
Variation in the Jaw Musculature of Ratsnakes and Their Allies (Serpentes: Colubridae)



Abstract
:1. Introduction

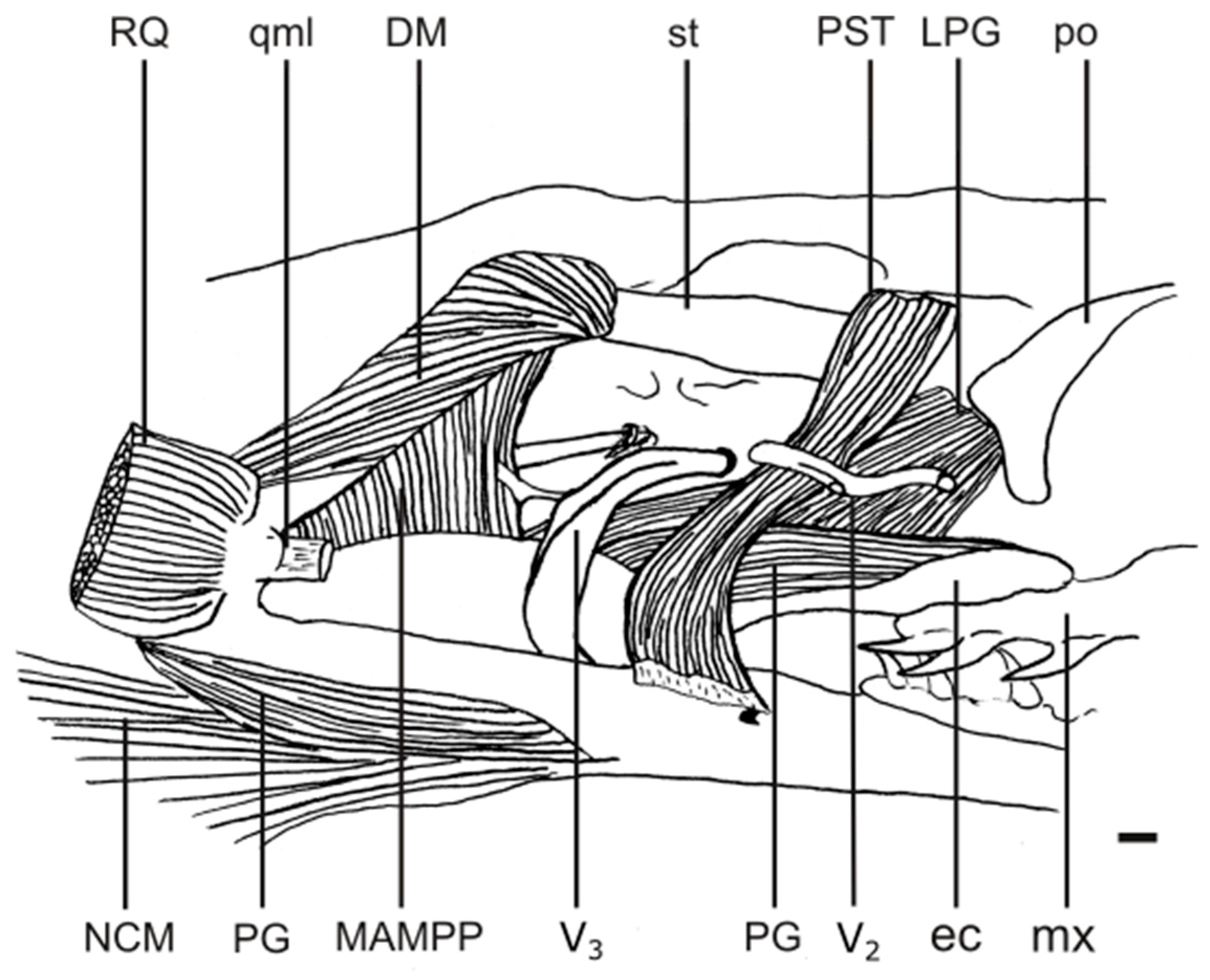
2. Materials and Methods
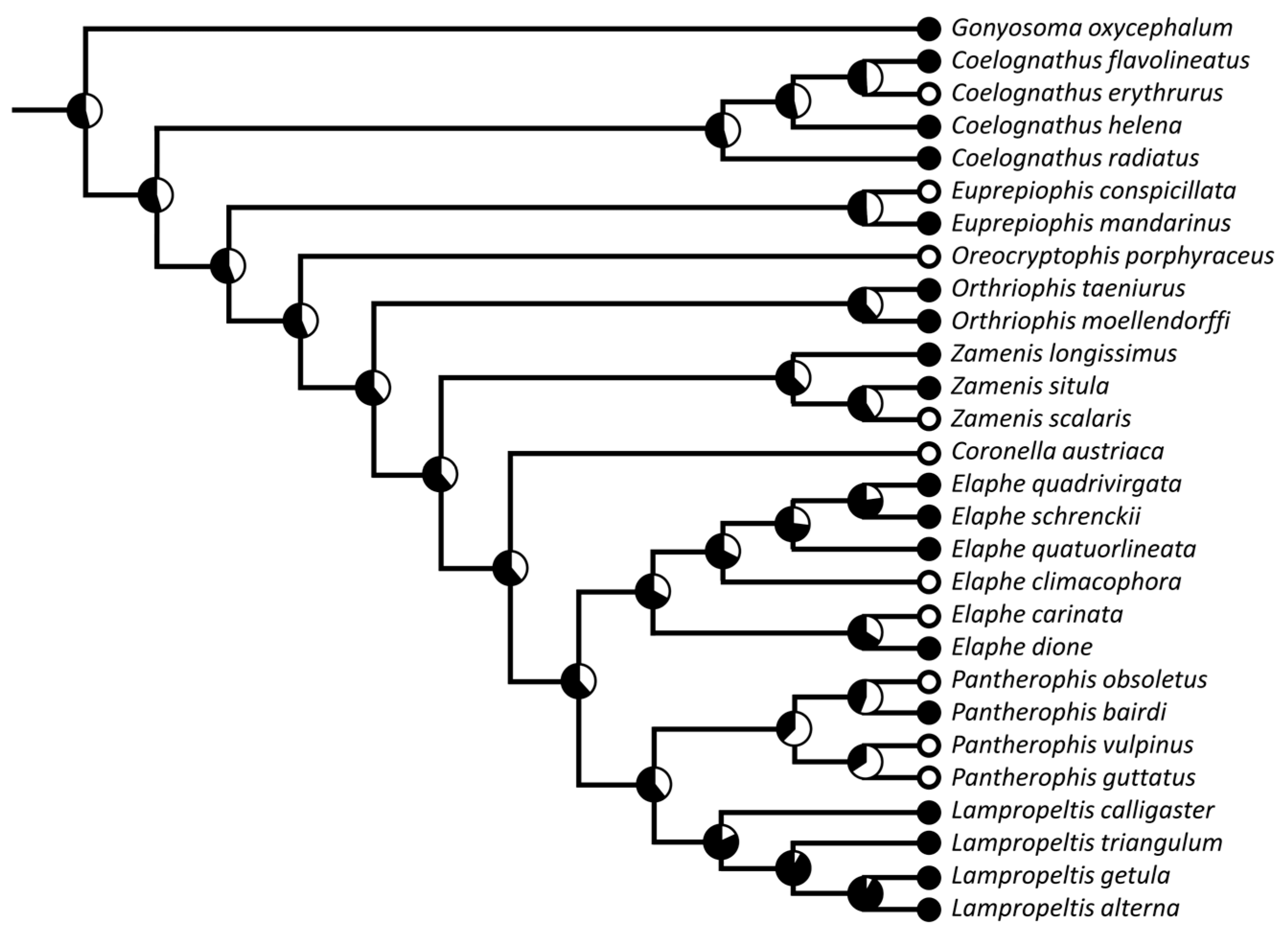
Phylogenetic Framework and Statistical Analyses
3. Results
3.1. Phylogenetic Signal and Ancestral State Reconstructions
3.2. Jaw Adductor Complex
3.2.1. Levator Anguli Oris (LAO)
3.2.2. Musculus Adductor Mandibulae Externus Superficialis (MAMES)
3.2.3. Musculus Adductor Mandibulae Externus Medialis (MAMEM)
3.2.4. Musculus Adductor Mandibulae Posterior Superficialis (MAMPS)
3.2.5. Musculus Adductor Mandibulae Posterior Profundus (MAMPP)
3.2.6. Musculus Pesudotemporalis (PST)
3.2.7. Musculus Pterygoideus (PG)
3.3. Dorsal Constrictors
3.3.1. Musculus Levator Pterygoidei (LPG)
3.3.2. Musculus Protractor Pterygoidei (PP)
3.3.3. Musculus Retractor Pterygoidei (RP)
3.3.4. Musculus Retractor Vomeris (RV)
3.3.5. Musculus Protractor Quadrati (PQ)
3.4. Intermandibular Musculature
3.4.1. Musculus Intermandibularis Anterior (MIA)
3.4.2. Musculus Intermandibularis Posterior (IP)
3.4.3. Musculus Transversus Branchialis (TB)
3.4.4. Musculus Geniotrachealis (GT)
3.4.5. Musculus Genioglossus (GG)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| a.l | aponeurosis lateralis |
