
An Illusion of Barriers to Gene Flow in Suburban Coyotes (Canis latrans): Spatial and Temporal Population Structure across a Fragmented Landscape in Southern California



Abstract
:1. Introduction
2. Materials and Methods
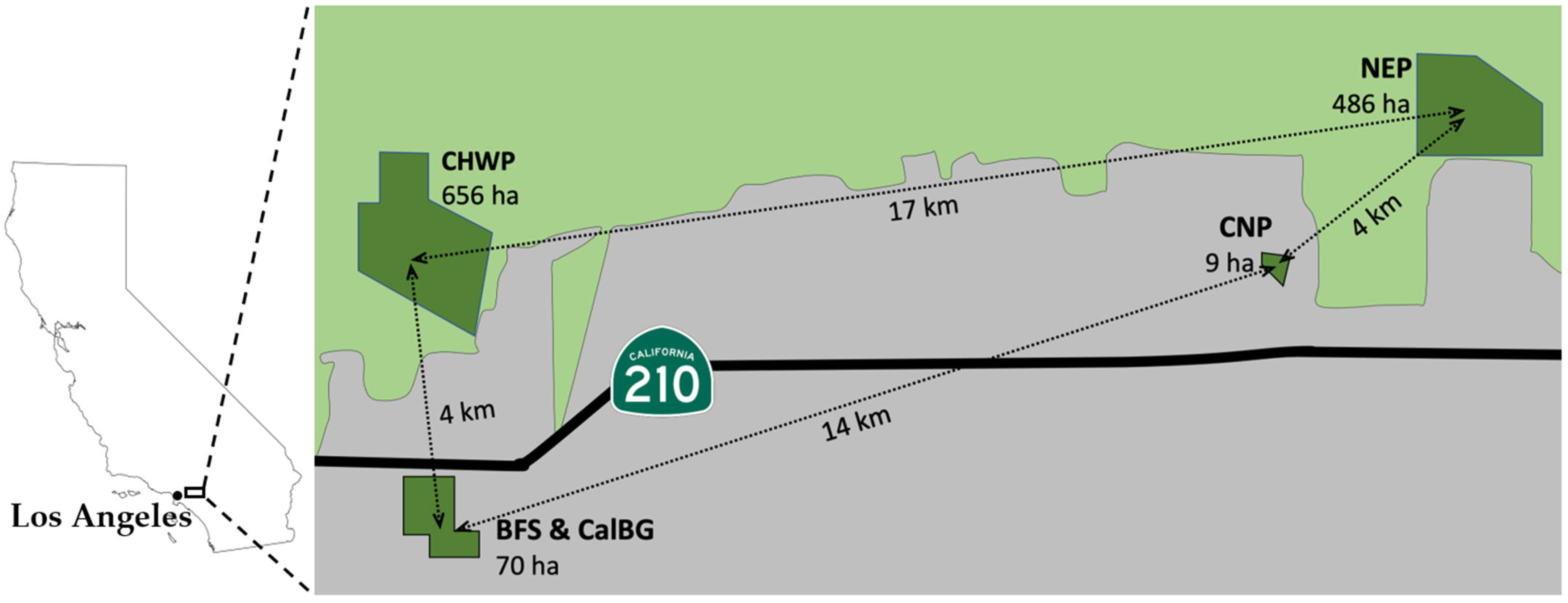
2.1. Study Sites
2.2. Scat Collection
2.3. Laboratory Methods
2.4. Genetic Analysis
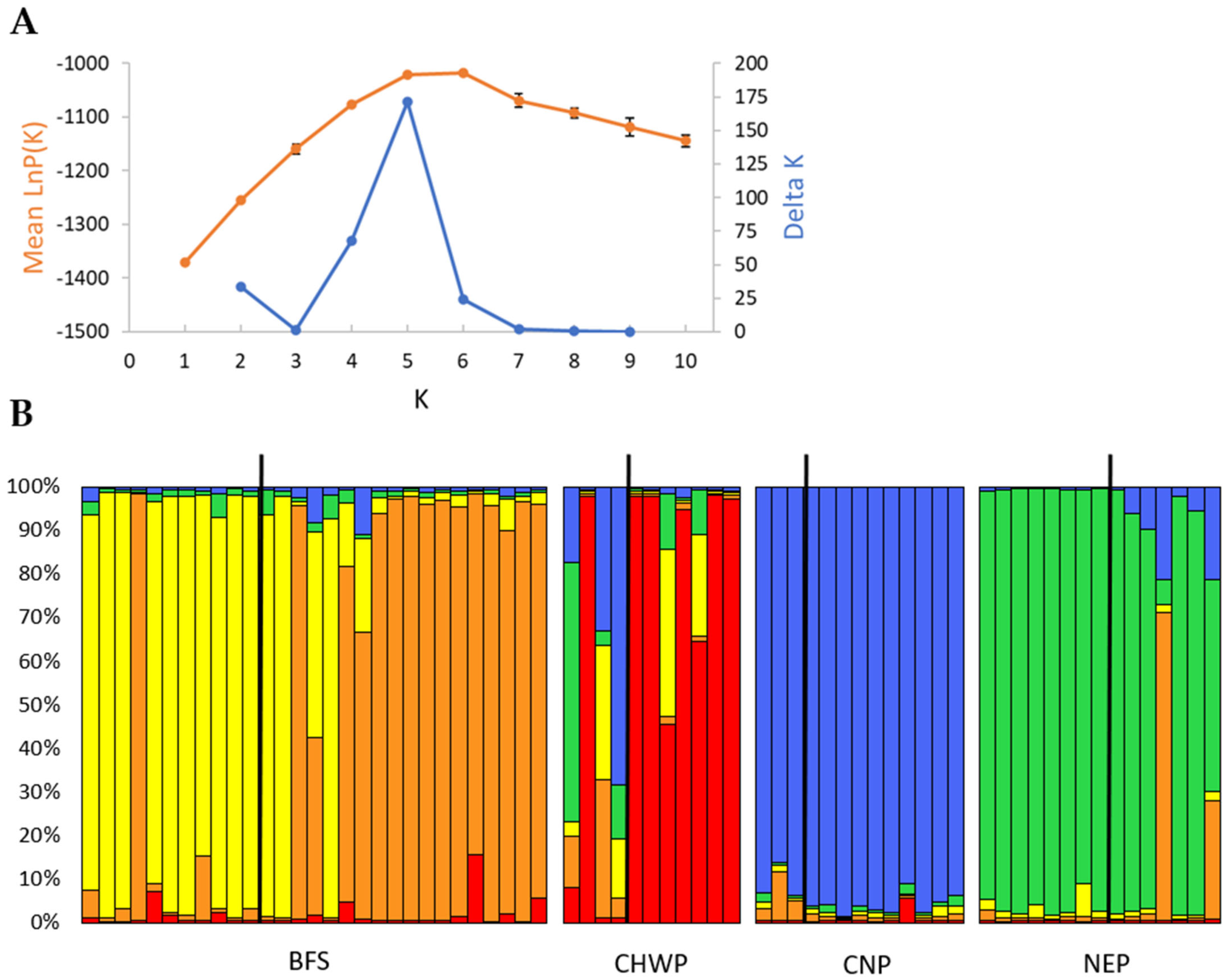
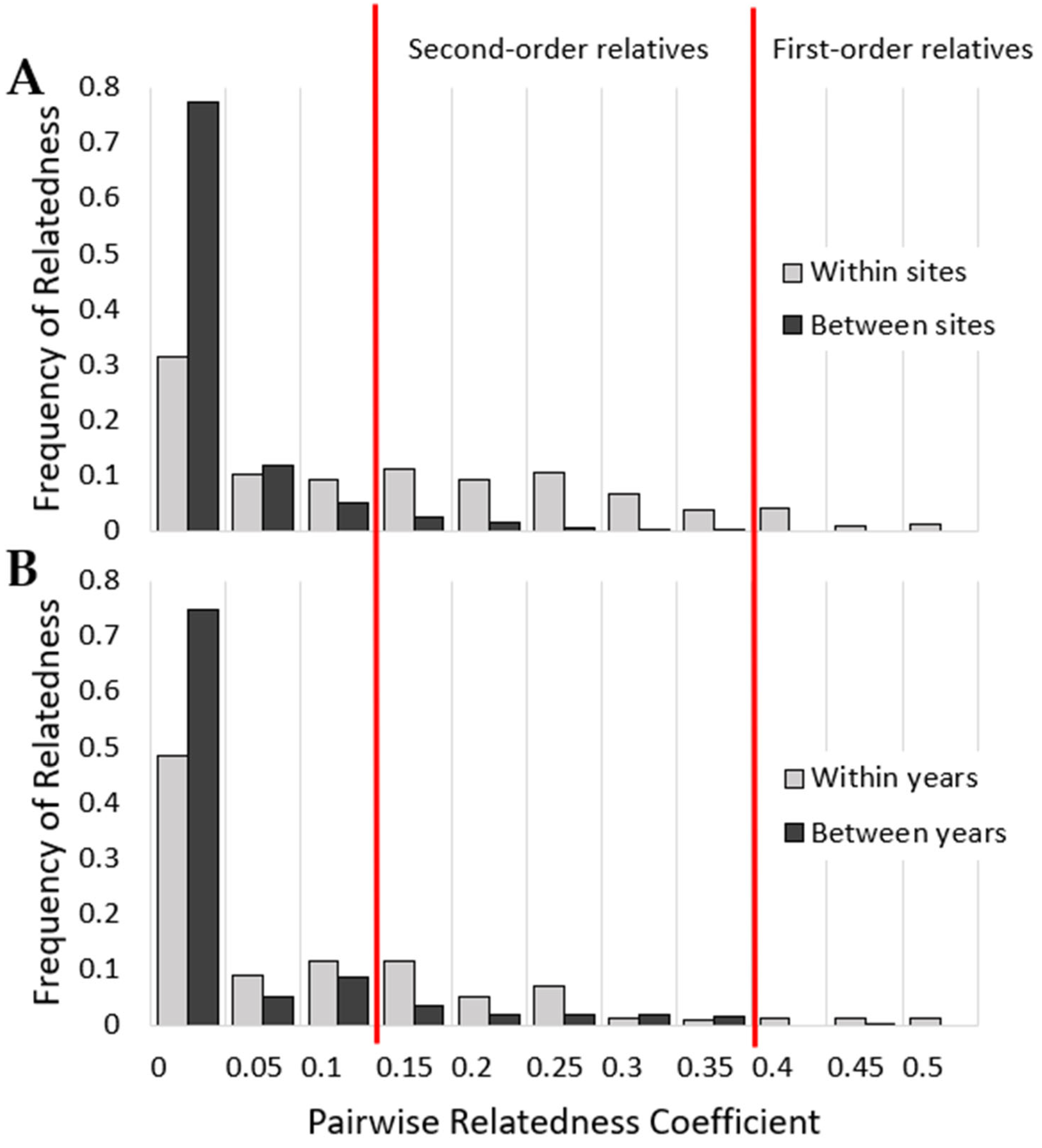
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ballou, J.D.; Rohner, C.; Breitenmoser, U.; Breitenmoser-Würsten, C. General Problems of Small (Carnivore) Populations: Minimum Viable Population Size and Inbreeding. In The Re-Introduction of the Lynx into the Alps: Proceedings of the 1st SCALP Conference, Engelberg, Switzerland, 7–9 December 1995; Council of Europe: Strasbourg, France, 1998; ISBN 978-92-871-3559-9. [Google Scholar]
- Liberg, O.; Andrén, H.; Pedersen, H.-C.; Sand, H.; Sejberg, D.; Wabakken, P.; Åkesson, M.; Bensch, S. Severe Inbreeding Depression in a Wild Wolf Canis Lupus Population. Biol. Lett. 2005, 1, 17–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, W.E.; Onorato, D.P.; Roelke, M.E.; Land, E.D.; Cunningham, M.; Belden, R.C.; McBride, R.; Jansen, D.; Lotz, M.; Shindle, D.; et al. Genetic Restoration of the Florida Panther. Science 2010, 329, 1641–1645. [Google Scholar] [CrossRef]
- Ruiz-López, M.J.; Gañan, N.; Godoy, J.A.; Del Olmo, A.; Garde, J.; Espeso, G.; Vargas, A.; Martinez, F.; Roldán, E.R.S.; Gomendio, M. Heterozygosity-Fitness Correlations and Inbreeding Depression in Two Critically Endangered Mammals. Conserv. Biol. 2012, 26, 1121–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fahrig, L. Effect of Habitat Fragmentation on the Extinction Threshold: A Synthesis. Ecol. Appl. 2002, 12, 346–353. [Google Scholar] [CrossRef]
- Delaney, K.S.; Riley, S.P.D.; Fisher, R.N. A Rapid, Strong, and Convergent Genetic Response to Urban Habitat Fragmentation in Four Divergent and Widespread Vertebrates. PLoS ONE 2010, 5, e12767. [Google Scholar] [CrossRef] [Green Version]
