

Effects of Bacillus cereus on Survival, Fecundity, and Host Adaptability of Pine Wood Nematode

Abstract
:1. Introduction
2. Materials and Methods
2.1. PWN Strains, Bacterial Strains, and Hosts
2.2. Pretreatment of Nematodes and Bacteria
2.3. Viability of Nematodes with Contrasting Virulence under Different Bacterial Treatments
2.4. Fecundity of Different Virulent Nematodes Treated with Different Concentrations of Bacteria
2.5. Inoculation of P. massoniana and L. kaempferi with Mixed Inoculation of Bacteria and Nematodes
2.6. RNA Isolation, cDNA Synthesis, and RNA Sequence
2.7. Quantitative Real-Time PCR
2.8. Sequencing Data Analysis
2.9. Statistical Analysis
3. Results
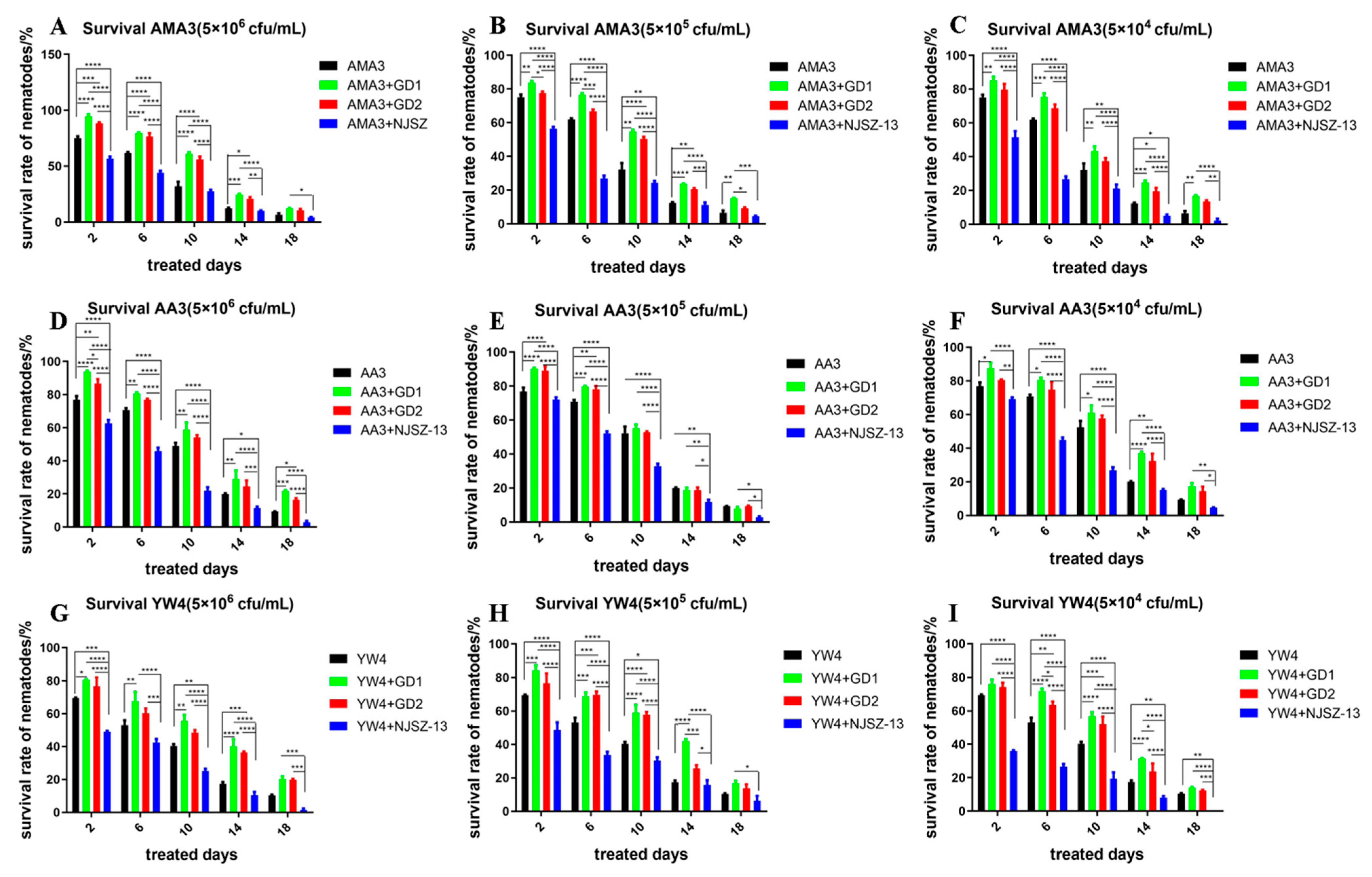
3.1. Survival of Nematodes with Different Virulence under Different Bacterial Treatments
3.2. Fecundity of Different Virulent Nematodes Treated with Different Concentrations of Bacteria
3.3. Mixed Inoculation of Bacteria and Nematodes to P. massoniana and L. kaempferi
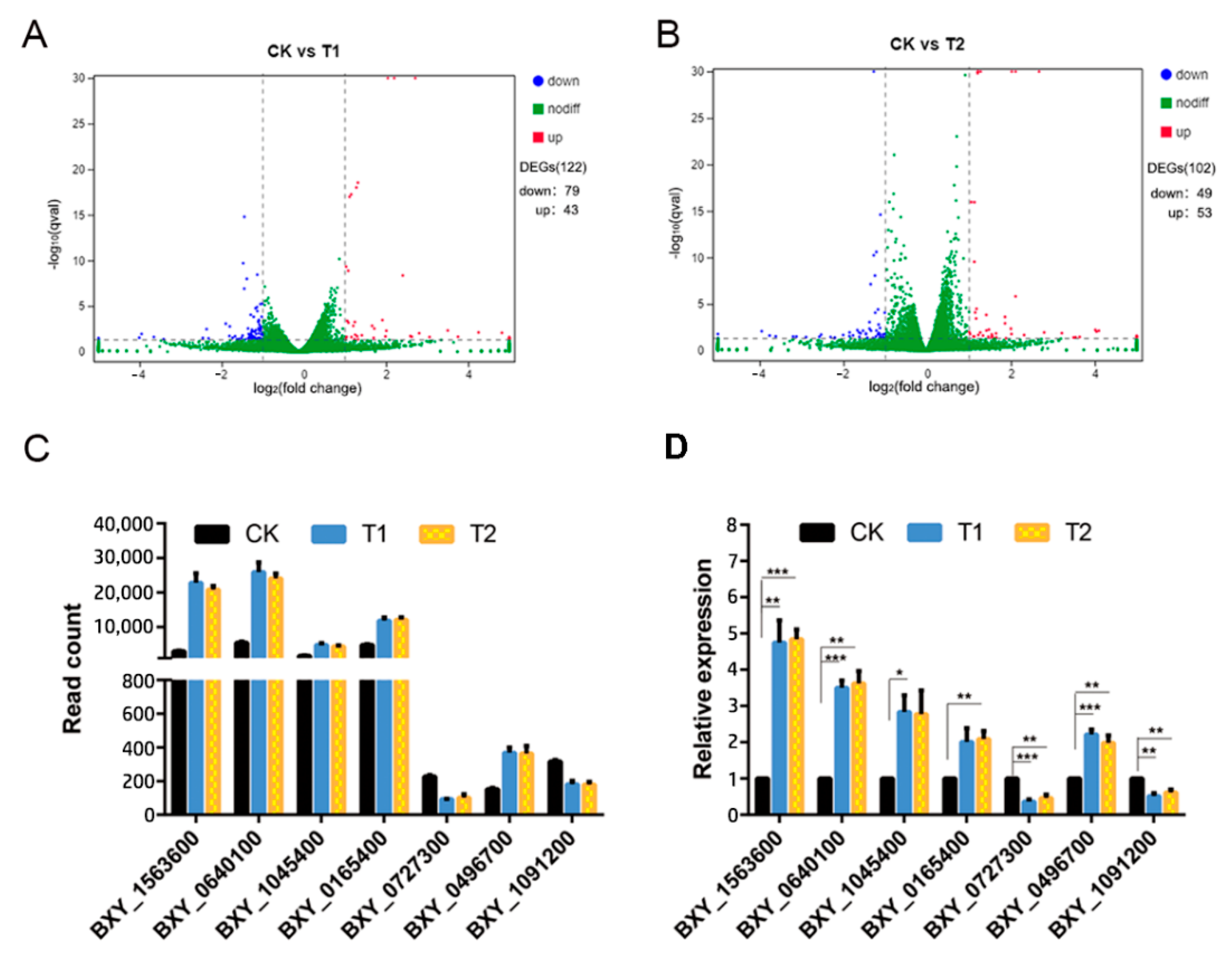
3.4. Differential Gene Expression Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, J.; Yao, H. Current situation and management countermeasures of pine wilt disease in China. For. Pest Dis. 2019, 38, 45–46. [Google Scholar]
- Tóth, Á. Bursaphelenchus xylophilus, the pinewood nematode: Its significance and a historical review. Acta Biol. Szeged. 2011, 55, 213–217. [Google Scholar]
- Dwinell, L. First report of pinewood nematode (Bursaphelenchus xylophilus) in Mexico. Plant Dis. 1993, 77, 846. [Google Scholar] [CrossRef]
- Ryss, A.Y.; Kulinich, O.A.; Sutherland, J.R. Pine wilt disease: A short review of worldwide research. For. Stud. China 2011, 13, 132–138. [Google Scholar] [CrossRef]
- Zhao, B.; Kazuyoshi, F.; Sutherland, J.; Takeuchi, Y. Pine Wilt Disease; Springer: Tokyo, Japan, 2008. [Google Scholar]
- Fonseca, L.; Cardoso, J.; Lopes, A.; Pestana, M.; Abreu, F.; Nunes, N.; Mota, M.; Abrantes, I. The pinewood nematode, Bursaphelenchus xylophilus, in Madeira Island. Helminthologia 2012, 49, 96–103. [Google Scholar] [CrossRef]
- Robertson, L.; Cobacho Arcos, S.; Escuer, M.; Santiago Merino, R.; Esparrago, G.; Abelleira, A.; Navas, A. Incidence of the pinewood nematode Bursaphelenchus xylophlius Steiner & Buhrer, 1934 (Nickle, 1970) in Spainv. Nematology 2011, 13, 755–757. [Google Scholar]
