
Biomonitoring for Watershed Protection from a Multiscale Land-Use Perspective

 , ,
, ,
Abstract
:1. Introduction
2. Biomonitoring in Different Freshwater Systems
2.1. Streams and Rivers
2.2. Lakes and Reservoirs
3. Biomonitoring the Effects of Different Land Uses on Freshwater Ecosystems
3.1. Agriculture
3.2. Urban and Industry
3.3. Mining
3.4. Forestry
3.5. Pasture
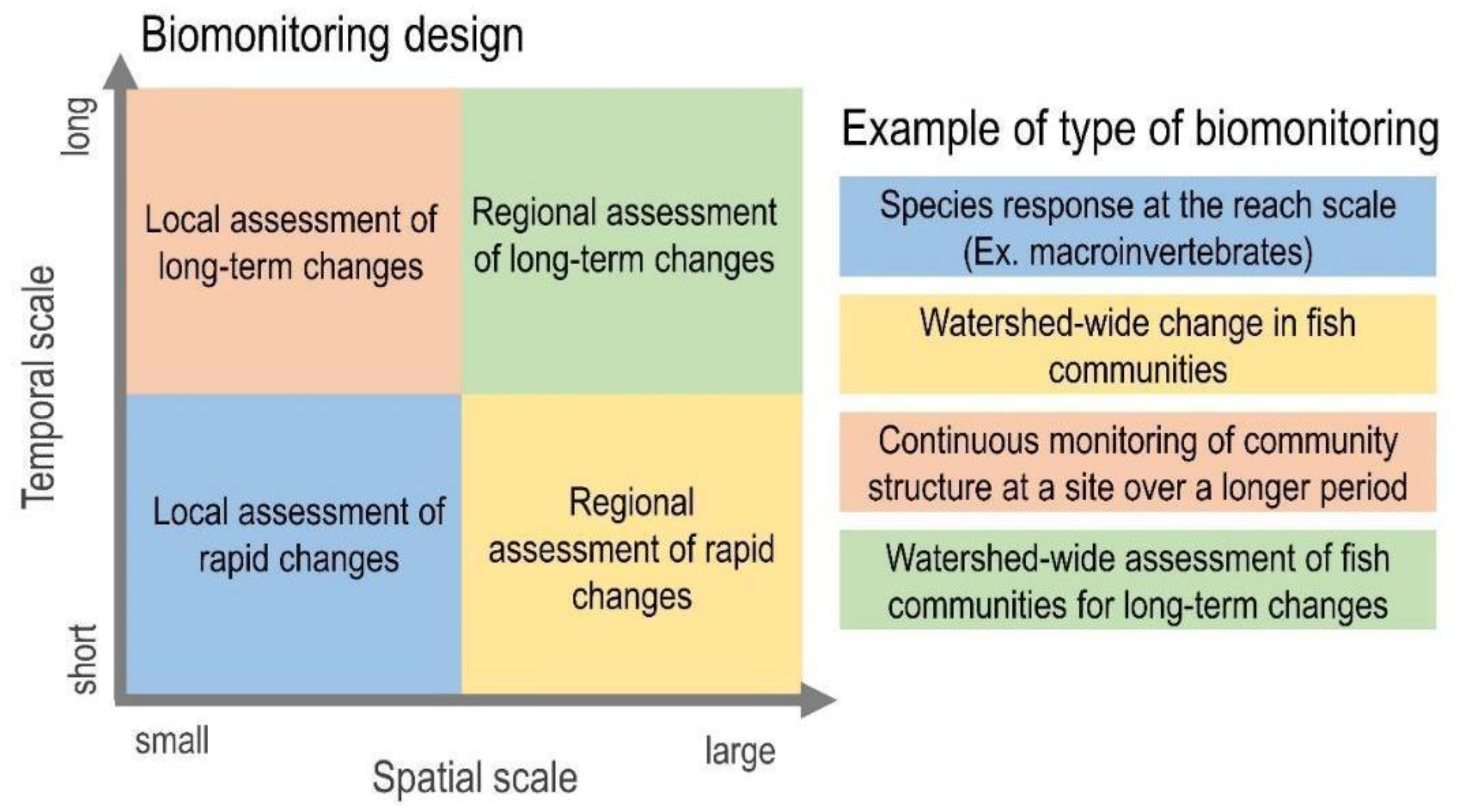
4. Multiscale Perspective of Biomonitoring
5. Planning Strategies and Future Predictions
5.1. How to Design Biomonitoring Programs
5.2. The Use of Models in Biomonitoring
5.3. Environmental DNA
5.4. Live-Cell Sensors
5.5. Stable Isotopes and Bioaccumulation
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Grill, G.; Lehner, B.; Thieme, M.; Geenen, B.; Tickner, D.; Antonelli, F.; Babu, S.; Borrelli, P.; Cheng, L.; Crochetiere, H.; et al. Mapping the World’s Free-Flowing Rivers. Nature 2019, 569, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Feio, M.J.; Hughes, R.M.; Serra, S.R.Q.; Nichols, S.J.; Kefford, B.J.; Lintermans, M.; Robinson, W.; Odume, O.N.; Callisto, M.; Macedo, D.R.; et al. Fish and Macroinvertebrate Assemblages Reveal Extensive Degradation of the World’s Rivers. Glob. Chang. Biol. 2023, 29, 355–374. [Google Scholar] [CrossRef]
- Li, F.; Zhang, Y.; Altermatt, F.; Zhang, X.; Cai, Y.; Yang, Z. Gap Analysis for DNA-Based Biomonitoring of Aquatic Ecosystems in China. Ecol. Indic. 2022, 137, 108732. [Google Scholar] [CrossRef]
- Ross, E.R.; Randhir, T.O. Effects of Climate and Land Use Changes on Water Quantity and Quality of Coastal Watersheds of Narragansett Bay. Sci. Total Environ. 2022, 807, 151082. [Google Scholar] [CrossRef]
- Vagheei, H.; Laini, A.; Vezza, P.; Palau-Salvador, G.; Boano, F. Ecohydrologic Modeling Using Nitrate, Ammonium, Phosphorus, and Macroinvertebrates as Aquatic Ecosystem Health Indicators of Albaida Valley (Spain). J. Hydrol. Reg. Stud. 2022, 42, 101155. [Google Scholar] [CrossRef]
- Vörösmarty, C.J.; McIntyre, P.B.; Gessner, M.O.; Dudgeon, D.; Prusevich, A.; Green, P.; Glidden, S.; Bunn, S.E.; Sullivan, C.A.; Liermann, C.R.; et al. Global Threats to Human Water Security and River Biodiversity. Nature 2010, 467, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Borges, A.C.P.; Piassão, J.F.G.; Albani, S.M.; Albertoni, E.F.; Martins, M.C.; Cansian, R.L.; Valduga, A.T.; Hepp, L.U.; Mielniczki-Pereira, A.A. Multiple Metals and Agricultural Use Affects Oxidative Stress Biomarkers in Freshwater Aegla Crabs. Braz. J. Biol. 2022, 82, 230147. [Google Scholar] [CrossRef]
- Mello, K.; Taniwaki, R.H.; de Paula, F.R.; Valente, R.A.; Randhir, T.O.; Macedo, D.R.; Leal, C.G.; Rodrigues, C.B.; Hughes, R.M. Multiscale Land Use Impacts on Water Quality: Assessment, Planning, and Future Perspectives in Brazil. J. Environ. Manag. 2020, 270, 110879. [Google Scholar] [CrossRef]
- Randhir, T.O.; Ekness, P. Water Quality Change and Habitat Potential in Riparian Ecosystems. Ecohydrol. Hydrobiol. 2013, 13, 192–200. [Google Scholar] [CrossRef]
- Rico-Sánchez, A.E.; Rodríguez-Romero, A.J.; Sedeño-Díaz, J.E.; López-López, E.; Sundermann, A. Aquatic Macroinvertebrate Assemblages in Rivers Influenced by Mining Activities. Sci. Rep. 2022, 12, 3209. [Google Scholar] [CrossRef]
- Randhir, T.O.; Hawes, A.G. Watershed Land Use and Aquatic Ecosystem Response: Ecohydrologic Approach to Conservation Policy. J. Hydrol. 2009, 364, 182–199. [Google Scholar] [CrossRef]
- Brooks, K.N.; Ffolliott, P.F.; Magner, J.A. Hydrology and the Management of Watersheds, 4th ed.; Wiley-Blackwell: Ames, IA, USA, 2012; ISBN 9780470963050. [Google Scholar]
- Calizza, E.; Favero, F.; Rossi, D.; Careddu, G.; Fiorentino, F.; Sporta Caputi, S.; Rossi, L.; Costantini, M.L. Isotopic Biomonitoring of N Pollution in Rivers Embedded in Complex Human Landscapes. Sci. Total Environ. 2020, 706, 136081. [Google Scholar] [CrossRef] [PubMed]
- Arini, A.; Feurtet-Mazel, A.; Morin, S.; Maury-Brachet, R.; Coste, M.; Delmas, F. Remediation of a Watershed Contaminated by Heavy Metals: A 2-Year Field Biomonitoring of Periphytic Biofilms. Sci. Total Environ. 2012, 425, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Garris, H.W.; Baldwin, S.A.; Taylor, J.; Gurr, D.B.; Denesiuk, D.R.; Van Hamme, J.D.; Fraser, L.H. Short-Term Microbial Effects of a Large-Scale Mine-Tailing Storage Facility Collapse on the Local Natural Environment. PLoS ONE 2018, 13, e0196032. [Google Scholar] [CrossRef]
- Isibor, P.O.; Imoobe, T.O.T.; Dedeke, G.A.; Adagunodo, T.A.; Taiwo, O.S. Health Risk Indices and Zooplankton-Based Assessment of a Tropical Rainforest River Contaminated with Iron, Lead, Cadmium, and Chromium. Sci. Rep. 2020, 10, 16896. [Google Scholar] [CrossRef]
- Rimet, F.; Bouchez, A. Biomonitoring River Diatoms: Implications of Taxonomic Resolution. Ecol. Indic. 2012, 15, 92–99. [Google Scholar] [CrossRef]