- Caspi, T.; Johnson, J.R.; Lambert, M.R.; Schell, C.J.; Sih, A. Behavioral Plasticity Can Facilitate Evolution in Urban Environments. Trends Ecol. Evol. 2022, 37, 1092–1103. [Google Scholar] [CrossRef]
- Caspi, T.; Hartz, L.A.; Soto Villa, A.E.; Loesberg, J.A.; Robins, C.R.; Meyer, W.M., III. Impacts of Invasive Annuals on Soil Carbon and Nitrogen Storage in Southern California Depend on the Identity of the Invader. Ecol. Evol. 2019, 9, 4980–4993. [Google Scholar] [CrossRef]
- Keeley, J.E.; Swift, C.C. Biodiversity and Ecosystem Functioning in Mediterranean-Climate California. In Mediterranean-Type Ecosystems; Davis, G.W., Richardson, D.M., Eds.; Ecological Studies; Springer: Berlin/Heidelberg, Germany, 1995; Volume 109, pp. 121–183. ISBN 978-3-642-78883-3. [Google Scholar]
- Nisi, A.C.; Benson, J.F.; King, R.; Wilmers, C.C. Habitat Fragmentation Reduces Survival and Drives Source–Sink Dynamics for a Large Carnivore. Ecol. Appl. 2023, e2822. [Google Scholar] [CrossRef] [PubMed]
- Gehrt, S.D.; Wilson, E.C.; Brown, J.L.; Anchor, C. Population Ecology of Free-Roaming Cats and Interference Competition by Coyotes in Urban Parks. PLoS ONE 2013, 8, e75718. [Google Scholar] [CrossRef] [PubMed]
- Kays, R.; Costello, R.; Forrester, T.; Baker, M.C.; Parsons, A.W.; Kalies, E.L.; Hess, G.; Millspaugh, J.J.; McShea, W. Cats Are Rare Where Coyotes Roam. J. Mammal. 2015, 96, 981–987. [Google Scholar] [CrossRef] [Green Version]
- Soulé, M.E.; Bolger, D.T.; Alberts, A.C.; Wrights, J.; Sorice, M.; Hill, S. Reconstructed Dynamics of Rapid Extinctions of Chaparral-Requiring Birds in Urban Habitat Islands. Conserv. Biol. 1988, 2, 75–92. [Google Scholar] [CrossRef] [Green Version]
- Crooks, K.R.; Soulé, M.E. Mesopredator Release and Avifaunal Extinctions in a Fragmented System. Nature 1999, 400, 563–566. [Google Scholar] [CrossRef]
- Loss, S.R.; Marra, P.P. Population Impacts of Free-Ranging Domestic Cats on Mainland Vertebrates. Front. Ecol. Environ. 2017, 15, 502–509. [Google Scholar] [CrossRef]