- Ye, J. Epidemic status of pine wilt disease in China and its prevention and control techniques and counter measures. Sci. Silvae Sin. 2019, 55, 1–10. [Google Scholar]
- Yan, Z.; Tan, J. Species and geographical distribution of parasitic nematode in pine in China. For. Pest Dis. 2019, 5, 38–42. [Google Scholar]
- Yu, H.; Wu, H.; Huang, R.; Wang, J.; Zhang, R.; Song, Y. Investigation, separation and identification of Bursaphelenchus xylophilus from Pinus sylvestris var. mongolica in Fushun city. For. Pest Dis. 2019, 39, 6–10. [Google Scholar]
- Yu, H.; Wu, H.; Zhang, X. Preliminary study on Larix spp. infected by Bursaphelenchus xylophilus in natural environment. For. Pest Dis. 2019, 38, 7–10. [Google Scholar]
- Oku, H.; Shiraishi, T.; Ouchi, S.; Kurozumi, S.; Ohta, H. Pine wilt toxin, the metabolite of a bacterium associated with a nematode. Naturwissenschaften 1980, 67, 198–199. [Google Scholar] [CrossRef]
- Kawazu, K.; Yamashita, H.; Kobayashi, A.; Kanzaki, H. Isolation of pine-wilting bacteria accompanying pine wood nematode, Bursaphelenchus xylophilus, and their toxic metabolites. Sci. Rep. Fac. Agric.-Okayama Univ. 1998, 87, 1–7. [Google Scholar]
- Zhao, B.; Wang, H.; Han, S.; Han, Z. Distribution and pathogenicity of bacteria species carried by Bursaphelenchus xylophilus in China. Nematology 2003, 5, 899–906. [Google Scholar] [CrossRef]
- Xie, L.; Zhao, B. Post-inoculation population dynamics of Bursaphelenchus xylophilus and associated bacteria in pine wilt disease on Pinus thunbergii. J. Phytopathol. 2008, 156, 385–389. [Google Scholar] [CrossRef]
- Tian, X.; Zhang, Q.; Chen, G.; Mao, Z.; Yang, J.; Xie, B. Diversity of bacteria associated with pine wood nematode revealed by metagenome. Acta Phytopathol. Sin. 2010, 50, 909–916. [Google Scholar]
- Vicente, C.S.L.; Nascimento, F.; Espada, M.; Barbosa, P.; Mota, M.; Glick, B.R.; Oliveira, S. Characterization of bacteria associated with pinewood nematode Bursaphelenchus xylophilus. PLoS ONE 2012, 7, e46661. [Google Scholar] [CrossRef]
