
Phylogenomics of One of the World’s Most Intriguing Groups of CAM Plants, the Opuntioids (Opuntioideae: Cactaceae): Adaptation to Tropical Dry Forests Helped Drive Prominent Morphological Features in the Clade

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Taxon Sampling, DNA Extraction and Sequencing
2.2. Phylogenetic Analyses and Divergence Time Estimation
2.3. Morphological Evolution across Cactaceae
2.4. Ancestral Range Reconstruction
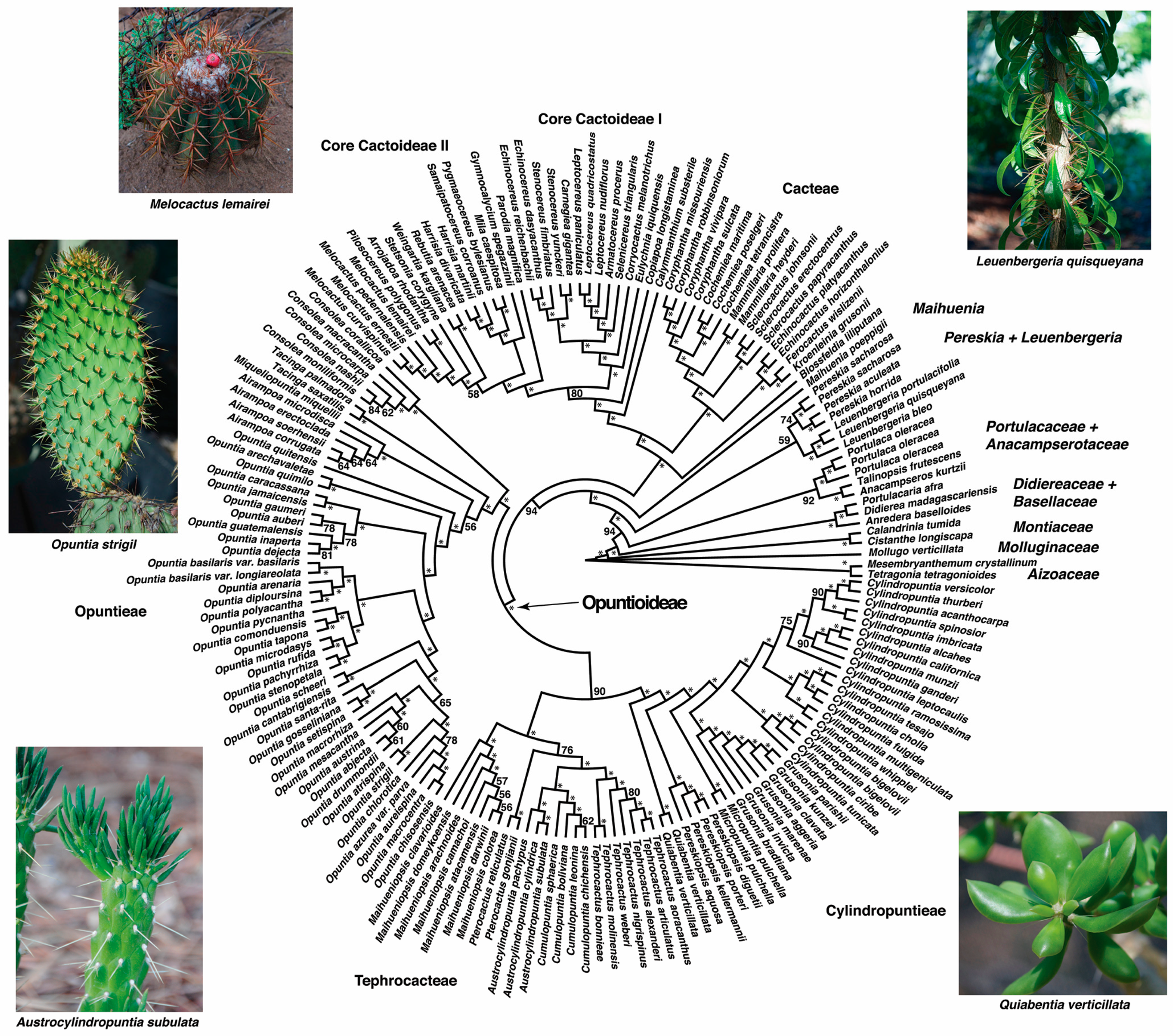
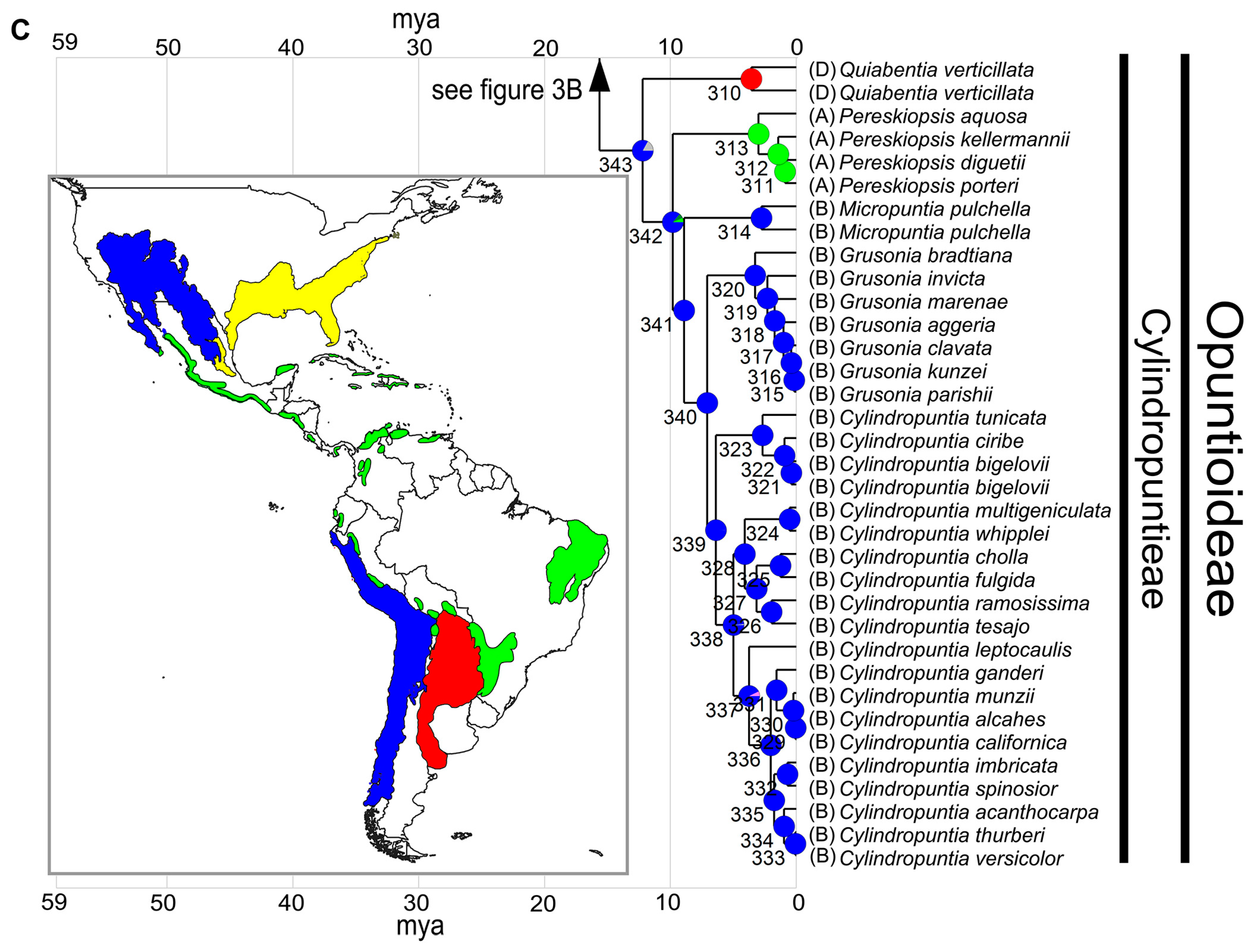
3. Results
3.1. Phylogeny and Divergence Time Estimation
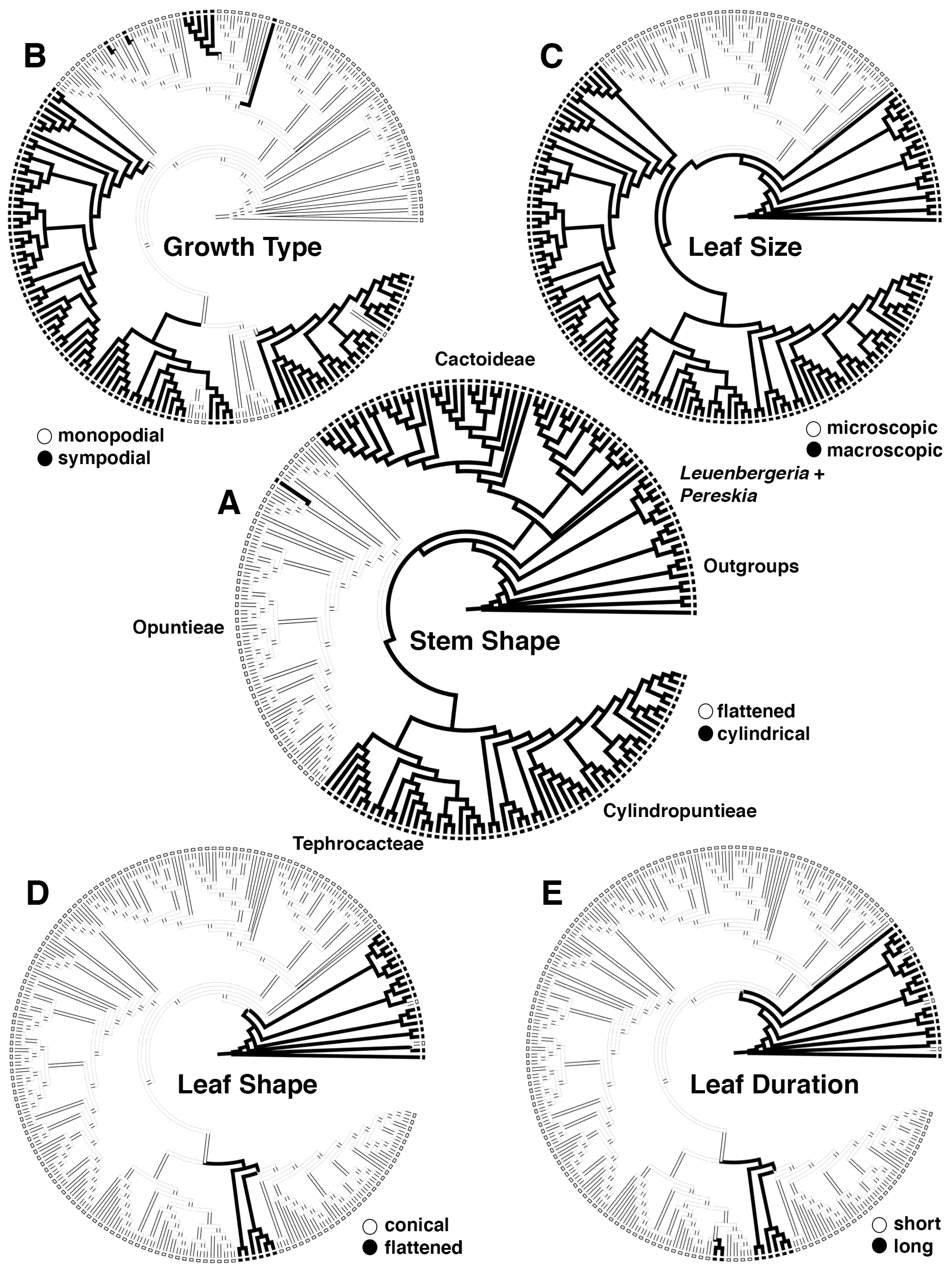
3.2. Morphological Evolution
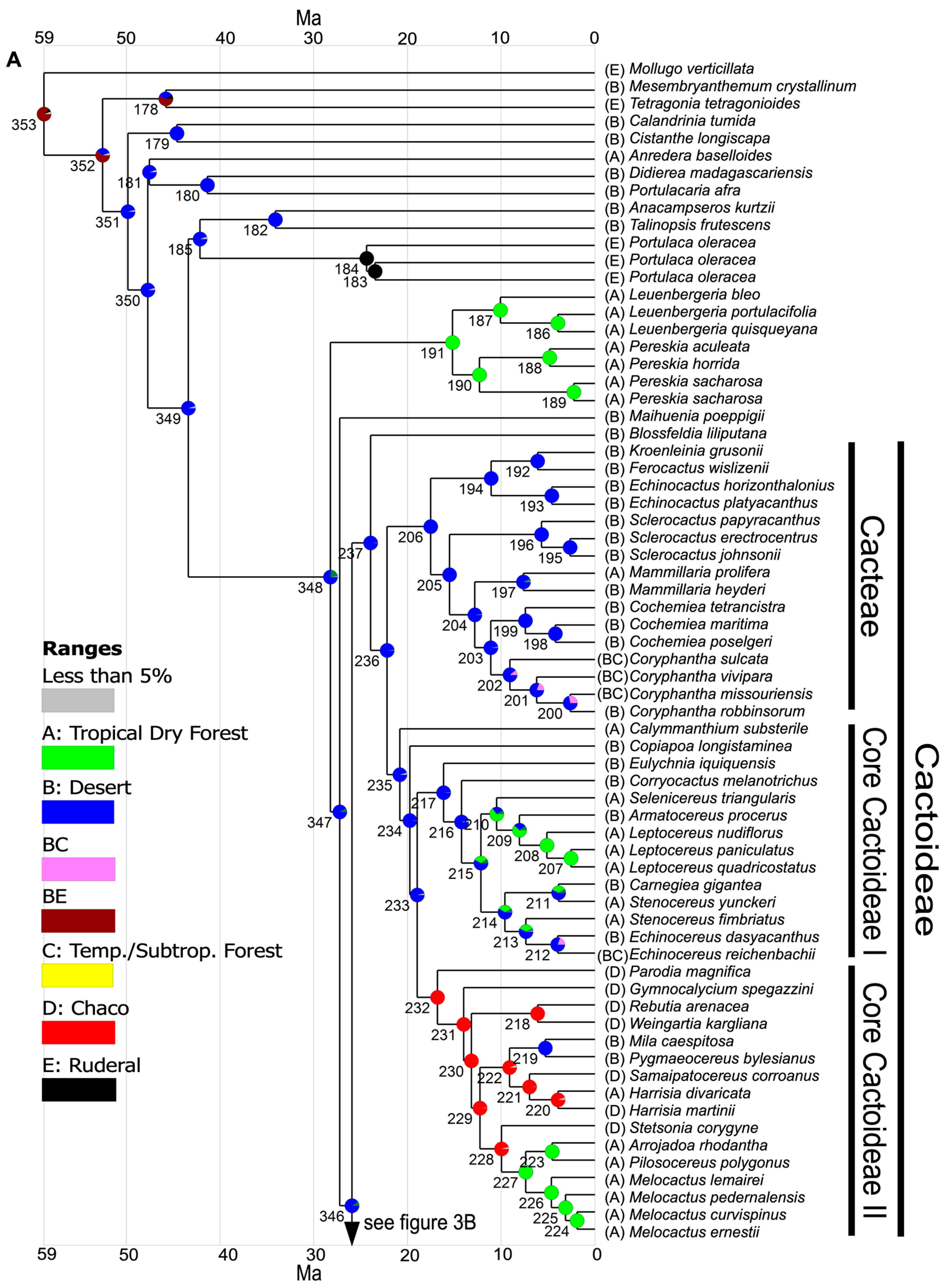
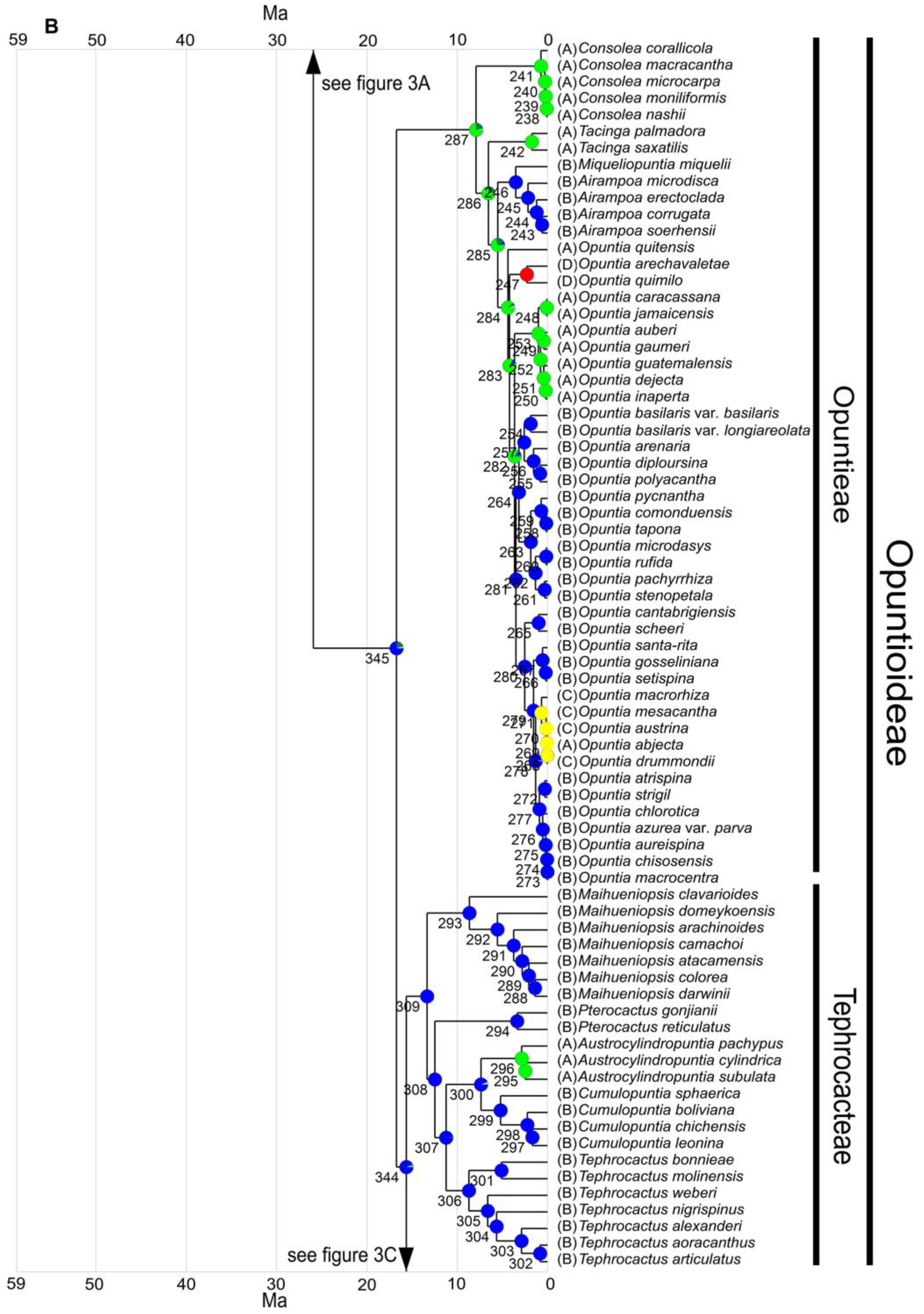
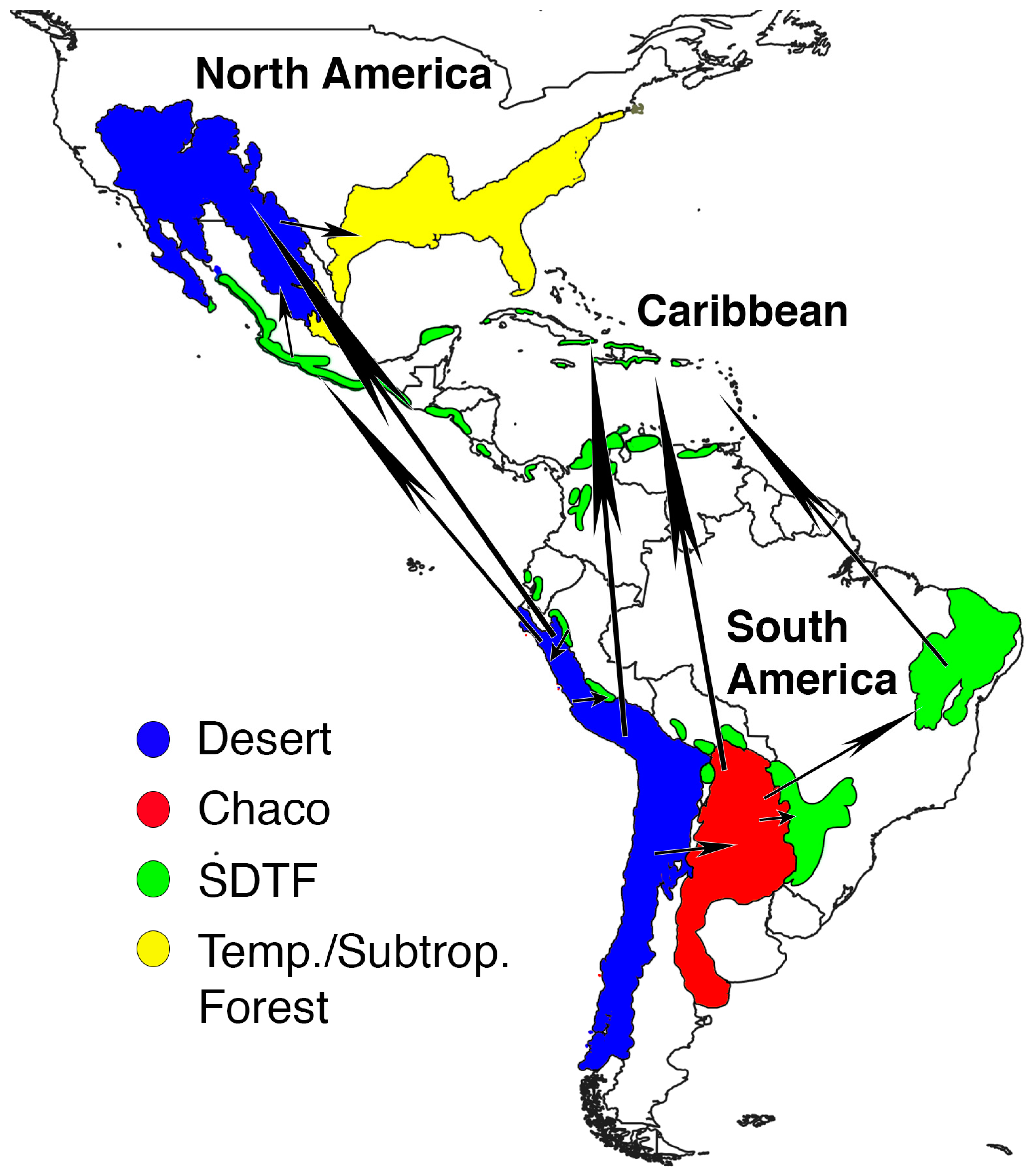
3.3. Ancestral Range Reconstruction
3.3.1. Range Defined Based on Habitat
3.3.2. Range Defined Based on Ecoregions
4. Discussion
4.1. Phylogenetic Relationships in Cactaceae
4.2. Morphological Evolution and Ancestral Habitats
4.3. Biogeography of Opuntioideae
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Britton, N.L.; Rose, J.N. The Cactaceae. Descriptions and Illustrations of Plants of the Cactus Family; Carnegie Institution of Washington D.C.: New York, NY, USA, 1919; Volume 1. [Google Scholar]
- Benson, L. The Cacti of the United States and Canada; Stanford University Press: Stanford, CA, USA, 1982. [Google Scholar]
- Anderson, E. The Cactus Family; Timber Press: Portland, OR, USA, 2001. [Google Scholar]
- Croizat, L. Manual of Phytogeography; Junk: Hague, The Netherlands, 1952. [Google Scholar]
- Buxbaum, F.; Kakteenleben. Eine Biologische Plauderei fur Jeden Naturfreund; A Philler Verlag: Minden, Germany, 1980. [Google Scholar]
- Barthlott, W. Biogeography and evolution in neo-and paleotropical Rhipsalinae. In Dispersal and Distribution; Kubitski, K., Ed.; Verlag Paul Parey: Hamburg, Germany, 1983; pp. 241–248. [Google Scholar]
- Backeberg, C. Cactaceae; Jahrbücher der DKG., Teil 2, Juni 1942: 55, Cactaceae Lindley; Systematische Übersicht (Neubearbeitung) mit Beschreibungsschlüssel. 1942. Available online: https://www.cactuspro.com/biblio_fichiers/pdf/CactJahr/CactJahr1941.pdf (accessed on 1 November 2022).
- Renner, S. Plant dispersal across the tropical Atlantic by wind and sea current. Int. J. Plant Sci. 2004, 165, S23–S33. [Google Scholar] [CrossRef]
- Cota-Sánchez, J.H.; Bomfim-Patrício, M.C. Seed morphology, polyploidy and the evolutionary history of the epiphytic cactus Rhipsalis baccifera (Cactaceae). Polibotánica 2010, 29, 107–129. [Google Scholar]
- Guerrero, P.; Majure, L.C.; Bustamante, E.; Hernández-Hernández, T. Phylogenetic relationships and evolutionary trends in the cactus family. J. Hered. 2018, 110, 4–21. [Google Scholar] [CrossRef] [PubMed]
- Ocampo, G.; Columbus, J.T. Molecular phylogenetics of suborder Cactineae (Caryophyllales), including insights into photosynthetic diversification and historical biogeography. Am. J. Bot. 2010, 97, 1827–1847. [Google Scholar] [CrossRef] [PubMed]
- Soltis, D.E.; Smith, S.A.; Cellinese, N.; Wurdack, K.J.; Tank, D.C.; Brockington, S.F.; Refulio-Rodriguez, N.F.; Walker, J.B.; Moore, M.J.; Carlsward, B.S.; et al. Angiosperm phylogeny: 17 genes, 640 taxa. Am. J. Bot 2011, 98, 704–730. [Google Scholar] [CrossRef]