- Larson, R.N.; Brown, J.L.; Karels, T.; Riley, S.P.D. Effects of Urbanization on Resource Use and Individual Specialization in Coyotes (Canis latrans) in Southern California. PLoS ONE 2020, 15, e0228881. [Google Scholar] [CrossRef] [PubMed]
- Fedriani, J.M.; Fuller, T.K.; Sauvajot, R.M. Does Availability of Anthropogenic Food Enhance Densities of Omnivorous Mammals? An Example with Coyotes in Southern California. Ecography 2001, 24, 325–331. [Google Scholar] [CrossRef]
- Gehrt, S.D.; Anchor, C.; White, L.A. Home Range and Landscape Use of Coyotes in a Metropolitan Landscape: Conflict or Coexistence? J. Mammal. 2009, 90, 1045–1057. [Google Scholar] [CrossRef] [Green Version]
- Tigas, L.A.; Van Vuren, D.H.; Sauvajot, R.M. Behavioral Responses of Bobcats and Coyotes to Habitat Fragmentation and Corridors in an Urban Environment. Biol. Conserv. 2002, 108, 299–306. [Google Scholar] [CrossRef]
- Riley, S.P.D.; Pollinger, J.P.; Sauvajot, R.M.; York, E.C.; Bromley, C.; Fuller, T.K.; Wayne, R.K. FAST-TRACK: A Southern California Freeway Is a Physical and Social Barrier to Gene Flow in Carnivores. Mol. Ecol. 2006, 15, 1733–1741. [Google Scholar] [CrossRef]
- Adducci, A., II; Jasperse, J.; Riley, S.; Brown, J.; Honeycutt, R.; Monzón, J. Urban Coyotes Are Genetically Distinct from Coyotes in Natural Habitats. J. Urban Ecol. 2020, 6, juaa010. [Google Scholar] [CrossRef]
- Salafsky, N.; Salzer, D.; Stattersfield, A.J.; Hilton-Taylor, C.; Neugarten, R.; Butchart, S.H.M.; Collen, B.; Cox, N.; Master, L.L.; O’connor, S.; et al. A Standard Lexicon for Biodiversity Conservation: Unified Classifications of Threats and Actions. Conserv. Biol. 2008, 22, 897–911. [Google Scholar] [CrossRef]
- Andrews, A. Fragmentation of Habitat by Roads and Utility Corridors: A Review. Aust. Zool. 1990, 26, 130–141. [Google Scholar] [CrossRef] [Green Version]
