

Distribution and Species Richness of Benthic Polychaeta and Sipuncula in the Northwestern Pacific



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
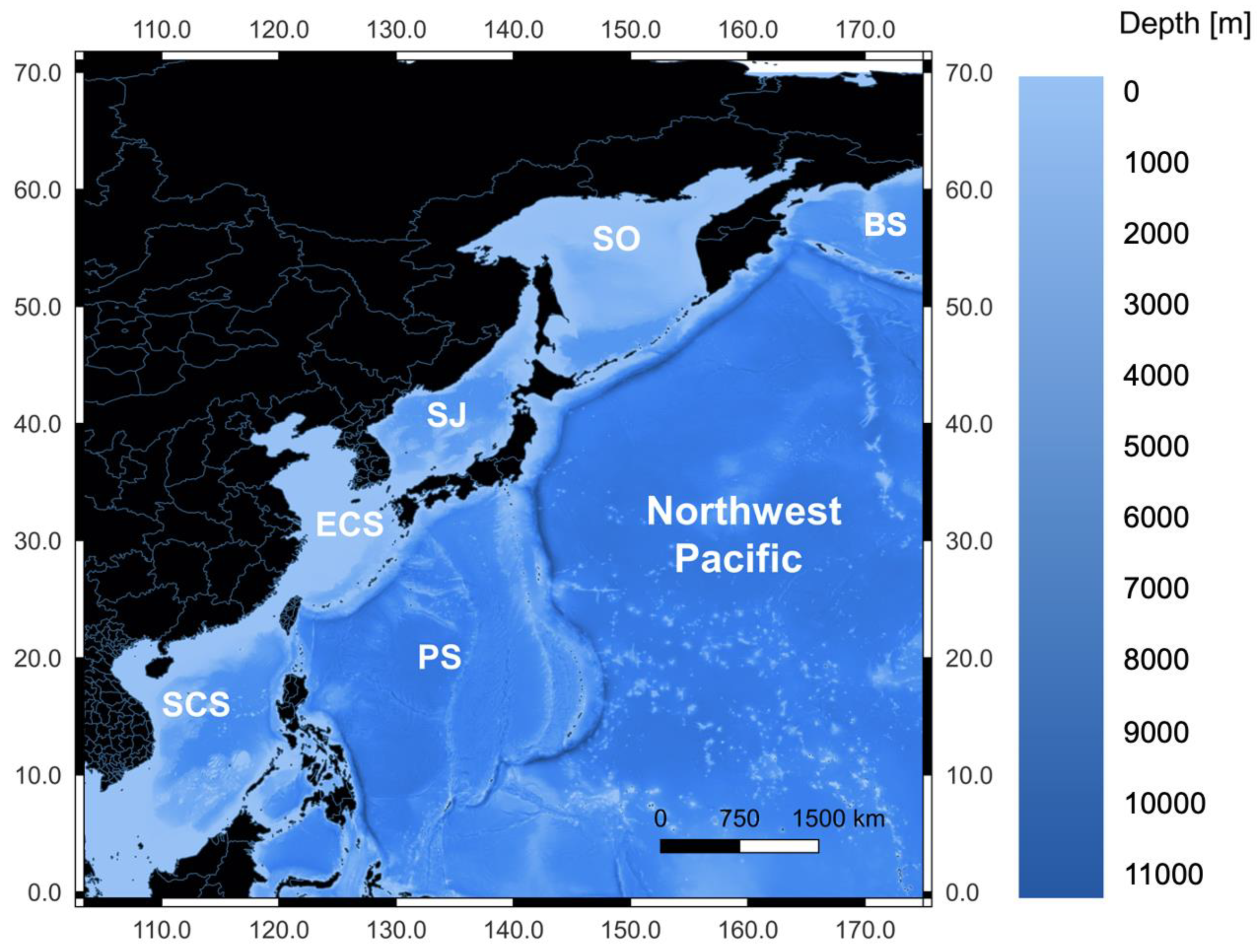
2.1. Study Area and Data Preparation
2.2. Data Analysis
3. Results
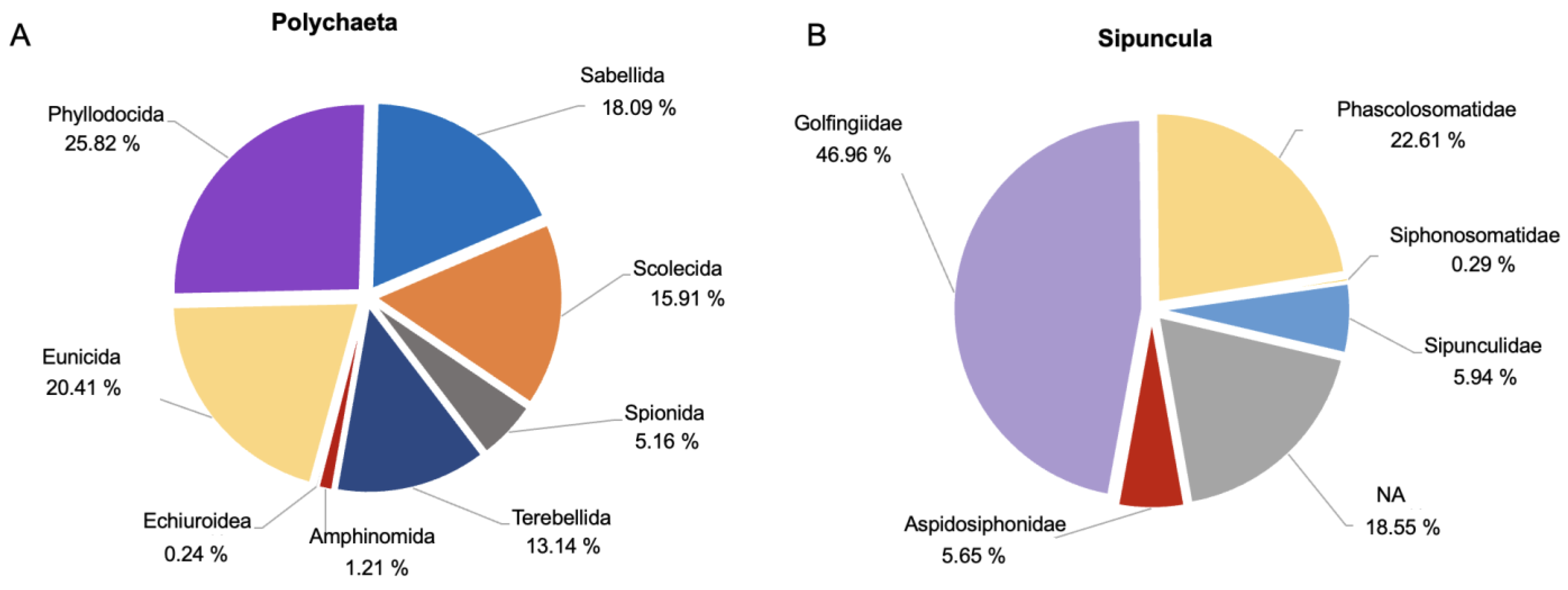
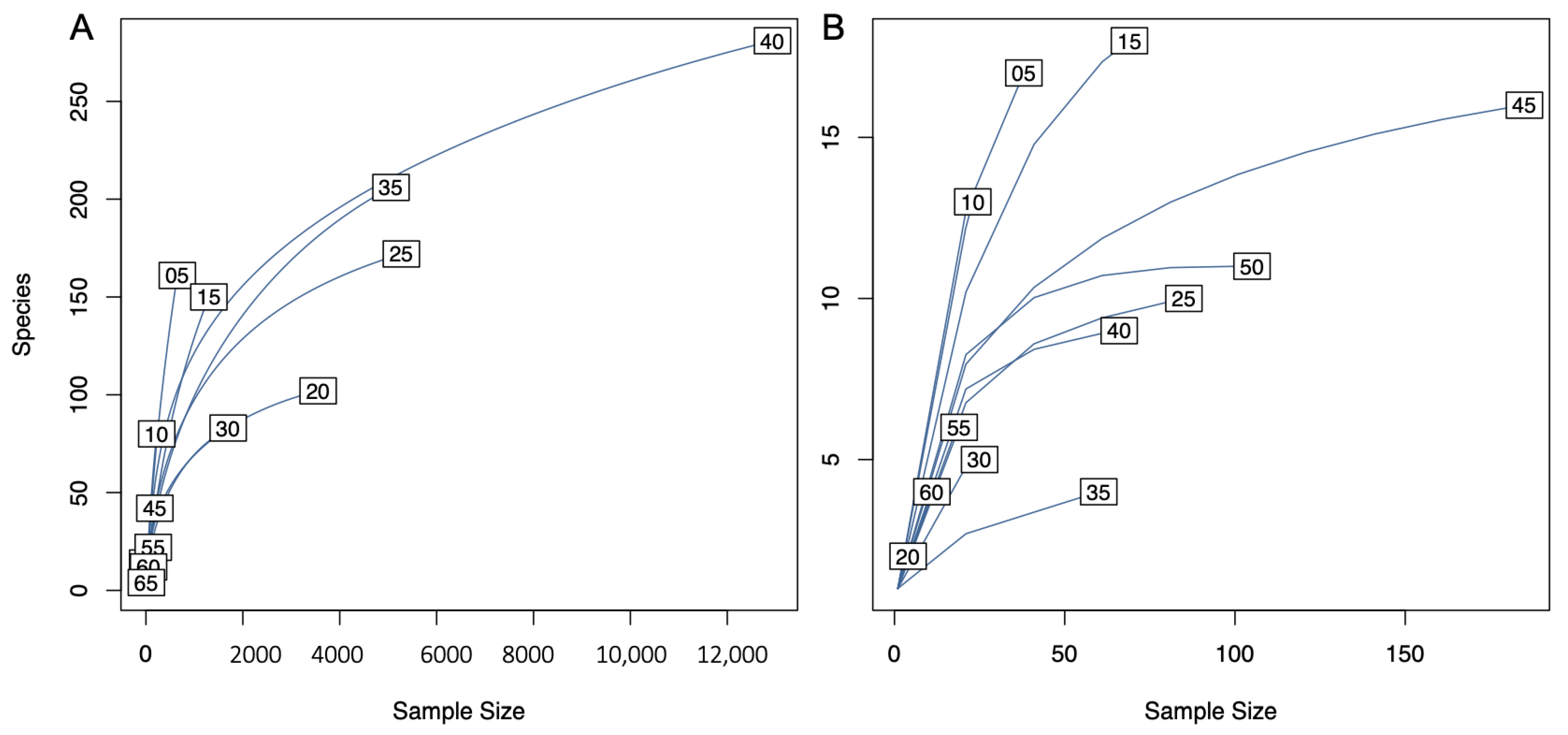