- Brockington, S.F.; Yang, Y.; Gandia-Herrero, F.; Covshoff, S.; Hibberd, J.M.; Sage, R.F.; Wong, G.K.S.; Moore, M.J.; Smith, S.A. Lineage-specific gene radiations underlie the evolution of novel betalain pigmentation in Caryophyllales. New Phytol. 2015, 207, 1170–1180. [Google Scholar] [CrossRef]
- Yang, Y.; Moore, M.J.; Brockington, S.F.; Soltis, D.E.; Wong, G.K.S.; Carpenter, E.J.; Zhang, Y.; Chen, L.; Yan, Z.; Xie, Y.; et al. Dissecting molecular evolution in the highly diverse plant clade Caryophyllales using transcriptome sequencing. Mol. Phylogent. Evol. 2015, 32, 2001–2014. [Google Scholar] [CrossRef]
- Yao, G.; Jin, J.J.; Li, H.T.; Yang, J.B.; Mandala, V.S.; Croley, M.; Mostow, R.; Douglas, N.A.; Chase, M.W.; Christenhusz, J.M.; et al. Plastid phylogenomic insights into the evolution of Caryophyllales. Mol. Phylogent. Evol. 2019, 134, 74–86. [Google Scholar] [CrossRef]
- Goolsby, E.W.; Moore, A.J.; Hancock, L.P.; De Vos, J.M.; Edwards, E.J. Molecular evolution of key metabolic genes during transition to C4 and CAM photosynthesis. Am. J. Bot. 2018, 105, 602–613. [Google Scholar] [CrossRef]
- Nyffeler, R. The closest relatives of cacti: Insights from phylogenetic analyses of chloroplast and mitochondrial sequences with special emphasis on relationships in the tribe Anacampseroteae. Am. J. Bot. 2007, 94, 89–101. [Google Scholar] [CrossRef]
- Moore, A.J.; de Vos, J.M.; Hancock, L.P.; Goolsby, E.; Edwards, E.J. Targeted enrichment of large gene families for phylogenetic inference: Phylogeny and molecular evolution of photosynthesis genes in the Portullugo clade (Caryophyllales). Syst. Biol. 2017, 67, 367–383. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Moore, M.J.; Brockington, S.F.; Mikenas, J.; Olivieri, J.; Walker, J.F.; Smith, S.A. Improved transcriptome sampling pinpoints 26 paleopolyploidy events in Caryophyllales, including two paleo-allopolyploidy events. New Phytol. 2018, 217, 855–870. [Google Scholar] [CrossRef]
- Walker, J.F.; Yang, Y.; Feng, T.; Timoneda, A.; Mikenas, J.; Hutchison, V.; Edwards, C.; Wang, N.; Ahluwalia, S.; Olivieri, J.; et al. From cacti to carnivores: Improved phylotranscriptomic sampling and hierarchical homology inference provides further insight to the evolution of Caryophyllales. Am. J. Bot. 2018, 105, 1–17. [Google Scholar] [CrossRef]
- Wang, N.; Yang, Y.; Moore, M.J.; Brockington, S.F.; Walker, J.F.; Brown, J.W.; Liang, B.; Feng, T.; Edwards, C.; Mikenas, J.; et al. Evolution of Portulacineae marked by adaptation to harsh environments, gene tree conflict, and gene family expansion. Mol. Biol. Evol. 2018, 36, 112–126. [Google Scholar] [CrossRef]
- Hershkovitz, M.A.; Zimmer, E.A. On the evolutionary origins of the cacti. Taxon 1997, 46, 217–232. [Google Scholar] [CrossRef]
- Arakaki, M.; Christin, P.-A.; Nyffeler, R.; Lendel, A.; Eggli, U.; Ogburn, R.M.; Spriggs, E.; Moore, M.J.; Edwards, E.J. Contemporaneous and recent radiations of the world’s major succulent plant lineages. Proc. Natl. Acad. Sci. USA 2011, 108, 8379–8384. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Hernández, T.; Brown, J.W.; Schlumpberger, B.O.; Eguiarte, L.E.; Magallón, S. Beyond aridification: Multiple explanations for the elevated diversification of cacti in the New World succulent biome. New Phytol. 2014, 202, 1382–1397. [Google Scholar] [CrossRef]
- Rodrigues Silva, G.S.; Antonelli, A.; Lendel, A.; de Moraes, E.M. The impact of Quaternary climate change on the diversification and population dynamics of a South American cactus species. J. Biogeogr. 2018, 45, 76–88. [Google Scholar] [CrossRef]