- Forman, R.T.T.; Alexander, L.E. Roads and Their Major Ecological Effects. Annu. Rev. Ecol. Syst. 1998, 29, 207–231. [Google Scholar] [CrossRef] [Green Version]
- Frantzen, M.A.J.; Silk, J.B.; Ferguson, J.W.H.; Wayne, R.K.; Kohn, M.H. Empirical Evaluation of Preservation Methods for Faecal DNA. Mol. Ecol. 1998, 7, 1423–1428. [Google Scholar] [CrossRef] [Green Version]
- Adams, J.R.; Kelly, B.T.; Waits, L.P. Using Faecal DNA Sampling and GIS to Monitor Hybridization between Red Wolves (Canis rufus) and Coyotes (Canis latrans). Mol. Ecol. 2003, 12, 2175–2186. [Google Scholar] [CrossRef] [Green Version]
- Prugh, L.R.; Ritland, C.E. Molecular Testing of Observer Identification of Carnivore Feces in the Field. Wildl. Soc. Bull. 2005, 33, 189–194. [Google Scholar] [CrossRef]
- Francisco, L.V.; Langsten, A.A.; Mellersh, C.S.; Neal, C.L.; Ostrander, E.A. A Class of Highly Polymorphic Tetranucleotide Repeats for Canine Genetic Mapping. Mamm. Genome 1996, 7, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Breen, M.; Jouquand, S.; Renier, C.; Mellersh, C.S.; Hitte, C.; Holmes, N.G.; Chéron, A.; Suter, N.; Vignaux, F.; Bristow, A.E.; et al. Chromosome-Specific Single-Locus FISH Probes Allow Anchorage of an 1800-Marker Integrated Radiation-Hybrid/Linkage Map of the Domestic Dog Genome to All Chromosomes. Genome Res. 2001, 11, 1784–1795. [Google Scholar] [CrossRef] [Green Version]
- Galpern, P.; Manseau, M.; Hettinga, P.; Smith, K.; Wilson, P. Allelematch: An R Package for Identifying Unique Multilocus Genotypes Where Genotyping Error and Missing Data May Be Present. Mol. Ecol. Resour. 2012, 12, 771–778. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic Analysis in Excel. Population Genetic Software for Teaching and Research—An Update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raymond, M.; Rousset, F. GENEPOP (Version 1.2): Population Genetics Software for Exact Tests and Ecumenicism. J. Hered. 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Rice, W.R. Analyzing Tables of Statistical Tests. Evolution 1989, 43, 223–225. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of Population Structure Using Multilocus Genotype Data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Earl, D.A.; vonHoldt, B.M. STRUCTURE HARVESTER: A Website and Program for Visualizing STRUCTURE Output and Implementing the Evanno Method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Jakobsson, M.; Rosenberg, N.A. CLUMPP: A Cluster Matching and Permutation Program for Dealing with Label Switching and Multimodality in Analysis of Population Structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, M.; Ritland, K. Estimation of Pairwise Relatedness With Molecular Markers. Genetics 1999, 152, 1753–1766. [Google Scholar] [CrossRef] [PubMed]
- Bekoff, M.; Wells, M.C. Social Ecology and Behavior of Coyotes. In Advances in the Study of Behavior; Elsevier: Amsterdam, The Netherlands, 1986; Volume 16, pp. 251–338. ISBN 978-0-12-004516-7. [Google Scholar]
- Rashleigh, R.M.; Krebs, R.A.; van Keulen, H. Population Structure of Coyote (Canis latrans) in the Urban Landscape of the Cleveland, Ohio Area. Ohio J Sci 2008, 108, 54–59. [Google Scholar]
- Damm, D.L.; Armstrong, J.B.; Arjo, W.M.; Piaggio, A.J. Assessment of Population Structure of Coyotes in East-Central Alabama Using Microsatellite DNA. Southeast. Nat. 2015, 14, 106–122. [Google Scholar] [CrossRef] [Green Version]
- DeCandia, A.L.; Henger, C.S.; Krause, A.; Gormezano, L.J.; Weckel, M.; Nagy, C.; Munshi-South, J.; vonHoldt, B.M. Genetics of Urban Colonization: Neutral and Adaptive Variation in Coyotes (Canis latrans ) Inhabiting the New York Metropolitan Area. J. Urban Ecol. 2019, 5, juz002. [Google Scholar] [CrossRef] [Green Version]
- Sacks, B.N.; Mitchell, B.R.; Williams, C.L.; Ernest, H.B. Coyote Movements and Social Structure along a Cryptic Population Genetic Subdivision. Mol. Ecol. 2005, 14, 1241–1249. [Google Scholar] [CrossRef]
- Gehrt, S.D. Ecology of coyotes in urban landscapes. Wildl. Damage Manag. Conf. Proc. 2007, 63, 303–311. [Google Scholar]