- Kikuchi, T.; Cotton, J.A.; Dalzell, J.J.; Hasegawa, K.; Kanzaki, N.; Mcveigh, P.; Takanashi, T.; Tsai, I.J.; Assefa, S.A.; Cock, P.J.A.; et al. Genomic insights into the origin of parasitism in the emerging plant pathogen Bursaphelenchus xylophilus. PLoS Pathog. 2011, 7, e1002219. [Google Scholar] [CrossRef]
- Nascimento, F.; Vicente, C.; Cock, P.; Tavares, M.; Mota, M. From plants to nematodes: Serratia grimesii BXF1 genome reveals an adaptation to the modulation of multi-species interac-tions. Microbial Genomics 2018, 4, e000178. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, T.; Pan, Z.; Lin, L.; Dong, G.; Wang, M.; Li, R. The alcohol dehydrogenase with a broad range of substrate specificity regulates vitality and reproduction of the plant-parasitic nematode Bursaphelenchus xylophilus. Parasitology 2019, 146, 497–505. [Google Scholar] [CrossRef]
- Yan, Z.; Zhai, N.; Tan, J.; Hao, D. Effects of three strains of Bacillus spp. On the survival and reproduction of pine wood nematode Bursaphelenchus xylophilus at low-temperature. J. Plant Prot. 2021, 48, 442–448. [Google Scholar]
- Ding, X.; Ye, J.; Lin, S.; Wu, X.; Bo, N. Deciphering the Molecular Variations of Pine Wood Nematode Bursaphelenchus xylophilus with Different Virulence. PLoS ONE 2016, 11, e0156040. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wu, F.; Wu, X.; Ye, J. Differential effects of rapamycin on Bursaphelenchus xylophilus with different virulence and differential expression of autophagy genes under stresses in nematodes. Acta Biochim. Biophys. Sin. 2019, 51, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Yuan, W.; Tian, X.; Fan, B.; Fang, X.; Ye, J.; Ding, X. Specific and functional diversity of endophytic bacteria from pine wood nematode Bursaphelenchus Xylophilus with different virulence. Int. J. Biol. Sci. 2013, 9, 34–44. [Google Scholar] [CrossRef]
- Zhu, L.; Ye, J.; Negi, S.; Xu, X.; Wang, Z.; Ji, J. Pathogenicity of aseptic Bursaphelenchus xylophilus. PLoS ONE 2012, 7, e38095. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Tan, J.; Chen, F.; Hao, D. Colonization of Bacillus cereus NJSZ-13, a species with nematicidal activity in Masson pine (Pinus massoniana Lamb.). J. For. Res. 2020, 31, 1025–1033. [Google Scholar] [CrossRef]
- Tan, J.; Feng, Z. Effect of two bacteria strains on the survival and reproduction of Bursaphelenchus xylophilus. Acta Phytopathol. Sin. 2003, 6, 557–558. [Google Scholar]