- Edwards, E.J.; Nyffeler, R.; Donoghue, M.J. Basal cactus phylogeny: Implications of Pereskia (Cactaceae) paraphyly for the transition to the cactus life form. Am. J. Bot. 2005, 92, 1177–1188. [Google Scholar] [CrossRef] [PubMed]
- Butterworth, C.; Edwards, E.J. Investigating Pereskia and the earliest divergences in Cactaceae. Haseltonia 2008, 14, 46–53. [Google Scholar] [CrossRef]
- Nyffeler, R.; Eggli, U.A. farewell to dated ideas and concepts: Molecular phylogenetics and a revised suprageneric classification of the family Cactaceae. Schummania 2010, 6, 109–149. [Google Scholar]
- Majure, L.C.; Baker, M.A.; Cloud-Hughes, M.; Salywon, A.; Neubig, K.M. Phylogenomics in Cactaceae: A case study using the chollas sensu lato (Cylindropuntieae, Opuntioideae) reveals a common pattern out of the Chihuahuan and Sonoran deserts. Am. J. Bot. 2019, 106, 1327–1345. [Google Scholar] [CrossRef]
- Hernández-Hernández, T.; Hernández, H.M.; De-Nova, J.A.; Puente, R.; Eguiarte, L.E.; Magallón, S. Phylogenetic relationships and evolution of growth form in Cactaceae (Caryophyllales, Eudicotyledoneae). Amer. J. Bot. 2011, 98, 44–61. [Google Scholar] [CrossRef]
- Wallace, R.S. Molecular systematic study of the Cactaceae: Using chloroplast DNA variation to elucidate cactus phylogeny. Bradleya 1995, 13, 1–12. [Google Scholar] [CrossRef]
- Wallace, R.S.; Dickie, S.L. Systematic implications of chloroplast DNA sequences variation in the Opuntioideae. In Studies in the Opuntioideae (Cactaceae); Hunt, D.R., Taylor, N.P., Eds.; David Hunt: Milborne Port, Sherborne, UK, 2002. [Google Scholar]
- Crozier, B. Systematics of Cactaceae Juss: Phylogeny, cpDNA Evolution, and Classification with Emphasis on the Genus Mammillaria Haw. Ph.D. Thesis, University of Texas, Austin, TX, USA, 2005. [Google Scholar]
- Bárcenas, R.T.; Yesson, C.; Hawkins, J. Molecular systematics of the Cactaceae. Cladistics 2011, 27, 470–489. [Google Scholar] [CrossRef]
- Köhler, M.; Reginato, M.; Souza-Chies, T.T.; Majure, L.C. Next-generation sequencing across Opuntioideae (Cactaceae): Insights into plastome structural variation and phylogenetic markers in a prickly group. Front. Pl. Sci. 2020, 11, 729. [Google Scholar] [CrossRef] [PubMed]
- Arias, S.; Terrazas, T.; Cameron, K. Phylogenetic analysis of Pachycereus (Cactaceae, Pachycereeae) based on chloroplast and nuclear sequences. Syst. Bot. 2003, 28, 547–557. [Google Scholar]
- Arias, S.; Terrazas, T.; Arreola-Nava, H.J.; Vazquez-Sanchez, M.; Cameron, K.M. Phylogenetic relationships in Peniocereus (Cactaceae) inferred from plastid DNA sequence data. J. Plant Res. 2005, 118, 317–328. [Google Scholar] [CrossRef]
- Butterworth, C.A.; Wallace, R.S. Phylogenetic studies of Mammillaria (Cactaceae)—Insights from chloroplast sequence variation and hypothesis testing using the parametric bootstrap. Am. J. Bot. 2004, 91, 1086–1098. [Google Scholar] [CrossRef]
- Calvente, A.; Zappi, D.C.; Forest, F.; Lohmann, L.G. Molecular phylogeny, evolution, and biogeography of South American epiphytic cacti. Int. J. Plant Sci. 2011, 172, 902–914. [Google Scholar] [CrossRef]
- Calvente, A.; Moraes, E.M.; Lavor, P.; Bonatelli, I.A.; Nacaguma, P.; Versieux, L.M.; Taylor, N.P.; Zappi, D.C. Phylogenetic analyses of Pilosocereus (Cactaceae) inferred from plastid and nuclear sequences. Bot. J. Lin. Soc. 2017, 183, 25–38. [Google Scholar] [CrossRef]