| a.lao | aponeurosis M. levator anguli oris |
| a.mv | aponeurosis medioventralis |
| a.ncm | aponeurosis neurocostomandibularis |
| an | angular |
| bd | bodenaponeurosis |
| bo | basioccipital |
| bs | basisphenoid |
| cs | compound bone (os compositum) |
| DM | depressor mandibulae |
| dt | dentary |
| ec | ectopterygoid |
| GGCL | M. genioglossus, caput lateralis |
| GGCM | M. genioglossus, caput medialis |
| GT | M. geniotrachealis |
| hg | glandula Harderii |
| hy | hyoid |
| IA | M. intermandibularis anterior (pp. posterior et anterior) |
| IAA | M. intermandibularis anterior, pars anterior |
| IAG | M. intermandibularis anterior, pars glandularis |
| IAP | intermandibularis anterior, pars posterior |
| ilg | glandula infralabialis |
| IPA | M. intermandibularis posterior, pars anterior |
| IPP | M. intermandibularis posterior, pars posterior |
| LAO | M. levator anguli oris |
| LPG | M. levator pterygoidei |
| lsg | glandula sublingualis lateralis |
| MAMEM | M. adductor mandibulae externus medialis |
| MAMEP | M. adductor mandibulae externus profundus |
| MAMES | M. adductor mandibulae externus superficialis |
| MAMPP | M. adductor mandibulae posterior profundus |
| MAMPS | M. adductor mandibulae posterior superficialis |
| mx | maxilla |
| NCM | M. neurocostomandibularis |
| pa | parietal |