- Lenart-Boroń, A.; Boroń, P.; Kulik, K.; Prajsnar, J.; Żelazny, M.; Chmiel, M.J. Anthropogenic Pollution Gradient along a Mountain River Affects Bacterial Community Composition and Genera with Potential Pathogenic Species. Sci. Rep. 2022, 12, 18140. [Google Scholar] [CrossRef]
- Persaud, A.D.; Paterson, A.M.; Dillon, P.J.; Winter, J.G.; Palmer, M.; Somers, K.M. Forecasting Cyanobacteria Dominance in Canadian Temperate Lakes. J. Environ. Manag. 2015, 151, 343–352. [Google Scholar] [CrossRef]
- Jabiol, J.; Chauvet, E.; Guérold, F.; Bouquerel, J.; Usseglio-Polatera, P.; Artigas, J.; Margoum, C.; Le Dréau, M.; Moreira, A.; Mazzella, N.; et al. The Combination of Chemical, Structural, and Functional Indicators to Evaluate the Anthropogenic Impacts on Agricultural Stream Ecosystems. Environ. Sci. Pollut. Res. 2022, 29, 29296–29313. [Google Scholar] [CrossRef]
- Qu, Y.; Wu, N.; Guse, B.; Makarevičiūtė, K.; Sun, X.; Fohrer, N. Riverine Phytoplankton Functional Groups Response to Multiple Stressors Variously Depending on Hydrological Periods. Ecol. Indic. 2019, 101, 41–49. [Google Scholar] [CrossRef]
- Gomes, A.C.A.M.; Gomes, L.F.; Roitman, I.; Pereira, H.R.; Junior, A.F.C.; da Costa, E.M.M.; da Silva, M.L.C.; Jacobson, T.K.B.; da Costa Ribeiro, R.J.; de Miranda Filho, R.J.; et al. Forest Cover Influences Zooplanktonic Communities in Amazonian Streams. Aquat. Ecol. 2020, 54, 1067–1078. [Google Scholar] [CrossRef]
- Brase, R.A.; Schwab, H.E.; Li, L.; Spink, D.C. Elevated Levels of Per- and Polyfluoroalkyl Substances (PFAS) in Freshwater Benthic Macroinvertebrates from the Hudson River Watershed. Chemosphere 2022, 291, 132830. [Google Scholar] [CrossRef]
- Edegbene, A.O. Assessing the Health of Forested Riverine Systems in the Niger Delta Area of Nigeria: A Macroinvertebrate-Based Multimetric Index Approach. Environ. Sci. Pollut. Res. 2022, 29, 15068–15080. [Google Scholar] [CrossRef]
- Dal Pont, G.; Duarte Ritter, C.; Agostinis, A.O.; Stica, P.V.; Horodesky, A.; Cozer, N.; Balsanelli, E.; Netto, O.S.M.; Henn, C.; Ostrensky, A.; et al. Monitoring Fish Communities through Environmental DNA Metabarcoding in the Fish Pass System of the Second Largest Hydropower Plant in the World. Sci. Rep. 2021, 11, 23167. [Google Scholar] [CrossRef]
- Johnson, S.L.; Ringler, N.H. The Response of Fish and Macroinvertebrate Assemblages to Multiple Stressors: A Comparative Analysis of Aquatic Communities in a Perturbed Watershed (Onondaga Lake, NY). Ecol. Indic. 2014, 41, 198–208. [Google Scholar] [CrossRef]
- Shah, Z.U.; Parveen, S. Oxidative, Biochemical and Histopathological Alterations in Fishes from Pesticide Contaminated River Ganga, India. Sci. Rep. 2022, 12, 3628. [Google Scholar] [CrossRef]
- Resh, V.H. Which Group Is Best? Attributes of Different Biological Assemblages Used in Freshwater Biomonitoring Programs. Environ. Monit. Assess. 2008, 138, 131–138. [Google Scholar] [CrossRef]
- Li, D.; Pi, J.; Zhang, T.; Tan, X.; Fraser, D.J. Evaluating a 5-Year Metal Contamination Remediation and the Biomonitoring Potential of a Freshwater Gastropod along the Xiangjiang River, China. Environ. Sci. Pollut. Res. 2018, 25, 21127–21137. [Google Scholar] [CrossRef]
- Blackman, R.C.; Ho, H.C.; Walser, J.C.; Altermatt, F. Spatio-Temporal Patterns of Multi-Trophic Biodiversity and Food-Web Characteristics Uncovered across a River Catchment Using Environmental DNA. Commun. Biol. 2022, 5, 259. [Google Scholar] [CrossRef]
- Emilson, C.E.; Thompson, D.G.; Venier, L.A.; Porter, T.M.; Swystun, T.; Chartrand, D.; Capell, S.; Hajibabaei, M. DNA Metabarcoding and Morphological Macroinvertebrate Metrics Reveal the Same Changes in Boreal Watersheds across an Environmental Gradient. Sci. Rep. 2017, 7, 12777. [Google Scholar] [CrossRef]
- Messager, M.L.; Davies, I.P.; Levin, P.S. Low-Cost Biomonitoring and High-Resolution, Scalable Models of Urban Metal Pollution. Sci. Total Environ. 2021, 767, 144280. [Google Scholar] [CrossRef]
- Tiegs, S.D.; Costello, D.M.; Isken, M.W.; Woodward, G.; Mcintyre, P.B.; Gessner, M.O.; Chauvet, E.; Griffiths, N.A.; Flecker, A.S.; Acuña, V.; et al. Global Patterns and Drivers of Ecosystem Functioning in Rivers and Riparian Zones. Sci. Adv. 2019, 5, eaav0486. [Google Scholar] [CrossRef]
- Dodds, W.K.; Gido, K.; Whiles, M.R.; Daniels, M.D.; Grudzinski, B.P. The Stream Biome Gradient Concept: Factors Controlling Lotic Systems across Broad Biogeographic Scales. Freshw. Sci. 2015, 34, 1. [Google Scholar] [CrossRef]
- Poff, N.L.; Allan, J.D.; Bain, M.B.; Karr, J.R.; Prestegaard, K.L.; Richter, B.D.; Sparks, R.E.; Stromberg, J.C. The Natural Flow Regime. Bioscience 1997, 47, 769–784. [Google Scholar] [CrossRef]
- Anbumozhi, V.; Radhakrishnan, J.; Yamaji, E. Impact of Riparian Buffer Zones on Water Quality and Associated Management Considerations. Ecol. Eng. 2005, 24, 517–523. [Google Scholar] [CrossRef]
- Auble, G.T.; Friedman, J.M.; Scott, M.L. Relating Riparian Vegetation to Present and Future Streamflows. Ecol. Appl. 1994, 4, 544–554. [Google Scholar] [CrossRef]
