


Evaluating the Impact of Headstarting on the Critically Endangered Spoon-Billed Sandpiper Calidris pygmaea

 , , ,
, , ,
Abstract
:1. Introduction
2. Methods
2.1. Ringing and Colour Marking
2.2. Collection of Eggs and Artificial Incubation for HS
2.3. Chick Rearing and Release
2.4. Calculations
3. Results
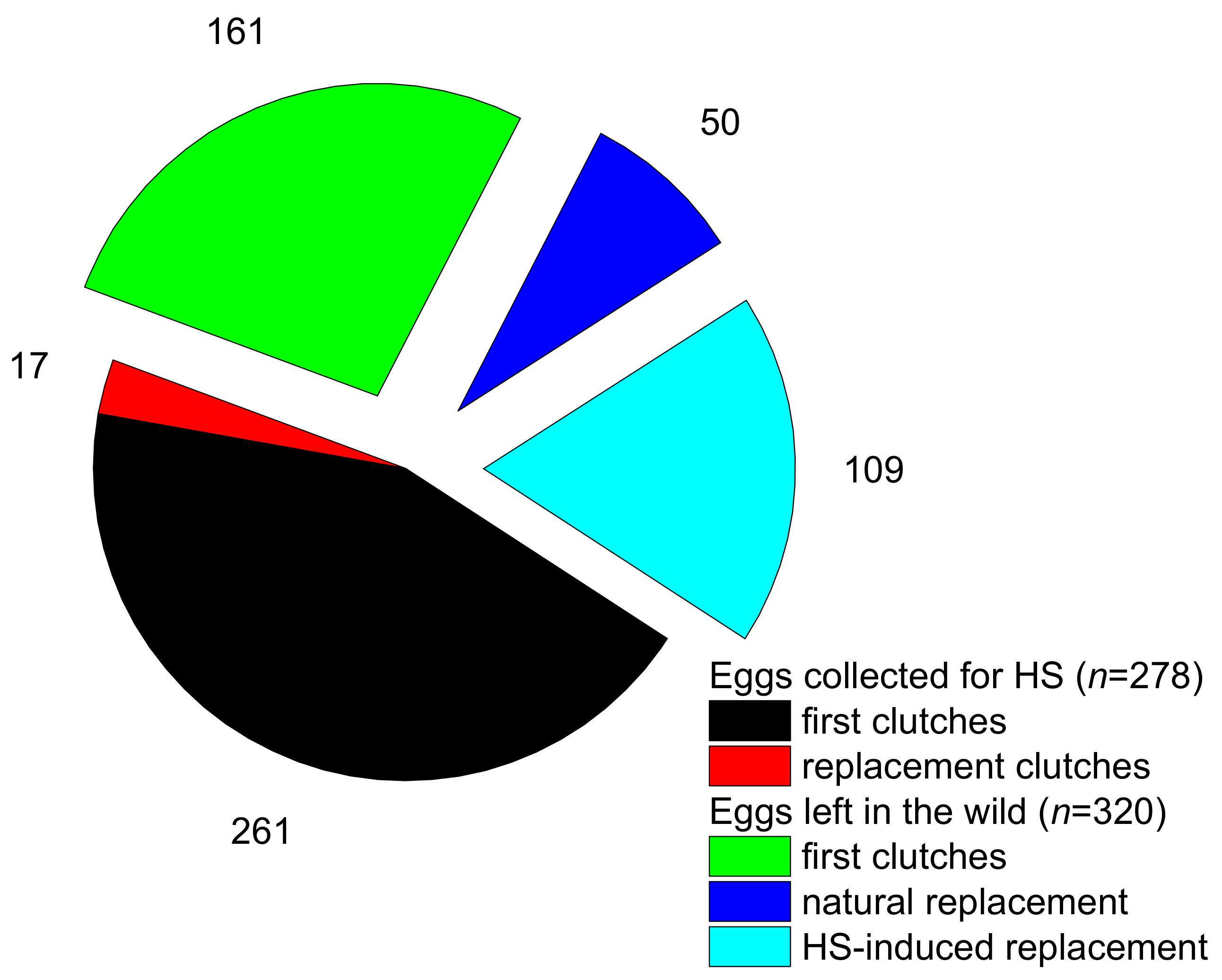
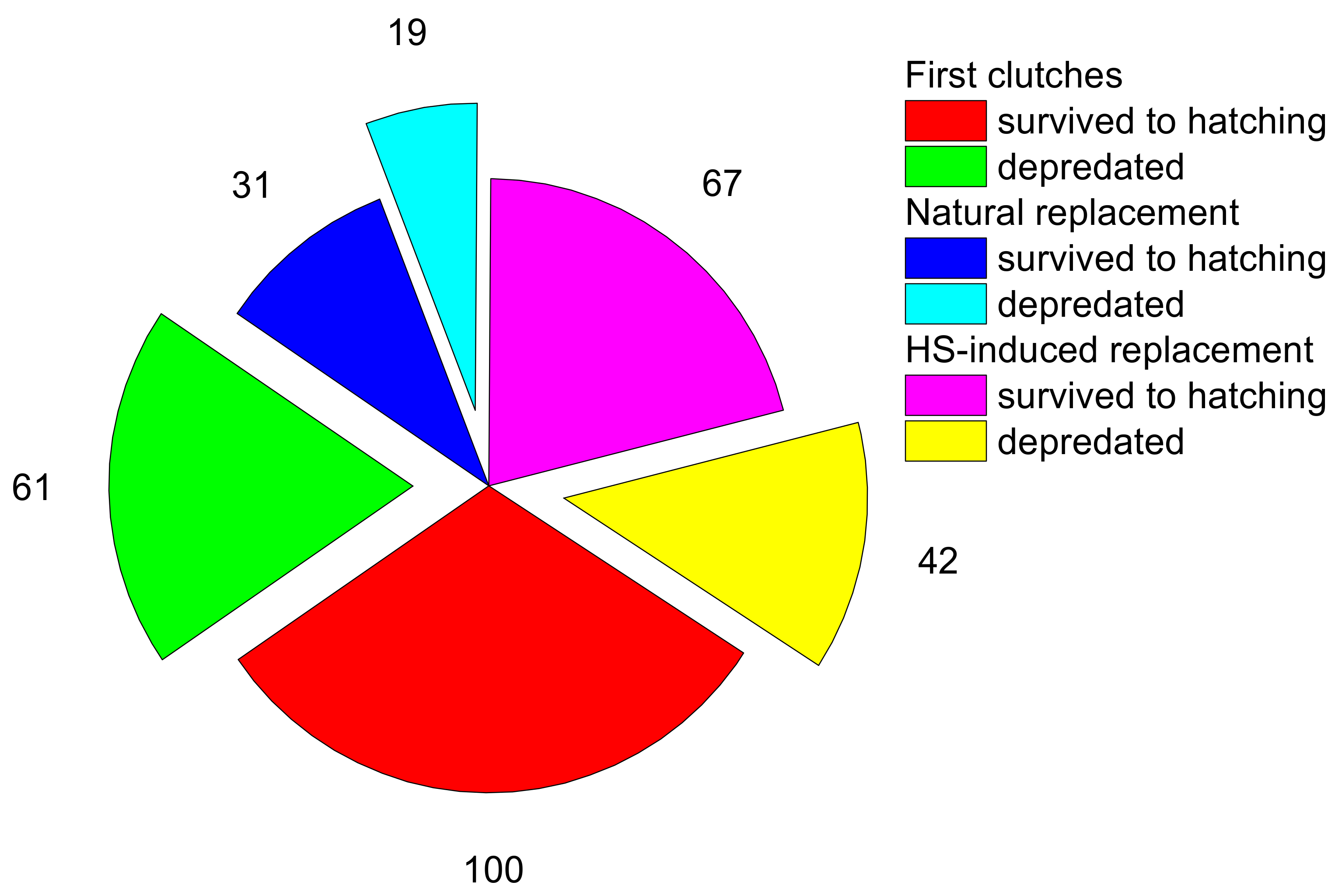
3.1. Nest Fate
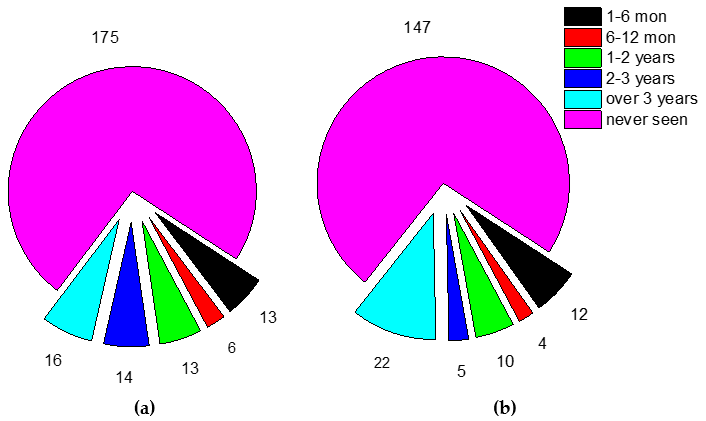
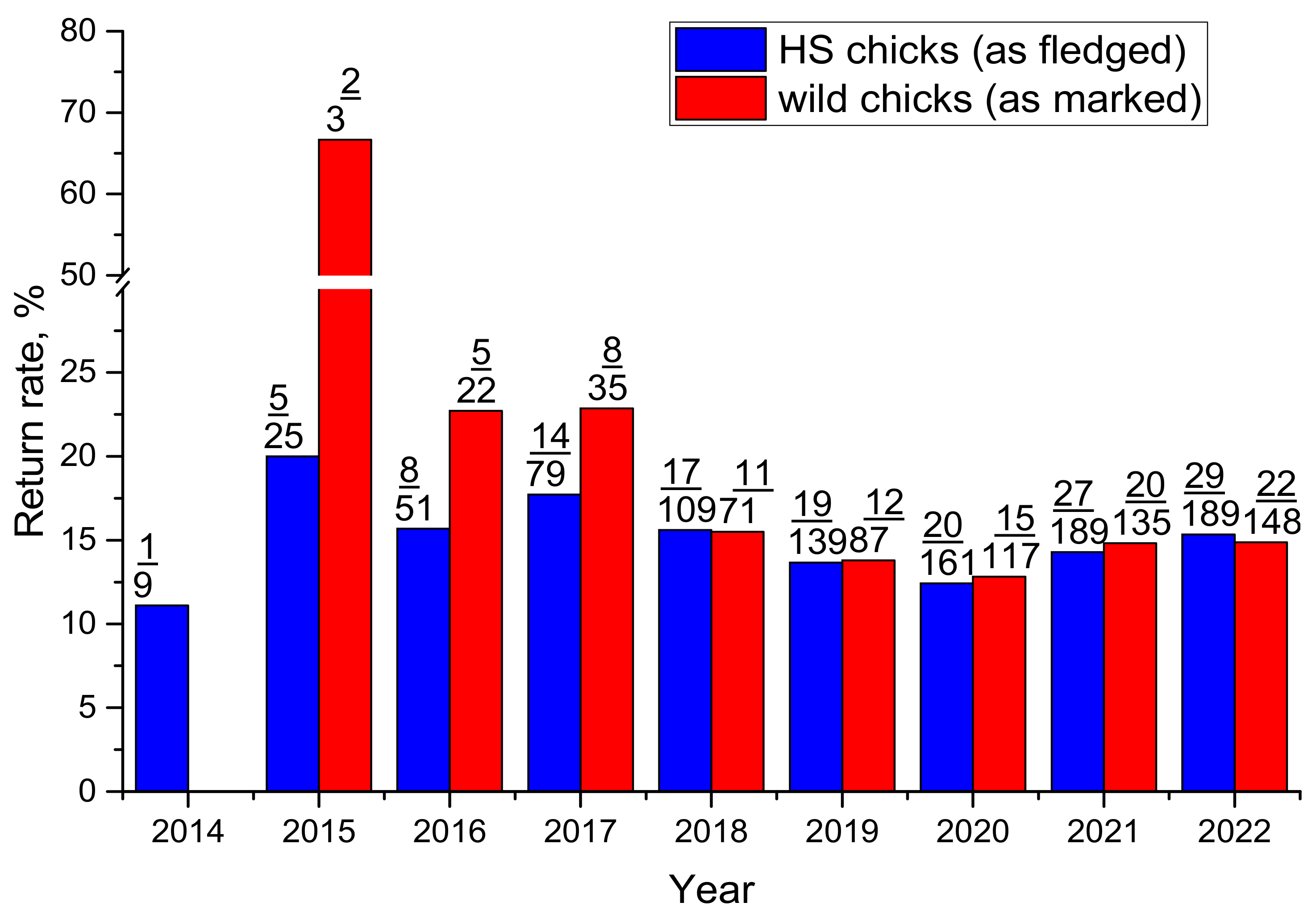
3.2. Fate of Chicks
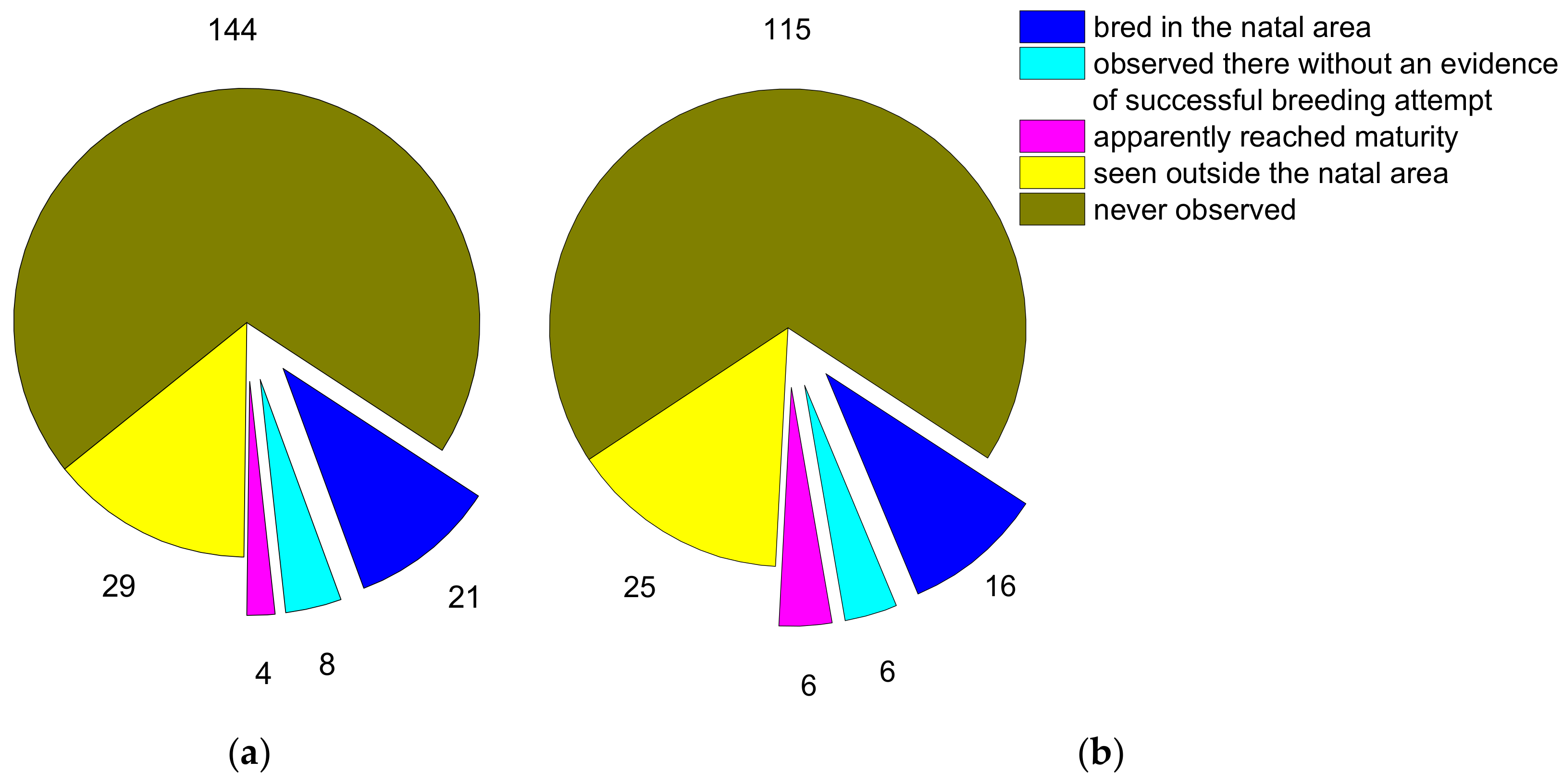
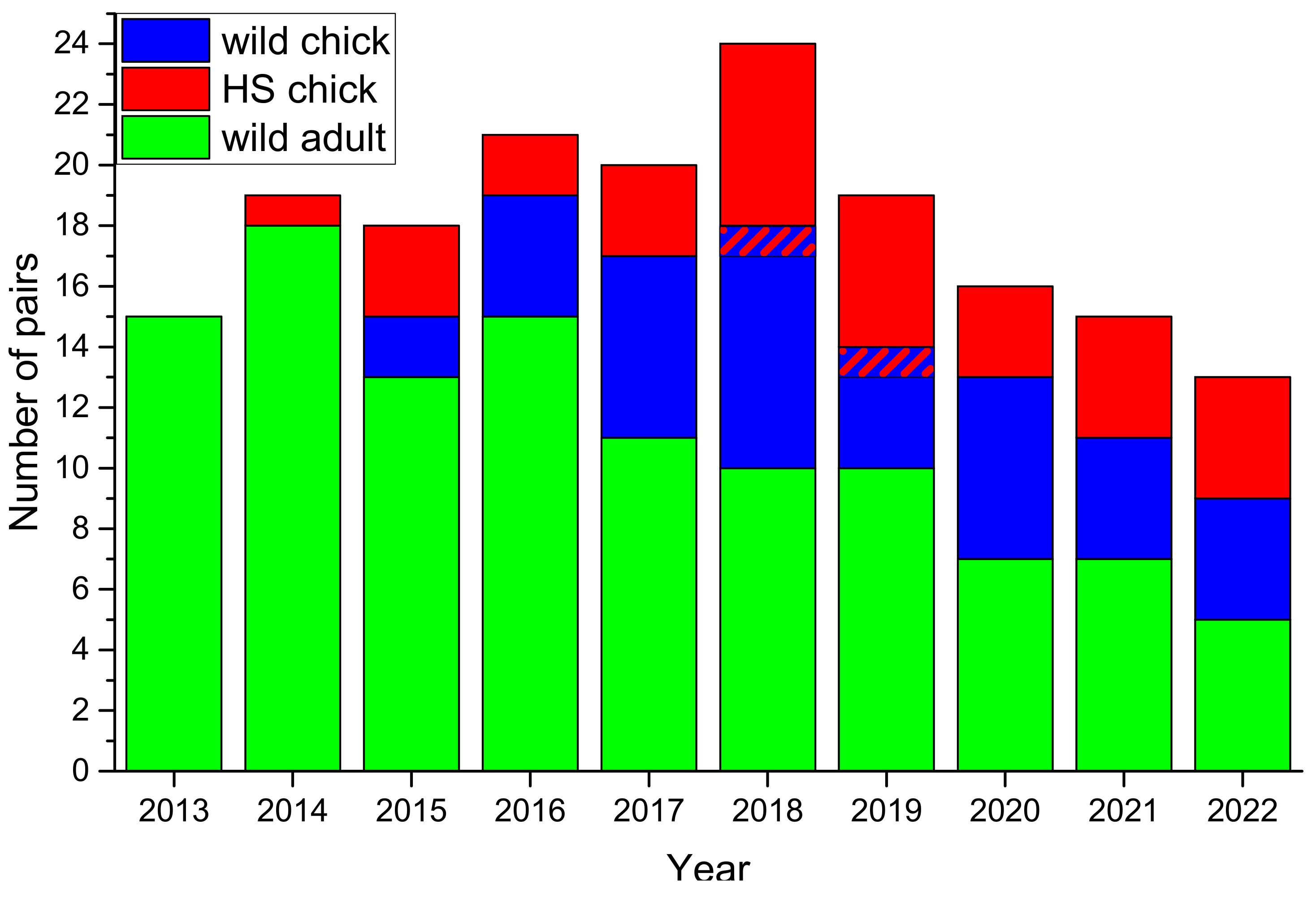
3.3. Population Level Impact
4. Discussion
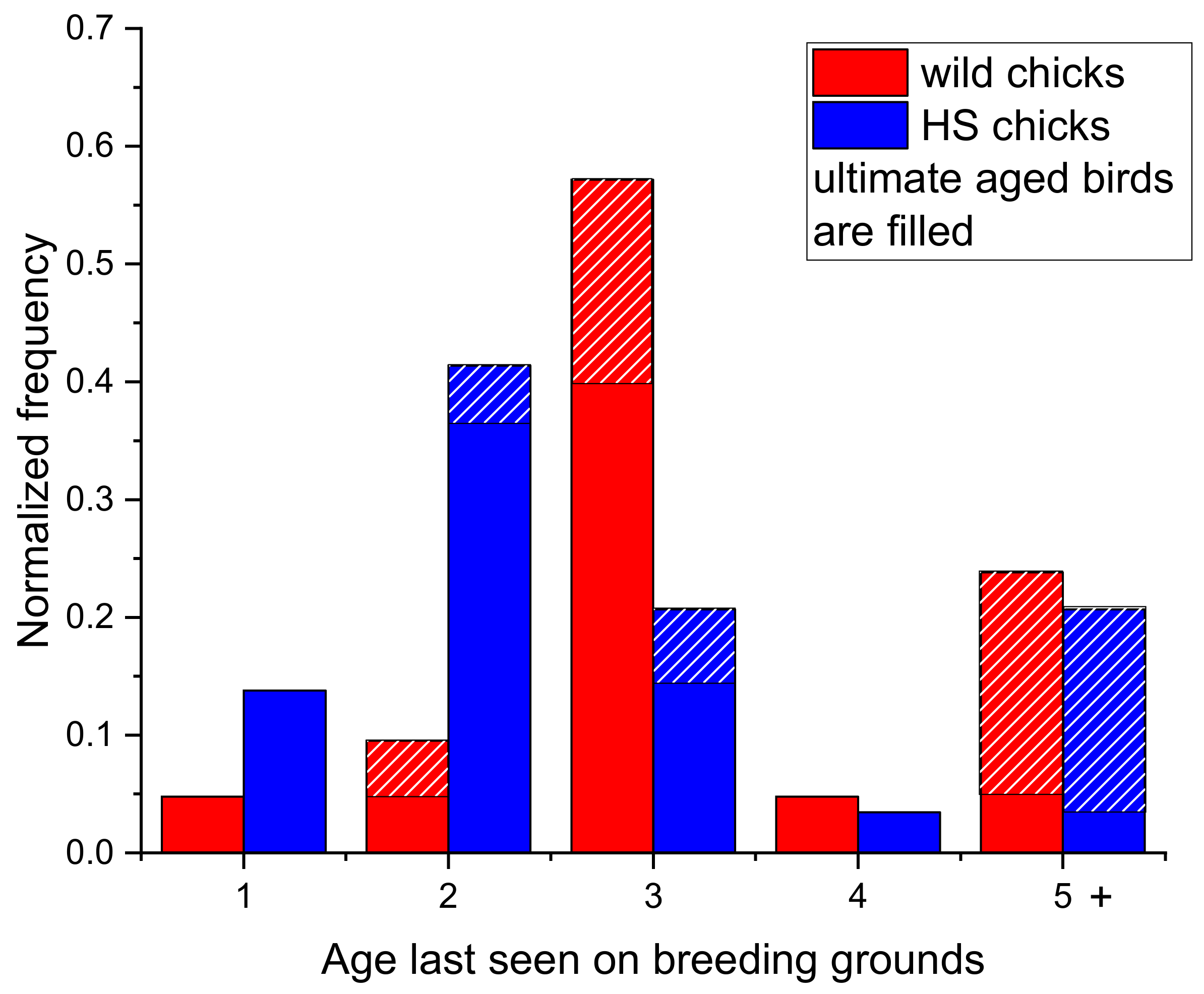
4.1. Survival of Chicks
4.2. Sex Issues in Chicks
4.3. Comparison to the Northern SbS Population
4.4. Comparison to Other Headstarted Species
4.5. Suggestions for Headstarting Improvement
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Payevsky, V.A. Demography of Birds; Nauka: Leningrad, Russia, 1985. [Google Scholar]
- Lack, D. The Natural Regulation of Animal Numbers; Clarendon Press: Oxford, UK, 1954; p. 343. [Google Scholar]
- Studds, C.E.; Kendall, B.E.; Murray, N.J.; Wilson, H.B.; Rogers, D.I.; Clemens, R.S.; Gosbell, K.; Hassell, C.J.; Jessop, R.; Melville, D.S.; et al. Rapid population decline in migratory shorebirds relying on Yellow Sea tidal mudflats as stopover sites. Nat. Commun. 2017, 8, 14895. [Google Scholar] [CrossRef] [PubMed]
- Piersma, T.; Lok, T.; Chen, Y.; Hassell, C.J.; Yang, H.-Y.; Boyle, A.; Slaymaker, M.; Chan, Y.-C.; Melville, D.S.; Zhang, Z.-W.; et al. Simultaneous declines in summer survival of three shorebird species signals a flyway at risk. J. Appl. Ecol. 2016, 53, 479–490. [Google Scholar] [CrossRef]
- Murray, N.J.; Marra, P.P.; Fuller, R.A.; Clemens, R.S.; Dhanjal-Adams, K.; Gosbell, K.B.; Hassell, C.J.; Iwamura, T.; Melville, D.; Minton, C.D.T.; et al. The large-scale drivers of population declines in a long-distance migratory shorebird. Ecography 2018, 41, 867–876. [Google Scholar] [CrossRef]
- Melville, D.S.; Chen, Y.; Ma, Z. Shorebirds along the Yellow Sea coast of China face an uncertain future—A review of threats. Emu-Austral Ornithol. 2016, 116, 100–110. [Google Scholar] [CrossRef]
- Thomas, P.; Boyer, D.M.; Oehler, D.A.; Silver, S.; Perrotti, L. Headstarting as a Conservation Strategy for Threatened and Endangered Species. In Scientific Foundations of Zoos and Aquariums: Their Role in Conservation and Research; Kaufman, A.B., Bashaw, M.J., Maple, T.L., Eds.; Cambridge University Press: Cambridge, UK, 2019; pp. 91–111. [Google Scholar] [CrossRef]
