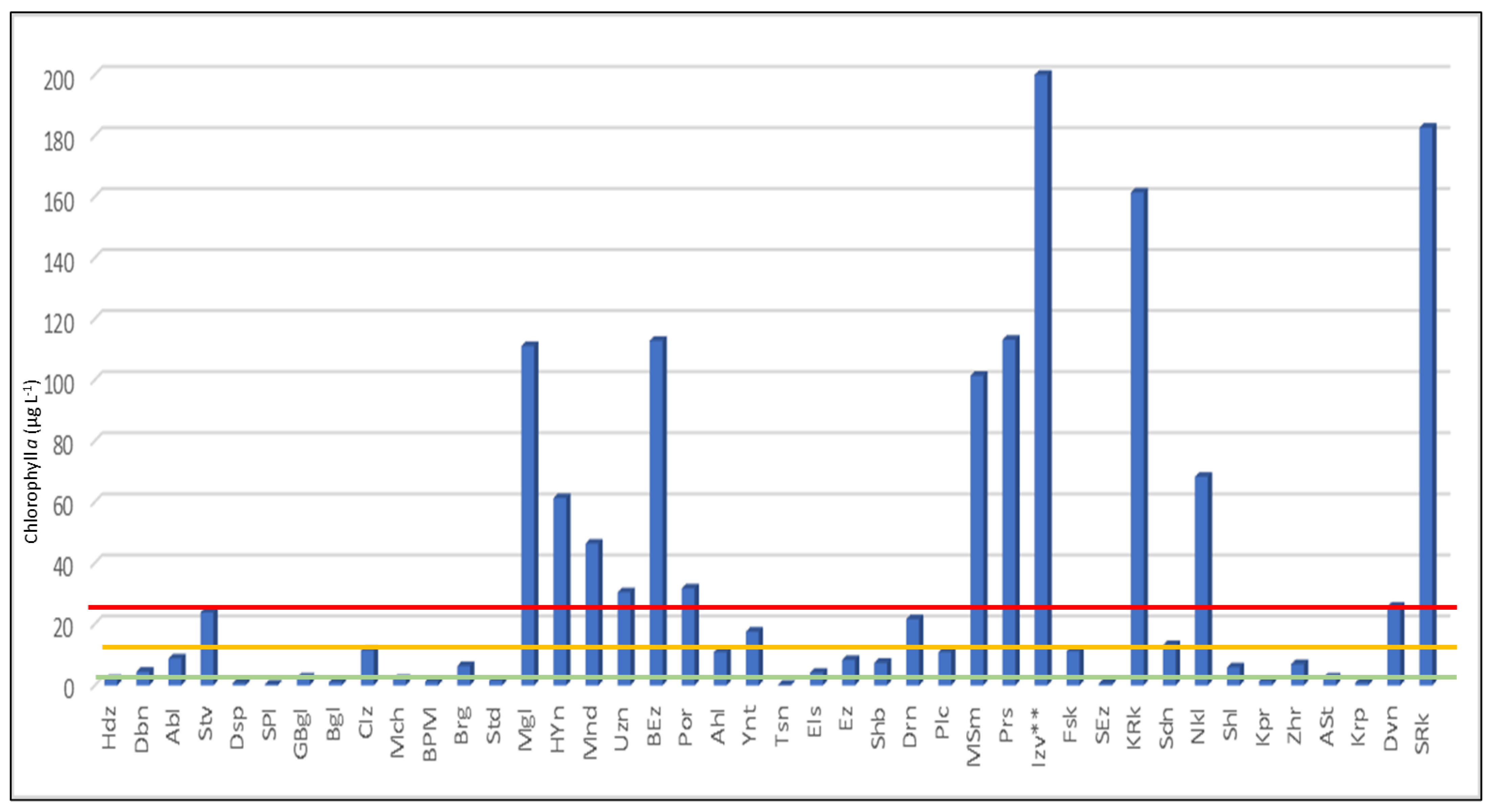
3.3. Algal Blooms and Toxic Species
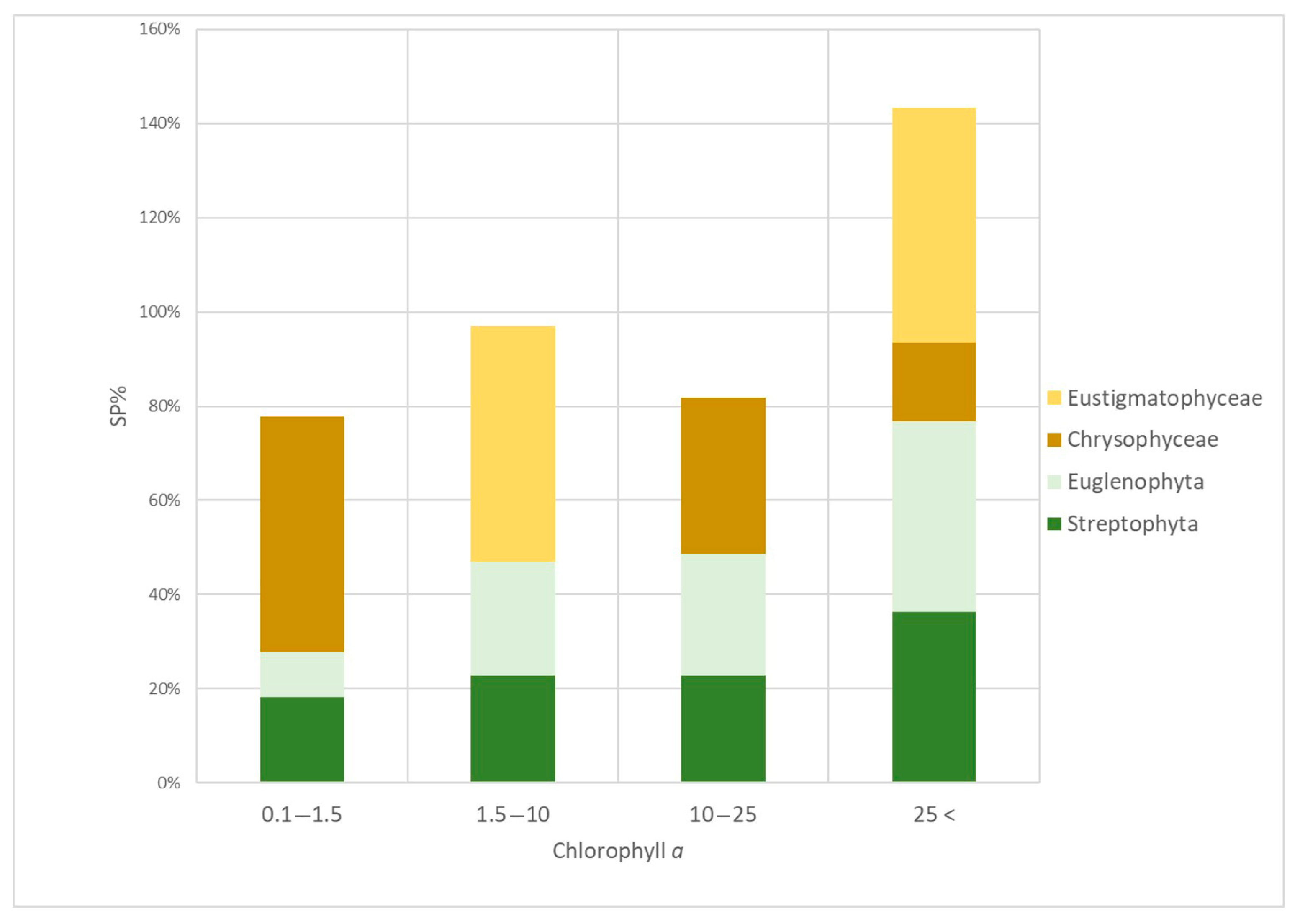
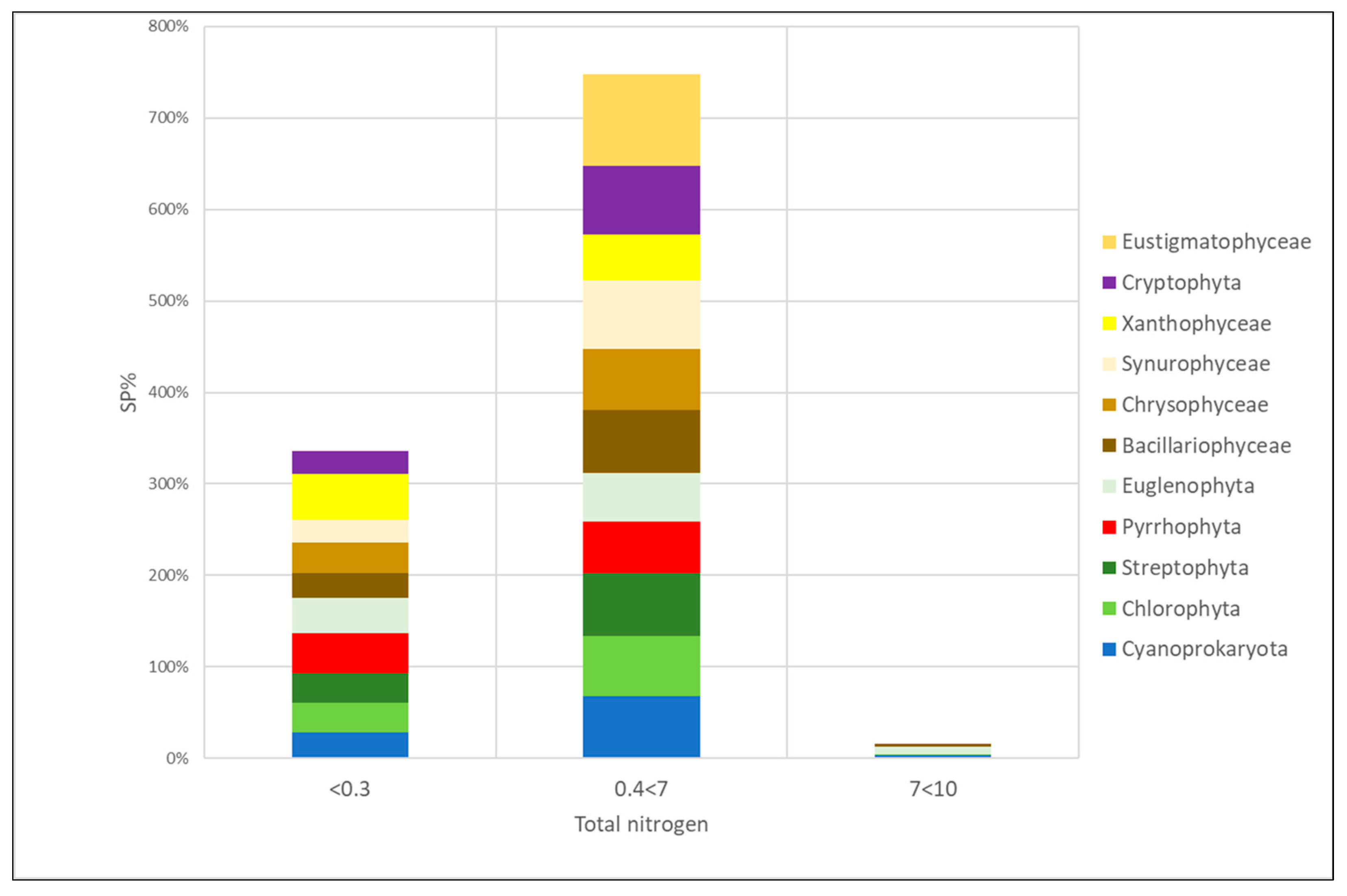
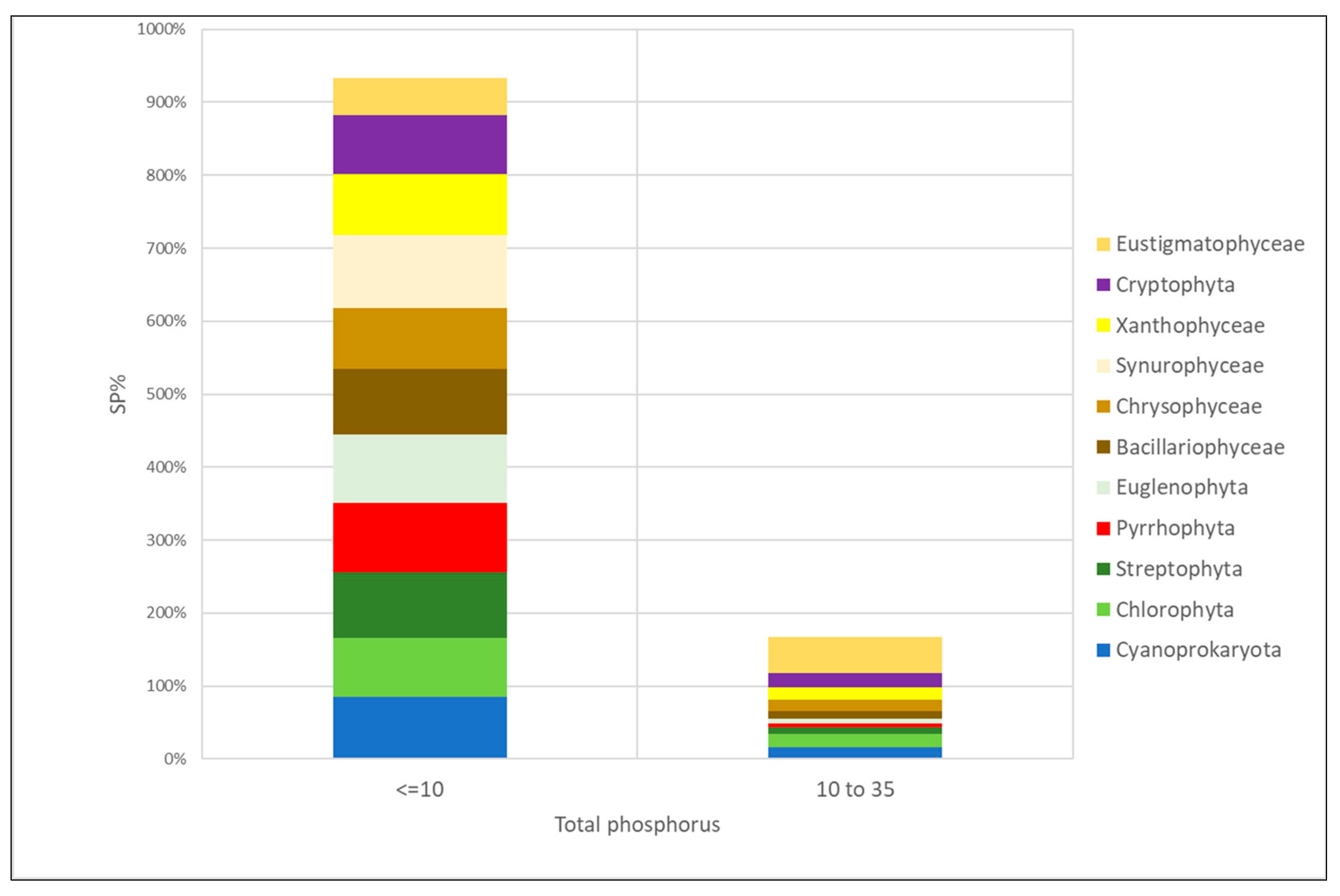
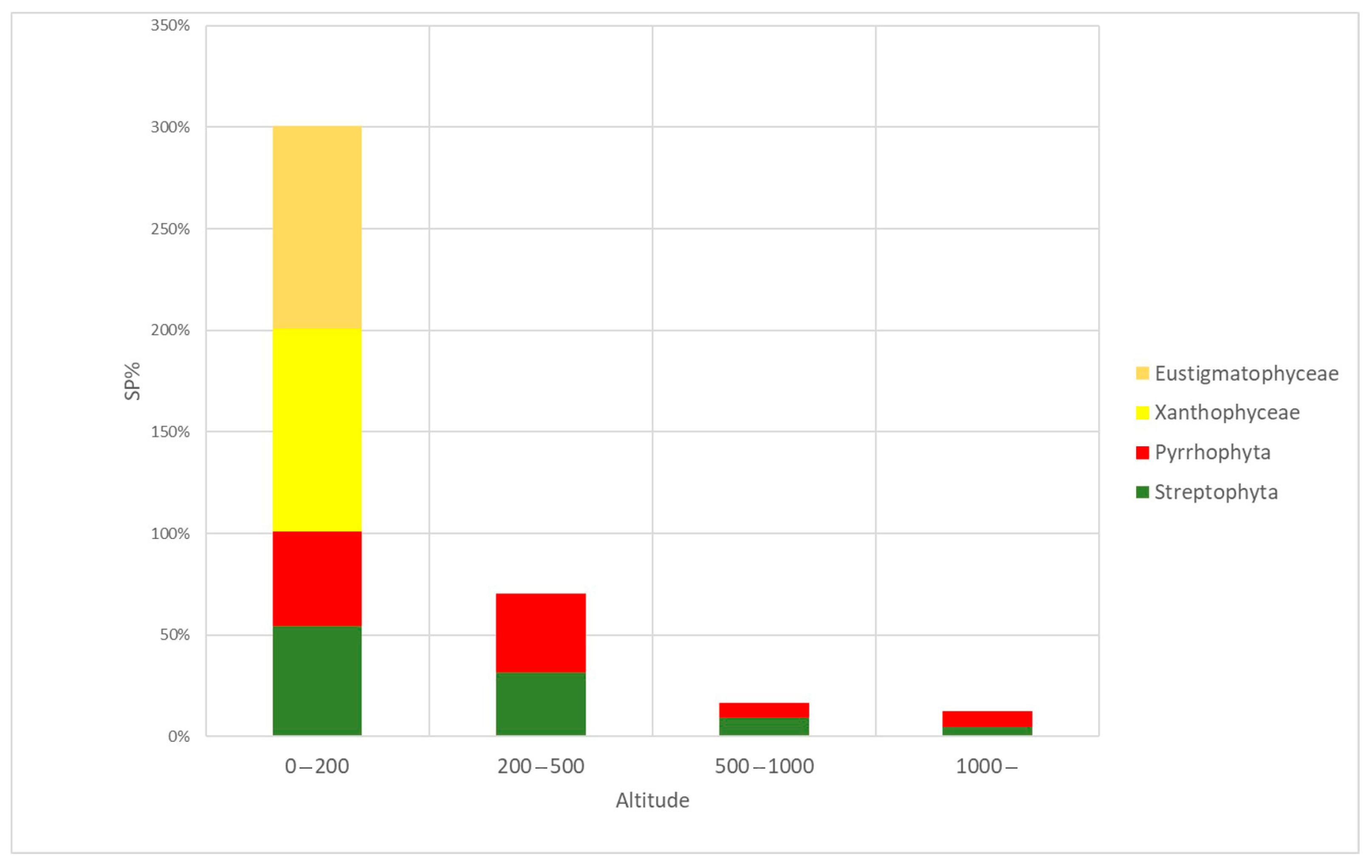
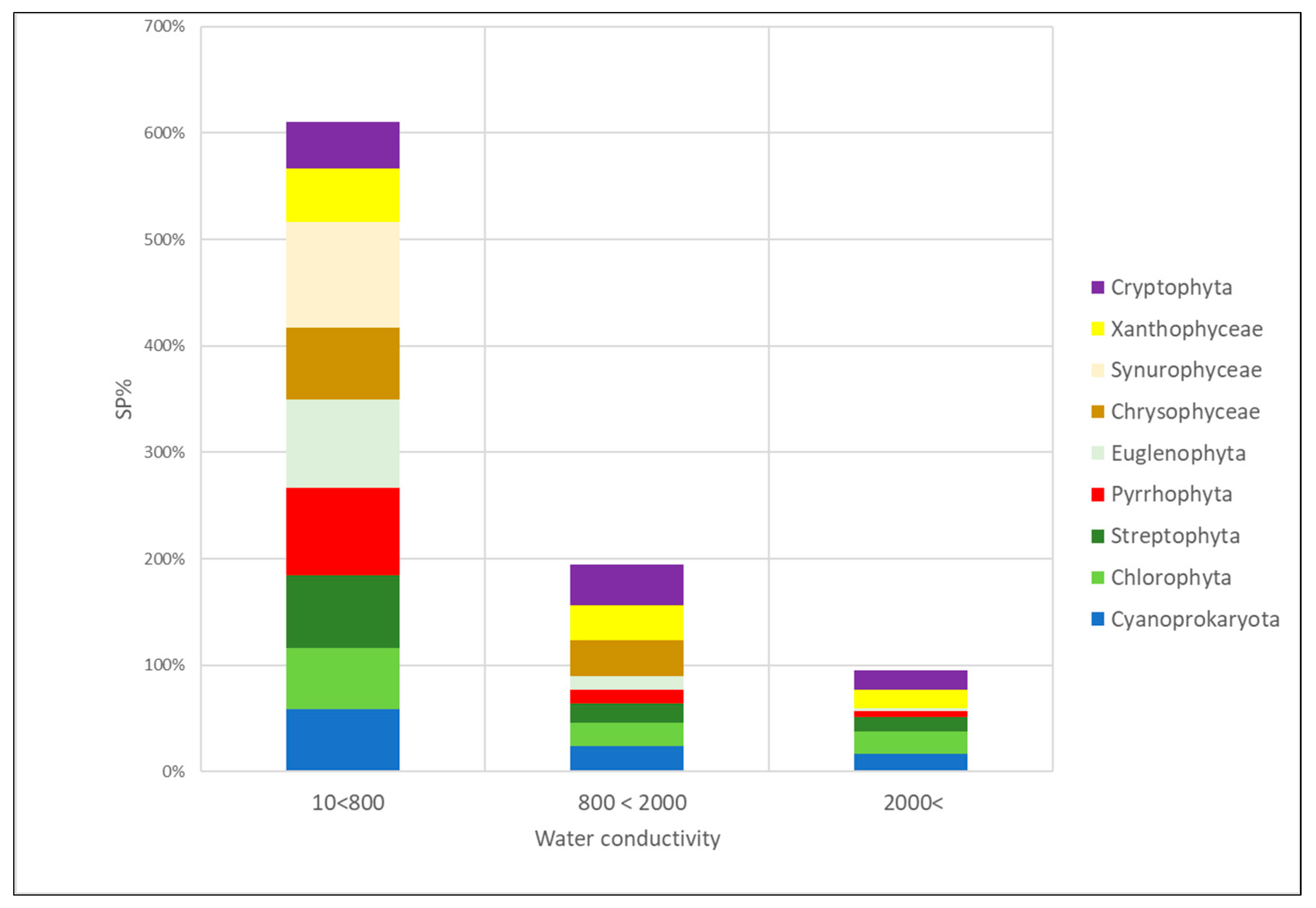
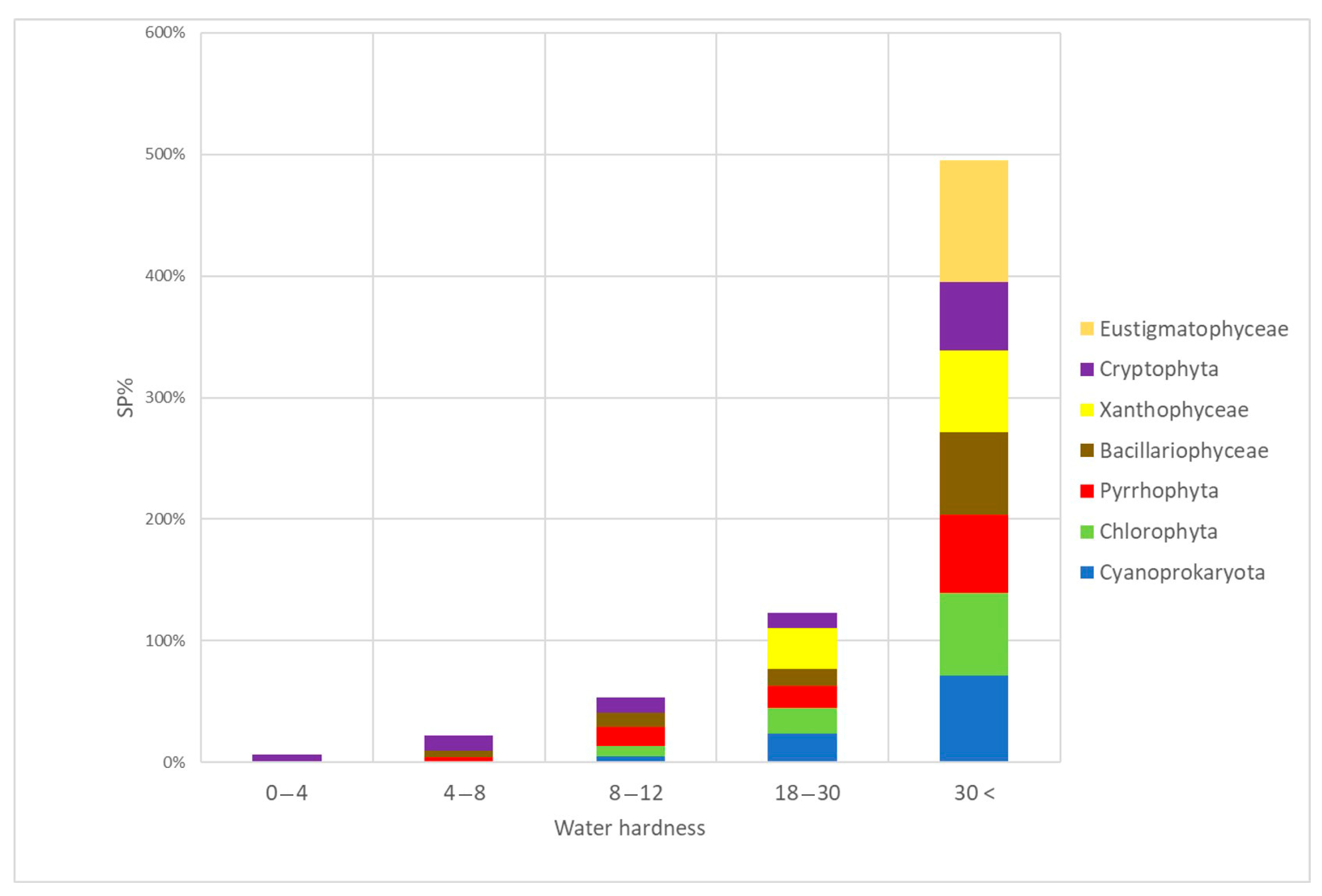
3.4. Algal Groups and Environmental Variables—Results from Statistical Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Delpla, I.; Jung, A.-V.; Baures, E.; Clement, M.; Thomas, O. Impacts of climate change on surface water quality in relation to drinking water production. Environ. Int. 2009, 35, 1225–1233. [Google Scholar] [CrossRef]
- Whitehead, P.G.; Wilby, R.L.; Battabee, R.W.; Kernan, M.; Wade, A.J. A review of the potential impacts of climate change on surface water quality. Hydrol. Sci. J. 2009, 54, 101–123. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, T.; Zounemat-Kermani, M.; Scholz, M. Climate Change, Water Quality and Water-Related Challenges: A Review with Focus on Pakistan. Int. J. Environ. Res. Public Health 2020, 17, 8518. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Guidelines for Drinking-Water Quality: Fourth Edition Incorporating the First and Second Addenda. 2022. Available online: https://www.who.int/publications/i/item/9789240045064 (accessed on 10 January 2023).
- Zeppernick, B.N.; Wilhelm, S.W.; Bullerjahn, G.S.; Paerl, H. Climate change and the aquatic continuum: A cyanobacterial comeback story. Environ. Microbiol. Rep. 2023, 15, 3–12. [Google Scholar] [CrossRef]
- Willén, E. Phytoplankton in Water Quality Assessment—An Indicator Concept. In Hydrological and Limnological Aspects in Lake Monitoring; Heinonen, P., Ziglio, G., Van der Beken, A., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2000. [Google Scholar]