- Boyero, L.; Pearson, R.G.; Dudgeon, D.; Ferreira, V.; Graca, M.A.S.; Gessner, M.O.; Boulton, A.J.; Chauvet, E.; Yule, C.M.; Albarino, R.J.; et al. Global Patterns of Stream Detritivore Distribution: Implications for Biodiversity Loss in Changing Climates. Glob. Ecol. Biogeogr. 2012, 21, 134–141. [Google Scholar] [CrossRef]
- Goncalves, J.F., Jr.; Callisto, M. Organic-Matter Dynamics in the Riparian Zone of a Tropical Headwater Stream in Southern Brasil. Aquat. Bot. 2013, 109, 8–13. [Google Scholar] [CrossRef]
- Tonin, A.M.; Gonçalves, J.F.; Bambi, P.; Couceiro, S.R.M.; Feitoza, L.A.M.; Fontana, L.E.; Hamada, N.; Hepp, L.U.; Lezan-Kowalczuk, V.G.; Leite, G.F.M.; et al. Plant Litter Dynamics in the Forest-Stream Interface: Precipitation Is a Major Control across Tropical Biomes. Sci. Rep. 2017, 7, 10799. [Google Scholar] [CrossRef]
- Paola, C.; Foufoula-Georgiou, E.; Dietrich, W.E.; Hondzo, M.; Mohrig, D.; Parker, G.; Power, M.E.; Rodriguez-Iturbe, I.; Voller, V.; Wilcock, P. Toward a Unified Science of the Earth’s Surface: Opportunities for Synthesis among Hydrology, Geomorphology, Geochemistry, and Ecology. Water Resour. Res. 2006, 42, 2336. [Google Scholar] [CrossRef]
- Limburg, K.E.; Swaney, D.P.; Strayer, D.L. River Ecosystems. In Encyclopedia of Biodiversity, 2nd ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2013; pp. 469–484. ISBN 9780123847195. [Google Scholar]
- Buss, D.F.; Carlisle, D.M.; Chon, T.-S.; Culp, J.; Harding, J.S.; Keizer-Vlek, H.E.; Robinson, W.A.; Strachan, S.; Thirion, C.; Hughes, R.M. Stream Biomonitoring Using Macroinvertebrates around the Globe: A Comparison of Large-Scale Programs. Environ. Monit. Assess. 2015, 187, 4132. [Google Scholar] [CrossRef]
- Vadas, R.L.; Hughes, R.M.; Bae, Y.J.; Baek, M.J.; Gonzáles, O.C.B.; Callisto, M.; de Carvalho, D.R.; Chen, K.; Ferreira, M.T.; Fierro, P.; et al. Assemblage-Based Biomonitoring of Freshwater Ecosystem Health via Multimetric Indices: A Critical Review and Suggestions for Improving Their Applicability. Water Biol. Secur. 2022, 1, 100054. [Google Scholar] [CrossRef]
- Hering, D.; Feld, C.K.; Moog, O.; Ofenböck, T. Cook Book for the Development of a Multimetric Index for Biological Condition of Aquatic Ecosystems: Experiences from the European AQEM and STAR Projects and Related Initiatives. In The Ecological Status of European Rivers: Evaluation and Intercalibration of Assessment Methods; Springer: Dordrecht, The Netherlands, 2006; pp. 311–324. [Google Scholar]
- Cooke, G.D.; Welch, E.B.; Peterson, S.A.; Newroth, P.R. Lake and Reservoir Restoration; Elsevier: Amsterdam, The Netherlands, 1986; ISBN 9780250406432. [Google Scholar]
- Menezes, S.; Baird, D.J.; Soares, A.M.V.M. Beyond Taxonomy: A Review of Macroinvertebrate Trait-Based Community Descriptors as Tools for Freshwater Biomonitoring. J. Appl. Ecol. 2010, 47, 711–719. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, X.; Xie, Y.; Song, C.; Zhang, Y.; Yu, H.; Burton, G.A. Zooplankton Community Profiling in a Eutrophic Freshwater Ecosystem-Lake Tai Basin by DNA Metabarcoding. Sci. Rep. 2017, 7, 1773. [Google Scholar] [CrossRef]
- Van den Broeck, M.; Waterkeyn, A.; Rhazi, L.; Grillas, P.; Brendonck, L. Assessing the Ecological Integrity of Endorheic Wetlands, with Focus on Mediterranean Temporary Ponds. Ecol. Indic. 2015, 54, 1–11. [Google Scholar] [CrossRef]
- Odountan, O.H.; Janssens de Bisthoven, L.; Abou, Y.; Eggermont, H. Biomonitoring of Lakes Using Macroinvertebrates: Recommended Indices and Metrics for Use in West Africa and Developing Countries. Hydrobiologia 2019, 826, 1–23. [Google Scholar] [CrossRef]
- Palma, P.; Matos, C.; Alvarenga, P.; Köck-Schulmeyer, M.; Simões, I.; Barceló, D.; López de Alda, M.J. Ecological and Ecotoxicological Responses in the Assessment of the Ecological Status of Freshwater Systems: A Case-Study of the Temporary Stream Brejo of Cagarrão (South of Portugal). Sci. Total Environ. 2018, 634, 394–406. [Google Scholar] [CrossRef]
- Marin, V.; Arranz, I.; Grenouillet, G.; Cucherousset, J. Fish Size Spectrum as a Complementary Biomonitoring Approach of Freshwater Ecosystems. Ecol. Indic. 2023, 146, 109833. [Google Scholar] [CrossRef]
- Metcalfe, J.L.; Hayton, A. Comparison of Leeches and Mussels as Biomonitors for Chlorophenol Pollution. J. Great Lakes Res. 1989, 15, 654–668. [Google Scholar] [CrossRef]
- Poikane, S.; Johnson, R.K.; Sandin, L.; Schartau, A.K.; Solimini, A.G.; Urbanič, G.; Arbačiauskas, K.; Aroviita, J.; Gabriels, W.; Miler, O.; et al. Benthic Macroinvertebrates in Lake Ecological Assessment: A Review of Methods, Intercalibration and Practical Recommendations. Sci. Total Environ. 2016, 543, 123–134. [Google Scholar] [CrossRef]
- Gerritsen, J.; Carlson, R.E.; Dycus, D.L.; Faulkner, C.; Gibson, G.R.; Harcum, J.; Markowitz, S.A. Lake and Reservoir Bioassessment and Biocriteria: Technical Guidance Document; USEPA Office of Water: Washington, DC, USA, 1998.