- Demaio, P.H.; Barfuss, M.H.; Kiesling, R.; Till, W.; Chiapella, J.O. Molecular phylogeny of Gymnocalycium (Cactaceae): Assessment of alternative infrageneric systems, a new subgenus, and trends in the evolution of the genus. Am. J. Bot. 2011, 98, 1841–1854. [Google Scholar] [CrossRef]
- Franck, A.R.; Cochrane, B.J.; Garey, J.R. Phylogeny, biogeography, and infrageneric classification of Harrisia (Cactaceae). Syst. Bot. 2013, 38, 210–223. [Google Scholar] [CrossRef]
- Vázquez-Sánchez, M.; Terrazas, T.; Arias, S.; Ochoterena, H. Molecular phylogeny, origin and taxonomic implications of the tribe Cacteae (Cactaceae). Syst. Biod. 2013, 11, 103–116. [Google Scholar] [CrossRef]
- Sánchez, D.; Arias, S.; Terrazas, T. Phylogenetic relationships in Echinocereus (Cactaceae, Cactoideae). Syst. Bot. 2014, 39, 1183–1196. [Google Scholar] [CrossRef]
- Vázquez-Lobo, A.; Morales, G.A.; Arias, S.; Golubov, J.; Hernández-Hernández, T.; Mandujano, M.C. Phylogeny and biogeographic history of Astrophytum (Cactaceae). Syst. Bot. 2016, 40, 1022–1030. [Google Scholar] [CrossRef]
- Vargas-Luna, M.D.; Hernández-Ledesma, P.; Majure, L.C.; Puente, R.; Hernández, H.M.; Bárcenas, R.T. Splitting Echinocactus: Morphological and molecular evidence support the recognition of Homalocephala as a distinct genus in the Cacteae. PhytoKeys 2018, 111, 31–59. [Google Scholar] [CrossRef]
- Barrios, D.; Arias, S.; González-Torres, L.R.; Majure, L.C. Phylogeny and taxonomy of the Antillean endemic genus Leptocereus (Cactaceae) inferred from chloroplast markers and morphological evidence. Pl. Syst. Evol. 2020, 306, 63. [Google Scholar] [CrossRef]
- Breslin, P.; Wojciechowski, M.; Majure, L.C. Molecular phylogeny of the Mammilloid clade (Cactaceae) of Baja California and adjacent regions: Resolving the monophyly of Mammillaria with new combinations in Cochemiea. Taxon 2021, 70, 308–323. [Google Scholar] [CrossRef]
- Majure, L.C.; Clase, T.; Encarnación, Y.; Peguero, B.; Ho, K.; Barrios, D. Phylogenetics of Leptocereus (Cactaceae) on Hispaniola: Clarifying species limits in the L. weingartianus complex and a new species from the Sierra de Bahoruco. Phytokeys 2021, 172, 17–37. [Google Scholar] [CrossRef]
- Majure, L.C.; Barrios, D.; Díaz, E.; Bacci, L.F.; Encarnación, Y. Phylogenomics of the Caribbean Melocacti: Cryptic species and multiple invasions. Taxon 2022, 71, 993–1012. [Google Scholar] [CrossRef]
- Hunt, D.R. That’s Opuntia, that was! Succul. Plant Res. 2002, 6, 245–248. [Google Scholar]
- Hunt, D.R. An interim checklist of the subfamily Opuntioideae (Cactaceae) in the Caribbean region and South America. Succul. Plant Res. 2014, 8, 201–223. [Google Scholar]
- Griffith, M.P. Grusonia pulchella reclassification and its impact on the genus Grusonia: Morphological and molecular evidence. Haseltonia 2002, 9, 86–93. [Google Scholar]
- Griffith, M.P.; Porter, J.M. Phylogeny of Opuntioideae. Int. J. Plant Sci. 2009, 170, 107–116. [Google Scholar] [CrossRef]
- Majure, L.C.; Puente, R.; Griffith, M.P.; Judd, W.S.; Soltis, P.S.; Soltis, D.E. Phylogeny of Opuntia s.s. (Cactaceae): Clade delineation, geographic origins, and reticulate evolution. Am. J. Bot. 2012, 99, 847–864. [Google Scholar] [CrossRef]
- Majure, L.C.; Soltis, D.E.; Soltis, P.S.; Judd, W.S. A case of mistaken identity, Opuntia abjecta, long-lost in synonymy under the Caribbean species, O. triacantha, and a reassessment of the enigmatic O. cubensis. Brittonia 2014, 66, 118–130. [Google Scholar] [CrossRef]