- Zhang, Y.; Gao, W.; Li, Y.; Jiang, Y.; Chen, X.; Yao, Y.; Messyasz, B.; Yin, K.; He, W.; Chen, Y. Characteristics of the Phytoplankton Community Structure and Water Quality Evaluation in Autumn in the Huaihe River (China). Int. J. Environ. Res. Public Health 2021, 18, 12092. [Google Scholar] [CrossRef]
- Cellamare, M.; Morin, S.; Coste, M.; Haury, J. Ecological assessment of French Atlantic lakes based on phytoplankton, phytobenthos and macrophytes. Environ. Monit. Assess 2012, 184, 4685–4708. [Google Scholar] [CrossRef]
- Ptachnik, R.; Solimini, A.; Bretum, P. Performance of a new phytoplankton composition metric along a eutrophication gradient in Nordic lakes. Hydrobiologia 2009, 633, 75–82. [Google Scholar] [CrossRef]
- Stoyneva, M.; Traykov, I.; Tosheva, A.; Uzunov, B.; Zidarova, R.; Descy, J.-P. Comparison of ecological state/potential assessment of 19 Bulgarian water bodies based on macrophytes and phytoplankton (2011–2012). Biotechnol. Biotechnol. Equip. 2015, 29 (Suppl. 1), S33–S38. [Google Scholar] [CrossRef] [Green Version]
- Bellinger, E.G.; Sigee, D.C. Freshwater Algae. Identification, Enumeration and Use as Bioindicators, 2nd ed.; Wiley Blackwell: Singapore, 2015. [Google Scholar]
- Huszar, V.L.M.; Silva, L.H.S.; Domingos, P.; Marinho, M.M.; Melo, S. Phytoplankton species composition is more sensitive than OECD criteria to the trophic status of three Brazilian lakes. Hydrobiologia 1998, 129, 59–71. [Google Scholar] [CrossRef]
- Huszar, V.L.M.; Carraco, N. The relationship between phytoplankton composition and physical-chemical variables: A comparison of taxonomic and morphological-functional groups approaches in six temperate lakes. Freshw. Biol. 1998, 40, 1–18. [Google Scholar]