- Maxwell, J.M.; Jamieson, I.G. Survival and Recruitment of Captive-Reared and Wild-Reared Takahe in Fiordland, New Zealand. Conserv. Biol. 1997, 11, 683–691. [Google Scholar] [CrossRef]
- Cunninghame, F.; Switzer, R.; Parks, B.; Young, G.; Carrion, A.; Medranda, P.; Sevilla, C. Conserving the Critically Endangered Mangrove Finch: Head-Starting to Increase Population Size. In Galapagos Report 2013–2014; GNPD; GCREG; CDF; GC: Puerto Ayora, Ecuador, 2015. [Google Scholar]
- Collins, S.A.; Sanders, F.J.; Jodice, P.G.R. Assessing conservation tools for an at-risk shorebird: Feasibility of headstarting for American Oystercatchers Haematopus palliatus. Bird Conserv. Int. 2016, 26, 451–465. [Google Scholar] [CrossRef]
- Hiscock, N.; Brand, E.; Calvo-Carrasco, D.; Donaldson, L. Headstarting the Black-Tailed Godwit Limosa Limosa Limosa at Key Breeding Sites for the UK Population: 2018 Annual Report; Wildfowl & Wetlands Trust: Slimbridge, UK, 2018. [Google Scholar]
- van Heezik, Y.; Maloney, R.F.; Seddon, P.J. Movements of translocated captive-bred and released Critically Endangered kaki (black stilts) Himantopus novaezelandiae and the value of long-term post-release monitoring. Oryx 2009, 43, 639–647. [Google Scholar] [CrossRef]
- Powell, A.N.; Cuthbert, F.J.; Wemmer, L.C.; Doolittle, A.W.; Feirer, S.T. Captive-rearing piping plovers: Developing techniques to augment wild populations. Zoo Biol. 1997, 16, 461–477. [Google Scholar] [CrossRef]
- Neuman, K.K.; Stenzel, L.E.; Warriner, J.C.; Page, G.W.; Erbes, J.L.; Eyster, C.R.; Miller, E.; Henkel, L.A. Success of captive-rearing for a threatened shorebird. Endanger. Species Res. 2013, 22, 85–94. [Google Scholar] [CrossRef]
- Boyd, H.; Piersma, T. Changing balance between survival and recruitment explains population trends in Red Knots Calidris canutus islandica wintering in Britain, 1969–1995. Ardea 2001, 89, 301–317. [Google Scholar]
- Lappo, E.G.; Tomkovich, P.S.; Syroechkovskiy, E.E. Atlas of Breeding Waders in the Russian Arctic; Publishing House “UF Ofsetnaya Pechat”: Moscow, Russia, 2012; p. 448. [Google Scholar]
- Zockler, C.; Beresford, A.E.; Bunting, G.; Chowdhury, S.U.; Clark, N.A.; Fu, V.W.K.; Htin Hla, T.; Morozov, V.V.; Syroechkovskiy, E.E.; Kashiwagi, M.; et al. The winter distribution of the Spoon-billed Sandpiper Calidris pygmaeus. Bird Conserv. Int. 2016, 26, 476–489. [Google Scholar] [CrossRef]