- Majure, L.C.; Puente, R. Phylogenetic relationships and morphological evolution in Opuntia s. str. and closely related members of tribe Opuntieae. Succul. Plant Res. 2014, 8, 9–30. [Google Scholar]
- Majure, L.C.D.; Barrios, E.; Díaz, B.; Zumwalde, W.T.; Negrón-Ortiz, V. Pleistocene drying underlies the evolutionary history of the Caribbean endemic, insular giant, Consolea (Opuntioideae). Am. J. Bot. 2021, 108, 200–215. [Google Scholar] [CrossRef]
- Ritz, C.M.; Reiker, J.; Charles, G.; Hoxey, P.; Hunt, D.; Lowry, M.; Stuppy, W.; Taylor, N. Molecular phylogeny and character evolution in terete-stemmed Andean opuntias (Cactaceae–Opuntioideae). Molec. Phylogen. Evol. 2012, 65, 668–681. [Google Scholar] [CrossRef]
- Las Peñas, M.L.; Kiesling, R.; Bernardello, G. Phylogenetic reconstruction of the genus Tephrocactus (Cactaceae) based on molecular, morphological and cytogenetical data. Taxon 2019, 68, 714–730. [Google Scholar] [CrossRef]
- Bárcenas, R.T. A molecular phylogenetic approach to the systematics of Cylindropuntieae (Opuntioideae, Cactaceae). Cladistics 2016, 32, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Mauseth, J.D. Structure-function relationships in highly modified shoots of Cactaceae. Ann. Bot. 2006, 98, 901–926. [Google Scholar] [CrossRef] [PubMed]
- Mauseth, J.D. Tiny but complex foliage leaves occur in many “leafless” cacti (Cactaceae). Int. J. Plant Sci. 2007, 168, 845–853. [Google Scholar] [CrossRef]
- Nobel, P.S.; Bobich, E.G. Environmental biology. In Cacti: Biology and Uses; Nobel, P.S., Ed.; University of California Press: Berkeley, CA, USA, 2002; pp. 57–74. [Google Scholar]
- Winter, K.; Garcia, M.; Holtum, J.A.M. Drought-stress-induced up-regulation of CAM in seedlings of a tropical cactus, Opuntia elatior, operating predominantly in the C3 mode. J. Exp. Bot. 2011, 62, 4037–4042. [Google Scholar] [CrossRef] [PubMed]
- Mauseth, J.D. An introduction to cactus areoles part II. Cactus Succ. J. 2017, 89, 219–229. [Google Scholar] [CrossRef]
- Majure, L.C.; Puente, R.; Pinkava, D.J. Miscellaneous chromosome counts in Opuntieae DC. (Cactaceae) with a compilation of counts for the group. Haseltonia 2012, 18, 67–78. [Google Scholar] [CrossRef]
- Baker, M.A.; Pinkava, D.J. Chromosome numbers in some cacti of Western North America—IX. Haseltonia 2018, 25, 4–28. [Google Scholar] [CrossRef]
- Liu, X.; Yang, H.; Zhao, J.; Zhou, B.; Li, T.; Xiang, B. The complete chloroplast genome sequence of the folk medicinal and vegetable plant purslane (Portulaca oleracea L.). J. Hort. Sci. Biotech. 2018, 93, 356–365. [Google Scholar] [CrossRef]
- Neubig, K.M.; Whitten, W.M.; Abbott, J.R.; Elliott, S.; Soltis, D.E.; Soltis, P.S. Variables affecting DNA preservation in archival plant specimens. In DNA Banking for the 21st Century: Proceedings of the US Workshop on DNA Banking; Applequist, W.L., Campbell., L.M., Eds.; Missouri Botanical Garden; William L. Brown Center: St. Louis, MI, USA, 2014; pp. 81–112. [Google Scholar]
- Katoh, K.; Standley, D.M. A simple method to control over-alignment in the MAFFT multiple sequence alignment program. Bioinformatics 2016, 32, 1933–1942. [Google Scholar] [CrossRef]