- Mamiya, Y. Pathology of the Pine Wilt Disease Caused by Bursaphelenchus xylophilus. Annu. Rev. Phytopathol. 1983, 21, 201–220. [Google Scholar] [CrossRef]
- Fang, Z. Research Methods of Plant Disease; China Agriculture Press: Beijing, China, 1998. [Google Scholar]
- Kawazu, K.; Nishii, Y.; Ishii, K.; Tada, M. A convenient screening method for nematicidal activity. Agric. Biol. Chem. 1980, 44, 631–635. [Google Scholar]
- Yu, L.; Wu, X.; Ye, J.; Zhang, S.; Wang, C. NOS-like-mediated nitric oxide is involved in Pinus thunbergii response to the invasion of Bursaphelenchus xylophilus. Plant Cell Rep. 2012, 31, 1813–1821. [Google Scholar] [CrossRef]
- Proenca, D.N.; Grass, G.; Morais, P.V. Understanding pine wilt disease: Roles of the pine endophytic bacteria and of the bacteria carried by the disease-causing pinewood nematode. MicrobiologyOpen 2017, 6, e00415. [Google Scholar] [CrossRef]
- Proenca, D.N.; Fonseca, L.; Powers, T.O.; Abrantes, I.M.O.; Morais, P.V. Diversity of bacteria carried by pinewood nematode in USA and phylogenetic comparison with isolates from other countries. PLoS ONE 2014, 9, e105190. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.; Moens, M.; Mota, M.; Li, H.; Kikuchi, T. Bursaphelenchus xylophilus: Opportunities in comparative genomics and molecular host-parasite interactions. Mol. Plant Pathol. 2008, 9, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Kazuyoshi, F. Pine Wood Nematode, Bursaphelenchus xylophilus. Annu. Rev. Phytopathol. 2013, 51, 61–83. [Google Scholar]
- Zhao, B.; Lin, F. Mutualistic symbiosis between Bursaphelenchus xylophilus and bacteria of the genus Pseudomonas. For. Pathol 2005, 35, 339–345. [Google Scholar] [CrossRef]
- Vicente, C.S.; Ikuyo, Y.; Mota, M.; Hasegawa, K. Pinewood nematode-associated bacteria contribute to oxidative stress resistance of Bursaphelenchus xylophilus. BMC Microbiol. 2013, 13, 299. [Google Scholar] [CrossRef]
- Chi, S.; He, Y.; Han, Z.; Zhang, Y. Studies on complex infecting of pine wood nematodes and the bacterium carried by it. J. Fujian Coll. For. 2008, 28, 92–96. [Google Scholar]
- Kotiranta, A.; Lounatmaa, K.; Haapasalo, M. Epidemiology and pathogenesis of Bacillus cereus infections. Microbes Infect. 2000, 2, 189–198. [Google Scholar] [CrossRef]