- Reynolds, C.S.; Huszar, V.; Kruk, C.; Naselli-Flores, L.; Melo, S. Towards a functional classification of the freshwater phytoplankton. J. Plankton Res. 2002, 24, 417–428. [Google Scholar] [CrossRef]
- Salmaso, N.; Naselli-Flores, L.; Padisák, J. Functional classifications and their application in phytoplankton ecology. Freshw. Biol. 2015, 60, 603–619. [Google Scholar] [CrossRef] [Green Version]
- Padisák, J.; Grigorszky, I.; Borics, G.; Soróczki-Pintér, É. Use of phytoplankton assemblages for monitoring ecological status of lakes within the Water Framework Directive: The assemblage index. Hydrobiologia 2006, 553, 1–14. [Google Scholar] [CrossRef]
- Kruk, C.; Peeters, E.; Van Nes, E.H.; Huszar, V.L.M.; Costa, L.S.; Scheffer, M. Phytoplankton community composition can be predicted best in terms of morphological groups. Limnol. Oceanogr. 2011, 56, 110–118. [Google Scholar] [CrossRef]
- Mischke, U.; Riedmüller, U.; Hoehn, E.; Schönfelder, I.; Nixdorf, B. Description of the German System for Phytoplankton-Based Assessment of Lakes for Implementation of the EU Water Framework Directive (WFD); Gewässereport 10, Aktuelle Reihe 2/2008; Univ. Cottbus, Lehrstuhl Gewässerschutz: Freiburg, Germany, 2008; pp. 117–146. [Google Scholar]
- Phillips, G.; Skjelbred, B.; Morabito, G.; Carvalho, L.; Lyche Solheim, A.; Andersen, T.; Mischke, U.; de Hoyos, C.; Borics, G. WISER Deliverable D3.1-1: Report on Phytoplankton Composition Metrics, Including a Common Metric Approach for Use in Intercalibration by All GIGs. 2010. Available online: http://www.wiser.eu/results/deliverables/ (accessed on 6 March 2023).