- Stamatakis, A. RaxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kuhnert, D.; De Maio, N.; et al. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [PubMed]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeifferand, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop; GCE: New Orleans, LA, USA, 2010; pp. 1–8. [Google Scholar]
- Smith, S.A.; O’Meara, B.C. treePL: Divergence time estimation using penalized likelihood for large phylogenies. Bioinformatics 2012, 28, 2689–2690. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A. FigTree: Tree Figure Drawing Tool, v1. 4.2; Institute of Evolutionary Biology, University of Edinburgh: Edinburgh, UK, 2012.
- Rambaut, A.; Drummond, A.J. LogCombiner, v2. 6.2.; Institute of Evolutionary Biology, University of Edinburgh: Edinburgh, UK, 2014.
- Rambaut, A.; Drummond, A.J. TreeAnnotator, v2.6.2. MCMC Output Analysis. 2020. Available online: https://www.beast2.org/treeannotator/ (accessed on 1 March 2021).
- Mauseth, J.D. Some cacti produce large, ephemeral foliage leaves when they flower. Int. J. Pl. Sci. 2022, 183, 119–127. [Google Scholar] [CrossRef]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis, Version 2.75. 2017. Available online: http://mesquiteproject.org (accessed on 1 March 2021).
- Yu, Y.; Harris, A.J.; Blair, C.; He, X. RASP (Reconstruct Ancestral State in Phylogenies): A tool for historical biogeography. Mol. Phylogenetics Evol. 2015, 87, 46–49. [Google Scholar] [CrossRef]
- Matzke, N.J. BioGeoBEARS: BioGeography with Bayesian (and Likelihood) Evolutionary Analysis in R Scripts; University of California: Berkeley, CA, USA, 2013. [Google Scholar]
- Matzke, N.J. Probabilistic historical biogeography: New models for founder-event speciation, imperfect detection, and fossils allow improved accuracy and model-testing. Frontiers Biogeogr. 2013, 54, 242–248. [Google Scholar]
- Matzke, N.J. Model Selection in Historical Biogeography Reveals that Founder-event Speciation is a Crucial Process in Island Clades. Syst. Biol. 2014, 63, 951–970. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria. 2020. Available online: https://www.R-project.org/ (accessed on 1 March 2021).
- Dupin, J.; Matzke, N.J.; Särkinen, T.; Knapp, S.; Olmstead, R.G.; Bohs, L.; Smith, S.D. Bayesian estimation of the global biogeographical history of the Solanaceae. J. Biogeogr. 2017, 44, 887–899. [Google Scholar] [CrossRef]
- Matzke, N.J. Stochastic Mapping under Biogeographical Models. PhyloWiki BioGeoBEARS website. 2016. Available online: https://phylo.wikidot.com/biogeobears#stochastic_mapping (accessed on 1 March 2021).
- Griffith, G.E.; Omernik, J.M.; Azevedo, S.H. Ecological Classification of the Western Hemisphere; Unpublished Report; Environmental Protection Agency, Western Ecology Division: Corvallis, OR, USA, 1998; 49p.
- Ree, R.H.; Smith, S.A. Maximum likelihood inference of geographic range evolution by dispersal, local extinction, and cladogenesis. Syst. Biol. 2008, 57, 4–14. [Google Scholar] [CrossRef]
- Massana, K.A.; Beaulieu, J.M.; Matzke, N.J.; O’Meara, B.C. Non-null effects of the null range in biogeographic models: Exploring parameter estimation in the dec model. BioRxiv 2015. [Google Scholar] [CrossRef]
- Ronquist, F. Duspersal-vicariance analysis: A new approach to the quantification of historical biogeography. Syst. Biol. 1997, 46, 195–203. [Google Scholar] [CrossRef]
- Landis, M.J.; Matzke, N.J.; Moore, B.R.; Huelsenbeck, J.P. Bayesian analysis of biogeography when the number of areas is large. Syst. Biol. 2013, 62, 789–804. [Google Scholar] [CrossRef] [PubMed]