3.1. Dataset Composition
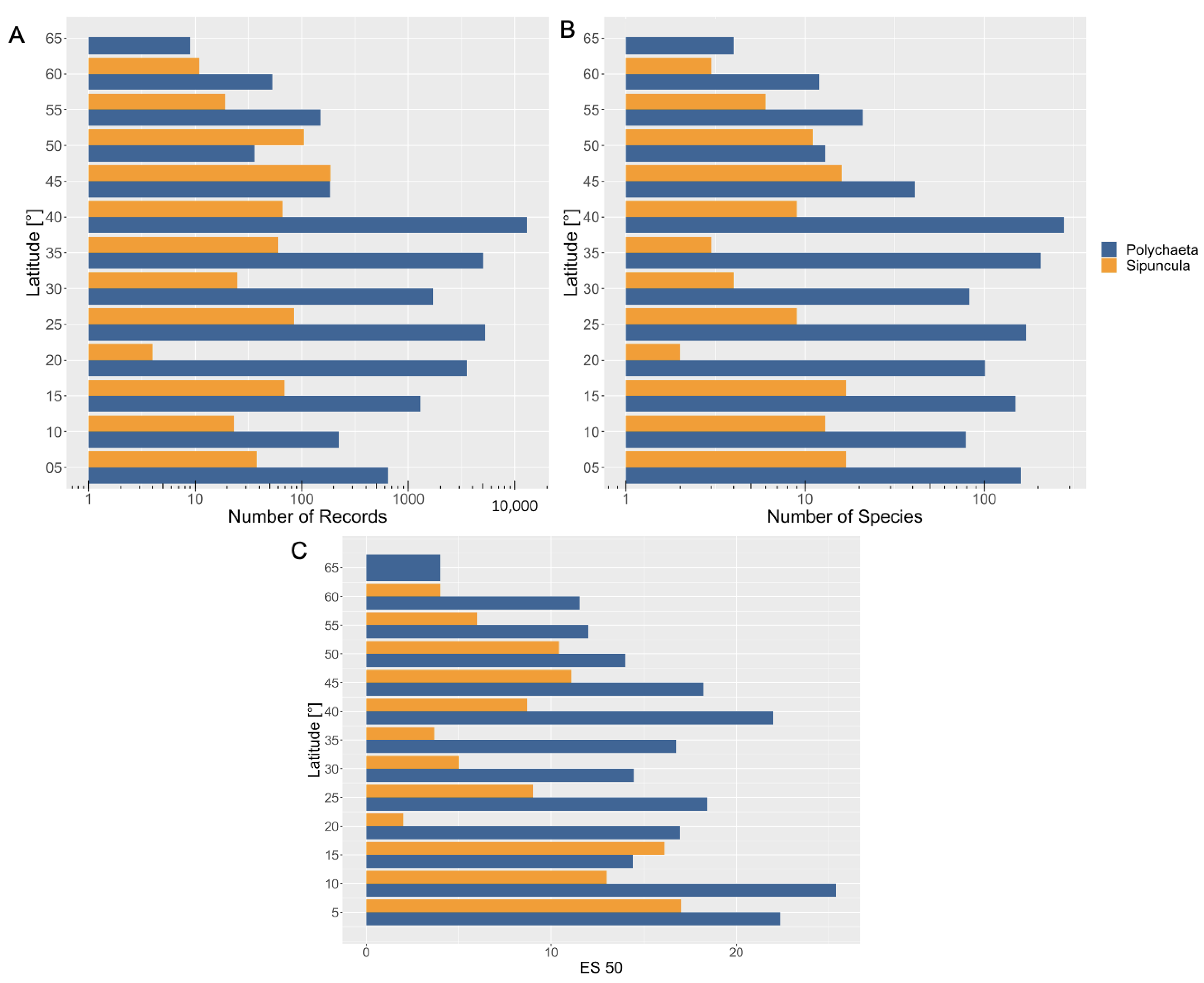
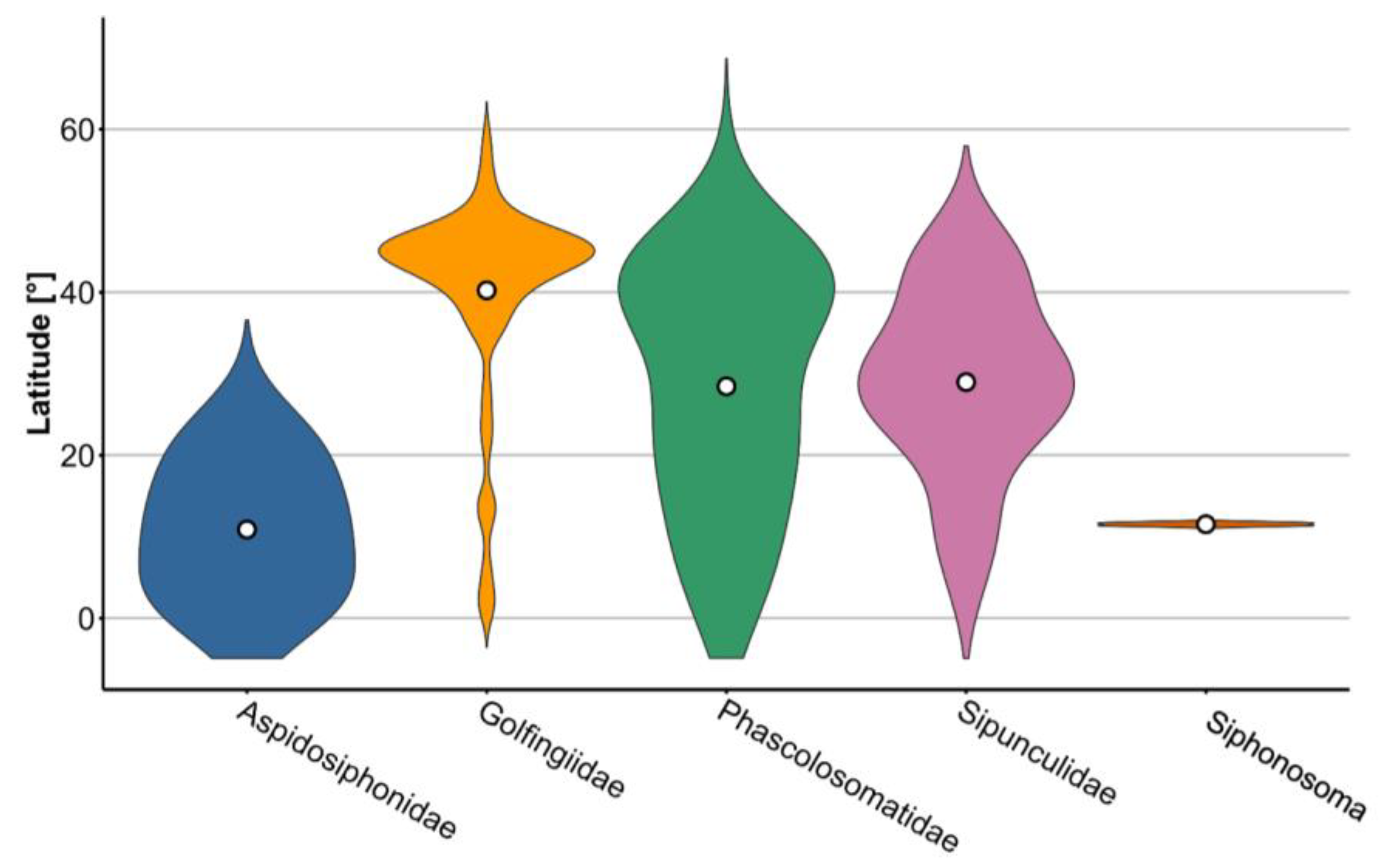
3.2. Latitudinal Distribution
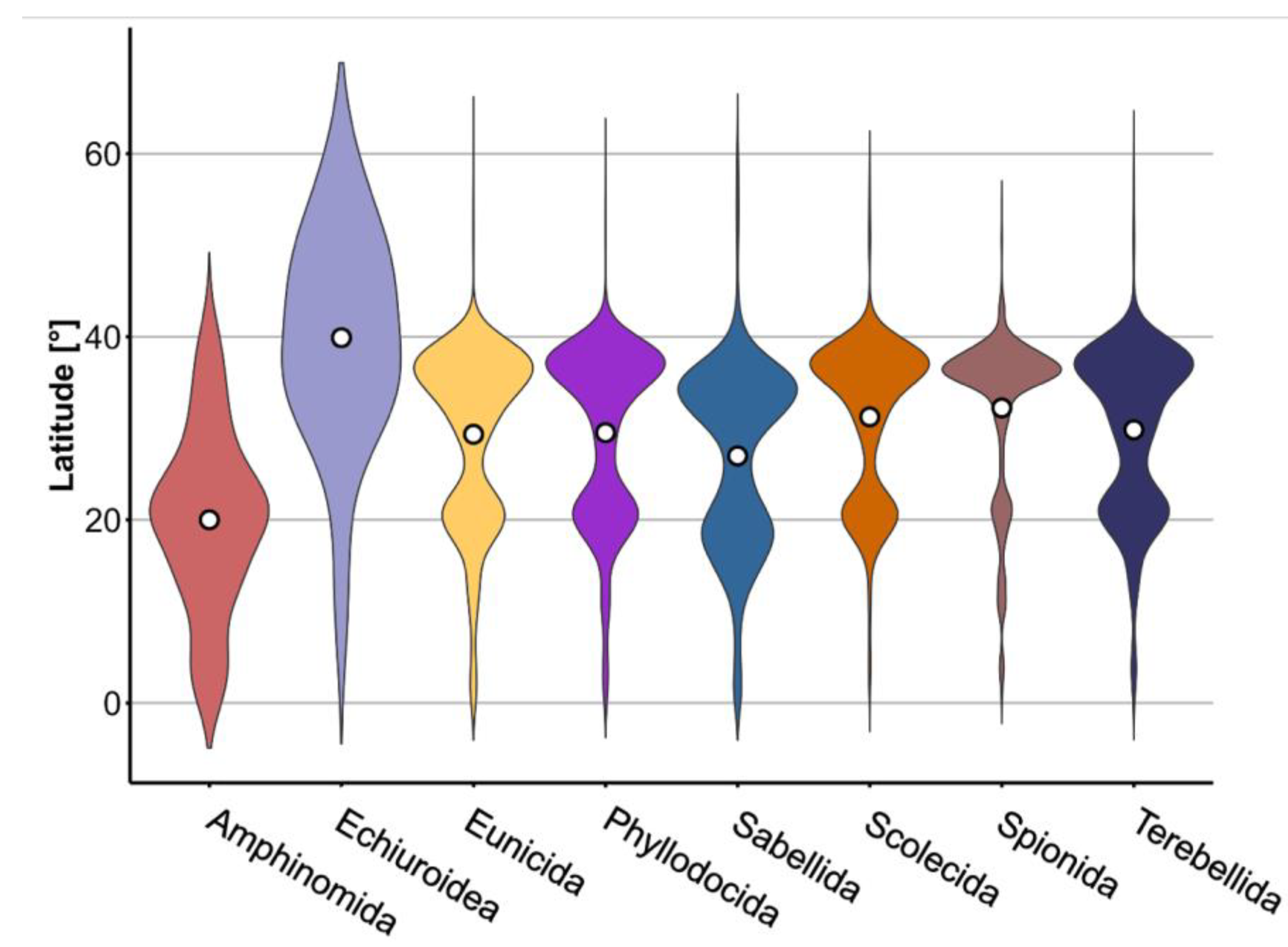
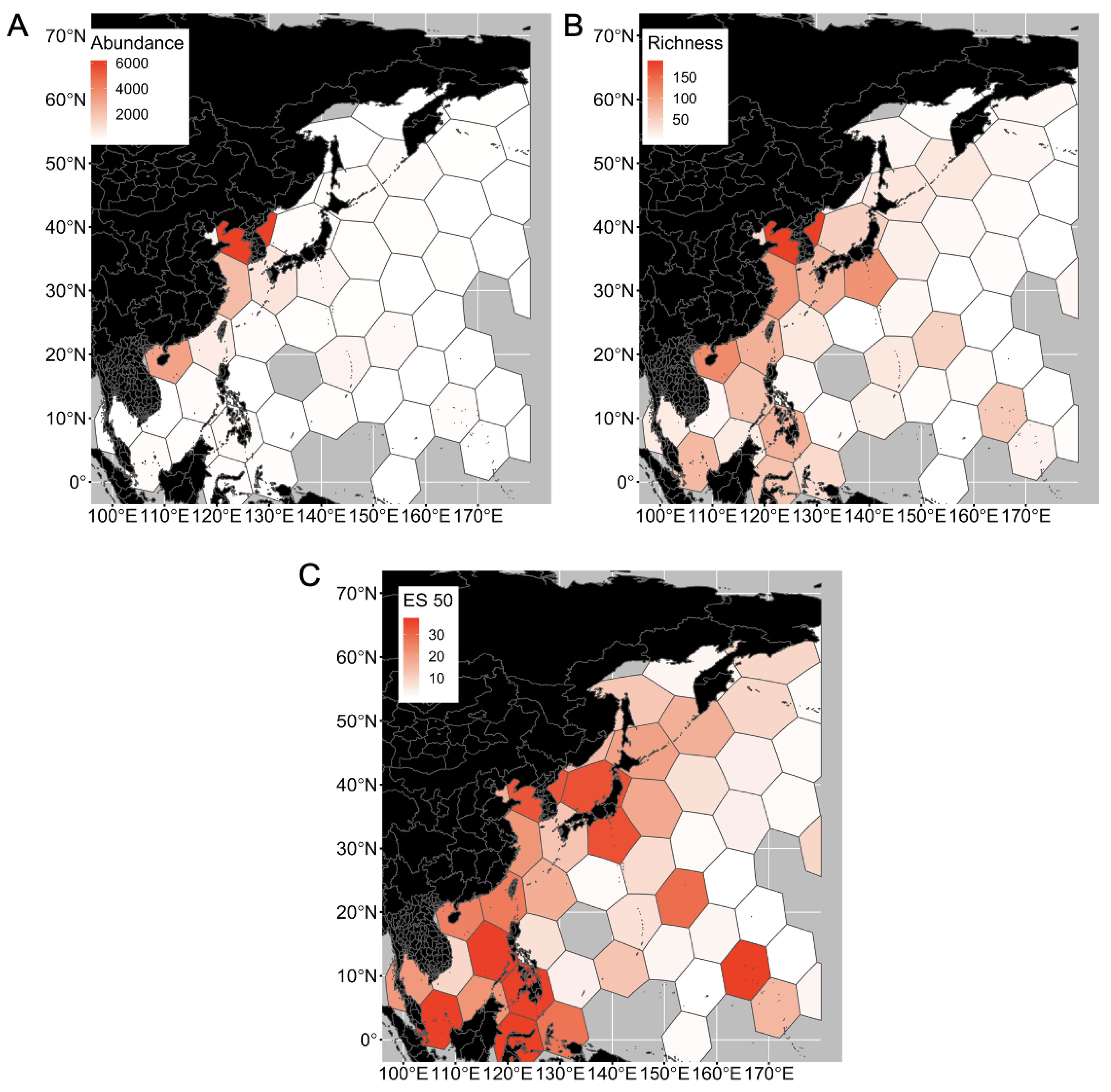
3.3. Distribution of Polychaeta and Sipuncula
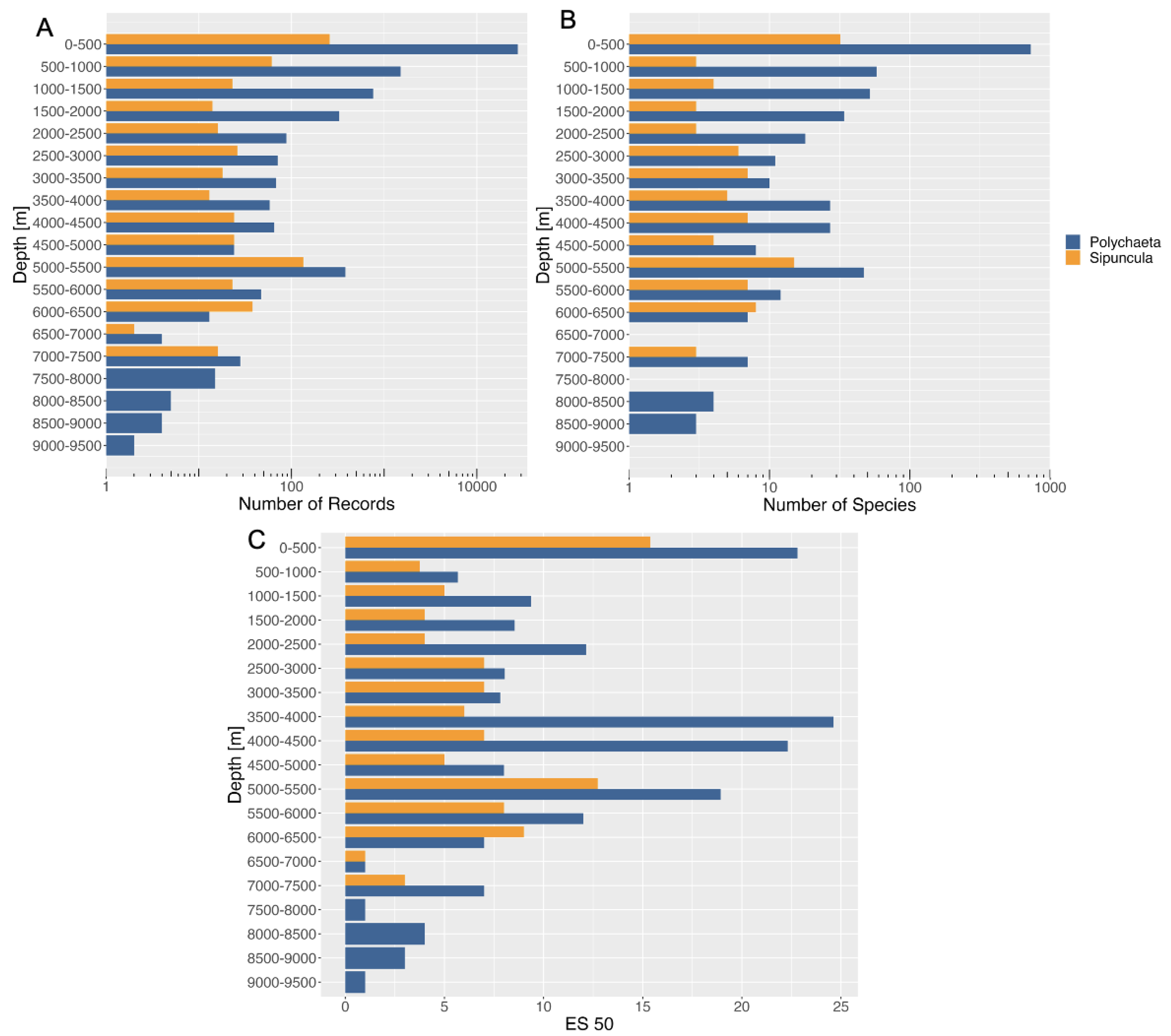
3.4. Bathymetric Distribution
