
Rediscovery of Five Rinodina Species Originally Described from Southwest China and One New Species

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Morphological and Chemical Study
2.2. DNA Extraction, Purification, and Sequencing
2.3. Phylogenetic Analysis
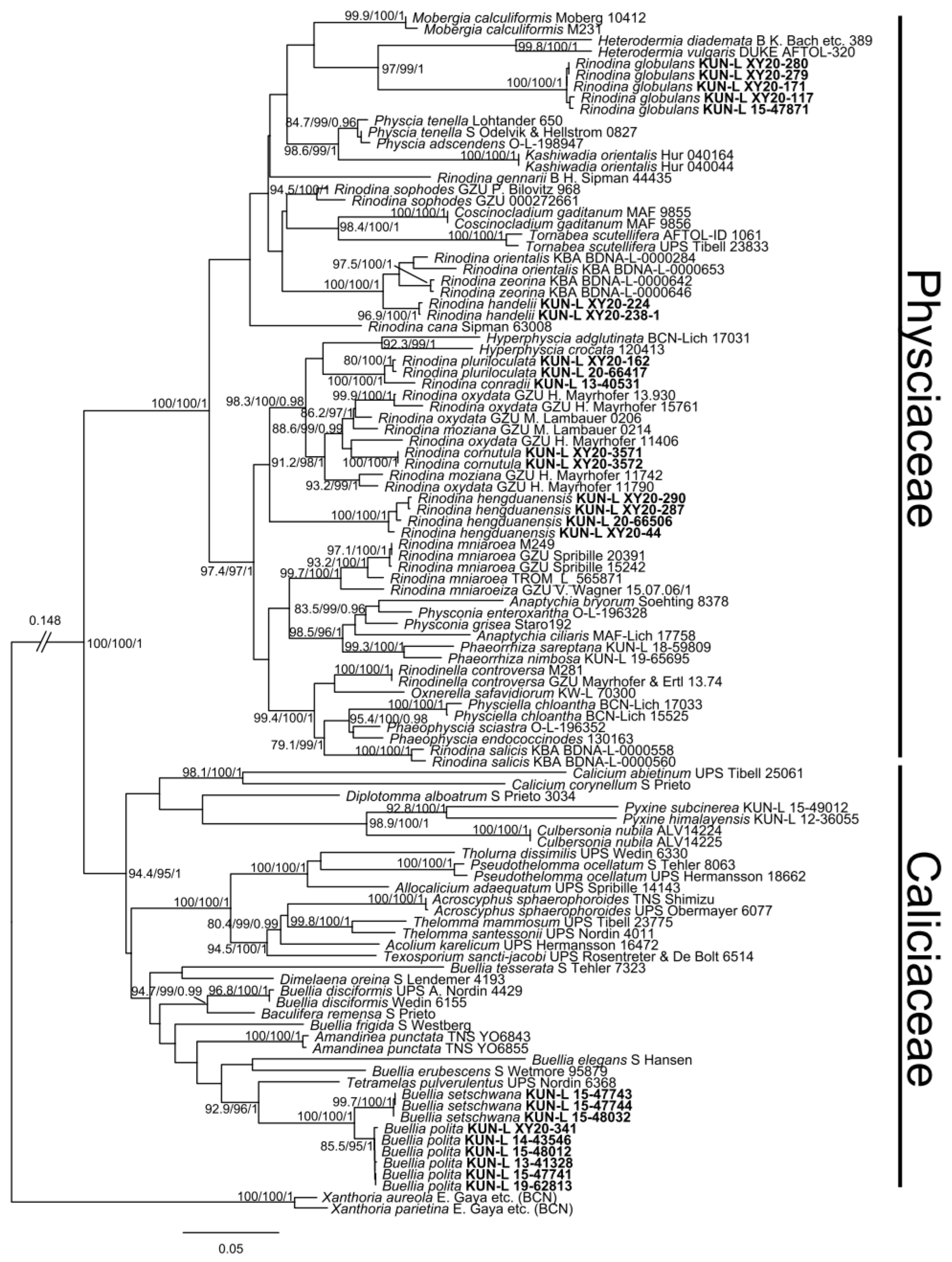
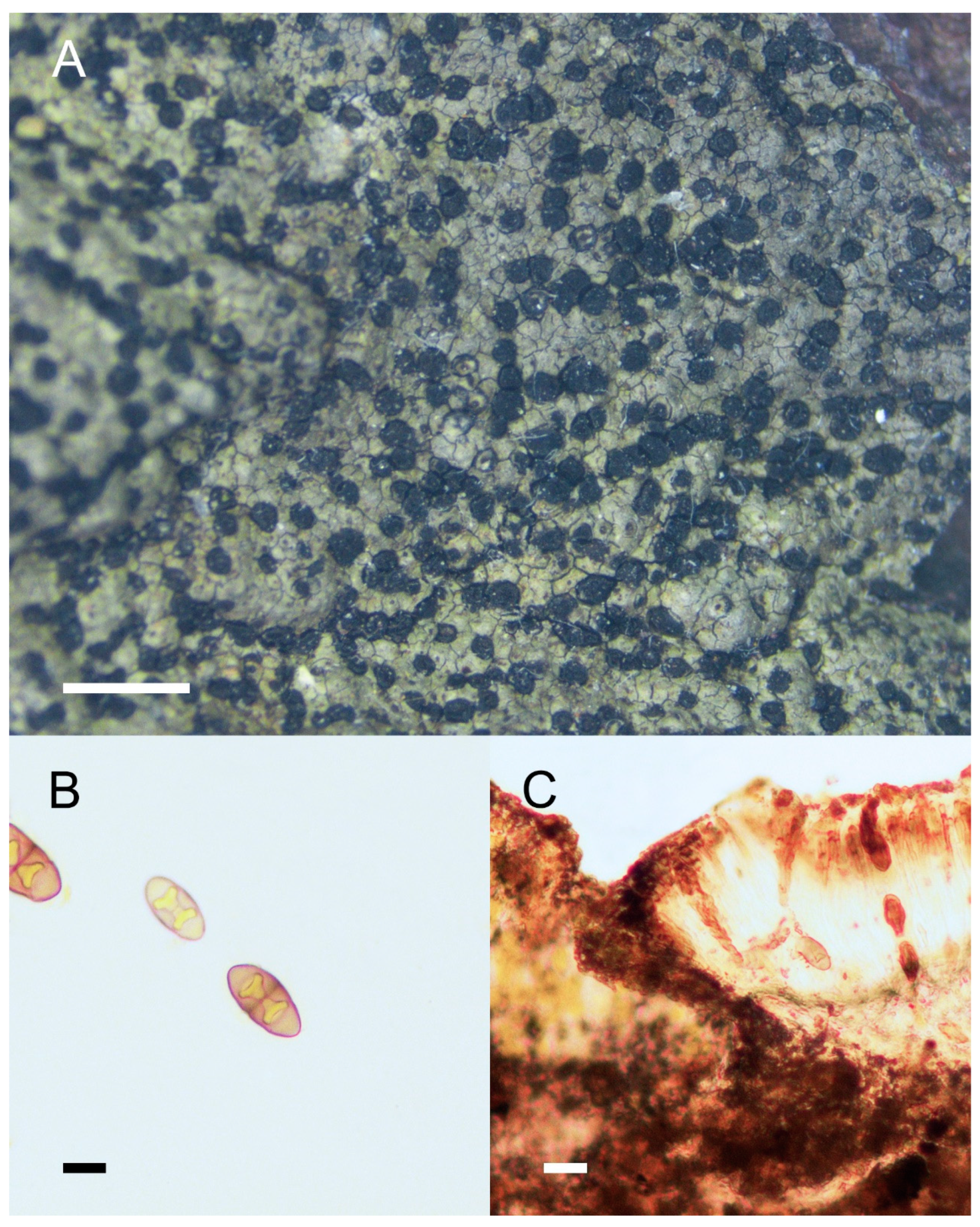