- Phillips, G.; Lyche Solheim, A.; Skjelbred, B.; Mischke, U.; Drakare, S.; Free, G.; Järvinen, M.; de Hoyos, C.; Morabito, G.; Poikane, S.; et al. A phytoplankton trophic index to assess the status of lakes for the Water Framework Directive. Hydrobiologia 2013, 704, 75–95. [Google Scholar] [CrossRef] [Green Version]
- Järvinen, M.; Drakare, S.; Free, G.; Lyche Solheim, A.; Phillips, G.; Skjelbred, B.; Mischke, U.; Ott, I.; Poikane, S.; Søndergaard, M.; et al. Phytoplankton indicator taxa for reference conditions in Northern and Central European lowland lakes. Hydrobiologia 2013, 704, 97–113. [Google Scholar] [CrossRef]
- Laplace-Treyture, C.; Feret, T. Performance of the Phytoplankton Index for Lakes (IPLAC): A multimetric phytoplankton index to assess the ecological status of water bodies in France. Ecol. Indic. 2016, 69, 686–698. [Google Scholar] [CrossRef] [Green Version]
- Gebler, D.; Kolada, A.; Pasztaleniec, A.; Szoszkiewicz, K. Modelling of ecological status of Polish lakes using deep learning techniques. Environ. Sci. Pollut. Res. 2020, 28, 5383–5397. [Google Scholar] [CrossRef]
- Evangelista, V.; Barsanti, L.; Frassanito, A.M.; Passarelli, V.; Gualtieri, P. (Eds.) Algal Toxins: Nature, Occurrence, Effect and Detection; NATO Advanced Study Institute on Sensor Systems for Biological Threats: The Algal Toxins Case; Springer: Dordrecht, The Netherlands, 2007. [Google Scholar]
- Svirčev, Z.; Lalić, D.; Savić, G.B.; Tokodi, N.; Backovic, D.D.; Chen, L.; Meriluoto, J.; Codd, G.A. Global geographical and historical overview of cyanotoxin distribution and cyanobacterial poisonings. Arch. Toxicol. 2019, 93, 2429–2481. [Google Scholar] [CrossRef]
- Lyche-Solheim, A.; Feld, C.K.; Birk, S.; Phillips, G.; Carvalho, L.; Morabito, G.; Mischke, U.; Willby, N.; Søndergaard, M.; Hellsten, S.; et al. Ecological status assessment of European lakes: A comparison of metrics for phytoplankton, macrophytes, benthic invertebrates and fish. Hydrobiologia 2013, 704, 57–74. [Google Scholar] [CrossRef] [Green Version]
- Mischke, U.; Thackeray, S.; Dunbar, M.; McDonald, C.; Carvalho, L.; de Hoyos, C.; Jarvinen, M.; Laplace-Treyture, C.; Morabito, G.; Skjelbred, B.; et al. WISER Deliverable D3.1-4: Guidance Document on Sampling, Analysis and Counting Standards for Phytoplankton in Lakes. 2012. Available online: https://nora.nerc.ac.uk/id/eprint/17466/ (accessed on 28 February 2023).
- CBD: The Convention on Biological Diversity. Available online: https://www.cbd.int/convention/text (accessed on 20 January 2023).
- Ohtani, S. How Is People’s Awareness of “Biodiversity” Measured? Using Sentiment Analysis and LDA Topic Modeling in the Twitter Discourse Space from 2010 to 2020. SN Comput. Sci. 2022, 3, 371. [Google Scholar] [CrossRef] [PubMed]
- Madgwick, G.; Jones, I.D.; Thackeray, S.J.; Elliott, J.A.; Miller, H.J. Phytoplankton communities and antecedent conditions: High resolution sampling in Esthwaite Water. Freshw. Biol. 2006, 51, 1798–1810. [Google Scholar] [CrossRef]
- Thackeray, S.; Nõges, P.; Dunbar, M.; Dudley, J.B.; Skjelbred, B.; Morabito, G.; Carvalho, L.; Phillips, G.; Mischke, U. WISER Deliverable D3.1-3: Uncertainty in Lake Phytoplankton Metrics. 2010. Available online: http://www.wiser.eu/download/D3.1-3.pdf (accessed on 27 February 2023).
- Thackeray, S.; Nõges, P.; Dunbar, M.; Dudley, J.B.; Skjelbred, B.; Morabito, G.; Carvalho, L.; Phillips, G.; Mischke, U.; Catalan, J.; et al. Quantifying uncertainties in biologicallybased water quality assessment: A pan-European analysis of lake phytoplankton community metrics. Ecol. Indic. 2013, 29, 34–47. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, L.; Poikane, S.; Lyche Solheim, A.; Phillips, G.; Borics, G.; Catalan, J.; De Hoyos, C.; Drakare, S.; Dudley, B.; Järvinen, M.; et al. Strength and uncertainty of phytoplankton metrics for assessing eutrophication impacts in lakes. Hydrobiologia 2013, 704, 127–140. [Google Scholar] [CrossRef] [Green Version]
- Maileht, K. Phytoplankton as Ecological Indicator of Lakes. Ph.D. Thesis, Estonian University of Life Sciences, Tartu, Estonia, 2021. [Google Scholar]
- Khalighi, M.; Sommeria-Klein, G.; Gonze, D.; Faust, K.; Lahti, L. Quantifying the impact of ecological memory on the dynamics of interacting communities. PLoS Comput. Biol. 2022, 18, e1009396. [Google Scholar] [CrossRef]
- Padisák, J. Seasonal Succession of Phytoplankton in a Large Shallow Lake (Balaton, Hungary)—A Dynamic Approach to Ecological Memory, Its Possible Role and Mechanisms. J. Ecol. 1992, 80, 217–230. [Google Scholar] [CrossRef]
- Michev, T.; Stoyneva, M. (Eds.) Inventory of Bulgarian Wetlands and Their Biodiversity; Elsi-M: Sofia, Bulgaria, 2007. [Google Scholar]
- Stoyneva-Gärtner, M.P.; Uzunov, B.A.; Descy, J.-P.; Gärtner, G.; Draganova, P.H.; Borisova, C.I.; Pavlova, V.; Mitreva, M. Pilot application of drone observations and pigment marker detection by HPLC in the studies of CyanoHABs in Bulgarian inland waters. Mar. Freshw. Res. 2019, 71, 606–616. [Google Scholar] [CrossRef]
- Radkova, M.; Stefanova, K.; Uzunov, B.; Gärtner, G.; Stoyneva-Gärtner, M. Morphological and Molecular Identification of Microcystin-Producing Cyanobacteria in Nine Shallow Bulgarian Water Bodies. Toxins 2020, 12, 39. [Google Scholar] [CrossRef] [Green Version]
- Stoyneva-Gärtner, M.; Stefanova, K.; Descy, J.-P.; Uzunov, B.; Radkova, M.; Pavlova, V.; Mitreva, M.; Gärtner, G. Microcystis aeruginosa and M. wesenbergii were the primary planktonic microcystin producers in several Bulgarian waterbodies (August 2019). Appl. Sci. 2021, 11, 357. [Google Scholar] [CrossRef]
- Uzunov, B.; Stefanova, K.; Radkova, M.; Descy, J.-P.; Gärtner, G.; Stoyneva-Gärtner, M. First Report on Microcystis as a Potential Microviridin Producer in Bulgarian Waterbodies. Toxins 2021, 13, 448. [Google Scholar] [CrossRef]
- Uzunov, B.; Stefanova, K.; Radkova, M.; Descy, J.-P.; Gärtner, G.; Stoyneva-Gärtner, M. Microcystis species and their toxigenic strains in phytoplankton of ten Bulgarian wetlands (August 2019). Botanica 2021, 27, 77–94. [Google Scholar] [CrossRef]
- Stoyneva-Gärtner, M.; Stefanova, K.; Uzunov, B.; Radkova, M.; Gärtner, G. Cuspidothrix Is the First Genetically Proved Anatoxin A Producer in Bulgarian Lakes and Reservoirs. Toxins 2022, 14, 778. [Google Scholar] [CrossRef] [PubMed]
- Stefanova, K.; Radkova, M.; Uzunov, B.; Gärtner, G.; Stoyneva-Gärtner, M. Pilot search for cylindrospermopsin-producers in nine shallow Bulgarian waterbodies reveals nontoxic strains of Raphidiopsis raciborskii, R. mediterranea and Chrysosporum bergii. Biotechnol. Biotechnol. Equip. 2020, 34, 384–394. [Google Scholar] [CrossRef]
- Valskys, V.; Gulbinas, Z.; Stoyneva-Gärtner, M.; Uzunov, B.; Skorupskas, R.; Karosienė, J.; Kasperovičienė, J.; Rašomavičius, V.; Uogintas, D.; Audzijonytė, A.; et al. Remote sensing in environmental studies: Advantages and challenges. Ann. Sof. Univ. 2022, 106, 31–44. [Google Scholar]
- OECD. Eutrophication of Waters—Monitoring, Assessment and Control; Organization for Economic Cooperation and Development: Paris, France, 1982. [Google Scholar]
- Poikāne, S.; Alves, M.H.; Argillier, C.; van den Berg, M.; Buzzi, F.; Hoehn, E.; de Hoyos, C.; Karottki, I.; Laplace-Treyture, C.; Solheim, A.L.; et al. Defining chlorophyll-a reference conditions in European lakes. Environ. Manag. 2010, 45, 1286–1298. [Google Scholar] [CrossRef] [Green Version]
- Descy, J.-P.; Stoyneva-Gärtner, M.P.; Uzunov, B.A.; Dimitrova, P.H.; Pavlova, V.T.; Gärtner, G. Studies on cyanoprokaryotes of the water bodies along the Bulgarian Black Sea Coast (1890–2017): A review, with special reference to new, rare and harmful taxa. Acta Zool. Bulgar. 2018, 11, 43–52. Available online: https://acta-zoologica-bulgarica.eu/supplement-11-2018/ (accessed on 10 December 2022).