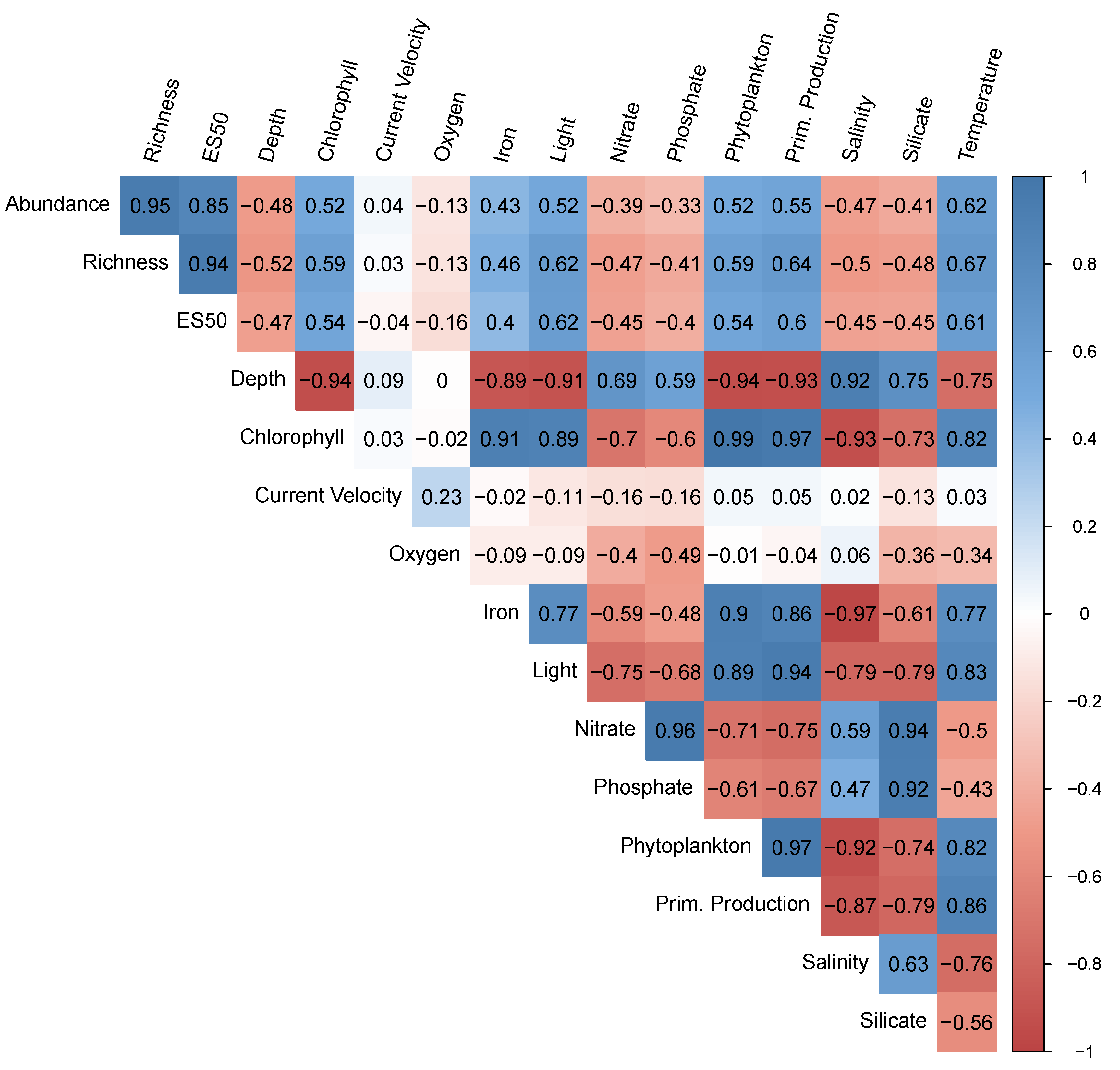
3.5. Environmental Variables
4. Discussion
4.1. Latitudinal Distribution
4.2. Bathymetric Distribution
4.3. Environmental Variables
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Birstein, J.A. Deep Water Isopods (Crustacea, Isopoda) of the North-Western Part of the Pacific Ocean; Indian National Scientific Documentation Centre: New Delhi, India, 1963. [Google Scholar]
- Brandt, A. RV Sonne SO-250 Cruise Report/Fahrtbericht. SO-250 KuramBio II (Kuril Kamchatka Biodiversity Studies) Tomakomai—Yokohama, Japan, 16.08.–26.09.2016. Available online: http://tristan.cordier.free.fr/files/SO250_KuramBio%20II_20161014.pdf (accessed on 4 November 2021).