- Burnham, K.P.; Anderson, D.R. Multimodel inference: Understanding AIC and BIC in model selection. Soc. Method Res. 2004, 33, 261–304. [Google Scholar] [CrossRef]
- Ree, R.H.; Sanmartín, I. Conceptual and statistical problems with the DEC+J model of founder-event speciation and its comparison with DEC via model selection. J. Biogeogr. 2018, 45, 741–749. [Google Scholar] [CrossRef]
- Matzke, N.J. Statistical comparison of DEC and DEC+J is identical to comparison of two ClaSSE submodels and is therefore valid. J. Biogeogr. 2022, 49, 1805–1824. [Google Scholar] [CrossRef]
- Sánchez, D.; Vázquez-Benítez, B.; Vázquez-Sánchez, M.; Aquino, D.; Arias, S. Phylogenetic relationsjhips in Coryphantha and implications on Pelecyphora and Escobaria (Cacteae, Cactoideae, Cactaceae). Phytokeys 2022, 188, 115–165. [Google Scholar] [CrossRef]
- Acha, S.; Majure, L.C. A new approach using targeted sequence capture for phylogenomic studies across Cactaceae. Genes 2022, 13, 350. [Google Scholar] [CrossRef]
- Las Peñas, M.L.; Bernardello, G. Divergence time estimation and mapping of morphological and cytogenetical data in the southern South American geophyte genus Pterocactus (Cactaceae). Taxon 2021, 70, 477–698. [Google Scholar] [CrossRef]
- Lewis, D.A.; Nobel, P.S. Thermal energy exchange model and water loss of a barrel cactus, Ferocactus acanthodes. Plant Physiol. 1977, 60, 609–616. [Google Scholar] [CrossRef] [PubMed]
- Nobel, P.S. Surface temperatures of cacti—Influences of environmental and morphological factors. Ecology 1978, 59, 986–996. [Google Scholar] [CrossRef]
- Nobel, P.S. Interception of photosynthetically active radiation by cacti of different morphologies. Oecologia 1980, 45, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Nobel, P.S. Morphology, surface temperatures, and northern limits of columnar cacti of the Sonoran Desert. Ecology 1980, 61, 1–7. [Google Scholar] [CrossRef]
- Nobel, P.S. Spine influences on PAR interception, stem temperature and nocturnal acid acumulation by cacti. Plant Cell Environ. 1983, 6, 153–159. [Google Scholar] [CrossRef]
- Ju, J.; Bai, H.; Zheng, Y.; Zhao, T.; Fang, R.; Jiang, L. A mult-structural and multi-functional integrated fog collection system in cactus. Nat. Commun. 2012, 3, 1247. [Google Scholar] [CrossRef]
- Gibson, A.C.; Nobel, P.S. The Cactus Primer; Harvard University Press: Cambridge, MA, USA, 1986. [Google Scholar]
- Boke, N.H. Histogenesis of the leaf and areole in Opuntia cylindrica. Am. J. Bot. 1944, 31, 299–316. [Google Scholar] [CrossRef]
- Bailey, I.W. Comparative anatomy of the leaf-bearing Cactaceae, I. Foliar vasculature of Pereskia, Pereskiopsis and Quiabentia. J. Arnold Arbor. 1960, 41, 341–356. [Google Scholar] [CrossRef]
- Bailey, I.W. Comparative anatomy of the leaf-bearing Cactaceae, XVII. Preliminary observations on the problem of transitions from broad to terete leaves. J. Arnold Arbor. 1968, 49, 370–376. [Google Scholar] [CrossRef]
- Tapia, H.J.; Bárcenas-Arguello, M.L.; Terrazas, T. Phylogeny and circumscription of Cephalocereus (Cactaceae) based on molecular and morphological evidence. Syst. Bot. 2017, 42, 1–15. [Google Scholar] [CrossRef]
- Nyffeler, R.; Eggli, U.; Ogburn, M.; Edwards, E. Variations on a theme: Repeated evolution of succulent life forms in the Portulacineae (Caryophyllales). Haseltonia 2008, 14, 26–36. [Google Scholar] [CrossRef]