| pal | palatine |
| PG | M. pterygoideus |
| po | postorbital |
| PGA | M. pterygoideus accesorius |
| PP | M. protractor pterygoidei |
| PQ | M. protractor quadrati |
| pr | prootic |
| PST | M. pseudotemporalis |
| ptg | pterygoid |
| qa | quadrate aponeurosis |
| qml | ligamentum quadrato-maxillare |
| RP | M. retractor pterygoidei |
| RQ | M. retractor quadrati |
| RV | M. retractor vomeris |
| slg | glandula supralabialis |
| sp | splenial |
| st | supratemporal |
| TBCL | M. transversus branchialis, caput lateralis |
| TBPG | M. transversus branchialis, pars glandularis |
| V2 | nervus trigeminus (ramus maxillaris) |
| V3 | nervus trigeminus (ramus mandibularis) |
| BB | personal collection of Bartosz Borczyk |
| FMNH | Field Museum of Natural History, Chicago, IL, USA |
| IZK | Department of Evolutionary Biology and Conservation of Vertebrates, University of Wrocław, Wrocław, Poland |
| MNHN | Museum National d’Histoire Naturelle, Paris, France |
| MNHW | Museum of Natural History, University of Wrocław, Wrocław, Poland |
| ZFMK | Museum Koenig Bonn, Bonn, Germany |
| ZMB | Museum für Naturkunde, Berlin, Germany |
References
- Uetz, P.; Freed, P.; Aguilar, R.; Reyes, F.; Hošek, J. (Eds.) The Reptile Database. Available online: http://www.reptile-database.org (accessed on 28 April 2023).