- Brandt, A.; Brix, S.; Riehl, T.; Malyutina, M. Biodiversity and biogeography of the abyssal and hadal Kuril-Kamchatka trench and adjacent NW Pacific deep-sea regions. Prog. Oceanogr. 2020, 181, 102232. [Google Scholar] [CrossRef]
- Brandt, A.; Malyutina, M. The German-Russian Deep-Sea Expedition KuramBio (Kuril Kamchatka Biodiversity Studies) to the Kuril Kamchatka Trench and Abyssal Plain on Board of the R/V Sonne, 223rd Expedition (21 July–7 September 2012). 2012. Available online: https://www.researchgate.net/publication/298305628_The_German-Russian_deep-sea_expedition_KuramBio_Kurile_Kamchatka_Biodiversity_Study_to_the_Kurile_Kamchatka_Trench_and_abyssal_plain_on_board_of_the_RV_Sonne_223rd_Expedition_July_21th_-_September_7th (accessed on 4 November 2021).
- Brandt, A.; Malyutina, M.; Majorova, N.; Bashmanov, A.; Brenke, N.; Chizhova, T.; Elsner, N.; Golovan, O.; Göcke, C.; Kaplunenko, D.; et al. The Russian-German Deep-Sea Expedition (SoJaBio) to the Sea of Japan Onboard of the R/V Akademik Lavrentyev. 51st Cruise, 11 August–5 September 2010, The Cruise Report. 2010. Available online: https://www.researchgate.net/publication/298305568_The_Russian-German_deep-sea_expedition_SoJaBio_to_the_Sea_of_Japan_onboard_of_the_RV_Akademik_Lavrentyev_51st_Cruise_August_11th_-_September_5th_2010_The_Cruise_Report (accessed on 4 November 2021).
- Bogorov, V.G. (Ed.) Fauna of the Kurile-Kamchatka Trench and Its Environment, 86th ed.; Based on Data from the 39th Cruise of the R/V “Vityaz”: Proceedings of the Institue of Oceanology; IPST: Bangkok, Thailand, 1970. [Google Scholar]
- Malyutina, M.; Brandt, A.; Ivin, V.V. The Russian-German Deep-Sea Expedition SokhoBio (Sea of Okhotsk Biodiversity Studies) to the Kurile Basin of the Sea of Okhotsk on board of the R/V Akademik M.A.Lavrentyev. The Cruise Report. 2015.
- Werner, R.; Hoernle, K.; Hauff, F. RV SONNE Fahrtbericht/Cruise Report SO249. In BERING—Origin and Evolution of the Bering Sea: An Integrated Geochronological, Volcanological, Petrological and Geochemical Approach; GEOMAR Helmholtz-Zentrum für Ozeanforschung Kiel: Kiel, Germany, 2016; Volume 30. [Google Scholar]
- WoRMS Editorial Board World Register of Marine Species (WoRMS): WoRMS Editorial Board. 2022. Available online: https://www.marinespecies.org (accessed on 4 November 2021).
- Saeedi, H.; Jacobsen, N.L.; Brandt, A. Biodiversity and distribution of Isopoda and Polychaeta along the Northwestern Pacific and the Arctic Ocean. Biodivers. Inform. 2022, 17, 10–26. [Google Scholar] [CrossRef]
- Alalykina, I.L. Composition of deep-sea polychaetes from the SokhoBio expedition with a description of a new species of Labioleanira (Annelida: Sigalionidae) from the Sea of Okhotsk. Deep Sea Res. Part II Top. Stud. Oceanogr. 2018, 154, 140–158. [Google Scholar] [CrossRef]
- Alalykina, I.L. Polychaete composition from the abyssal plain adjacent to the Kuril–Kamchatka Trench with the description of a new species of Sphaerephesia (Polychaeta: Sphaerodoridae). Deep Sea Res. Part II Top. Stud. Oceanogr. 2015, 111, 166–174. [Google Scholar] [CrossRef]
- Fauchald, K. The polychaete worms. definition and keys to the orders, Families, and Genera. In Natural History Museum of Los Angeles County, Science Series; Los Angeles County Museum: Los Angeles, CA, USA, 1977; Volume 28. [Google Scholar]
- Saeedi, H.; Brandt, A. Biogeographic Atlas of the Deep NW Pacific Fauna; Pensoft Publishers: Sofia, Bulgaria, 2020. [Google Scholar]
- Kamenev, G.M.; Mordukhovich, V.V.; Alalykina, I.L.; Chernyshev, A.V.; Maiorova, A.S. Macrofauna and Nematode Abundance in the Abyssal and Hadal Zones of Interconnected Deep-Sea Ecosystems in the Kuril Basin (Sea of Okhotsk) and the Kuril-Kamchatka Trench (Pacific Ocean). Front. Mar. Sci. 2022, 9, 812464. [Google Scholar] [CrossRef]
- Hutchings, P. Biodiversity and functioning of polychaetes in benthic sediments. Biodivers. Conserv. 1998, 7, 1133–1145. [Google Scholar] [CrossRef]
- Sikorski, A.; Pavlova, L. Three new species of Laonice (Polychaete: Spionidae) from West and Southwest Africa. Zootaxa 2016, 4097, 353–368. [Google Scholar] [CrossRef]
- Pamungkas, J.; Glasby, C.J.; Read, G.B.; Wilson, S.P.; Costello, M.J. Progress and perspectives in the discovery of polychaete worms (Annelida) of the world. Helgol. Mar. Res. 2019, 73, 4. [Google Scholar] [CrossRef]
- Moreno, R.A.; Rivadeneira, M.M.; Hernandez, C.E.; Sampértegui, S.; Rozbaczylo, N. Do Rapoport´s rule, the mid-domain effect or the source-sink hypotheses predict bathymetric patterns of polychaete richness on the Pacific coast of South America? Glob. Ecol. Biogeogr. 2008, 17, 415–423. [Google Scholar] [CrossRef]
- Ellingsen, K.E.; Brandt, A.; Ebbe, B.; Linse, K. Diversity and species distribution of polychaetes, isopods and bivalves in the Atlantic sector of the deep Southern Ocean. Polar Biol. 2007, 30, 1265–1273. [Google Scholar] [CrossRef]
- Hilbig, B.; Blake, J.A. Deep-sea polychaete communities in the northeast Pacific Ocean off the Gulf of the Farallones, California. Bull. Mar. Sci. 2006, 78, 243–269. [Google Scholar]
- Brandt, A.; De Broyer, C.; De Mesel, I.; Ellingsen, K.E.; Gooday, A.J.; Hilbig, B.; Linse, K.; Thomson, M.R.; Tyler, P.A. The biodiversity of the deep Southern Ocean benthos. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2007, 362, 39–66. [Google Scholar] [CrossRef] [Green Version]
- Cutler, E.B. The Sipuncula: Their Systematics, Biology, and Evolution; Ithaka and London: Ithaca, NY, USA, 1994. [Google Scholar]
- Schulze, A.; Maiorova, A.; Timm, L.E.; Rice, M.E. Sipunculan Larvae and “Cosmopolitan” Species. Integr. Comp. Biol. 2012, 52, 497–510. [Google Scholar] [CrossRef] [Green Version]
- Dordel, J.; Fisse, F.; Purschke, G.; Struck, T.H. Phylogenetic position of Sipuncula derived from multi-gene and phylogenomic data and its implication for the evolution of segmentation. J. Zool. Syst. Evol. Res. 2010, 48, 197–207. [Google Scholar] [CrossRef]
- Mwinyi, A.; Meyer, A.; Bleidorn, C.; Lieb, B.; Bartolomaeus, T.; Podsiadlowski, L. Mitochondrial genome sequence and gene order of Sipunculus nudus give additional support for an inclusion of Sipuncula into Annelida. BMC Genom. 2009, 10, 27. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.; Ma, X.; Ren, J.; Zhao, F. A close phylogenetic relationship between Sipuncula and Annelida evidenced from the complete mitochondrial genome sequence of Phascolosoma esculenta. BMC Genom. 2009, 10, 136. [Google Scholar] [CrossRef] [Green Version]
- Sung, J.-M.; Karagozlu, M.Z.; Kim, C.-B. The complete mitogenome of a peanut worm Phascolosoma pacificum (Sipuncula, Phascolosomatida, Phascolosomatidea). Mitochondrial DNA Part B 2017, 2, 618–619. [Google Scholar] [CrossRef] [Green Version]
- Schulze, A.; Kawauchi, G.Y. How Many Sipunculan Species Are Hiding in Our Oceans? Diversity 2021, 13, 43. [Google Scholar] [CrossRef]
- Kędra, M.; Grebmeier, J.M.; Cooper, L.W. Sipunculan fauna in the Pacific Arctic region: A significant component of benthic infaunal communities. Polar Biol. 2018, 41, 163–174. [Google Scholar] [CrossRef] [Green Version]