- Zockler, C.; Syroechkovskiy, E.E.; Atkinson, P.W. Rapid and continued population decline in the Spoon-billed Sandpiper Eurynorhynchus pygmeus indicates imminent extinction unless conservation action is taken. Bird Conserv. Int. 2010, 20, 95–111. [Google Scholar] [CrossRef]
- Tomkovich, P.S.; Syroechkovski, J.E.E.; Lappo, E.G.; Zöckler, C. First indications of a sharp population decline in the globally threatened Spoon-billed Sandpiper Eurynorhynchus pygmeus. Bird Conserv. Int. 2002, 12, 1–18. [Google Scholar] [CrossRef]
- Syroechkovski, E.E.; Tomkovich, P.S.; Kashiwagi, M.; Taldenkov, I.A.; Buzin, V.A.; Lappo, E.G.; Zöckler, C. Population decline in the spoon-billed sandpiper (Eurynorhynchus pygmeus) in northern Chukotka based on monitoring on breeding grounds. Biol. Bull. 2010, 37, 941–951. [Google Scholar] [CrossRef]
- Green, R.E.; Syriechkovsliy, E.E.; Anderson, G.Q.A.; Chang, Q.; Chowdhury, S.U.; Clark, J.A.; Foysal, M.; Gerasimov, Y.; Hughes, B.; Kelly, C.; et al. New estimated of the size and trend of the world population of the Spoon-billed Sandpiper using three independent statistical models. Wader Study 2021, 128, 22–35. [Google Scholar] [CrossRef]
- Clark, N.A.; Anderson, G.Q.A.; Li, J.; Syroechkovskiy, E.E.; Tomkovich, P.S.; Zöckler, C.; Lee, R.; Green, R.E. First formal estimate of the world population of the Critically Endangered spoon-billed sandpiper Calidris pygmaea. Oryx 2018, 52, 137–146. [Google Scholar] [CrossRef]
- Peng, H.-B.; Anderson, G.Q.A.; Chang, Q.; Choi, C.-Y.; Chowdhury, S.U.; Clark, N.A.; Gan, X.; Hearn, R.D.; Li, J.; Lappo, E.G.; et al. The intertidal wetlands of southern Jiangsu Province, China—Globally important for Spoon-billed Sandpipers and other threatened waterbirds, but facing multiple serious threats. Bird Conserv. Int. 2017, 27, 305–322. [Google Scholar] [CrossRef]
- Menxiu, T.; Lin, Z.; Li, J.; Zöckler, C.; Clark, N.A. The critical importance of the Rudong mudflats, Jiangsu Province, China in the annual cycle of the Spoon-billed Sandpiper Calidris pygmeus. Wader Study Group Bull. 2012, 119, 208–211. [Google Scholar]
- Clark, N.A.; Pain, D.; Green, R.E. Saving the Spoon-billed Sandpiper: An update on the conservation programme. Br. Birds 2014, 107, 467–475. [Google Scholar]