- Kamler, J.F.; Gipson, P.S. Space and Habitat Use by Resident and Transient Coyotes. Can. J. Zool. 2000, 78, 2106–2111. [Google Scholar] [CrossRef]
- Hennessy, C.A.; Dubach, J.; Gehrt, S.D. Long-Term Pair Bonding and Genetic Evidence for Monogamy among Urban Coyotes (Canis latrans). J. Mammal. 2012, 93, 732–742. [Google Scholar] [CrossRef] [Green Version]
- Henger, C.S.; Herrera, G.A.; Nagy, C.M.; Weckel, M.E.; Gormezano, L.J.; Wultsch, C.; Munshi-South, J. Genetic Diversity and Relatedness of a Recently Established Population of Eastern Coyotes (Canis latrans) in New York City. Urban Ecosyst. 2020, 23, 319–330. [Google Scholar] [CrossRef]
- Atwood, T.; Weeks, H. Spatial Home-Range Overlap and Temporal Interaction in Eastern Coyotes: The Influence of Pair Types and Fragmentation. Can. J. Zool. 2011, 81, 1589–1597. [Google Scholar] [CrossRef]
- Grubbs, S.E.; Krausman, P.R. Use of Urban Landscape by Coyotes. Southwest. Nat. 2009, 54, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Riley, S.P.D.; Sauvajot, R.M.; Fuller, T.K.; York, E.C.; Kamradt, D.A.; Bromley, C.; Wayne, R.K. Effects of Urbanization and Habitat Fragmentation on Bobcats and Coyotes in Southern California. Conserv. Biol. 2003, 17, 566–576. [Google Scholar] [CrossRef] [Green Version]
- Sasmal, I.; Moorman, C.E.; Swingen, M.B.; Datta, S.; DePerno, C.S. Seasonal Space Use of Transient and Resident Coyotes (Canis latrans) in North Carolina, USA. Can. J. Zool. 2019, 97, 326–331. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, C.E.; Ruprecht, J.; Levi, T. More Affordable and Effective Noninvasive Single Nucleotide Polymorphism Genotyping Using High-Throughput Amplicon Sequencing. Mol. Ecol. Resour. 2020, 20, 1505–1516. [Google Scholar] [CrossRef]
- Pence, D.B.; Windberg, L.A.; Pence, B.C.; Sprowls, R. The Epizootiology and Pathology of Sarcoptic Mange in Coyotes, Canis latrans, from South Texas. J. Parasitol. 1983, 69, 1100–1115. [Google Scholar] [CrossRef] [Green Version]