- Kędra, M.; Murina, G.V. The sipunculan fauna of Svalbard. Polar Res. 2007, 26, 37–47. [Google Scholar] [CrossRef]
- Maiorova, A.S.; Adrianov, A.V. Distribution of peanut worms (Sipuncula) in the West Pacific. In Proceedings of the China-Russian Bilateral symposium on “Comparison on Marine Biodiversityin the Northwest Pacific Ocean”, Qingdao, China, 10–11 November 2010; pp. 10–11. [Google Scholar]
- Maiorova, A.S.; Adrianov, A.V. Peanut worms of the phylum Sipuncula from the Sea of Japan with a key to species. Deep Sea Res. Part II Top. Stud. Oceanogr. 2013, 86–87, 140–147. [Google Scholar] [CrossRef]
- Maiorova, A.S.; Adrianov, A.V. Deep-sea sipunculans from the Kuril-Kamchatka Trench and adjacent abyssal plain. Deep Sea Res. Part II Top. Stud. Oceanogr. 2015, 111, 128–134. [Google Scholar] [CrossRef]
- Wallace, A.R. Tropical Nature, and Other Essays; Macmillan and Company: New York, NY, USA, 1878. [Google Scholar]
- Menegotto, A.; Rangel, T.F. Mapping knowledge gaps in marine diversity reveals a latitudinal gradient of missing species richness. Nat. Commun. 2018, 9, 4713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macpherson, E.; Duarte, C.M. Patterns in species richness, size, and latitudinal range of East Atlantic fishes. Ecography 1994, 17, 242–248. [Google Scholar] [CrossRef]
- Fischer, A.G. Latitudinal variations in organic diversity. Evolution 1960, 14, 64–81. [Google Scholar] [CrossRef]
- Saeedi, H.; Dennis, T.E.; Costello, M.J. Bimodal latitudinal species richness and high endemicity of razor clams (Mollusca). J. Biogeogr. 2017, 44, 592–604. [Google Scholar] [CrossRef]
- Hillebrand, H. On the generality of the latitudinal diversity gradient. Am. Nat. 2004, 163, 192–211. [Google Scholar] [CrossRef] [Green Version]
- Crame, J.A. Evolution of taxonomic diversity gradients in the marine realm: A comparison of Late Jurassic and Recent bivalve faunas. Paleobiology 2002, 28, 184–207. [Google Scholar] [CrossRef]
- Böhm, M.; Mayhew, P.J. Historical biogeography and the evolution of the latitudinal gradient of species richness in the Papionini (Primata: Cercopithecidae). Biol. J. Linn. Soc. 2005, 85, 235–246. [Google Scholar] [CrossRef] [Green Version]
- Chaudhary, C.; Saeedi, H.; Costello, M.J. Bimodality of Latitudinal Gradients in Marine Species Richness. Trends Ecol. Evol. 2016, 31, 670–676. [Google Scholar] [CrossRef]
- Willig, M.R.; Kaufman, D.M.; Stevens, R.D. Latitudinal Gradients of Biodiversity: Pattern, Process, Scale, and Synthesis. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 273–309. [Google Scholar] [CrossRef]
- Zhao, F.; Filker, S.; Wang, C.; Xu, K. Bathymetric gradient shapes the community composition rather than the species richness of deep-sea benthic ciliates. Sci. Total Environ. 2021, 755, 142623. [Google Scholar] [CrossRef]
- Danovaro, R.; Gambi, C.; Lampadariou, N.; Tselepides, A. Deep-sea nematode biodiversity in the Mediterranean basin: Testing for longitudinal, bathymetric and energetic gradients. Ecography 2008, 31, 231–244. [Google Scholar] [CrossRef]
- Cartes, J.; Sardà, F. Abundance and diversity of decapod crustaceans in the deep-Catalan Sea (Western Mediterranean). J. Nat. Hist. 1992, 26, 1305–1323. [Google Scholar] [CrossRef] [Green Version]
- Saeedi, H.; Costello, M.J.; Warren, D.; Brandt, A. Latitudinal and bathymetrical species richness patterns in the NW Pacific and adjacent Arctic Ocean. Sci. Rep. 2019, 9, 9303. [Google Scholar] [CrossRef] [Green Version]
- Tittensor, D.P.; Mora, C.; Jetz, W.; Lotze, H.K.; Ricard, D.; Berghe, E.V.; Worm, B. Global patterns and predictors of marine biodiversity across taxa. Nature 2010, 466, 1098–1101. [Google Scholar] [CrossRef]
- Woolley, S.N.C.; Tittensor, D.P.; Dunstan, P.K.; Guillera-Arroita, G.; Lahoz-Monfort, J.J.; Wintle, B.A.; Worm, B.; O’Hara, T.D. Deep-sea diversity patterns are shaped by energy availability. Nature 2016, 533, 393–396. [Google Scholar] [CrossRef] [Green Version]
- Saeedi, H.; Simões, M.; Brandt, A. Biodiversity and distribution patterns of deep-sea fauna along the temperate NW Pacific. Prog. Oceanogr. 2020, 183, 102296. [Google Scholar] [CrossRef]
- Levin, L.A.; Wei, C.; Dunn, D.C.; Amon, D.J.; Ashford, O.S.; Cheung, W.W.L.; Colaço, A.; Dominguez-Carrió, C.; Escobar, E.G.; Harden-Davies, H.R.; et al. Climate change considerations are fundamental to management of deep-sea resource extraction. Glob. Chang. Biol. 2020, 26, 4664–4678. [Google Scholar] [CrossRef]
- McClain, C.R.; Hardy, S.M. The dynamics of biogeographic ranges in the deep sea. Proc. R. Soc. B Biol. Sci. 2010, 277, 3533–3546. [Google Scholar] [CrossRef] [Green Version]
- Hoegh-Guldberg, O.; Cai, R.; Poloczanska, E.S.; Brewer, P.G.; Sundby, S.; Hilmi, K.; Fabry, V.J.; Jun, S.E. The Ocean, in Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part B: Regional Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Barros, V.R., Field, C.B., Dokken, D.J., Mastrandrea, M.D., Mach, K.J., Bilir, T.E., Chatterjee, M., Ebi, K.L., Estrada, Y.O., Genova, R.C., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; pp. 1655–1731. [Google Scholar]
- Abram, N.; Gattuso, J.P.; Prakash, A.; Cheng, L.; Chidichimo, M.P.; Crate, S.; Enomoto, H.; Garschagen, M.; Gruber, N.; Harper, S.; et al. Special Report on the Ocean and Cryosphere in a Changing Climate; Cambridge University Press: Cambridge, UK, 2019; pp. 73–129. [Google Scholar] [CrossRef]
- Simões, M.V.P.; Saeedi, H.; Cobos, M.E.; Brandt, A. Environmental matching reveals non-uniform range-shift patterns in benthic marine Crustacea. Clim. Chang. 2021, 168, 31. [Google Scholar] [CrossRef]
- Sweetman, A.K.; Thurber, A.R.; Smith, C.R.; Levin, L.; Mora, C.; Wei, C.-L.; Gooday, A.J.; Jones, D.; Rex, M.; Yasuhara, M.; et al. Major impacts of climate change on deep-sea benthic ecosystems. Elementa: Sci. Anthr. 2017, 5, 2–23. [Google Scholar] [CrossRef]
- Danovaro, R.; Corinaldesi, C.; Dell’Anno, A.; Snelgrove, P.V.R. The deep-sea under global change. Curr. Biol. 2017, 27, R461–R465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez-Llodra, E.; Brandt, A.; Danovaro, R.; De Mol, B.; Escobar, E.; German, C.R.; Levin, L.A.; Martinez Arbizu, P.; Menot, L.; Buhl-Mortensen, P.; et al. Deep, diverse and definitely different: Unique attributes of the world’s largest ecosystem. Biogeosciences 2010, 7, 2851–2899. [Google Scholar] [CrossRef] [Green Version]
- Dreutter, S.; Steffen, M.; Arbizu, P.M.; Brandt, A. Will the “top five” deepest trenches lose one of their members? Prog. Oceanogr. 2020, 181, 102258. [Google Scholar] [CrossRef]
- Grassle, J.F. The Ocean Biogeographic Information System (OBIS): An on-line, worldwide atlas for accessing, modeling and mapping marine biological data in a multidimensional geographic context. Oceanography 2000, 13, 5–7. [Google Scholar] [CrossRef]
- Global Biodiversity Information Facility. 2021. Available online: www.gbif.org (accessed on 4 November 2021).