- Chang, Q.; Anderson, G.Q.A.; Brides, K.; Clark, J.A.; Clark, N.A.; Hearn, R.; Leung, K.; Melville, D.S.; Weston, E.; Weston, J.; et al. A high proportion of the world population of the Spoon-billed Sandpiper occurs at Tiaozini, China, during the post-breeding moult. Wader Study 2019, 126, 35–42. [Google Scholar] [CrossRef]
- Zöckler, C.; Chowdhury, S.U.; Sun, L.; Qing, C.; Aung, P.P.; Clements, J.; Klokov, K.; Lappo, E.G.; Syroechkovskiy, E. The Spoon-billed Sandpiper Calidris pygmaea conservation project in 2019 and 2020: Population trends continue to be negative. BirdingASIA 2020, 33, 51–56. [Google Scholar]
- Chowdhury, S.U.; Foysal, M.; Green, R.E. Accelerating decline of an important wintering population of the critically endangered Spoon-billed Sandpiper Calidris pygmaea at Sonadia Island, Bangladesh. J. Ornithol. 2022, 163, 891–901. [Google Scholar] [CrossRef]
- Syroechkovskiy, E.E.; Lappo, E.G. Information from regions: Chukotka. Inf. Mater. Work. Group Waders 2002, 15, 17–18. (In Russian) [Google Scholar]
- Tomkovich, P.; Syroechkovskiy, E.; Yakushev, N.; Loktionov, E.; Digby, R.; Shepelev, I. Spoon-billed Sandpipers in Meinypilgyno: Update of the 2016 breeding season. Spoon-Bill. Sandpipers Task Force News Bull. 2016, 16, 13–15. [Google Scholar]
- Tomkovich, P.S. Maximum life longevity of some waders in Chukotka. Inf. Mater. Work. Group Waders 2003, 16, 55–56. [Google Scholar]
- Tomkovich, P.S. Breeding biology and breeding success of the Spoon-billed Sandpiper Eurynorhynchus pygmeus. Russ. J. Ornithol. 1995, 4, 77–91. [Google Scholar]
- Tomkovich, P.S. Spatial structure of the Spoon-billed Sandpiper (Eurynorhynchus pygmeus) population at the breeding grounds. In Modern Ornithology; Kurochkin, E.N., Ed.; Nauka: Moscow, Russia, 1994; pp. 130–148. [Google Scholar]
- Tomkovich, P.S.; Loktionov, E.Y. Age of first breeding of Spoon-billed Sandpipers Calidris pygmaea. Wader Study 2021, 128, 96–98. [Google Scholar] [CrossRef]
- Tomkovich, P.S. Migration of the Spoon-billed Sandpiper Eurynorhynchus pygmeus in the Far East of the Russian Federation. Stilt 1992, 21, 29–33. [Google Scholar]
- Tomkovich, P.S. External morphology of the Spoon-billed Sandpiper (Eurynorhynchus pygmeus) at Chukotski Peninsula. Ornithologia 1991, 25, 135–144. (In Russian) [Google Scholar]
- Cook, M.I.; Beissinger, S.R.; Toranzos, G.A.; Arendt, W.J. Incubation reduces microbial growth on eggshells and the opportunity for trans-shell infection. Ecol. Lett. 2005, 8, 532–537. [Google Scholar] [CrossRef]