- Stoyneva-Gärtner, M.P.; Descy, J.-P.; Latli, A.; Uzunov, B.; Pavlova, V.; Bratanova, Z.l.; Babica, P.; Maršálek, B.; Meriluoto, J.; Spoof, L. Assessment of cyanoprokaryote blooms and of cyanotoxins in Bulgaria in a 15-years period (2000–2015). Adv. Oceanogr. Limnol. 2017, 8, 131–152. [Google Scholar] [CrossRef] [Green Version]
- Google Earth. Available online: https:/?earth.google.com (accessed on 25 January 2023).
- Ginkgo Maps—Free Digital Maps. Available online: https://www.ginogomaps.com (accessed on 25 January 2023).
- Komárek, J. Cyanoprokaryota. In 3rd Part: Heterocytous Genera; Büdel, B., Krienitz, L., Gärtner, G., Schagerl, M., Eds.; Süßwasserflora von Mitteleuropa; Elsevier, Spektrum Akad. Verl.: Heidelberg, Germany, 2014; Volume 19. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota. 1. Teil: Chroococcales. In Süßwasserflora von Mitteleuropa. Bd. 19/1; Ettl, H., Gärtner, G., Heynig, G., Mollenhauer, D., Eds.; Gustav Fischer: Jena, Germany; Stuttgart, Germany; Lübeck Germany, 1999. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota. 2. Teil: Oscillatoriales. In Süßwasserflora von Mitteleuropa. Bd. 19/2; Büdel, B., Gärtner, G., Krienitz, L., Schagerl, M., Eds.; Elsevier, Spektrum Akad. Verl.: Heidelberg, Germany; München, Germany, 2005. [Google Scholar]
- Komárek, J.; Fott, B. Chlorophyceae (Grünalgen). Ordnung: Chlorococcales. In Das Phytoplankton des Süßwassers, 7/1; Schweizerbart’sche Verlagsbuchhandlung: Stuttgart, Germany, 1983; pp. 1–1044. [Google Scholar]
- Moestrup, Ø.; Calado, A.J. Süßwasserflora von Mitteleuropa, Bd. 6—Freshwater Flora of Central Europe, Vol. 6: Dinophyceae; Springer Spektrum: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. Available online: http://www.algaebase.org/ (accessed on 26 December 2022).
- Stoyneva-Gärtner, M.; Uzunov, B. Bases of Systematics of Algae and Fungi; JAMG Books: Sofia, Bulgaria, 2017. [Google Scholar]
- Starmach, K. Metody Badania Planktonu; PWRiL: Warszawa, Poland, 1955; pp. 1–135. [Google Scholar]
- Stoyneva, M.P. Planktic green algae of Bulgarian coastal wetlands. Hydrobiologia 2000, 438, 25–41. [Google Scholar] [CrossRef]
- Descy, J.P. SOP5: Estimation of cyanobacteria biomass by marker pigment analysis. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, J., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 343–349. [Google Scholar]
- Sarmento, H.; Descy, J.-P. Use of marker pigments and functional groups for assessing the status of phytoplankton assemblages in lakes. J. Appl. Phycol. 2008, 20, 1001–1011. [Google Scholar] [CrossRef]
- Mackey, M.D.; Mackey, D.J.; Higgins, H.W.; Wright, S.W. CHEMTAX—A program for estimating class abundances from chemical markers: Application to HPLC measurements of phytoplankton. Mar. Ecol. Prog. Ser. 1996, 144, 265–283. [Google Scholar] [CrossRef] [Green Version]
- Wright, S.W.; Jeffrey, S.W. Pigment markers for phytoplankton production. In Marine Organic Matter: Biomarkers, Isotopes and DNA; Volkman, J.K., Ed.; The Handbook of Environmental Chemistry; Springer: Berlin/Heidelberg, Germany, 2006; Volume 2N, pp. 71–104. [Google Scholar]
- Christensen, V.G.; Khan, E. Freshwater neurotoxins and concerns for human, animal, and ecosystem health: A review of anatoxin-a and saxitoxin. Sci. Total Environ. 2020, 736, 139515. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Cyanobacterial Toxins: Anatoxin-a and Analogues. Background Document for Development of WHO Guidelines for Drinking-Water Quality and Guidelines for Safe Recreational Water Environments; World Health Organization: Geneva, Switzerland, 2020.
- Legrand, B.; Lesobre, J.; Colombet, J.; Latour, D.; Sabart, M. Molecular tools to detect anatoxin-a genes in aquatic ecosystems: Toward a new nested PCR-based method. Harmful Algae 2016, 58, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Ballot, A.; Scherer, P.I.; Wood, S.A. Variability in the anatoxin gene clusters of Cuspidothrix issatschenkoi from Germany, New Zealand, China and Japan. PLoS ONE 2018, 13, e0200774. [Google Scholar] [CrossRef]
- Rantala-Ylinen, A.; Känä, S.; Wang, H.; Rouhiainen, L.; Wahlsten, M.; Rizzi, E.; Berg, K.; Gugger, M.; Sivonen, K. Anatoxin-a synthetase gene cluster of the cyanobacterium Anabaena sp. strain 37 and molecular methods to detect potential producers. Appl. Environ. Microbiol. 2011, 77, 7271–7278. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Basic Local Alignment Search Tool (BLAST). Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 24 January 2023).
- NCBI: National Centre for Biotechnology Information. Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 12 January 2023).
- World Health Organization (WHO). Cyanobacterial Toxins: Microcystins. Background Document for Development of WHO Guidelines for Drinking-Water Quality and Guidelines for Safe Recreational Water Environments; WHO: Geneva, Switzerland, 2020.
- Humbert, J.-F. Molecular tools for the detection of toxigenic Cyanobacteria in natural ecosystems. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, J., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 280–283. [Google Scholar]
- Mbukwa, E.; Boussiba, S.; Wepener, V.; Leu, S.; Kaye, Y.; Msagati, T.A.M.; Mamba, B.B. PCR amplification and DNA sequence of mcyA gene: The distribution profile of a toxigenic Microcystis aeruginosa in the Hartbeespoort Dam, South Africa. J. Water Health 2013, 11, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Mbukwa, E.; Msagati, T.A.M.; Mamba, B.B.; Boussiba, S.; Wepener, V.; Leu, S.; Kaye, Y. Toxic Microcystis novacekii T20-3 from Phakalane Ponds, Botswana: PCR Amplifications of Microcystin Synthetase (mcy) Genes, Extraction and LC-ESI-MS Identification of Microcystins. J. Environ. Anal. Toxicol. 2015, S7, 010. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Choi, J.; Fatka, M.; Swanner, E.; Ikuma, K.; Liang, X.; Leung, T.; Howe, A. Improved detection of mcyA genes and their phylogenetic origins in harmful algal blooms. Water Res. 2020, 176, 115730. [Google Scholar] [CrossRef] [PubMed]
- Statistical Analysis Software Programs in Biomedical Research. Available online: https://www.labome.com/method/Statistical-Analysis-Software-Programs-in-Biomedical-Research.html (accessed on 26 January 2023).