- Glover, A.G.; Higgs, N.; Horton, T. World Register of Deep-Sea Species (WoRDSS). 2021. Available online: http://www.marinespecies.org/deepsea (accessed on 12 November 2021).
- R Core Team. R. A Language and Environment for Statistical Computing, Version 2021.09.1; R Foundation for Statistical Computing: Vienna, Austria, 2021.
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.; Francois, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef] [Green Version]
- Chamberlain, S. Scrubr: Clean Biological Occurence Records. 2021.
- Provoost, P.; Bosch, S. Obistools: Tools for Data Enhancement and Quality Control: Ocean Biogeographic Information System. Intergovernmental Oceanographic Commission of UNESCO. 2022. Available online: https://cran.r-project.org/package=obistools (accessed on 4 November 2021).
- Group, G.C. GEBCO 2021 Grid. [2021 11.11.21]. Available online: https://www.gebco.net (accessed on 4 November 2021).
- Assis, J.; Tyberghein, L.; Bosch, S.; Verbruggen, H.; Serrão, E.A.; de Clerck, O. Extending marine data layers for bioclimatic modelling. Glob. Ecol. Biogeogr. 2018, 27, 277–284. [Google Scholar] [CrossRef]
- Tyberghein, L.; Verbruggen, H.; Pauly, K.; Troupin, C.; Mineur, F.; de Clerck, O. Bio-ORACLE: A global environmental dataset for marine species distribution modelling. Glob. Ecol. Biogeogr. 2012, 21, 272–281. [Google Scholar] [CrossRef]
- Pebesma, E. Simple Features for R: Standardized Support for Spatial Vector Data. R J. 2018, 10, 439–446. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. Package R Package Version 2018, 2, 5–7. Available online: https://CRAN.R-project.org/package=vegan (accessed on 4 November 2021).
- Suzuki, R.; Terada, Y.; Shimodaira, H. Pvclust: Hierarchical Clustering with p-Values via Multiscale Bootstrap. 2019. Available online: https://CRAN.R-project.org/package=pvclust (accessed on 4 November 2021).
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix. 2021. Available online: https://github.com/taiyun/corrplot (accessed on 4 November 2021).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis. 2016. Available online: https://ggplot2.tidyverse.org (accessed on 4 November 2021).
- Costello, M.J.; Chaudhary, C. Marine Biodiversity, Biogeography, Deep-Sea Gradients, and Conservation. Curr. Biol. 2017, 27, R511–R527. [Google Scholar] [CrossRef]
- Brandt, A.; Linse, K.; Schüller, M. Bathymetric distribution patterns of Southern Ocean macrofaunal taxa: Bivalvia, Gastropoda, Isopoda and Polychaeta. Deep Sea Res. Part I Oceanogr. Res. Pap. 2009, 56, 2013–2025. [Google Scholar] [CrossRef]
- Pamungkas, J.; Glasby, C.; Costello, M. Biogeography of polychaete worms (Annelida) of the world. Mar. Ecol. Prog. Ser. 2021, 657, 147–159. [Google Scholar] [CrossRef]
- Moreno, R.A.; Labra, F.A.; Cotoras, D.D.; Camus, P.A.; Gutiérrez, D.; Aguirre, L.; Rozbaczylo, N.; Poulin, E.; Lagos, N.A.; Zamorano, D.; et al. Evolutionary drivers of the hump-shaped latitudinal gradient of benthic polychaete species richness along the Southeastern Pacific coast. PeerJ 2021, 9, e12010. [Google Scholar] [CrossRef]
- Giangrande, A.; Licciano, M. Factors influencing latitudinal pattern of biodiversity: An example using Sabellidae (Annelida, Polychaeta). Biodivers. Conserv. 2004, 13, 1633–1646. [Google Scholar] [CrossRef]
- Powell, M.G.; Beresford, V.P.; Colaianne, B.A. The latitudinal position of peak marine diversity in living and fossil biotas. J. Biogeogr. 2012, 39, 1687–1694. [Google Scholar] [CrossRef]
- Yasuhara, M.; Wei, C.-L.; Kucera, M.; Costello, M.J.; Tittensor, D.P.; Kiessling, W.; Bonebrake, T.C.; Tabor, C.R.; Feng, R.; Baselga, A.; et al. Past and future decline of tropical pelagic biodiversity. Proc. Natl. Acad. Sci. USA 2020, 117, 12891–12896. [Google Scholar] [CrossRef]
- Molinos, J.G.; Halpern, B.S.; Schoeman, D.S.; Brown, C.J.; Kiessling, W.; Moore, P.J.; Pandolfi, J.M.; Poloczanska, E.S.; Richardson, A.J.; Burrows, M.T. Climate velocity and the future global redistribution of marine biodiversity. Nat. Clim. Chang. 2016, 6, 83–88. [Google Scholar] [CrossRef]
- Saiz, J.I.; Bustamante, M.; Tajadura, J. A census of deep-water sipunculans (Sipuncula). Mar. Biodivers. 2018, 48, 449–464. [Google Scholar] [CrossRef]
- Khim, J.S.; Lee, C.; Song, S.J.; Bae, H.; Noh, J.; Lee, J.; Kim, H.-G.; Choi, J.-W. Marine biodiversity in Korea: A review of macrozoobenthic assemblages, their distributions, and long-term community changes from human impacts. Oceanogr. Mar. Biol. Annu. Rev. 2021, 59, 483–532. [Google Scholar] [CrossRef]
- Shin, P.K. Biodiversity of subtidal benthic polychaetes in Hong Kong coastal waters. In The Marine Biology of the South China Sea. In Proceedings of the Third International Conference on the Marine Biology of Hong Kong and the South China Sea, Hong Kong, China, 28 October–1 November 1998; Volume 28, pp. 57–74. [Google Scholar]
- Sun, Y.; ten Hove, H.A.; Qiu, J.-W. Serpulidae (Annelida: Polychaeta) from Hong Kong. Zootaxa 2012, 3424, 1–42. [Google Scholar] [CrossRef] [Green Version]
- Golovan, O.A.; Błażewicz, M.; Brandt, A.; Jażdżewska, A.; Jóźwiak, P.; Lavrenteva, A.V.; Malyutina, M.V.; Petryashov, V.V.; Riehl, T.; Sattarova, V.V. Diversity and distribution of peracarid crustaceans (Malacostraca) from the abyss adjacent to the Kuril-Kamchatka Trench. Mar. Biodivers. 2019, 49, 1343–1360. [Google Scholar] [CrossRef]
- Murina, G.V. Geographical distribution of marine worms of the phylum Sipuncula in the world ocean. Zool. Zh. 1971, 50, 184–192. [Google Scholar]
- Murina, G.V. Novye taksony morskikh chervei sipunkulid roda Golfingia. Zool. Zh. 1975, 54, 1085–1089. [Google Scholar]
- Murina, G.V. Morskie chervy sipunkulidy arkticheskikh i boreal’nykh vod Evrazii. Oprediteli Po Faune SSSR 1977, 111. Available online: https://agris.fao.org/agris-search/search.do?recordID=US201300379732 (accessed on 4 November 2021).