- Clements, J.; Loktionov, E.Y.; Yakushev, N.; Clark, N.A.; Digby, R.; Jarrett, N.; Shepelev, I.; Tomkovich, P.S.; Green, R.E. Effect of age at release on the post-release survival of head-started Spoon-billed Sandpipers. Wader Study 2022, 129, 100–104. [Google Scholar] [CrossRef]
- Tomkovich, P.S.; Loktionov, E.Y.; Syroechkovskiy, E.E. Predators of wader nests in Southern Chukotka, Russia, as leant with camera traps. In Actual Issues of Wader Studies in Northern Eurasia, Proceedings of the XI International Scientific and Practical Conference, Minsk, Belarus, 29 January–2 February 2019; pp. 5–11. (In Russin). [Google Scholar]
- Li, J.; Lin, Z. Spoon-billed Sandpiper in Jiangsu Province, China and Korea in spring and summer. Spoon-Bill. Sandpiper Task Force News Bull. 2016, 16, 22–23. [Google Scholar]
- Moores, N.; Rogers, D.I.; Rogers, K.; Hansbro, P.M. Reclamation of tidal flats and shorebird declines in Saemangeum and elsewhere in the Republic of Korea. Emu 2016, 116, 136–146. [Google Scholar] [CrossRef]
- Ma, Z.; Melville, D.S.; Liu, J.; Chen, Y.; Yang, H.; Ren, W.; Zhang, Z.; Piersma, T.; Li, B. Rethinking China’s new great wall. Science 2014, 346, 912–914. [Google Scholar] [CrossRef]
- Heg, D.; Dingemanse, N.J.; Lessells, C.M.; Mateman, A.C. Parental Correlates of Offspring Sex Ratio in Eurasian Oystercatchers. Auk 2000, 117, 980–986. [Google Scholar] [CrossRef]
- Liker, A.; Freckleton, R.P.; Székely, T. The evolution of sex roles in birds is related to adult sex ratio. Nat. Commun. 2013, 4, 1587. [Google Scholar] [CrossRef]
- Riordan, M.M.; Lukacs, P.M.; Huyvaert, K.P.; Dreitz, V.J. Sex ratios of Mountain Plovers from egg production to fledging. Avian Conserv. Ecol. 2015, 10, 3. [Google Scholar] [CrossRef]
- Saunders, S.P.; Cuthbert, F.J. Chick mortality leads to male-biased sex ratios in endangered Great Lakes Piping Plovers. J. Field Ornithol. 2015, 86, 103–114. [Google Scholar] [CrossRef]
- Eberhart-Phillips, L.J.; Küpper, C.; Carmona-Isunza, M.C.; Vincze, O.; Zefania, S.; Cruz-López, M.; Kosztolányi, A.; Miller, T.E.X.; Barta, Z.; Cuthill, I.C.; et al. Demographic causes of adult sex ratio variation and their consequences for parental cooperation. Nat. Commun. 2018, 9, 1651. [Google Scholar] [CrossRef]