- Cross, P.C.; Almberg, E.S.; Haase, C.G.; Hudson, P.J.; Maloney, S.K.; Metz, M.C.; Munn, A.J.; Nugent, P.; Putzeys, O.; Stahler, D.R.; et al. Energetic Costs of Mange in Wolves Estimated from Infrared Thermography. Ecology 2016, 97, 1938–1948. [Google Scholar] [CrossRef] [Green Version]
- Hill, J.E.; DeVault, T.L.; Wang, G.; Belant, J.L. Anthropogenic Mortality in Mammals Increases with the Human Footprint. Front. Ecol. Environ. 2020, 18, 13–18. [Google Scholar] [CrossRef]
- Hein, E.W.; Andelt, W.F. Estimating Coyote Density from Mark-Resight Surveys. J. Wildl. Manag. 1995, 59, 164–169. [Google Scholar] [CrossRef]
- Gese, E.M.; Morey, P.S.; Gehrt, S.D. Influence of the Urban Matrix on Space Use of Coyotes in the Chicago Metropolitan Area. J. Ethol. 2012, 30, 413–425. [Google Scholar] [CrossRef] [Green Version]
- Zepeda, E.; Payne, E.; Wurth, A.; Sih, A.; Gehrt, S. Early Life Experience Influences Dispersal in Coyotes (Canis latrans). Behav. Ecol. 2021, 32, 728–737. [Google Scholar] [CrossRef]
- Crooks, K.R. Relative Sensitivities of Mammalian Carnivores to Habitat Fragmentation. Conserv. Biol. 2002, 16, 488–502. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Ho (A)(H-W) * | |||||||
|---|---|---|---|---|---|---|---|
| Site | FH2001 | FH2010 | FH2062 | FH2096 | FH2137 | FH2140 | PEZ19 |
| BFS | 0.59 (4) | 0.53 (5) | 0.48 (4) | 0.62 (3) | 0.76 (7) | 0.82 (4) | 0.57 (5) |
| p = 0.005 | p = 0.139 | p = 0.776 | p = 0.521 | p = 0.118 | p = 0.002 | p = 0.021 | |
| CHWP | 1.00 (5) | 0.70 (4) | 0.57 (6) | 0.91 (6) | 0.33 (3) | 0.18 (5) (−) | 0.27 (5) (−) |
| p = 0.237 | p = 0.250 | p = 0.161 | p = 0.776 | p = 0.225 | p = 0.001 | p < 0.0001 | |
| CNP | 0.46 (3) | 0.30 (3) | 0.62 (4) | 0.62 (3) | 0.55 (4) | 0.54 (3) | 0.23 (2) |
| p = 1.000 | p = 1.000 | p = 0.260 | p = 0.850 | p = 0.596 | p = 1.000 | p = 1.000 | |
| NEP | 0.40 (3) | 0.92 (5) | 0.93 (4) | 0.73 (4) | 0.85 (4) | 0.87 (7) | 0.79 (5) |
| p = 0.589 | p = 0.837 | p = 0.429 | p = 0.706 | p = 0.141 | p = 0.200 | p = 0.181 | |