- Maiorova, A.S.; Adrianov, A.V. Deep-sea sipunculans (Sipuncula) of the northwestern Pacific. Russ. J. Mar. Biol. 2017, 43, 181–189. [Google Scholar] [CrossRef]
- Chaudhary, C.; Saeedi, H.; Costello, M.J. Marine Species Richness Is Bimodal with Latitude: A Reply to Fernandez and Marques. Trends Ecol. Evol. 2017, 32, 234–237. [Google Scholar] [CrossRef]
- Angel, M.; Gage, J.; Tyler, P. Deep-Sea Biology: A Natural History of Organisms at the Deep-Sea Floor. J. Anim. Ecol. 1991, 61, 233. [Google Scholar] [CrossRef]
- Fernández-Romero, A.; Moreira, J.; Guerra-García, J.M. Marinas: An overlooked habitat for exploring the relation among polychaete assemblages and environmental factors. Mar. Pollut. Bull. 2019, 138, 584–597. [Google Scholar] [CrossRef]
- Dafforn, K.A.; Kelaher, B.P.; Simpson, S.L.; Coleman, M.A.; Hutchings, P.A.; Clark, G.F.; Knott, N.A.; Doblin, M.A.; Johnston, E.L. Polychaete Richness and Abundance Enhanced in Anthropogenically Modified Estuaries Despite High Concentrations of Toxic Contaminants. PLoS ONE 2013, 8, e77018. [Google Scholar] [CrossRef] [Green Version]
- Arvanitidis, C.; Koutsoubas, D.; Dounas, C.; Eleftheriou, A. Annelid fauna of a Mediterranean lagoon (Gialova Lagoon, south-west Greece): Community structure in a severely fluctuating environment. J. Mar. Biol. Assoc. United Kingd. 1999, 79, 849–856. [Google Scholar] [CrossRef]
- Campanyà-Llovet, N.; Snelgrove, P.V.; Parrish, C.C. Rethinking the importance of food quality in marine benthic food webs. Prog. Oceanogr. 2017, 156, 240–251. [Google Scholar] [CrossRef]
- Boyd, P.W.; Jickells, T.; Law, C.S.; Blain, S.; Boyle, E.A.; Buesseler, K.O.; Coale, K.H.; Cullen, J.J.; de Baar, H.J.W.; Follows, M.; et al. Mesoscale Iron Enrichment Experiments 1993-2005: Synthesis and Future Directions. Science 2007, 315, 612–617. [Google Scholar] [CrossRef] [Green Version]
- De Baar, H.J.W.; de Jong, J.; Bakker, D.; Löscher, B.M.; Veth, C.; Bathmann, U.; Smetacek, V. Importance of iron for plankton blooms and carbon dioxide drawdown in the Southern Ocean. Nature 1995, 373, 412–415. [Google Scholar] [CrossRef]
- Glover, A.G. Abyssal Polychaete Assemblages along Latitudinal Gradients of Productivity in the Equatorial Pacific and North Atlantic Oceans. Ph.D. Thesis, University of Southampton, Southampton, UK, 2000. [Google Scholar]
- Neal, L.; Hardy, S.L.M.; Smith, C.R.; Glover, A.G. Polychaete species diversity on the West Antarctic Peninsula deep continental shelf. Mar. Ecol. Prog. Ser. 2011, 428, 119–134. [Google Scholar] [CrossRef] [Green Version]
- Ambrose, W.G., Jr.; Renaud, P.E.; Cochrane, S.K.J.; Denisenko, S.G.; Skardhamar, J. Polychaete diveristy patterns on two Artic shelves: Impacts of ice and primary production? Zoosymposia 2009, 2, 457–485. [Google Scholar] [CrossRef] [Green Version]
- Rogers, A.D. The role of oceanic oxygen minima in generating biodiversity in the deep sea. Deep Sea Reserach II Top. Stud. Oceanogr. 2000, 47, 119–148. [Google Scholar] [CrossRef]
- Jayaraj, K.A.; Jayalakshmi, K.V.; Saraladevi, K. Influence of environmental properties on macrobenthos in the northwest Indian shelf. Environ. Monit. Assess. 2007, 127, 459–475. [Google Scholar] [CrossRef] [Green Version]
- Llanso, R.J.; Diaz, R.J. Tolerance to low dissolved oxygen by the tubicolous polychaete Loimia medusa. J. Mar. Biol. Assoc. United Kingd. 1994, 74, 143–148. [Google Scholar] [CrossRef]
- Lamont, P.A.; Gage, J.D. Morphological responses of macrobenthic polychaetes to low oxygen on the Oman continental slope, NW Arabian Sea. Deep Sea Res. Part II 2000, 47, 9–24. [Google Scholar] [CrossRef]
- Levin, L.A.; Etter, R.J.; Rex, M.A.; Gooday, A.J.; Smith, C.R.; Pineda, J.; Stuart, C.T.; Hessler, R.R.; Pawson, D. Environmental Influences on Regional Deep-Sea Species Diversity. Annu. Rev. Ecol. Syst. 2001, 32, 51–93. [Google Scholar] [CrossRef] [Green Version]
- Muniz, P.; Pires, A.M.S. Polychaete associations in a subtropical environment (São Sebastião Channel, Brazil): A structural analysis. Mar. Ecol. 2000, 21, 145–160. [Google Scholar] [CrossRef]
- Musale, A.S.; Desai, D.V. Distribution and abundance of macrobenthic polychaetes along the South Indian coast. Environ. Monit. Assess. 2011, 178, 423–436. [Google Scholar] [CrossRef]
- Halpin, P.N.; Read, A.J.; Fujioka, E.; Best, B.D.; Donnely, B.; Hazen, L.J.; Kot, C.; Urian, K.; LaBrecque, E.; Dimatteo, A.; et al. OBIS-SEAMAP: The world data center for marine mammal, sea bird, and sea turtle distributions. Oceanography 2009, 22, 104–115. Available online: http://www.jstor.org/stable/24860963 (accessed on 4 November 2021). [CrossRef]
- Bosch, S.; Tyberghein, L.; Deneudt, K.; Hernandez, F.; de Clerck, O. In search of relevant predictors for marine species distribution modelling using the MarineSpeed benchmark dataset. Divers. Distrib. 2018, 24, 144–157. [Google Scholar] [CrossRef] [Green Version]
- Coro, G.; Magliozzi, C.; Berghe, E.V.; Bailly, N.; Ellenbroek, A.; Pagano, P. Estimating absence locations of marine species from data of scientific surveys in OBIS. Ecol. Model. 2016, 323, 61–76. [Google Scholar] [CrossRef]
- Ready, J.; Kaschner, K.; South, A.B.; Eastwood, P.D.; Rees, T.; Rius, J.; Agbayani, E.; Kullander, S.; Froese, R. Predicting the distributions of marine organisms at the global scale. Ecol. Model. 2010, 221, 467–478. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kohlenbach, K.; Knauber, H.; Brandt, A.; Saeedi, H. Distribution and Species Richness of Benthic Polychaeta and Sipuncula in the Northwestern Pacific. Diversity 2023, 15, 557. https://doi.org/10.3390/d15040557
Kohlenbach K, Knauber H, Brandt A, Saeedi H. Distribution and Species Richness of Benthic Polychaeta and Sipuncula in the Northwestern Pacific. Diversity. 2023; 15(4):557. https://doi.org/10.3390/d15040557
Chicago/Turabian StyleKohlenbach, Katharina, Henry Knauber, Angelika Brandt, and Hanieh Saeedi. 2023. "Distribution and Species Richness of Benthic Polychaeta and Sipuncula in the Northwestern Pacific" Diversity 15, no. 4: 557. https://doi.org/10.3390/d15040557
APA StyleKohlenbach, K., Knauber, H., Brandt, A., & Saeedi, H. (2023). Distribution and Species Richness of Benthic Polychaeta and Sipuncula in the Northwestern Pacific. Diversity, 15(4), 557. https://doi.org/10.3390/d15040557