- Ward, M.H.; Jones, R.R.; Brender, J.D.; de Kok, T.M.; Weyer, P.J.; Nolan, B.T.; Villanueva, C.M.; van Breda, S.G. Drinking Water Nitrate and Human Health: An Updated Review. Int. J. Environ. Res. Public Health 2018, 15, 1557. [Google Scholar] [CrossRef] [Green Version]
- Haralampiev, K. SPSS for Advanced; University Press, St. Kliment Ohridski: Sofia, Bulgaria, 2007. [Google Scholar]
- Stoyneva, M. Algal flora of the Danube River (Bulgarian sector) and adjoined water basins. V. Algal flora of the water bodies adjacent to the Lake of Srebarna. Ann. Univ. Sof. 1995, 88, 5–19. [Google Scholar]
- Stoyneva, M.P. Development of the phytoplankton of the shallow Srebarna Lake (North-Eastern Bulgaria) across the trophic gradient. Hydrobiologia 1998, 369/370, 259–367. [Google Scholar] [CrossRef]
- Stoyneva, M. Algae. In Biodiversity of the Srebarna Biosphere Reserve. Checklist and Bibliography; Michev, T.M., Georgiev, B.B., Petrova, A.V., Stoyneva, M.P., Eds.; Co-Publ. Context & Pensoft: Sofia, Bulgaria, 1998; pp. 10–371. [Google Scholar]
- Stoyneva, M.P. Steady-state phytoplankton assemblages in shallow Bulgarian wetlands. Hydrobiologia 2003, 502, 169–176. [Google Scholar] [CrossRef]
- Stoyneva, M.P. Contribution to the Studies of the Biodiversity of Hydro- and Aerobiontic Prokaryotic and Eukaryotic Algae in Bulgaria. Ph.D. Thesis, Sofia University “St. Kliment Ohridski”, Sofia, Bulgaria, 2014. [Google Scholar]
- Stoyanov, P.; Teneva, I.; Mladenov, R.; Belkinova, D. Diversity and Ecology of the Phytoplankton of Filamentous Blue-Green Algae (Cyanoprokaryota, Nostocales) in Bulgarian Standing Waters. Ecol. Balk 2013, 5, 1–6. [Google Scholar]
- Dimitrova, R.E.; Nenova, E.P.; Uzunov, B.A.; Shihiniova, M.D.; Stoyneva, M.P. Phytoplankton composition of Vaya Lake (2004–2006). Bulg. J. Agric. Sci. 2014, 20 (Suppl. 1), 165–172. [Google Scholar]
- Pavlova, V.; Stoyneva, M.; Georgieva, V.; Donchev, D.; Spoof, L.; Meriluoto, J.; Bratanova, Z.; Karadjova, I. New records of microcystins in some Bulgarian water bodies of health and conservational importance. J. Water Resour. Prot. 2014, 6, 446–453. [Google Scholar] [CrossRef] [Green Version]
- Stoyneva, M.P. Allochthonous planktonic algae recorded during the last 25 years in Bulgaria and their possible dispersal agents. Hydrobiologia 2016, 764, 53–64. [Google Scholar] [CrossRef]
- Kokociński, M.; Akçaalan, R.; Salmaso, N.; Stoyneva-Gärtner, M.P.; Sukenik, A. Expansion of alien and invasive cyanobacteria. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, J., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 28–39. [Google Scholar]
- Dochin, K. The dominance of invasive algae Raphidiopsis raciborskii in lowland reservoirs in Bulgaria. BJAS 2022, 28, 158–165. [Google Scholar]
- Watanabe, M. Studies on the planktonic blue-green algae 3. Some Aphanizomenon species in Hokkaido, northern Japan. Bull. Natl. Sci. Mus. Nat. Sci. Ser. B Bot. 1991, 17, 141–150. [Google Scholar]
- Teiling, E. Schwedische Planktonalgen. II. Tetrallantos, eine neue Gattung der Protococcoideen. Svenka Bot. Tidskr. Upps. 1916, 10, 59–66. [Google Scholar]
- Tasinov, O.B.; Vankova, D.G.; Nazifova-Tasinova, N.F.; Pasheva, M.G.; Kiselova, Y.D.; Sokrateva, T.D.; Ivanov, D.L.; Uzunov, B.A.; Stoyneva-Gärtner, M.P.; Ivanova, D.G. Cytotoxicity of water from five Bulgarian wetlands contaminated by toxigenic cyanobacteria and cyanotoxins. Bulg. Chem. Commun. 2020, 52, 257–262. [Google Scholar]
- Chichova, M.; Tasinov, O.; Shkodrova, M.; Mishonova, M.; Sazdova, I.; Ilieva, B.; Doncheva-Stoimenova, D.; Kiselova-Kaneva, Y.; Raikova, N.; Uzunov, B.; et al. New Data on Cylindrospermopsin Toxicity. Toxins 2021, 13, 41. [Google Scholar] [CrossRef]
- Teneva, I.; Belkinova, D.; Mladenov, R.; Stoyanov, P.; Moten, D.; Basheva, D.; Kazakov, S.; Dzhambazov, B. Phytoplankton composition with an emphasis of Cyanobacteria and their toxins as an indicator for the ecological status of Lake Vaya (Bulgaria)—Part of the Via Pontica migration route. BDJ 2020, 8, e57507. [Google Scholar] [CrossRef] [PubMed]
- Maileht, K.; Nõges, T.; Nõges, P.; Ott, I.; Mischke, U.; Carvalho, L.; Dudley, B. Water colour, phosphorus and alkalinity are the major determinants of the dominant phytoplankton species in European lakes. Hydrobiologia 2013, 704, 115–126. [Google Scholar] [CrossRef]
- Nõges, P.; van de Bund, W.; Cardoso, A.C.; Solimini, A.G.; Heiskanen, A.-S. Assessment of the ecological status of European surface waters: A work in progress. Hydrobiologia 2009, 633, 197–211. [Google Scholar] [CrossRef]
- Reynolds, C.S. The Ecology of Phytoplankton; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Dochin, K.T.; Stoyneva, M.P. Effect of long-term cage fish-farming on the phytoplankton biodiversity in two large Bulgarian reservoirs. Ber. Naturwiss.-Med. Ver. Innsbr. 2014, 99, 49–66. [Google Scholar]
- Dochin, K.T.; Stoyneva, M.P. Phytoplankton of the Dospat Reservoir (Rhodopi Mts, Bulgaria)—Indicator of negative trend in reservoir development due to long-term cage fish farming. Ann. Sof. Univ. 2016, 99, 47–60. [Google Scholar]
- Teneva, I.; Mladenov, R.; Belkinova, D.; Dimitrova-Dyulgerova, I.; Dzhambazov, B. Phytoplankton community of the drinking water supply reservoir Borovitsa (South Bulgaria) with an emphasis on cyanotoxins and water quality. Open Life Sci. 2010, 5, 231–239. [Google Scholar] [CrossRef]
- Stoyneva, M.P. Algological studies of Bulgarian coastal wetlands. I. Species composition of the phytoplankton of Durankulak and Shabla-Ezeretz lakes. Ann. Univ. Sof. 2000, 91, 27–48. [Google Scholar]
- Stoyneva, M. Algological studies of Bulgarian coastal wetlands. II. Quantitative structure of the phytoplankton of Durankulak and Shabla-Ezeretz lakes. Ann. Univ. Sof. 2002, 92, 91–109. [Google Scholar]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WBN and IBW | Abbr | Type | Year | Alt | Latitude | Longitude | WT | pH | CN | TDS | DO | TP | TN | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | * Hadzhidimovo | Hdz | M | 2021 | 156 | 41°29.8933 | 23°50.1890 | 29.1 | 9.5 | 300 | 192 | 17.00 | 0.1 | 0.1 |
| 2 | * Dubnitsa (IBW3698) | Dbn | M | 2021 | 600 | 41°33.8500 | 23°50.7500 | 25.2 | 9.6 | 246 | 159 | 9.21 | 0.1 | 0.1 |
| 3 | * Ablanitsa (IBW6013) | Abl | M | 2021 | 682 | 41°32.8594 | 23°55.5869 | 27.2 | 8.8 | 242 | 157 | 8.54 | 1.0 | 0.5 |
| 4 | * Satovcha 2 (IBW1197) | Stv | M | 2021 | 1017 | 41°36.8222 | 23°58.1446 | 27.4 | 8.70 | 272 | 176 | 9.00 | 0.5 | 0.1 |
| 5 | Dospat (IBW3155) | Dsp | R | 2021 | 1214 | 41°39.1495 | 24°89.5596 | 25.9 | 9.9 | 81 | 52 | 8.73 | 0.1 | 0.5 |
| 2021 | 1212 | 41°39.1493 | 24°89.5918 | 25.6 | 9.5 | 83 | 52 | 8.70 | 0.3 | 0.5 | ||||
| 6 | Golyam Beglik (IBW1314) | GBg | R | 2021 | 1540 | 41°48.8927 | 24°07.8725 | 22.0 | 9.1 | 99 | 63 | 8.92 | 1.5 | 1.0 |
| 7 | Shiroka Polyana (IBW3144) | SPl | R | 2021 | 1550 | 41°46.1776 | 24°08.8201 | 25.3 | 8.9 | 66 | 42 | 8.70 | 0.5 | 0.5 |
| 8 | Beglika (IBW3141) | Bgl | M | 2021 | 1535 | 41°49.7963 | 24°07.8196 | 21.7 | 9.1 | 242 | 157 | 9.11 | 1.0 | 0.8 |
| 9 | * Chetiridesette Izvora | 246 | 42°00.5510 | |||||||||||
| (IBW1523) | CIz | M | 2021 | 24°56.2819 | 28.7 | 7.5 | 402 | 263 | 8.66 | 1.0 | 0.5 | |||
| 10 | * Mechka (IBW1584) | Mck | M | 2021 | 319 | 41°55.8970 | 25°06.1595 | 27.1 | 9.0 | 241 | 154 | 8.50 | 1.5 | 1.0 |
| 11 | * Byalata Prust-Mezek | BPM | M | 2021 | 167 | 41°45.1080 | 26°05.2403 | 29.7 | 8.5 | 291 | 188 | 9.37 | 2.0 | 1.0 |