| All ** | 0.61 (7) | 0.61 (6) | 0.65 (6) | 0.72 (6) | 0.62 (7) | 0.60 (11) | 0.46 (6) |
| p = 0.074 | p = 0.047 | p = 0.160 | p = 0.069 | p = 0.0190 | p = 0.181 | p = 0.008 | |
| Site | N | Ho | He | A | H-W |
|---|---|---|---|---|---|
| BFS | 29 | 0.62 ± 0.05 | 0.63 ± 0.05 | 4.6 ± 0.48 | (−) p = 0.006 |
| CHWP | 11 | 0.57 ± 0.12 | 0.65 ± 0.05 | 4.9 ± 0.40 | p = 0.062 |
| CNP | 13 | 0.47 ± 0.06 | 0.46 ± 0.07 | 3.1 ± 0.26 | p = 0.212 |
| NEP | 15 | 0.78 ± 0.07 | 0.69 ± 0.05 | 4.6 ± 0.48 | p = 0.014 |
| All | 68 | 0.61 ± 0.04 | 0.61 ± 0.03 | 4.3 ± 0.24 | p = 0.0528 |
| BFS | CHWP | NEP | |
|---|---|---|---|
| CHWP | 0.112 | ||
| p < 0.001 | |||
| NEP | 0.085 | 0.128 | |
| p < 0.001 | p < 0.001 | ||
| CNP | 0.165 | 0.180 | 0.176 |
| p < 0.001 | p < 0.001 | p < 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bird, S.; Monzón, J.D.; Meyer, W.M., III; Moore, J.E. An Illusion of Barriers to Gene Flow in Suburban Coyotes (Canis latrans): Spatial and Temporal Population Structure across a Fragmented Landscape in Southern California. Diversity 2023, 15, 498. https://doi.org/10.3390/d15040498
Bird S, Monzón JD, Meyer WM III, Moore JE. An Illusion of Barriers to Gene Flow in Suburban Coyotes (Canis latrans): Spatial and Temporal Population Structure across a Fragmented Landscape in Southern California. Diversity. 2023; 15(4):498. https://doi.org/10.3390/d15040498
Chicago/Turabian StyleBird, Savanah, Javier D. Monzón, Wallace M. Meyer, III, and Jonathan E. Moore. 2023. "An Illusion of Barriers to Gene Flow in Suburban Coyotes (Canis latrans): Spatial and Temporal Population Structure across a Fragmented Landscape in Southern California" Diversity 15, no. 4: 498. https://doi.org/10.3390/d15040498
APA StyleBird, S., Monzón, J. D., Meyer, W. M., III, & Moore, J. E. (2023). An Illusion of Barriers to Gene Flow in Suburban Coyotes (Canis latrans): Spatial and Temporal Population Structure across a Fragmented Landscape in Southern California. Diversity, 15(4), 498. https://doi.org/10.3390/d15040498