- Eberhart-Phillips, L.J.; Küpper, C.; Miller, T.E.X.; Cruz-López, M.; Maher, K.H.; dos Remedios, N.; Stoffel, M.A.; Hoffman, J.I.; Krüger, O.; Székely, T. Sex-specific early survival drives adult sex ratio bias in snowy plovers and impacts mating system and population growth. Proc. Natl. Acad. Sci. USA 2017, 114, E5474–E5481. [Google Scholar] [CrossRef]
- Loonstra, A.H.J.; Verhoeven, M.A.; Senner, N.R.; Hooijmeijer, J.C.E.W.; Piersma, T.; Kentie, R. Natal habitat and sex-specific survival rates result in a male-biased adult sex ratio. Behav. Ecol. 2019, 30, 843–851. [Google Scholar] [CrossRef] [PubMed]
- Hallgrimsson, G.T.; Palsson, S.; Summers, R.W.; Benediktsson, G.Ö. Sex ratio and sexual size dimorphism in Purple Sandpiper Calidris maritima chicks. Bird Study 2011, 58, 44–49. [Google Scholar] [CrossRef]
- Székely, T.; Cuthill, I.C.; Yezerinac, S.; Griffiths, R.; Kis, J. Brood sex ratio in the Kentish plover. Behav. Ecol. 2004, 15, 58–62. [Google Scholar] [CrossRef]
- Clark, J.A. Selective mortality of waders during severe weather. Bird Study 2009, 56, 96–102. [Google Scholar] [CrossRef]
- Meissner, W. Sex determination of juvenile Dunlins migrating through the Polish Baltic region. J. Field Ornithol. 2005, 76, 368–372. [Google Scholar] [CrossRef]
- Tomkovich, P.S.; Soloviev, M.Y. Numbers of the Spoon-billed Sandpiper Eurynorhynchus pygmeus at the north of Kolyuchinskaya Gulf, Chukotka, and count methods for the species on breeding grounds. Russ. J. Ornithol. 2000, 8, 3–10. [Google Scholar]
- Rönkä, N.; Pakanen, V.-M.; Pauliny, A.; Thomson, R.L.; Nuotio, K.; Pehlak, H.; Thorup, O.; Lehikoinen, P.; Rönkä, A.; Blomqvist, D.; et al. Genetic differentiation in an endangered and strongly philopatric, migrant shorebird. BMC Ecol. Evol. 2021, 21, 125. [Google Scholar] [CrossRef]
- Valdebenito, J.O.; Maher, K.H.; Zachár, G.; Huang, Q.; Zhang, Z.; Young, L.J.; Székely, T.; Que, P.; Liu, Y.; Urrutia, A.O. Sex differences in immune gene expression in the brain of a small shorebird. Immunogenetics 2022, 74, 487–496. [Google Scholar] [CrossRef]
- Galli, R.; Preusse, G.; Schnabel, C.; Bartels, T.; Cramer, K.; Krautwald-Junghanns, M.-E.; Koch, E.; Steiner, G. Sexing of chicken eggs by fluorescence and Raman spectroscopy through the shell membrane. PLoS ONE 2018, 132, e0192554. [Google Scholar] [CrossRef]