| 12 | * Birgo (Shtit) | Brg | M | 2021 | 215 | 41°49.7743 | 26°22.1889 | 27.3 | 8.0 | 594 | 385 | 8.75 | 1.5 | 1.8 |
| 13 | * Studena (Fishera) | 282 | 41°54.2136 | 1 | ||||||||||
| (IBW2421) | Std | M | 2021 | 26°24.5964 | 29.3 | 9.0 | 652 | 423 | 3.35 | 1.0 | 0.3 | |||
| 14 | * Mogila (Kaynaka) | 166 | 42°29.8310 | |||||||||||
| (IBW2626) | Mgl | M | 2021 | 26°36.1043 | 29.2 | 9.5 | 682 | 442 | 15.75 | 4.0 | 1.0 | |||
| 15 | * Hadzhi Yani (Lozenets) | 12 | 42°12.0333 | |||||||||||
| (IBW2893) | HYn | M | 2021 | 27°47.3000 | 26.1 | 7.5 | 751 | 488 | 8.42 | 1.5 | 0.8 | |||
| 16 | Mandra (IBW1720) | Mnd | R | 2018 | 12 | 42°24.0643′ | 27°26.1120′ | 25.9 | 8.3 | 649 | 421 | 6.81 | 3.0 | 3.0 |
| 2018 | 42°24.0670′ | 27°19.1310′ | 26.2 | 8.2 | 663 | 461 | 5.89 | 6.0 | 4.0 | |||||
| 2018 | 42°26.1420′ | 27°26.5860′ | 24.9 | 8.5 | 639 | 415 | 7.91 | 4.0 | 3.3 | |||||
| 2019 | 42°24.0295′ | 27°19.1194′ | 25.88 | 7.9 | 676 | 436 | 7.93 | 0.7 | 0.5 | |||||
| 2019 | 42°25.9303′ | 27°26.7652′ | 27.2 | 8.5 | 578 | 375 | 7.87 | 1.5 | 1.8 | |||||
| 2021 | 42°24.2370 | 27°19.1205′ | 27.3 | 9.0 | 513 | 333 | 9.32 | 7.0 | 4.0 | |||||
| 2021 | 42°25.9282 | 27°26.7675′ | 27.3 | 9.0 | 513 | 333 | 10.70 | 7.5 | 4.0 | |||||
| 17 | Uzungeren (IBW0710) | Uzn | L | 2018 | 7 | 42°26.1782′ | 27°27.1998′ | 25.9 | 8.1 | 1458 | 9351 | 7.83 | 5.0 | 2.8 |
| 2019 | 42°26.1551′ | 27°27.2235′ | 27.6 | 8.5 | 1748 | 1132 | 9.70 | 0.4 | 0.3 | |||||
| 2021 | 42°26.1532′ | 27°27.2214′ | 28.1 | 9.0 | 18520 | 12000 | 11.21 | 5.5 | 4.0 | |||||
| 18 | Burgasko Ezero (Vaya) (IBW0191) | BEz | L | 2018 | 0 | 42°30.5940′ | 27°22.0750′ | 26.9 | 9.7 | 2588 | 1682 | 12.51 | 13 | 5.4 |
| 2018 | 42°28.4540′ | 27°25.4820′ | 28.28 | 8.9 | 1183 | 768 | 11.94 | 11 | 3.7 | |||||
| 2018 | 42°29.1850′ | 27°26.5310′ | 23.7 | 9.5 | 1024 | 665 | 7.01 | 12 | 4.6 | |||||
| 2019 | 42°30.5940′ | 27°22.0750′ | 27.9 | 9.2 | 490 | 170 | 7.69 | 0.5 | 0.3 | |||||
| 2021 | 42°30.7934′ | 27°24.2425′ | 26.6 | 9.0 | 4421 | 2873 | 1.26 | 12 | 5.3. | |||||
| 19 | Poroy (IBW3038) | Por | M | 2018 | 41 | 42°43.0190′ | 27°37.3160′ | 25.10 | 8.3 | 762 | 495 | 9.45 | 1.0 | 2.8 |
| 2019 | 42°43.3403′ | 27°37.5255′ | 27.5 | 8.1 | 644 | 416 | 7.60 | 0.1 | 0.3 | |||||
| 2021 | 42°43.4683′ | 27°36.8757′ | 26.1 | 9.0 | 792 | 514 | 11.68 | 2.1 | 1.5 | |||||
| 20 | Aheloy (IBW3032) | Ahl | M | 2018 | 144 | 42°42.8230′ | 27°30.9740′ | 25.4 | 8.5 | 614 | 399 | 8.92 | 1 | 4.1 |
| 21 | * Yunets | Ynt | M | 2021 | 79 | 42°55.6700′ | 27°45.3074′ | 27.4 | 8.5 | 965 | 765 | 11.00 | 2.5 | 1.8 |
| 22 | Tsonevo (IBW3022) | Tsn | R | 2019 | 75 | 43°01.8055′ | 27°24.3965′ | 24.8 | 8.8 | 355 | 231 | 8.20 | 0.1 | 0.1 |
| 2021 | 43°01.8278′ | 27°24.3954′ | 26.6 | 8.0 | 417 | 272 | 10.65 | 0.1 | 0.1 | |||||
| 23 | Eleshnitsa (IBW3023) | Els | M | 2019 | 44 | 43°00.3344′ | 27°28. 0744′ | 26.7 | 8.4 | 532 | 347 | 6.78 | 0.1 | 0.3 |
| 24 | Ezerets (IBW0233) | Ezr | L | 2018 | 0 | 43°35.2770′ | 28°33.2290′ | 26.4 | 8.4 | 1084 | 10 | 9.94 | 0.5 | 5.3 |
| 2019 | 6 | 43°35.2681′ | 28°33.2096′ | 25.9 | 8.6 | 1669 | 1739 | 8.58 | 0.1 | 0.1 | ||||
| 25 | Shabla (IBW0219) | Shb | L | 2018 | 0 | 43°33.8180′ | 28°34.1860′ | 27.1 | 8.5 | 1087 | 706 | 9.97 | 0.1 | 5.1 |
| 2019 | 43°33.8212′ | 28°34.8204′ | 25.9 | 8.7 | 1842 | 1196 | 9.64 | 0.1 | 1.0 | |||||
| 26 | Durankulak (IBW0216) | Drn | L | 2018 | 4 | 43°40.3240′ | 28°32.0470′ | 24.03 | 8.5 | 1111 | 722 | 7.35 | 21 | 2.8 |
| 2018 | 43°40.3340′ | 28°32.0220′ | 24.7 | 8.2 | 1094 | 711 | 7.79 | 20 | 4.0 | |||||
| 2018 | 43°40.5300′ | 28°32.9930′ | 24.6 | 8.5 | 1075 | 698 | 6.19 | 24 | 3.9 | |||||
| 2018 | 43°40.6950′ | 28°32.6000′ | 26.5 | 8.5 | 1087 | 706 | 9.60 | 20 | 3.2 | |||||
| 2019 | 43°40.0006′ | 29°32.6166′ | 26.5 | 8.9 | 974 | 631 | 7.86 | 0.3 | 0.7 | |||||
| 2019 | 43°40.5355′ | 28°33.0806′ | 26.7 | 8.9 | 1048 | 680 | 6.04 | 0.3 | 0.6 | |||||
| 2021 | 43°40.6935′ | 28°32.6000′ | 25.5 | 9.0 | 2960 | 736 | 10.70 | 14 | 4.5 | |||||
| 2021 | 43°40.5300′ | 28°33.0826′ | 25.5 | 9.0 | 3008 | 1952 | 7.40 | 11 | 2.0 | |||||
| 27 | * Plachidol 2 (IBW5073) | Plc | M | 2019 | 220 | 43°33.3504′ | 27°52.6338′ | 24.6 | 9.0 | 1225 | 793 | 9.13 | 0.2 | 0.4 |
| 28 | * Malka Smolnitsa (IBW3107) | MSm | M | 2019 | 211 | 43°36.2606′ | 27°44.5367′ | 25.2 | 9.1 | 755 | 490 | 7.05 | 0.6 | 0.6 |
| 29 | * Preselka (IBW3078) | Prs | M | 2019 | 281 | 43°25.3767′ | 27°16.6214′ | 24.1 | 9.0 | 138 | 282 | 10.05 | 0.6 | 2.8 |
| 30 | * Izvornik 2 (IBW3082) | Izv | M | 2019 | 255 | 43°27.3838′ | 27°21.1110′ | 24.5 | 9.4 | 389 | 253 | 13.26 | 9.0 | 4.8 |
| 31 | * Fisek (IBW2674) | Fsk | M | 2019 | 182 | 43°18.8453′ | 26°44. 3765′ | 27.2 | 8.7 | 690 | 397 | 7.52 | 0.2 | 0.1 |
| 32 | * Shumensko Ezero (IBW2754) | SEz | M | 2019 | 152 | 43°14.8140′ | 26°57.5675′ | 25.2 | 8.5 | 471 | 445 | 6.32 | 0.2 | 0.5 |
| 33 | * Kriva Reka (IBW3071) | KRk | M | 2019 | 133 | 43°22.6573′ | 27°10.9807′ | 23.7 | 8.4 | 662 | 428 | 6.24 | 1.0 | 9.0 |
| 34 | Suedinenie (IBW2642) | Sdn | R | 2019 | 133 | 43°20.0734′ | 26°33.6368′ | 28.1 | 7.6 | 739 | 481 | 6.77 | 0.1 | 0.3 |
| 35 | * Nikolovo (IBW3176) | Nkl | M | 2021 | 89 | 43°50.9768 | 26°05.1796 | 26.0 | 9.8 | 2156 | 1400 | 11.88 | 11 | 2.0 |
| 36 | Shilkovtsi (Iovkovtsi) (IBW2105) | Shl | R | 2019 | 410 | 42°55.2320′ | 25°47.6743′ | 27.2 | 8.9 | 746 | 479 | 7.48 | 0.03 | 0.1 |
| 37 | Koprinka (IBW2062) | Kpr | R | 2019 | 450 | 42°37.0172′ | 25°19.4795′ | 27.2 | 8.2 | 250 | 163 | 7.21 | 0.1 | 0.2 |
| 38 | Zhrebchevo (IBW2545) | Zhr | R | 2019 | 253 | 42°36.6024′ | 25°51.2345′ | 27.6 | 7.7 | 358 | 233 | 8.01 | 0.1 | 0.2 |
| 39 | Al. Stamboliyski (IBW2056) | ASt | R | 2019 | 190 | 43°07.0000′ | 25°07.3936′ | 29.4 | 8.9 | 670 | 433 | 9.82 | 1.4 | 3.5 |
| 40 | Krapets (IBW2000) | Krp | M | 2019 | 410 | 43°04.0316′ | 24°52.3835′ | 28.7 | 8.3 | 870 | 564 | 7.74 | 0.1 | 1.0 |
| 41 | Sopot (IBW1437) | Spt | R | 2019 | 376 | 40°00.7017′ | 24°52.6045′ | 29.0 | 8.3 | 779 | 490 | 3.44 | 0.1 | 0.1 |
| 42 | * Duvanli (IBW1483) | Dvn | M | 2019 | 250 | 42°23.1851′ | 24°43.1000′ | 26.3 | 8.8 | 4050 | 291 | 7.09 | 0.1 | 0.3 |
| 43 | Sinyata Reka (IBW1890) | SRk | M | 2018 | 317 | 42°28.1480′ | 24°42.2170 | 27.4 | 9.7 | 470 | 305 | 9.36 | 25 | 4.8 |
| 2018 | 42°28.1473′ | 24°42.2175 | 26.7 | 9.4 | 468 | 306 | 9.21 | 27 | 4.3 | |||||
| 2019 | 42°28.1518′ | 24°42.0159′ | 28.2 | 10.4 | 490 | 317 | 14.76 | 1.0 | 0.2 |
| Species | Abundance | Waterbody |
|---|---|---|
| Cyanoprokaryota | ||
| Anabaenopsis elenkinii + Cuspidothrix issatschenkoi | Codominants | Mogila |
| Aphanizomenon klebahnii | Dominant/Subdominant | Mandra (2019), Poroy (2019, 2021)/Hadzhi Yani |
| Aphanizomenon yezoense + Sphaerospermopsis aphanizomenoides | Codominants | Studena |
| Chrysosporum minor + Raphidiopsis mediterranea | Codominants | Plachidol 2 |
| Chrysosporum ovalisporum | Dominant | Shabla (2019) |
| Dolichospermum compactum | Dominant | Izvornik 2 |
| Dolichospermum perturbatum + Planktothrix isothrix | Codominants | Burgasko Ezero (2018) |
| Dolichospermum planctonicum | Dominant/Codominant | Golyam Beglik/Ablanitsa |
| Dolichospermum scheremetieviae | Dominant | Yunets |
| Limnothrix redekei | Dominant | Preselka |
| Limnothrix mirabilis | Codominant | Poroy (2018) |
| Microcystis wesenbergii | Dominant | Kriva Reka, Nikolovo, Sinya Reka (2018) |
| Planktothrix isothrix + Planktothrix suspensa | Codominants | Burgasko Ezero (2019) |