- de Paula, F.R.; Leal, C.G.; Leitão, R.P.; Ferraz, S.F.D.B.; Pompeu, P.S.; Zuanon, J.A.S.; Hughes, R.M. The Role of Secondary Riparian Forests for Conserving Fish Assemblages in Eastern Amazon Streams. Hydrobiologia 2022, 849, 4529–4546. [Google Scholar] [CrossRef]
- Macaulay, S.J.; Buchwalter, D.B.; Matthaei, C.D. Water Temperature Interacts with the Insecticide Imidacloprid to Alter Acute Lethal and Sublethal Toxicity to Mayfly Larvae. N. Z. J. Mar. Freshw. Res. 2020, 54, 115–130. [Google Scholar] [CrossRef]
- Piggott, J.J.; Lange, K.; Townsend, C.R.; Matthaei, C.D. Multiple Stressors in Agricultural Streams: A Mesocosm Study of Interactions among Raised Water Temperature, Sediment Addition and Nutrient Enrichment. PLoS ONE 2012, 7, e49873. [Google Scholar] [CrossRef]
- Piggott, J.J.; Niyogi, D.K.; Townsend, C.R.; Matthaei, C.D. Multiple Stressors and Stream Ecosystem Functioning: Climate Warming and Agricultural Stressors Interact to Affect Processing of Organic Matter. J. Appl. Ecol. 2015, 52, 1126–1134. [Google Scholar] [CrossRef]
- Egler, M.; Buss, D.F.; Moreira, J.C.; Baptista, D.F. Influence of Agricultural Land-Use and Pesticides on Benthic Macroinvertebrate Assemblages in an Agricultural River Basin in Southeast Brazil. Braz. J. Biol. 2012, 72, 437–443. [Google Scholar] [CrossRef]
- Maltby, L.; Hills, L. Spray Drift of Pesticides and Stream Macroinvertebrates: Experimental Evidence of Impacts and Effectiveness of Mitigation Measures. Environ. Pollut. 2008, 156, 1112–1120. [Google Scholar] [CrossRef]
- Schäfer, R.B.; Caquet, T.; Siimes, K.; Mueller, R.; Lagadic, L.; Liess, M. Effects of Pesticides on Community Structure and Ecosystem Functions in Agricultural Streams of Three Biogeographical Regions in Europe. Sci. Total Environ. 2007, 382, 272–285. [Google Scholar] [CrossRef]
- Helson, J.E.; Williams, D.D. Development of a Macroinvertebrate Multimetric Index for the Assessment of Low-Land Streams in the Neotropics. Ecol. Indic. 2013, 29, 167–178. [Google Scholar] [CrossRef]
- Sutherland, A.B.; Culp, J.M.; Benoy, G.A. Evaluation of Deposited Sediment and Macroinvertebrate Metrics Used to Quantify Biological Response to Excessive Sedimentation in Agricultural Streams. Environ. Manag. 2012, 50, 50–63. [Google Scholar] [CrossRef]
- Taniwaki, R.H.; Piggott, J.J.; Ferraz, S.F.B.; Matthaei, C.D. Climate Change and Multiple Stressors in Small Tropical Streams. Hydrobiologia 2017, 793, 41–53. [Google Scholar] [CrossRef]
- Vitecek, S.; Johnson, R.; Poikane, S. Assessing the Ecological Status of European Rivers and Lakes Using Benthic Invertebrate Communities: A Practical Catalogue of Metrics and Methods. Water 2021, 13, 346. [Google Scholar] [CrossRef]
- Debenest, T.; Pinelli, E.; Coste, M.; Silvestre, J.; Mazzella, N.; Madigou, C.; Delmas, F. Sensitivity of Freshwater Periphytic Diatoms to Agricultural Herbicides. Aquat. Toxicol. 2009, 93, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Fore, L.S.; Grafe, C. Using Diatoms to Assess the Biological Condition of Large Rivers in Idaho (U.S.A.). Freshw.iol. 2002, 47, 2015–2037. [Google Scholar] [CrossRef]
- Schneck, F.; Schwarzbold, A.; Melo, A.S. Substrate Roughness Affects Stream Benthic Algal Diversity, Assemblage Composition, and Nestedness. J. N. Am. Benthol. Soc. 2011, 30, 1049–1056. [Google Scholar] [CrossRef]
- Taniwaki, R.H.; Matthaei, C.D.; Cardoso, T.K.M.; Ferraz, S.F.B.; Martinelli, L.A.; Piggott, J.J. The Effect of Agriculture on the Seasonal Dynamics and Functional Diversity of Benthic Biofilm in Tropical Headwater Streams. Biotropica 2019, 51, 18–27. [Google Scholar] [CrossRef]
- Mangadze, T.; Bere, T.; Mwedzi, T. Choice of Biota in Stream Assessment and Monitoring Programs in Tropical Streams: A Comparison of Diatoms, Macroinvertebrates and Fish. Ecol. Indic. 2016, 63, 128–143. [Google Scholar] [CrossRef]
- Edegbene, A.O.; Akamagwuna, F.C.; Odume, O.N.; Arimoro, F.O.; Edegbene Ovie, T.T.; Akumabor, E.C.; Ogidiaka, E.; Kaine, E.A.; Nwaka, K.H. A Macroinvertebrate-Based Multimetric Index for Assessing Ecological Condition of Forested Stream Sites Draining Nigerian Urbanizing Landscapes. Sustainability 2022, 14, 11289. [Google Scholar] [CrossRef]
- Gebrehiwot, M.; Awoke, A.; Beyene, A.; Kifle, D.; Triest, L. Macroinvertebrate Community Structure and Feeding Interactions along a Pollution Gradient in Gilgel Gibe Watershed, Ethiopia: Implications for Biomonitoring. Limnologica 2017, 62, 68–76. [Google Scholar] [CrossRef]
- Grabicová, K.; Vojs Staňová, A.; Švecová, H.; Nováková, P.; Kodeš, V.; Leontovyčová, D.; Brooks, B.W.; Grabic, R. Invertebrates Differentially Bioaccumulate Pharmaceuticals: Implications for Routine Biomonitoring. Environ. Pollut. 2022, 309, 119715. [Google Scholar] [CrossRef]
- Park, C.B.; Kim, G.E.; Kim, D.W.; Kim, S.; Yeom, D.H. Biomonitoring the Effects of Urban-Stream Waters on the Health Status of Pale Chub (Zacco Platypus): A Comparative Analysis of Biological Indexes and Biomarker Levels. Ecotoxicol. Environ. Saf. 2021, 208, 111452. [Google Scholar] [CrossRef]