- Groombridge, B.C. Comments on the intermandibular muscles of snakes. J. Nat. Hist. 1979, 13, 477–498. [Google Scholar] [CrossRef]
- Cundall, D. Functional Morphology. In Snakes: Ecology and Evolutionary Biology; Seigel, R.A., Collins, J.T., Novak, S.S., Eds.; The Blackburn Press: Caldwell, NJ, USA, 1987; pp. 106–140. [Google Scholar]
- Greene, H.W. Snakes: The Evolution of Mystery in Nature; University of California Press: Berkeley, CA, USA, 1997. [Google Scholar]
- Lee, M.S.Y.; Bell, G.L.; Caldwell, M.W. The origin of snake feeding. Nature 1999, 400, 655–659. [Google Scholar] [CrossRef]
- Cundall, D.; Greene, H.W. Feeding in snakes. In Feeding: Form, and Evolution in Tetrapod Vertebrates; Schwenk, K., Ed.; Academic Press: Cambridge, MA, USA, 2000; pp. 293–333. [Google Scholar]
- Cundall, D.; Irish, F. The snake skull. In Biology of the Reptilia; Gans, C., Gaunt, A.S., Adler, K., Eds.; Society for the Study of Amphibians and Reptiles: Ithaca, NY, USA, 2008; Volume 20, pp. 349–692. [Google Scholar]
- Haas, G. Muscles of the jaws and associated structures in the Rhynchocephalia and Squamata. In Biology of the Reptilia; Gans, C., Parsons, T.S., Eds.; Academic Press: Cambridge, MA, USA, 1973; Volume 4, pp. 285–490. [Google Scholar]
- Albright, G.B.; Nelson, E.M. Cranial kinetics of the generalized colubrid snake Elaphe obsoleta quadrivittata. I. Descriptive morphology. J. Morphol. 1959, 105, 193–239. [Google Scholar] [CrossRef] [PubMed]
- Kardong, K.V. Morphology of the respiratory system and its musculature in different snake genera. (Part I) Crotalus and Elaphe. Morphol. Jahrb. 1972, 117, 285–302. [Google Scholar]
- Groombridge, B.C. Variations in morphology of the superficial palate of henophidian snakes and some possible systematic implications. J. Nat. Hist. 1979, 13, 447–475. [Google Scholar] [CrossRef]
- Kardong, K.V. Jaw musculature of the West Indian snake Alsophis cantherigerus brooksi (Colubridae, Reptilia). Breviora 1980, 463, 1–26. [Google Scholar]
- Rieppel, O. The trigeminal jaw adductors of primitive snakes and their homologies with the lacertilian jaw adductors. J. Zool. 1980, 190, 447–471. [Google Scholar] [CrossRef]
- McDowell, S.B. The architecture of the corner of the mouth of colubroid snakes. J. Herpetol. 1986, 20, 353–407. [Google Scholar] [CrossRef]
- Cundall, D.; Rossman, D.E. Cephalic anatomy of the rare Indonesian snake Anomochilus weberi. Zool. J. Linn. Soc. 1993, 109, 235–273. [Google Scholar] [CrossRef]
- Zaher, H. Comments on the evolution of the jaw adductor musculature of snakes. Zool. J. Linn. Soc. 1994, 111, 339–384. [Google Scholar] [CrossRef]
- Das, S.; Pramanick, K. Comparative anatomy and homology of jaw adductor muscles of some South Asian colubroid snakes (Serpentes: Colubroidea). Vertebr. Zool. 2019, 69, 93–102. [Google Scholar]
- Daza, J.D.; Diogo, R.; Johnston, P.; Abdala, V. Jaw adductor muscles across Lepidosaurs: A reappraisal. Anat. Rec. 2011, 294, 1765–1782. [Google Scholar] [CrossRef]
- Johnston, P. Homology of the jaw muscles in lizards and snakes—A solution from a comparative gnathostome approach. Anat. Rec. 2014, 297, 574–585. [Google Scholar] [CrossRef]
- Diogo, R.; Abdala, V. Muscles of Vertebrates: Comparative Anatomy, Evolution, Homologies and Development; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar]