| Pseudanabaena limnetica | Codominant/Subdominant | Duvanli, Malka Smolnitsa/Preselka |
| Raphidiopsis raciborskii | Codominant/Subdominant | Malka Smolnitsa/Byalata Prust, Poroy (2018), Preselka |
| Raphidiopsis raciborskii + R. acuminato-crispa + R. gangetica | Codominants | Mechka |
| Romeria simplex | Codominant | Duvanli |
| Sphaerospermopsis aphanizomenoides | Dominant | Burgasko Ezero (2021) |
| Sphaerospermopsis torques-reginae | Dominant | Sinyata Reka (2019) |
| Anabaenopsis milleri | Subdominant | Izvornik 2 |
| Aphanizomenon yezoense + Microcystis aeruginosa+Pseudanabaena mucicola + Synechocystis endobiotica | Subdominants | Durankulak (2021) |
| Aphanocapsa delicatissima | Subdominant | Shumensko Ezero |
| Aphanocapsa holsatica | Subdominant | Durankulak (2018), Hadzhi Yani |
| Coelomoron pusillum | Subdominant | Kriva Reka |
| Dolichospermum perturbatum | Subdominant | Izvornik 2 |
| Microcystis aeruginosa | Subdominant | Mandra (2021) |
| Microcystis sp. (separate cells) | Subdominant | Duvanli |
| Oscillatoria cf. simplicissima | Subdominant | Burgasko Ezero (2021) |
| Planktolyngbya limnetica | Subdominant | Eleshnitsa |
| Pseudanabaena mucicola | Subdominant | Nikolovo |
| Raphidiopsis raciborskii + Pseudanabaena limnetica | Subdominants | Shabla (2019) |
| Chlorophyta | ||
| Binuclearia lauterbornii | Dominant/Subdominant | Sopot, Tsonevo (2019)/Durankulak (2019), Uzungeren (2018) |
| Gloeocystis sp. | Dominant | Ezerets (2018), Shabla (2018) |
| Monactinus simplex | Dominant | Hadzhi Yani |
| Oocystis sp. | Dominant | Al. Stamboliyski, Zhrebchevo |
| Siderocystopsis pseudoblonga | Codominant | Shilkovtsi |
| Coelastrum astroideum + Tetrallantos lagerheimii | Subdominants | Hadzhidimovo |
| Didymocystis inconspicua + Pediastrum duplex | Subdominants | Poroy (2018) |
| Elakatothrix lacustris | Subdominant | Al. Stamboliyski |
| Golenkinia radiata | Subdominant | Plachidol 2 |
| Hariotina polychorda | Subdominant | Suedinenie |
| Lauterborniella appendiculata + Lobocystis sp. | Subdominants | Durankulak (2019) |
| Scenedesmus ellipticus | Subdominant | Aheloy |
| Streptophyta | ||
| Cosmarium neodepressum var. planctonicum | Dominant | Fisek |
| Closterium acerosum | Subdominant | Uzungeren (2018) |
| Cosmarium phaseolus var. elevatum | Subdominant | Dubnitsa |
| Pyrrhophyta | ||
| Ceratium rhomvoides | Dominant | Al. Stamboliyski |
| Peridinium volzii var. cinctiforme | Dominant | Suedinenie |
| Parvodinium elpatiewskyi | Dominant/Subdominant | Birgo, Dubnitsa/Satovcha 2 |
| Parvodinium cunningtonii | Codominant | Ablanitsa |
| Parvodinium umbonatum | Dominant | Hadzhidimovo |
| Sphaerodinium polonicum | Dominant | Duvanli, Eleshnitsa (2019) |
| Ceratium furcoides | Subdominant | Mandra (2018), Suedinenie |
| Parvodinium goslaviense | Subdominant | Mechka, Mogila |
| Euglenophyta | ||
| Euglena adhaerens | Dominant | Uzungeren (2021) |
| Euglenaria clavata | Dominant | Satovcha 2 |
| Phacus rotundus | Codominant | Hadzhi Yani |
| Discoplastis spathirhyncha | Subdominant | Kriva Reka |
| Euglena sp. | Subdominant | Uzungeren (2018) |
| Trachelomonas hispida | Subdominant | Birgo |
| Trachelomonas intermedia | Subdominant | Satovcha 2 |
| Ochrophyta | ||
| Bacillariophyceae | ||
| Asterionella formosa | Codominant | Dospat |
| Coscinodiscus sp. | Dominant/Subdominant | Durankulak (2021)/Mandra (2021), Poroy (2021) |
| Ctenophora pulchella | Dominant | Shumensko Ezero |
| Cymbella cf. cymbiformis | Dominant | Mandra (2021), Tsonevo (2018) |
| Fragilaria crotonensis | Codominant | Shilkovtsi |
| Lindavia comta | Dominant | Beglika, Chetiridesette Izvora |
| Nitzschia acicularis | Dominant | Uzungeren (2018) |
| Ulnaria acus | Dominant | Ezerets (2019) |
| Ulnaria ulna | Codominant | Eleshnitsa (2021) |
| Stephanocyclus meneghinianus | Subdominant | Koprinka |
| Chrysophyceae | ||
| Dinobryon bavaricum | Dominant/Codominant | Shiroka Polyana/Eleshnitsa (2021) |
| Dinobryon korschikovii | Dominant | Sopot |
| Synurophyceae | ||
| Mallomonas akrokomos | Codominant | Dospat |
| Xanthophyceae | ||
| Nephrodiella cf. acuta | Dominant | Uzungeren (2019) |
| Cryptophyta | ||
| Cryptomonas erosa | Dominant/Subdominant | Koprinka/Durankulak (2021) |
| Cryptomonas cf. ovata | Subdominant | Shabla (2018) |
| Cryptomonas sp. | Subdominant | Uzungeren (2019) |
| Waterbody/Year | Toxins | Species | Reference |
|---|---|---|---|
| 2018 | |||
| Burgasko Ezero | CPS | Microcystis aeruginosa, Microcystis wesenbergii, Microcystis novacekii; Cuspidothrix issatschenkoi, Cuspidothrix tropicalis | [38,39,43] |
| Durankulak | MC-LR, MC-RR, MC-YR, SXT | Microcystis aeruginosa, Microcystis wesenbergii | [38,39] |
| Mandra | Microcystis novacekii | [39] | |
| Poroy | Microcystis novacekii; Cuspidothrix issatschenkoi, Cuspidothrix tropicalis | [39,43] | |
| Sinyata Reka | MC-LR, MC-RR | Microcystis wesenbergii; Cuspidothrix issatschenkoi, Cuspidothrix tropicalis | [38,39,43] |
| 2019 | |||
| Burgasko Ezero | MV-CBJ | Microcystis aeruginosa, Microcystis wesenbergii, Cuspidothrix elenkinii, Cuspidothrix issatschenkoi, Cuspidothrix tropicalis | [40,41,43] |
| Durankulak | MC-LR, MV-CBJ | Microcystis aeruginosa | [40,41,42] |
| Duvanli | Microcystis aeruginosa, Microcystis viridis (?), Cuspidothrix issatschenkoi | [42,43] | |
| Ezerets | Microcystis aeruginosa, Microcystis viridis (?) | [42] | |
| Izvornik 2 | Microcystis wesenbergii | [40] | |
| Koprinka | Microcystis aeruginosa, Microcystis viridis (?), Microcystis wesenbergii, Cuspidothrix elenkinii | [42,43] | |
| Malka Smolnitsa | Microcystis aeruginosa, Microcystis viridis (?) | [42] | |
| Mandra | MC-LR, MV-CBJ | Microcystis aeruginosa | [40,41] |
| Plachidol 2 | Cuspidothrix issatschenkoi | [43] | |
| Preselka | Microcystis aeruginosa, Microcystis viridis | [42] | |
| Poroy | MV-A, MV/MC19 | Microcystis aeruginosa, Microcystis viridis, Microcystis wesenbergii | [40,41] |
| Sinyata Reka | MV-B/C | Microcystis aeruginosa, Microcystis wesenbergii, Cuspidothrix tropicalis | [40,41,43] |
| Uzungeren | Microcystis aeruginosa | [40] | |
| Zhrebchevo | Microcystis aeruginosa, Microcystis viridis (?), Microcystis wesenbergii | [42] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stoyneva-Gärtner, M.P.; Descy, J.-P.; Uzunov, B.A.; Miladinov, P.; Stefanova, K.; Radkova, M.; Gärtner, G. Diversity of the Summer Phytoplankton of 43 Waterbodies in Bulgaria and Its Potential for Water Quality Assessment. Diversity 2023, 15, 472. https://doi.org/10.3390/d15040472
Stoyneva-Gärtner MP, Descy J-P, Uzunov BA, Miladinov P, Stefanova K, Radkova M, Gärtner G. Diversity of the Summer Phytoplankton of 43 Waterbodies in Bulgaria and Its Potential for Water Quality Assessment. Diversity. 2023; 15(4):472. https://doi.org/10.3390/d15040472
Chicago/Turabian StyleStoyneva-Gärtner, Maya P., Jean-Pierre Descy, Blagoy A. Uzunov, Peter Miladinov, Katerina Stefanova, Mariana Radkova, and Georg Gärtner. 2023. "Diversity of the Summer Phytoplankton of 43 Waterbodies in Bulgaria and Its Potential for Water Quality Assessment" Diversity 15, no. 4: 472. https://doi.org/10.3390/d15040472
APA StyleStoyneva-Gärtner, M. P., Descy, J. -P., Uzunov, B. A., Miladinov, P., Stefanova, K., Radkova, M., & Gärtner, G. (2023). Diversity of the Summer Phytoplankton of 43 Waterbodies in Bulgaria and Its Potential for Water Quality Assessment. Diversity, 15(4), 472. https://doi.org/10.3390/d15040472