- McGrane, S.J. Impacts of Urbanisation on Hydrological and Water Quality Dynamics, and Urban Water Management: A Review. Hydrol. Sci. J. 2016, 61, 2295–2311. [Google Scholar] [CrossRef]
- Amoatey, P.; Baawain, M.S. Effects of Pollution on Freshwater Aquatic Organisms. Water Environ. Res. 2019, 91, 1272–1287. [Google Scholar] [CrossRef] [PubMed]
- Hughes, R.M.; Dunham, S.; Maas-Hebner, K.G.; Yeakley, J.A.; Schreck, C.; Harte, M.; Molina, N.; Shock, C.C.; Kaczynski, V.W.; Schaeffer, J. A Review of Urban Water Body Challenges and Approaches: (1) Rehabilitation and Remediation. Fisheries 2014, 39, 18–29. [Google Scholar] [CrossRef]
- Feio, M.J.; Hughes, R.M.; Callisto, M.; Nichols, S.J.; Odume, O.N.; Quintella, B.R.; Kuemmerlen, M.; Aguiar, F.C.; Almeida, S.F.P.; Alonso-EguíaLis, P.; et al. The Biological Assessment and Rehabilitation of the World’s Rivers: An Overview. Water 2021, 13, 371. [Google Scholar] [CrossRef]
- Callisto, M.; Massara, R.L.; Linares, M.S.; Hughes, R.M. Benthic Macroinvertebrate Assemblages Detect the Consequences of a Sewage Spill: A Case Study of a South American Environmental Challenge. Limnology 2022, 23, 181–194. [Google Scholar] [CrossRef]
- Macedo, D.R.; Callisto, M.; Linares, M.S.; Hughes, R.M.; Romano, B.M.L.; Rothe-Neves, M.; Silveira, J.S. Urban Stream Rehabilitation in a Densely Populated Brazilian Metropolis. Front. Environ. Sci. 2022, 10, 921934. [Google Scholar] [CrossRef]
- Horton, A.A.; Walton, A.; Spurgeon, D.J.; Lahive, E.; Svendsen, C. Microplastics in Freshwater and Terrestrial Environments: Evaluating the Current Understanding to Identify the Knowledge Gaps and Future Research Priorities. Sci. Total Environ. 2017, 586, 127–141. [Google Scholar] [CrossRef]
- França, J.S.; Solar, R.; Hughes, R.M.; Callisto, M. Student Monitoring of the Ecological Quality of Neotropical Urban Streams. Ambio 2019, 48, 867–878. [Google Scholar] [CrossRef]
- Krabbenhoft, C.A.; Kashian, D.R. Citizen Science Data Are a Reliable Complement to Quantitative Ecological Assessments in Urban Rivers. Ecol. Indic. 2020, 116, 106476. [Google Scholar] [CrossRef]
- Bonnail, E.; Sarmiento, A.M.; DelValls, T.A.; Nieto, J.M.; Riba, I. Assessment of Metal Contamination, Bioavailability, Toxicity and Bioaccumulation in Extreme Metallic Environments (Iberian Pyrite Belt) Using Corbicula Fluminea. Sci. Total Environ. 2016, 544, 1031–1044. [Google Scholar] [CrossRef]
- Albuquerque, F.E.A.; Herrero-Latorre, C.; Miranda, M.; Barrêto Júnior, R.A.; Oliveira, F.L.C.; Sucupira, M.C.A.; Ortolani, E.L.; Minervino, A.H.H.; López-Alonso, M. Fish Tissues for Biomonitoring Toxic and Essential Trace Elements in the Lower Amazon. Environ. Pollut. 2021, 283, 117024. [Google Scholar] [CrossRef] [PubMed]
- Condini, M.V.; Pichler, H.A.; de Oliveira-Filho, R.R.; Cattani, A.P.; Andrades, R.; Vilar, C.C.; Joyeux, J.-C.; Soeth, M.; De Biasi, J.B.; Eggertsen, L.; et al. Marine Fish Assemblages of Eastern Brazil: An Update after the World’s Largest Mining Disaster and Suggestions of Functional Groups for Biomonitoring Long-Lasting Effects. Sci. Total Environ. 2022, 807, 150987. [Google Scholar] [CrossRef] [PubMed]
- de Almeida, P.I.N.; de Jesus, H.E.; Pereira, P.H.F.; Vieira, C.E.D.; Bianchini, A.; Martins, C.D.M.G.; dos Santos, H.F. The Microbial Profile of Rivers and Lagoons Three Years after the Impact of the World’s Largest Mining Disaster (Fundão Dam, Brazil). Environ. Res. 2023, 216, 114710. [Google Scholar] [CrossRef] [PubMed]
- Bevitório, L.Z.; da Silva, N.G.; Pirovani, J.C.M.; Marques, J.A.; Vieira, C.E.D.; Zebral, Y.D.; Damasceno, E.M.; Lopes, L.L.R.; de Sant’Ana, L.P.; Marube, L.C.; et al. Impacts of Tailings of Fundão Dam (Brazil) Rupture on Marine Fish: Metals Bioaccumulation and Physiological Responses. Mar. Pollut. Bull. 2022, 177, 113511. [Google Scholar] [CrossRef] [PubMed]
- de Matos, A.R.; da Silva, H.T.; da Faria, M.C.S.; Freire, B.M.; Pereira, R.M.; Batista, B.L.; Rodrigues, J.L. Environmental Disaster in Mining Areas: Routes of Exposure to Metals in the Doce River Basin. Int. J. Environ. Sci. Technol. 2022, 19, 12091–12102. [Google Scholar] [CrossRef]
- Maraschi, A.C.; Marques, J.A.; Costa, S.R.; Vieira, C.E.D.; Geihs, M.A.; Costa, P.G.; Martins, C.d.M.G.; Sandrini, J.Z.; Bianchini, A.; Souza, M.M. Marine Shrimps as Biomonitors of the Fundão (Brazil) Mine Dam Disaster: A Multi-Biomarker Approach. Environ. Pollut. 2022, 305, 119245. [Google Scholar] [CrossRef]
- Rodrigues, C.B.; Taniwaki, R.H.; Lane, P.; de Paula Lima, W.; de Barros Ferraz, S.F. Eucalyptus Short-Rotation Management Effects on Nutrient and Sediments in Subtropical Streams. Forests 2019, 10, 519. [Google Scholar] [CrossRef]
- van Dijk, A.I.J.M.; Keenan, R.J. Planted Forests and Water in Perspective. For. Ecol. Manag. 2007, 251, 1–9. [Google Scholar] [CrossRef]
- Trayler, K.M.; Davis, J.A. Forestry Impacts and the Vertical Distribution of Stream Invertebrates in South-Western Australia. Freshw. Biol. 1998, 40, 331–342. [Google Scholar] [CrossRef]
- USDA—United States Department of Agriculture. Best Management Practices to Minimize Agricultural Phosphorus Impacts on Water Quality; USDA: Washington, DC, USA, 2006.