- Dutton, C.J.; Tieber, A. A Modified Protocol for Sex Identification of in Ovo Avian Embryos and Its Application as a Management TOOL for Endangered Species Conservation Programs. J. Zoo Wildl. Med. 2001, 32, 176–180. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year Species | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | 2021 | 2022 | Total |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Spoon-billed Sandpiper Calidris pygmaea | 66.7% | 62.5% | 50.0% | 50.0% | 50.0% | 52.9% | 41.7% | 41.7% | 55.6% | 57.1% | 51.4% |
| (n = 6) | (n = 8) | (n = 10) | (n = 12) | (n = 12) | (n = 17) | (n = 12) | (n = 12) | (n = 9) | (n = 7) | (n = 105) | |
| Other locally breeding waders [39] | 74.0% | 64.0% | 54.5% | 65.4% | 46.0% | 68.6% | 51.1% | 49.1% | 61.0% | 68.6% | 60.0% |
| (n = 46) | (n = 44) | (n = 55) | (n = 52) | (n = 50) | (n = 51) | (n = 47) | (n = 57) | (n = 59) | (n = 51) | (n = 512) |
| Year | 2011 * | 2012 * | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | 2021 | 2022 | Total |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HS | 80.0% | 75.7% | 87.5% | 87.1% | 87.9% | 86.1% | 83.3% | 71.4% | 68.4% | 90.0% | 74.4% | 0 | 80.5% |
| n = 25 | n = 37 | n = 24 | n = 31 | n = 33 | n = 36 | n = 36 | n = 35 | n = 38 | n = 30 | n = 39 | n = 364 | ||
| Wild | 100% | 100% | 85.7% | 85.7% | 84.6% | 92.9% | 72.7% | 91.7% | 94.7% | 94.4% | 78.9% | 86.7% | 88.2% |
| n = 4 | n = 4 | n = 7 | n = 21 | n = 13 | n = 14 | n = 11 | n = 24 | n = 19 | n = 18 | n = 19 | n = 15 | n = 169 | |
| Total | 82.8% | 78.0% | 87.1% | 86.5% | 87.0% | 88.0% | 80.9% | 79.7% | 77.2% | 91.7% | 75.9% | 86.7% | 82.9% |
| n = 29 | n = 41 | n = 31 | n = 52 | n = 46 | n = 50 | n = 47 | n = 59 | n = 57 | n = 48 | n = 57 | n = 15 | n = 533 |
| Year | Headstarting | Wild Chicks Marked | Birds Returned (HS/Wild) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Eggs Coll. | Chicks Hatched | Young Released | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | 2021 | 2022 | ||
| 2012 | 11 | 9 | 9 | 5 | 1/0 | 1/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 |
| 2013 | 24 | 21 | 18 | 3 | 0/0 | 3/1 | 2/1 | 0/1 | 0/1 | 0/0 | 0/1 | 0/1 | 0/1 |
| 2014 | 31 | 27 | 26 | 19 | – | 1/1 | 2/4 | 2/3 | 2/1 | 2/0 | 0/0 | 0/0 | 0/0 |
| 2015 | 33 | 29 | 28 | 12 | – | – | 1/0 | 4/3 | 1/3 | 1/1 | 0/1 | 1/1 | 0/1 |
| 2016 | 35 | 31 | 30 | 38 | – | – | – | 2/1 | 3/4 | 2/4 | 2/3 | 1/3 | 1/2 |
| 2017 | 38 * | 30 | 30 | 16 | – | – | – | – | 0/0 | 1/0 | 1/0 | 2/0 | 2/0 |
| 2018 | 35 | 25 | 22 | 30 | – | – | – | – | – | 0/1 | 0/3 | 1/3 | 0/0 |
| 2019 | 38 | 26 | 23 | 26 | – | – | – | – | – | – | 0/0 | 5/4 | 2/4 |
| 2020 | 30 | 27 | 22 | 20 | – | – | – | – | – | – | – | 0/0 | 2/1 |