- Lin, C.; Zhang, X.; Xiao, J.; Zhong, Q.; Kuang, Y. Effects on longevity extension and mechanism of action of carnosic acid in Caenorhabditis elegans. Food Funct. 2019, 10, 1398–1410. [Google Scholar] [CrossRef]
- Treinin, M.; Shliar, J.; Jiang, H.; Powell-Coffman, J.A.; Bromberg, Z.; Horowitz, M. HIF-1 is required for heat acclimation in the nematode Caenorhabditis elegans. Physiol. Genom. 2003, 14, 17–24. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disease Grade | Grading Standard | Representative Value |
|---|---|---|
| I | Healthy, normal growth of plants | 0 |
| II | A few needles turned yellow | 1 |
| III | Half of the needles turned yellow and the branches were bent | 2 |
| IV | Most of the needles turned yellow, tree became wilted | 3 |
| V | All the needles turned yellow, tree wilted | 4 |
| ID | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|
| BXY_1563600 | AAGGGCCGTCTCTCACAAAG | TCTCTGCCGTCTGGTTGTTC |
| BXY_0640100 | TCAATGGGTGGAGAGCAACC | CAGTAGGTCCACTGGCTTGG |
| BXY_1045400 | TGGAGGCAATTCAGGCTCAG | ACTCGGAGCCCAACGAATTT |
| BXY_0165400 | ACCGACACATCAGGATTCCG | GGGCTTCACTTGAATGGGGA |
| BXY_0727300 | CCCACAATGTCGCCAATCCT | CCACATCAGCGGGAAGGAAA |
| BXY_0496700 | GCCTTTCGCTGGAAGACCC | AACCCTCGTCGCACTGTCG |
| BXY_1091200 | CCGTGCCTGCTCATCATTCT | ATCCCGACCTGCTTACAACG |
| EU100952 | GCAACACGGAGTTCGTTGTA | GTATCGTCACCAACTGGGAT |
| Days after Inoculation/d. | AMA3 | AMA3 + GD1 | AMA3 + GD2 | GD1 | GD2 | CK | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Disease Incidence/% | DI | Disease Incidence/% | DI | Disease Incidence/% | DI | Disease Incidence/% | DI | Disease Incidence/% | DI | Disease Incidence/% | DI | |
| 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | N/A | 0 | N/A | 0 | N/A |
| 10 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | N/A | 0 | N/A | 0 | N/A |
| 15 | 10 | 2.5 | 10 | 2.5 | 20 | 10 | 0 | N/A | 0 | N/A | 0 | N/A |
| 20 | 10 | 7.5 | 10 | 10 | 30 | 22.5 | 0 | N/A | 0 | N/A | 0 | N/A |
| 25 | 20 | 20 | 40 | 35 | 40 | 35 | 0 | N/A | 0 | N/A | 0 | N/A |
| 30 | 20 | 20 | 40 | 35 | 50 | 37.5 | 0 | N/A | 0 | N/A | 0 | N/A |
| 35 | 20 | 20 | 40 | 40 | 70 | 67.5 | 0 | N/A | 0 | N/A | 0 | N/A |
| 40 | 40 | 35 | 40 | 40 | 70 | 70 | 0 | N/A | 0 | N/A | 0 | N/A |