- Vowell, J.L.; Frydenborg, R.B. A Biological Assessment of Best Management Practice Effectiveness during Intensive Silviculture and Forest Chemical Application. Water Air Soil Pollut. Focus 2004, 4, 297–307. [Google Scholar] [CrossRef]
- Erdozain, M.; Kidd, K.A.; Emilson, E.J.S.; Capell, S.S.; Luu, T.; Kreutzweiser, D.P.; Gray, M.A. Forest Management Impacts on Stream Integrity at Varying Intensities and Spatial Scales: Do Biological Effects Accumulate Spatially? Sci. Total Environ. 2021, 763, 144043. [Google Scholar] [CrossRef] [PubMed]
- Prudente, B.S.; Pompeu, P.S.; Juen, L.; Montag, L.F.A. Effects of Reduced-Impact Logging on Physical Habitat and Fish Assemblages in Streams of Eastern Amazonia. Freshw. Biol. 2017, 62, 303–316. [Google Scholar] [CrossRef]
- Dias, M.S.; Magnusson, W.E.; Zuanon, J. Effects of Reduced-Impact Logging on Fish Assemblages in Central Amazonia. Conserv. Biol. 2010, 24, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Mello, K.D.; Valente, R.A.; Randhir, T.O.; dos Santos, A.C.A.; Vettorazzi, C.A. Effects of Land Use and Land Cover on Water Quality of Low-Order Streams in Southeastern Brazil: Watershed versus Riparian Zone. Catena 2018, 167, 130–138. [Google Scholar] [CrossRef]
- Hepp, L.U.; Santos, S. Benthic Communities of Streams Related to Different Land Uses in a Hydrographic Basin in Southern Brazil. Environ. Monit. Assess. 2009, 157, 305–318. [Google Scholar] [CrossRef]
- Sunohara, M.D.; Topp, E.; Wilkes, G.; Gottschall, N.; Neumann, N.; Ruecker, N.; Jones, T.H.; Edge, T.A.; Marti, R.; Lapen, D.R. Impact of Riparian Zone Protection from Cattle on Nutrient, Bacteria, F-Coliphage, Cryptosporidium, and Giardia Loading of an Intermittent Stream. J. Environ. Qual. 2012, 41, 1301–1314. [Google Scholar] [CrossRef]
- Griffith, M.B.; McManus, M.G. Consideration of Spatial and Temporal Scales in Stream Restorations and Biotic Monitoring to Assess Restoration Outcomes: A Literature Review, Part 1. River Res. Appl. 2020, 36, 1385–1397. [Google Scholar] [CrossRef]
- Hermoso, V.; Linke, S.; Prenda, J.; Possingham, H.P. Addressing Longitudinal Connectivity in the Systematic Conservation Planning of Fresh Waters. Freshw. Biol. 2011, 56, 57–70. [Google Scholar] [CrossRef]
- Leal, C.G.; Pompeu, P.S.; Gardner, T.A.; Leitão, R.P.; Hughes, R.M.; Kaufmann, P.R.; Zuanon, J.; de Paula, F.R.; Ferraz, S.F.B.; Thomson, J.R.; et al. Multi-Scale Assessment of Human-Induced Changes to Amazonian Instream Habitats. Landsc. Ecol. 2016, 31, 1725–1745. [Google Scholar] [CrossRef]
- Brejão, G.L.; Hoeinghaus, D.J.; Pérez-Mayorga, M.A.; Ferraz, S.F.B.; Casatti, L. Threshold Responses of Amazonian Stream Fishes to Timing and Extent of Deforestation. Conserv. Biol. 2018, 32, 860–871. [Google Scholar] [CrossRef]
- Green, R.H. Sampling Design and Statistical Methods for Environmental Biologists; John Wiley & Sons: Hoboken, NJ, USA, 1979. [Google Scholar]
- Wright, J.; Sutcliffe, D.; Furse, M. Assessing the Biological Quality of Fresh Water: RIVPACS and Other Techniques; Freshwater Biological Association: Cumbria, UK, 2000. [Google Scholar]
- Bailey, R.; Norris, R.; Reynoldson, T. Bioassessment of Freshwater Ecosystems Using the Reference Condition Approach; Kluwer Academic Publishers: Boston, UK, 2004. [Google Scholar]
- Ward, J.V. The Four-Dimensional Nature of Lotic Ecosystems. J. North Am. Benthol. Soc. 1989, 8, 2–8. [Google Scholar] [CrossRef]
- Allan, D.; Erickson, D.; Fay, J. The Influence of Catchment Land Use on Stream Integrity across Multiple Spatial Scales. Freshw. Biol. 1997, 37, 149–161. [Google Scholar] [CrossRef]
- Altermatt, F. Diversity in Riverine Metacommunities: A Network Perspective. Aquat. Ecol. 2013, 47, 365–377. [Google Scholar] [CrossRef]
- Heino, J. The Importance of Metacommunity Ecology for Environmental Assessment Research in the Freshwater Realm. Biol. Rev. 2013, 88, 166–178. [Google Scholar] [CrossRef]
- Paulsen, S.G.; Peck, D.V.; Kaufmann, P.R.; Herlihy, A.T. Rivers and Streams: Upgrading Monitoring of the Nation’s Freshwater Resources—Meeting the Spirit of the Clean Water Act. In Water Quality—Science, Assessments and Policy; IntechOpen: Rijeka, Croatia, 2020. [Google Scholar]
- Poikane, S.; Zampoukas, N.; Borja, A.; Davies, S.P.; van de Bund, W.; Birk, S. Intercalibration of Aquatic Ecological Assessment Methods in the European Union: Lessons Learned and Way Forward. Environ. Sci. Policy 2014, 44, 237–246. [Google Scholar] [CrossRef]
- Clapcott, J.E.; Goodwin, E.O.; Snelder, T.H.; Collier, K.J.; Neale, M.W.; Greenfield, S. Finding Reference: A Comparison of Modelling Approaches for Predicting Macroinvertebrate Community Index Benchmarks. N. Z. J. Mar. Freshw. Res. 2017, 51, 44–59. [Google Scholar] [CrossRef]
- Lee, J.-Y.; Kwon, K. Current Status of Groundwater Monitoring Networks in Korea. Water 2016, 8, 168. [Google Scholar] [CrossRef]
- Reynolds, J.H.; Knutson, M.G.; Newman, K.B.; Silverman, E.D.; Thompson, W.L. A Road Map for Designing and Implementing a Biological Monitoring Program. Environ. Monit. Assess. 2016, 188, 399. [Google Scholar] [CrossRef] [PubMed]
- Barbour, C.D.M.T.; Gerritsen, J.; Snyder, B.D.; Stribling, J.B. Rapid Bioassessment Protocols for Use in Streams and Wadeable Rivers: Periphyton, Benthic Macroinvertebrates and Fish, 2nd ed.; Environmental Protection Agency, Office of Water: Washington, DC, USA, 1999.