| 2021 | 39 | 29 | 28 | 18 | – | – | – | – | – | – | – | – | 0/0 |
| 2022 | 0 | 0 | 0 | 13 | – | – | – | – | – | – | – | – | – |
| Total | 314 | 254 | 236 | 200 | 1/0 | 5/2 | 5/5 | 8/8 | 6/9 | 6/6 | 3/8 | 10/12 | 7/9 |
| Wild * | HS from Aviary | |||||
|---|---|---|---|---|---|---|
| ♂ | ♀ | ♂ + ♀ | ♂ | ♀ | ♂ + ♀ | |
| n | 11 | 4 | 15 | 10 | 9 | 19 |
| mean | 9251 | 5720 | 8309 | 5191 | 8833 | 6916 |
| s.d. | 10,866 | 4671 | 9572 | 7960 | 10,646 | 9249 |
| median | 4370 | 6002 | 4370 | 1781 | 3180 | 2502 |
| min | 42 | 146 | 42 | 650 | 763 | 650 |
| max | 30,594 | 10,730 | 30,594 | 26,216 | 26,846 | 26,846 |
| Eggs Produced | Chicks Hatched | Chicks Fledged | Mature Birds | Recruits in Natal Area | Successful First-Breeders | Successful Second-Breeders | |
|---|---|---|---|---|---|---|---|
| HS | 5.15 | 4.29 | 3.67 | 0.681 | 0.496 | 0.416 | 0.299 |
| Nature | 3.08 | 2.73 | 1.71 | 0.42 | 0.32 | 0.3 | 0.282 |
| Gain | 1.68 | 1.57 | 2.14 | 1.62 | 1.55 | 1.39 | 1.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loktionov, E.Y.; Digby, R.A.; Yakushev, N.N.; Shepelev, I.A.; Clements, J.P.; Tomkovich, P.S.; Jarrett, N.S.; Clark, N.A.; Green, R.E.; Lappo, E.G.; et al. Evaluating the Impact of Headstarting on the Critically Endangered Spoon-Billed Sandpiper Calidris pygmaea. Diversity 2023, 15, 584. https://doi.org/10.3390/d15040584
Loktionov EY, Digby RA, Yakushev NN, Shepelev IA, Clements JP, Tomkovich PS, Jarrett NS, Clark NA, Green RE, Lappo EG, et al. Evaluating the Impact of Headstarting on the Critically Endangered Spoon-Billed Sandpiper Calidris pygmaea. Diversity. 2023; 15(4):584. https://doi.org/10.3390/d15040584
Chicago/Turabian StyleLoktionov, Egor Y., Roland A. Digby, Nickolay N. Yakushev, Ivan A. Shepelev, Jodie P. Clements, Pavel S. Tomkovich, Nigel S. Jarrett, Nigel A. Clark, Rhys E. Green, Elena G. Lappo, and et al. 2023. "Evaluating the Impact of Headstarting on the Critically Endangered Spoon-Billed Sandpiper Calidris pygmaea" Diversity 15, no. 4: 584. https://doi.org/10.3390/d15040584
APA StyleLoktionov, E. Y., Digby, R. A., Yakushev, N. N., Shepelev, I. A., Clements, J. P., Tomkovich, P. S., Jarrett, N. S., Clark, N. A., Green, R. E., Lappo, E. G., & Syroechkovskiy, E. E. (2023). Evaluating the Impact of Headstarting on the Critically Endangered Spoon-Billed Sandpiper Calidris pygmaea. Diversity, 15(4), 584. https://doi.org/10.3390/d15040584