- Cowan, I.M.; Hick, W.B.M. A comparative study of the myology of the head region in three species of Thamnophis (Reptilia, Ophidia). Trans. R. Soc. Can. 1961, 45, 19–60. [Google Scholar]
- Weaver, W.G. The cranial anatomy of the hog-nosed snakes (Heterodon). Bull. Florida State Mus. 1965, 9, 275–304. [Google Scholar]
- Varkey, A. Comparative cranial myology of North American Natricinae snakes. Milwaukee Public Mus. Publ. Biol. Geol. 1979, 4, 1–70. [Google Scholar]
- Cundall, D. Variations of the cephalic muscles in the colubrid snake genera Entechinus, Opheodrys, Symphimus. J. Morphol. 1986, 187, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Utiger, U.; Schatti, B.; Helfenberger, N. The Oriental Colubrinae genus Coelognathus Fitzinger, 1843 and classification of Old and New World racers and ratsnakes (Reptilia, Squamata, Colubridae, Colubrinae). Russ. J. Herpetol. 2005, 12, 39–60. [Google Scholar]
- Burbrink, F.T.; Lawson, R. How and when did Old World ratsnakes disperse into New World? Mol. Phylogenet. Evol. 2009, 43, 173–189. [Google Scholar] [CrossRef] [PubMed]
- Pyron, R.A.; Burbrink, F.T.; Wiens, J.J. A phylogeny and revised classification of Squamata, including 4161 species of lizards and snakes. BMC Evol. Biol. 2013, 13, 93. [Google Scholar] [CrossRef]
- Chen, X.; McKelvy, A.D.; Grismer, L.L.; Matsui, M.; Nishikawa, K.; Burbrink, F.T. The phylogenetic position and taxonomic status of the Rainbow Tree Snake Gonyophis margaritatus (Peters, 1871) (Squamata: Colubridae). Zootaxa 2014, 3881, 532–548. [Google Scholar] [CrossRef] [PubMed]
- Zaher, H.; Murphy, R.W.; Arredondo, J.C.; Graboski, R.; Machado-Filho, P.R.; Mahlow, K.; Montingelli, G.G.; Quadros, A.B.; Orlov, N.L.; Wilkinson, M.; et al. Large-scale molecular phylogeny, morphology, divergence-time estimation, and the fossil record of advanced caenophidian snakes (Squamata: Serpentes). PLoS ONE 2019, 14, e0216148. [Google Scholar] [CrossRef]
- Schultz, K.-D. A Monograph of the Colubrid Snakes of the Genus Elaphe Fitzinger; Koeltz Scientific Books: Havlíčkův Brod, Czech Republic, 1996. [Google Scholar]
- Pyron, R.A.; Burbrink, F.T. Body size as a primary determinant of ecomorphological diversification and the evolution of mimicry in the lampropeltinine snakes (Serpentes: Colubridae). J. Evol. Biol. 2009, 22, 2057–2067. [Google Scholar] [CrossRef]
- Albright, G.B.; Nelson, E.M. Cranial kinetics of generalized colubrid snake Elaphe obsoleta quadrivittata. II. Functional morphology. J. Morphol. 1959, 105, 241–291. [Google Scholar] [CrossRef] [PubMed]
- Radovanović, M. Anatomische Studien am Schlangenkopf. Jena. Z. Naturwiss. 1935, 71, 179–312. [Google Scholar]
- Kumar, S.; Stecher, G.; Suleski, M.; Hedges, S.B. TimeTree: A resource for timelines, timetrees, and divergence dates. Mol. Biol. Evol. 2017, 34, 1812–1819. [Google Scholar] [CrossRef]
- Bock, W.; Shear, C.R. A staining method for gross dissection of vertebrate muscles. Anat. Anz. 1972, 130, 222–227. [Google Scholar]
- Oliveira, L.D.; Grazziotin, F.G.; Sánchez-Martínez, P.M.; Sasa, M.; Flores-Villela, O.; Prudente, A.L.D.C.; Zaher, H. Phylogenetic and morphological evidence reveals the association between diet and the evolution of the venom delivery system in Neotropical goo-eating snakes. Syst. Biodivers. 2023, 21, 2153944. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org/ (accessed on 28 April 2023).