- Chessman, B.C. What’s Wrong with the Australian River Assessment System (AUSRIVAS)? Mar. Freshw. Res. 2021, 72, 1110–1117. [Google Scholar] [CrossRef]
- De Souza, N.F.; Baptista, D.F.; Buss, D.F. A Predictive Index Based on Environmental Filters for the Bioassessment of River Basins without Reference Areas in Atlantic Forest Biome, Brazil. Biota Neotrop. 2019, 19, e20180601. [Google Scholar] [CrossRef]
- Feio, M.J.; Poquet, J.M. Predictive Models for Freshwater Biological Assessment: Statistical Approaches, Biological Elements and the Iberian Peninsula Experience: A Review. Int. Rev. Hydrobiol. 2011, 96, 321–346. [Google Scholar] [CrossRef]
- Waite, I.R.; Van Metre, P.C. Multistressor Predictive Models of Invertebrate Condition in the Corn Belt, USA. Freshw. Sci. 2017, 36, 901–914. [Google Scholar] [CrossRef]
- Stoddard, J.L.; Larsen, D.P.; Hawking, C.P.; Johnson, R.K.; Norris, R.H. Setting Expectations for the Ecological Condition of Streams: The Concept of Reference Condition. Ecol. Appl. 2006, 16, 1267–1276. [Google Scholar] [CrossRef]
- Ambelu, A.; Mekonen, S.; Koch, M.; Addis, T.; Boets, P.; Everaert, G.; Goethals, P. The Application of Predictive Modelling for Determining Bio-Environmental Factors Affecting the Distribution of Blackflies (Diptera: Simuliidae) in the Gilgel Gibe Watershed in Southwest Ethiopia. PLoS ONE 2014, 9, e112221. [Google Scholar] [CrossRef] [PubMed]
- Gezie, A.; Mulat, W.L.; Anteneh, W.; Dejen, E.; Kloos, H.; Mereta, S.T. Habitat Suitability Modelling of Benthic Macroinvertebrate Community in Wetlands of Lake Tana Watershed, Northwest Ethiopia. Wetlands 2020, 40, 853–864. [Google Scholar] [CrossRef]
- Feio, M.J.; Viana-Ferreira, C.; Costa, C. Testing a Multiple Machine Learning Tool (HYDRA) for the Bioassessment of Fresh Waters. Freshw. Sci. 2014, 33, 1286–1296. [Google Scholar] [CrossRef]
- Leitão, R.P.; Zuanon, J.; Mouillot, D.; Leal, C.G.; Hughes, R.M.; Kaufmann, P.R.; Villéger, S.; Pompeu, P.S.; Kasper, D.; de Paula, F.R.; et al. Disentangling the Pathways of Land Use Impacts on the Functional Structure of Fish Assemblages in Amazon Streams. Ecography 2018, 41, 219–232. [Google Scholar] [CrossRef]
- Waite, I.R.; Brown, L.R.; Kennen, J.G.; May, J.T.; Cuffney, T.F.; Orlando, J.L.; Jones, K.A. Comparison of Watershed Disturbance Predictive Models for Stream Benthic Macroinvertebrates for Three Distinct Ecoregions in Western US. Ecol. Indic. 2010, 10, 1125–1136. [Google Scholar] [CrossRef]
- Keith, D.A.; Akçakaya, H.R.; Thuiller, W.; Midgley, G.F.; Pearson, R.G.; Phillips, S.J.; Regan, H.M.; Araújo, M.B.; Rebelo, T.G. Predicting Extinction Risks under Climate Change: Coupling Stochastic Population Models with Dynamic Bioclimatic Habitat Models. Biol. Lett. 2008, 4, 560–563. [Google Scholar] [CrossRef]
- Ruaro, R.; Gubiani, É.A.; Hughes, R.M.; Mormul, R.P. Global Trends and Challenges in Multimetric Indices of Biological Condition. Ecol. Indic. 2020, 110, 105862. [Google Scholar] [CrossRef]
- Karr, J.R. Assessment of Biotic Integrity Using Fish Communities. Fisheries 1981, 6, 21–27. [Google Scholar] [CrossRef]
- Ndiritu, G.G.; Terer, T.; Njoroge, P.; Muiruri, V.M.; Njagi, E.L.; Kosgei, G.; Njoroge, L.; Kamau, P.W.; Malonza, P.K.; Muchane, M.; et al. Using the Biological Condition Gradient Model as a Bioassessment Framework to Support Rehabilitation and Restoration of the Upper Tana River Watershed in Kenya. Front. Environ. Sci. 2021, 9, 671051. [Google Scholar] [CrossRef]
- De Oliveira, R.B.D.S.; Mugnai, R.; Pereira, P.D.S.; De Souza, N.F.; Baptista, D.F. A Predictive Multimetric Index Based on Macroinvetebrates for Atlantic Forest Wadeable Streams Assessment. Biota Neotrop. 2019, 19, e20180541. [Google Scholar] [CrossRef]
- Martins, I.; Macedo, D.R.; Hughes, R.M.; Callisto, M. Major Risks to Aquatic Biotic Condition in a Neotropical Savanna River Basin. River Res. Appl. 2021, 37, 858–868. [Google Scholar] [CrossRef]
- Silva, D.R.O.; Herlihy, A.T.; Hughes, R.M.; Macedo, D.R.; Callisto, M. Assessing the Extent and Relative Risk of Aquatic Stressors on Stream Macroinvertebrate Assemblages in the Neotropical Savanna. Sci. Total Environ. 2018, 633, 179–188. [Google Scholar] [CrossRef]
- USEPA—United States Environmental Protection Agency. The Biological Condition Gradient (BCG): A Model for Interpreting Anthropogenic Stress on the Aquatic Environment; USEPA: Washington, DC, USA, 2021.