- Wilson, J.S.; Pitts, J.P. Illuminating the lack of consensus among descriptions of earth history data in the North American deserts: A resource for biologists. Prog. Phys. Geogr. 2010, 34, 419–441. [Google Scholar] [CrossRef]
- Breslin, P.; Wojciechowski, M.; Majure, L.C. Remarkably rapid, recent diversification of Cochemiea and Mammillaria (Cactaceae) in the Baja California, Mexico Region. Am. J. Bot. 2022, 109, 1472–1487. [Google Scholar] [CrossRef] [PubMed]
- Lavor, P.; Calvente, A.; Versieux, L.M.; Sanmartin, I. Bayesian spatio-temporal reconstruction reveals rapid diversification and Pleistocene range expansion in the widespread columnar cactus Pilosocereus. J. Biogeogr. 2018, 46, 238–250. [Google Scholar] [CrossRef]
- Pennington, R.T.; Prado, D.E.; Pendry, C.A. Neotropical seasonally dry forests and Quaternary vegetation changes. J. Biogeogr. 2000, 27, 261–273. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clade | Crown Age | CI |
|---|---|---|
| Pereskia + Leuenbergeria | 15.19 | 7.24–27.83 |
| Maihuenia + the rest | 27.22 | 23.35–31.23 |
| Cactoideae | 25.93 | 19.88–28.16 |
| Cacteae | 17.53 | 12.32–23.25 |
| Core Cactoideae I | 17.53 | 12.32–23.25 |
| Core Cactoideae II | 20.83 | 16.39–25.72 |
| Opuntioideae | 16.73 | 11.32–22.18 |
| Opuntieae | 7.92 | 3.98–12.11 |
| Tacinga | 1.75 | 0.16–4.51 |
| Miqueliopuntia + Tunilla | 3.53 | 1.51–5.88 |
| Consolea | 0.73 | 0.14–1.64 |
| Opuntia | 4.39 | 2.52–6.43 |
| Tephrocacteae | 13.34 | 8.74–18-86 |
| Tephrocactus | 8.72 | 4.65–13.08 |
| Pterocactus | 3.37 | 0.45–6.95 |
| Austrocylindropuntia + Cumulopuntia | 7.37 | 3.73–11.54 |
| Maihueniopsis | 8.65 | 3.83–13.78 |
| Cylindropuntieae | 12.2 | 7.86–16.55 |
| Quiabentia | 3.58 | 0.37–7.66 |
| Pereskiopsis | 3 | 0.87–5.82 |
| Micropuntia | 2.77 | 0.38–5.54 |
| Cylindropuntia | 6.4 | 3.86–9.05 |
| Grusonia | 3.26 | 1.41–5.77 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Majure, L.C.; Achá, S.; Baker, M.A.; Puente-Martínez, R.; Köhler, M.; Fehlberg, S. Phylogenomics of One of the World’s Most Intriguing Groups of CAM Plants, the Opuntioids (Opuntioideae: Cactaceae): Adaptation to Tropical Dry Forests Helped Drive Prominent Morphological Features in the Clade. Diversity 2023, 15, 570. https://doi.org/10.3390/d15040570
Majure LC, Achá S, Baker MA, Puente-Martínez R, Köhler M, Fehlberg S. Phylogenomics of One of the World’s Most Intriguing Groups of CAM Plants, the Opuntioids (Opuntioideae: Cactaceae): Adaptation to Tropical Dry Forests Helped Drive Prominent Morphological Features in the Clade. Diversity. 2023; 15(4):570. https://doi.org/10.3390/d15040570
Chicago/Turabian StyleMajure, Lucas C., Serena Achá, Marc A. Baker, Raul Puente-Martínez, Matias Köhler, and Shannon Fehlberg. 2023. "Phylogenomics of One of the World’s Most Intriguing Groups of CAM Plants, the Opuntioids (Opuntioideae: Cactaceae): Adaptation to Tropical Dry Forests Helped Drive Prominent Morphological Features in the Clade" Diversity 15, no. 4: 570. https://doi.org/10.3390/d15040570
APA StyleMajure, L. C., Achá, S., Baker, M. A., Puente-Martínez, R., Köhler, M., & Fehlberg, S. (2023). Phylogenomics of One of the World’s Most Intriguing Groups of CAM Plants, the Opuntioids (Opuntioideae: Cactaceae): Adaptation to Tropical Dry Forests Helped Drive Prominent Morphological Features in the Clade. Diversity, 15(4), 570. https://doi.org/10.3390/d15040570