- Paradis, E.; Schliep, K. ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef] [PubMed]
- Borges, R.; Machado, J.P.; Gomes, C.; Rocha, A.P.; Antunes, A. Measuring phylogenetic signal between categorical traits and phylogenies. Bioinformatics 2019, 35, 1862–1869. [Google Scholar] [CrossRef]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. Version 3.61 (build 927). Available online: http://www.mesquiteproject.org (accessed on 28 April 2023).
- Borczyk, B. The adductor mandibulae in Elaphe and related genera (Serpentes: Colubridae). In Herpetologia Bonnensis II., Proceedings of the 13th Congress of the Societas Europaea Herpetologica, Bonn, Germany, 27 September–2 October 2005; Vences, M., Köhler, J., Ziegler, T., Böhme, W., Eds.; Societas Europaea Herpetologica: Bonn, Germany, 2006; pp. 19–22. [Google Scholar]
- Smith, K.T. The evolution of mid-latitude faunas during the Eocene: Late Eocene lizards of the Medicine Pole Hills reconsidered. Bull. Peabody Mus. Nat. Hist. 2011, 52, 3–105. [Google Scholar] [CrossRef]
- Witmer, L.M. The Extant Phylogenetic Bracket and the importance of reconstructing soft tissue in fossils. In Functional Morphology in Vertebrate Paleontology; Thomason, J.J., Ed.; Cambridge University Press: Cambridge, UK, 1995; pp. 19–33. [Google Scholar]
- Young, B.A. The anatomy of the head of the red-sided garter snake, Thamnophis sirtalis parietalis. Part II: Myology. Zool. Jahrb. Abh. Anat. 1989, 118, 325–354. [Google Scholar]
- Elzanowski, A. Interconnection of muscles in the adductor mandibulae complex of birds. Ann. Anat. 1993, 1975, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Bell, C.J.; Mead, J.I. Not enough skeletons in the closet: Collection-based anatomical research in an age of conservation conscience. Anat. Rec. 2014, 297, 344–348. [Google Scholar] [CrossRef]
- Olori, J.C.; Bell, C.J. Comparative skull morphology of uropeltid snakes (Alethinophidia: Uropeltidae) with special reference to disarticulated bone elements and variation. PLoS ONE 2012, 7, e32450. [Google Scholar] [CrossRef]
- Cundall, D. Activity of head muscles during feeding by snakes: A comparative study. Am. Zool. 1983, 23, 383–396. [Google Scholar] [CrossRef]
- Pyron, R.A.; Burbrink, F.T. Early origin of viviparity and multiple reversions to oviparity in squamate reptiles. Ecol. Lett. 2014, 17, 13–21. [Google Scholar] [CrossRef]
























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Character | δ | p |
|---|---|---|
| M1 * | 2.239 | 0.01 |
| M2 | 0.327 | 0.4 |
| M3 * | 4.145 | 0.02 |
| M4 | 12.280 | 0.01 |
| M5 * | 1.003 | 0.58 |
| M6 | 0.8719 ** 1.380 *** | 0.17 ** 0.08 *** |
| M7 | 0.2348 | 0.38 |
| M8 | 0.7048 | 0.24 |
| M9 | 1.850 | - |
| M10 | 1.270 | - |
| M11 * | 2.626 | 0.03 |
| M12 | 0.7748 | 0.05 |
| M13 | 0.5753 | 0.1 |
| M14 * | 4.109 | 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borczyk, B.; Skawiński, T. Variation in the Jaw Musculature of Ratsnakes and Their Allies (Serpentes: Colubridae). Diversity 2023, 15, 628. https://doi.org/10.3390/d15050628
Borczyk B, Skawiński T. Variation in the Jaw Musculature of Ratsnakes and Their Allies (Serpentes: Colubridae). Diversity. 2023; 15(5):628. https://doi.org/10.3390/d15050628
Chicago/Turabian StyleBorczyk, Bartosz, and Tomasz Skawiński. 2023. "Variation in the Jaw Musculature of Ratsnakes and Their Allies (Serpentes: Colubridae)" Diversity 15, no. 5: 628. https://doi.org/10.3390/d15050628
APA StyleBorczyk, B., & Skawiński, T. (2023). Variation in the Jaw Musculature of Ratsnakes and Their Allies (Serpentes: Colubridae). Diversity, 15(5), 628. https://doi.org/10.3390/d15050628