- Mello, K.D.; Randhir, T.O.; Valente, R.A.; Vettorazzi, C.A. Riparian Restoration for Protecting Water Quality in Tropical Agricultural Watersheds. Ecol. Eng. 2017, 108, 514–524. [Google Scholar] [CrossRef]
- Blackman, R.C.; Osathanunkul, M.; Brantschen, J.; Di Muri, C.; Harper, L.R.; Mächler, E.; Hänfling, B.; Altermatt, F. Mapping Biodiversity Hotspots of Fish Communities in Subtropical Streams through Environmental DNA. Sci. Rep. 2021, 11, 10375. [Google Scholar] [CrossRef]
- Carraro, L.; Mächler, E.; Wüthrich, R.; Altermatt, F. Environmental DNA Allows Upscaling Spatial Patterns of Biodiversity in Freshwater Ecosystems. Nat. Commun. 2020, 11, 3585. [Google Scholar] [CrossRef]
- Carvalho, D.C.; Leal, C.G. Steps Forward in Biomonitoring 2.0: EDNA Metabarcoding and Community-level Modelling Allow the Assessment of Complex Drivers of Neotropical Fish Diversity. Glob. Chang. Biol. 2023, 29, 1688–1690. [Google Scholar] [CrossRef]
- Zizka, V.M.A.; Geiger, M.F.; Leese, F. DNA Metabarcoding of Stream Invertebrates Reveals Spatio-Temporal Variation but Consistent Status Class Assessments in a Natural and Urban River. Ecol. Indic. 2020, 115, 106383. [Google Scholar] [CrossRef]
- Pérez-Burillo, J.; Trobajo, R.; Vasselon, V.; Rimet, F.; Bouchez, A.; Mann, D.G. Evaluation and Sensitivity Analysis of Diatom DNA Metabarcoding for WFD Bioassessment of Mediterranean Rivers. Sci. Total Environ. 2020, 727, 138445. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Zhang, S.; Xu, S.; Xiong, P.; Cao, Y.; Chen, Z.; Li, M. Comparison of Environmental DNA Metabarcoding and Bottom Trawling for Detecting Seasonal Fish Communities and Habitat Preference in a Highly Disturbed Estuary. Ecol. Indic. 2023, 146, 109754. [Google Scholar] [CrossRef]
- Skelton, J.; Cauvin, A.; Hunter, M.E. Environmental DNA Metabarcoding Read Numbers and Their Variability Predict Species Abundance, but Weakly in Non-Dominant Species. Environ. DNA 2022, 1–13. [Google Scholar] [CrossRef]
- Deiner, K.; Fronhofer, E.A.; Mächler, E.; Walser, J.C.; Altermatt, F. Environmental DNA Reveals That Rivers Are Conveyer Belts of Biodiversity Information. Nat. Commun. 2016, 7, 12544. [Google Scholar] [CrossRef] [PubMed]
- Seymour, M.; Edwards, F.K.; Cosby, B.J.; Bista, I.; Scarlett, P.M.; Brailsford, F.L.; Glanville, H.C.; de Bruyn, M.; Carvalho, G.R.; Creer, S. Environmental DNA Provides Higher Resolution Assessment of Riverine Biodiversity and Ecosystem Function via Spatio-Temporal Nestedness and Turnover Partitioning. Commun. Biol. 2021, 4, 512. [Google Scholar] [CrossRef] [PubMed]
- Mächler, E.; Walser, J.; Altermatt, F. Decision-making and Best Practices for Taxonomy-free Environmental DNA Metabarcoding in Biomonitoring Using Hill Numbers. Mol. Ecol. 2021, 30, 3326–3339. [Google Scholar] [CrossRef]
- Wlodkowic, D.; Karpiński, T.M. Live-Cell Systems in Real-Time Biomonitoring of Water Pollution: Practical Considerations and Future Perspectives. Sensors 2021, 21, 7028. [Google Scholar] [CrossRef]
- Gu, M.B.; Mitchell, R.J.; Kim, B.C. Whole-Cell-Based Biosensors for Environmental Biomonitoring and Application. In Biomanufacturing. Advances in Biochemical Engineering; Zhong, J.J., Ed.; Springer: Berlin/Heidelberg, Germany, 2004; Volume 87. [Google Scholar]
- Eltzov, E.; Marks, R.S. Whole-Cell Aquatic Biosensors. Anal. Bioanal. Chem. 2011, 400, 895–913. [Google Scholar] [CrossRef]
- Dalu, T.; Cuthbert, R.N.; Taylor, J.C.; Magoro, M.L.; Weyl, O.L.F.; Froneman, P.W.; Wasserman, R.J. Benthic Diatom-Based Indices and Isotopic Biomonitoring of Nitrogen Pollution in a Warm Temperate Austral River System. Sci. Total Environ. 2020, 748, 142452. [Google Scholar] [CrossRef]
- de Carvalho, D.R.; Alves, C.B.M.; Pompeu, P.S. Uncertainty in Estimating Fish Trophic Positions and Food Web Structure in Highly Polluted River Basins. Environ. Biol. Fishes 2022, 105, 119–137. [Google Scholar] [CrossRef]
- de Carvalho, D.R.; Flecker, A.S.; Alves, C.B.M.; Sparks, J.P.; Pompeu, P.S. Trophic Responses to Aquatic Pollution of Native and Exotic Livebearer Fishes. Sci. Total Environ. 2019, 681, 503–515. [Google Scholar] [CrossRef] [PubMed]
- Morrissey, C.A.; Boldt, A.; Mapstone, A.; Newton, J.; Ormerod, S.J. Stable Isotopes as Indicators of Wastewater Effects on the Macroinvertebrates of Urban Rivers. Hydrobiologia 2013, 700, 231–244. [Google Scholar] [CrossRef]
- Sekabira, K.; Origa, H.O.; Basamba, T.A.; Mutumba, G.; Kakudidi, E. Application of Algae in Biomonitoring and Phytoextraction of Heavy Metals Contamination in Urban Stream Water. Int. J. Environ. Sci. Technol. 2011, 8, 115–128. [Google Scholar] [CrossRef]
- Yan, Z.; Chen, Y.; Bao, X.; Zhang, X.; Ling, X.; Lu, G.; Liu, J.; Nie, Y. Microplastic Pollution in an Urbanized River Affected by Water Diversion: Combining with Active Biomonitoring. J. Hazard. Mater. 2021, 417, 126058. [Google Scholar] [CrossRef]
- Bertoli, M.; Pastorino, P.; Lesa, D.; Renzi, M.; Anselmi, S.; Prearo, M.; Pizzul, E. Microplastics Accumulation in Functional Feeding Guilds and Functional Habit Groups of Freshwater Macrobenthic Invertebrates: Novel Insights in a Riverine Ecosystem. Sci. Total Environ. 2022, 804, 150207. [Google Scholar] [CrossRef]


{kind=link}
| Land Use Type | Main Stressors |
|---|---|
| Agriculture | Sedimentation, nutrient loading, changes in pH, increased temperature range (due to deforestation), and the increased runoff of pesticides and herbicides |
| Urban and industry | Industrial effluents, domestic wastewaters, the complex mixture of pollutants from urban drainage, heavy metals, temperature range increases, microplastics, increased nutrient loading, and changes in pH |
| Mining | Metal contamination from mining processes; sedimentation |
| Forestry | Fluctuations in water yield and nutrient load, changes in pH, pest control, and increased sediment inputs from harvesting |
| Pasture | Fertilizer runoff, organic matter loads, increased temperature range, and sedimentation |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Mello, K.; Taniwaki, R.H.; Macedo, D.R.; Leal, C.G.; Randhir, T.O. Biomonitoring for Watershed Protection from a Multiscale Land-Use Perspective. Diversity 2023, 15, 636. https://doi.org/10.3390/d15050636
de Mello K, Taniwaki RH, Macedo DR, Leal CG, Randhir TO. Biomonitoring for Watershed Protection from a Multiscale Land-Use Perspective. Diversity. 2023; 15(5):636. https://doi.org/10.3390/d15050636
Chicago/Turabian Stylede Mello, Kaline, Ricardo Hideo Taniwaki, Diego Rodrigues Macedo, Cecília Gontijo Leal, and Timothy O. Randhir. 2023. "Biomonitoring for Watershed Protection from a Multiscale Land-Use Perspective" Diversity 15, no. 5: 636. https://doi.org/10.3390/d15050636
APA Stylede Mello, K., Taniwaki, R. H., Macedo, D. R., Leal, C. G., & Randhir, T. O. (2023). Biomonitoring for Watershed Protection from a Multiscale Land-Use Perspective. Diversity, 15(5), 636. https://doi.org/10.3390/d15050636