| Days after Inoculation/d | AMA3 | AMA3 + GD1 | AMA3 + GD2 | GD1 | GD2 | CK | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Disease Incidence/% | DI | Disease Incidence/% | DI | Disease Incidence/% | DI | Disease Incidence/% | DI | Disease Incidence/% | DI | Disease Incidence/% | DI | |
| 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | N/A | 0 | N/A | 0 | N/A |
| 10 | 40 | 10 | 40 | 10 | 40 | 40 | 0 | N/A | 0 | N/A | 0 | N/A |
| 15 | 60 | 15 | 40 | 35 | 60 | 45 | 0 | N/A | 0 | N/A | 0 | N/A |
| 20 | 60 | 15 | 40 | 35 | 60 | 60 | 0 | N/A | 0 | N/A | 0 | N/A |
| 25 | 60 | 25 | 40 | 40 | 60 | 60 | 0 | N/A | 0 | N/A | 0 | N/A |
| 30 | 60 | 45 | 40 | 45 | 80 | 65 | 0 | N/A | 0 | N/A | 0 | N/A |
| 35 | 60 | 50 | 60 | 60 | 80 | 70 | 0 | N/A | 0 | N/A | 0 | N/A |
| 40 | 60 | 60 | 60 | 60 | 80 | 70 | 0 | N/A | 0 | N/A | 0 | N/A |
| 45 | 60 | 60 | 60 | 60 | 100 | 85 | 0 | N/A | 0 | N/A | 0 | N/A |
| ID | Gene Name | KEGG-ko | Pathway |
|---|---|---|---|
| BXY_1563600 | Hsp 72 | ko04010 | MAPK signaling |
| BXY_0640100 | Hsp 70 | ko04010 | MAPK signaling |
| BXY_0165400 | sHsp 21 | ko04212 | Longevity-regulating pathway worm |
| BXY_0727300 | DI09 29p80 | ko00982 | Drug-metabolism cytochrome P450 |
| BXY_0496700 | osta-1 | ko04976 | Bile secretion |
| BXY_1091200 | CBG 01395 | ko04976 | Bile secretion |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, Y.; Yan, Z.; Chen, Y.; Ye, J.; Tan, J. Effects of Bacillus cereus on Survival, Fecundity, and Host Adaptability of Pine Wood Nematode. Diversity 2023, 15, 566. https://doi.org/10.3390/d15040566
Yuan Y, Yan Z, Chen Y, Ye J, Tan J. Effects of Bacillus cereus on Survival, Fecundity, and Host Adaptability of Pine Wood Nematode. Diversity. 2023; 15(4):566. https://doi.org/10.3390/d15040566
Chicago/Turabian StyleYuan, Yuchao, Zhengmei Yan, Yangxue Chen, Jianren Ye, and Jiajin Tan. 2023. "Effects of Bacillus cereus on Survival, Fecundity, and Host Adaptability of Pine Wood Nematode" Diversity 15, no. 4: 566. https://doi.org/10.3390/d15040566
APA StyleYuan, Y., Yan, Z., Chen, Y., Ye, J., & Tan, J. (2023). Effects of Bacillus cereus on Survival, Fecundity, and Host Adaptability of Pine Wood Nematode. Diversity, 15(4), 566. https://doi.org/10.3390/d15040566