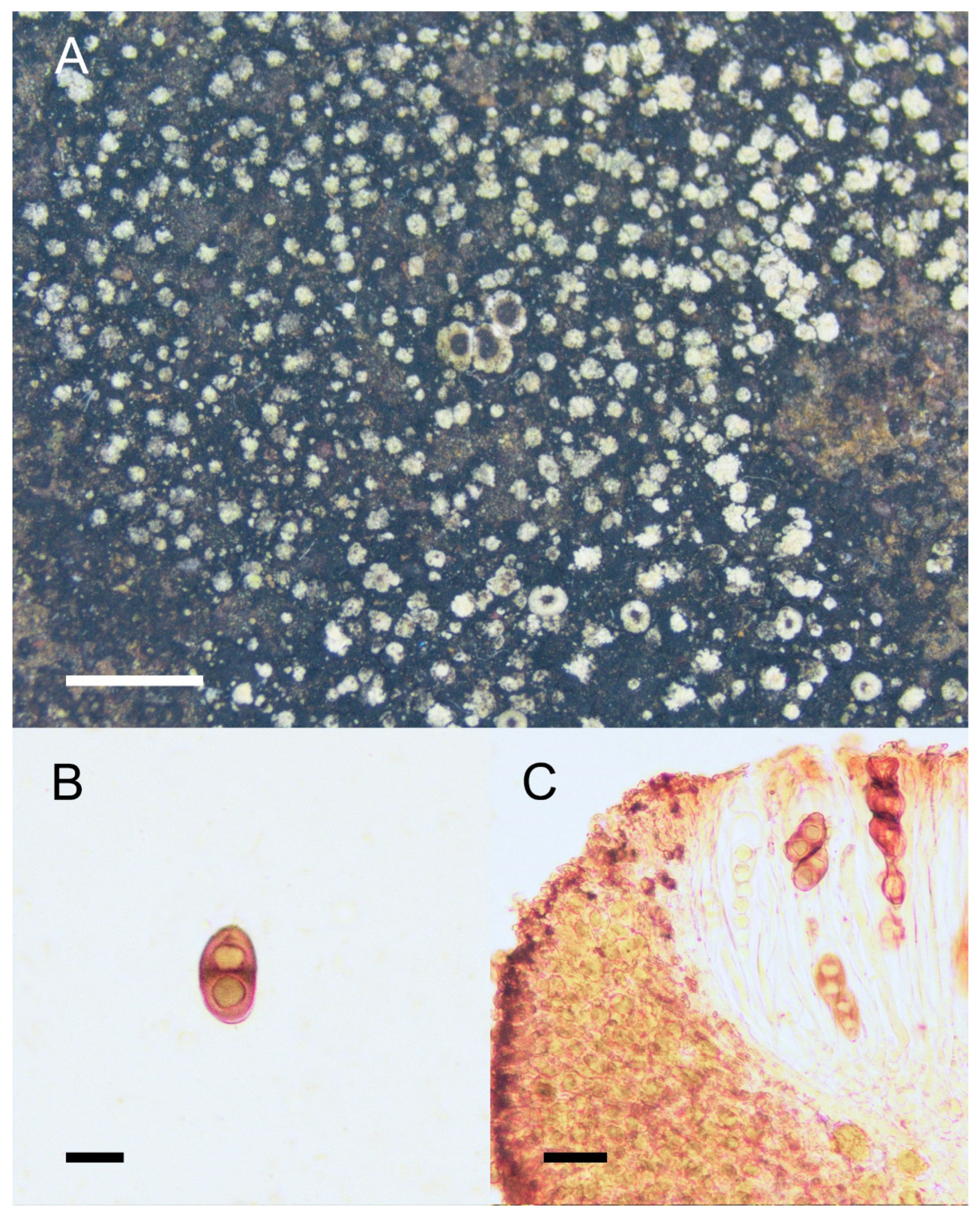
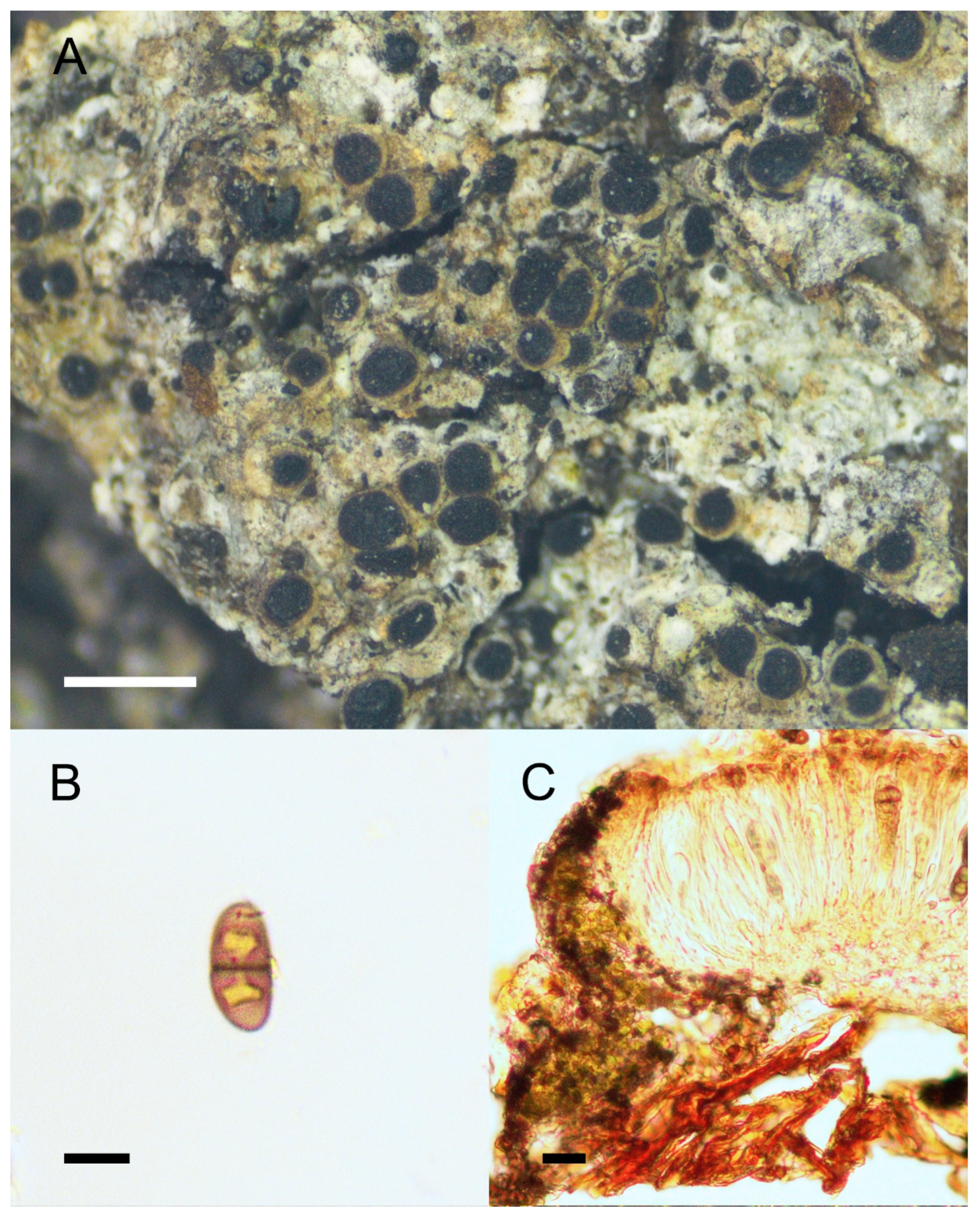
3. Results and Discussion
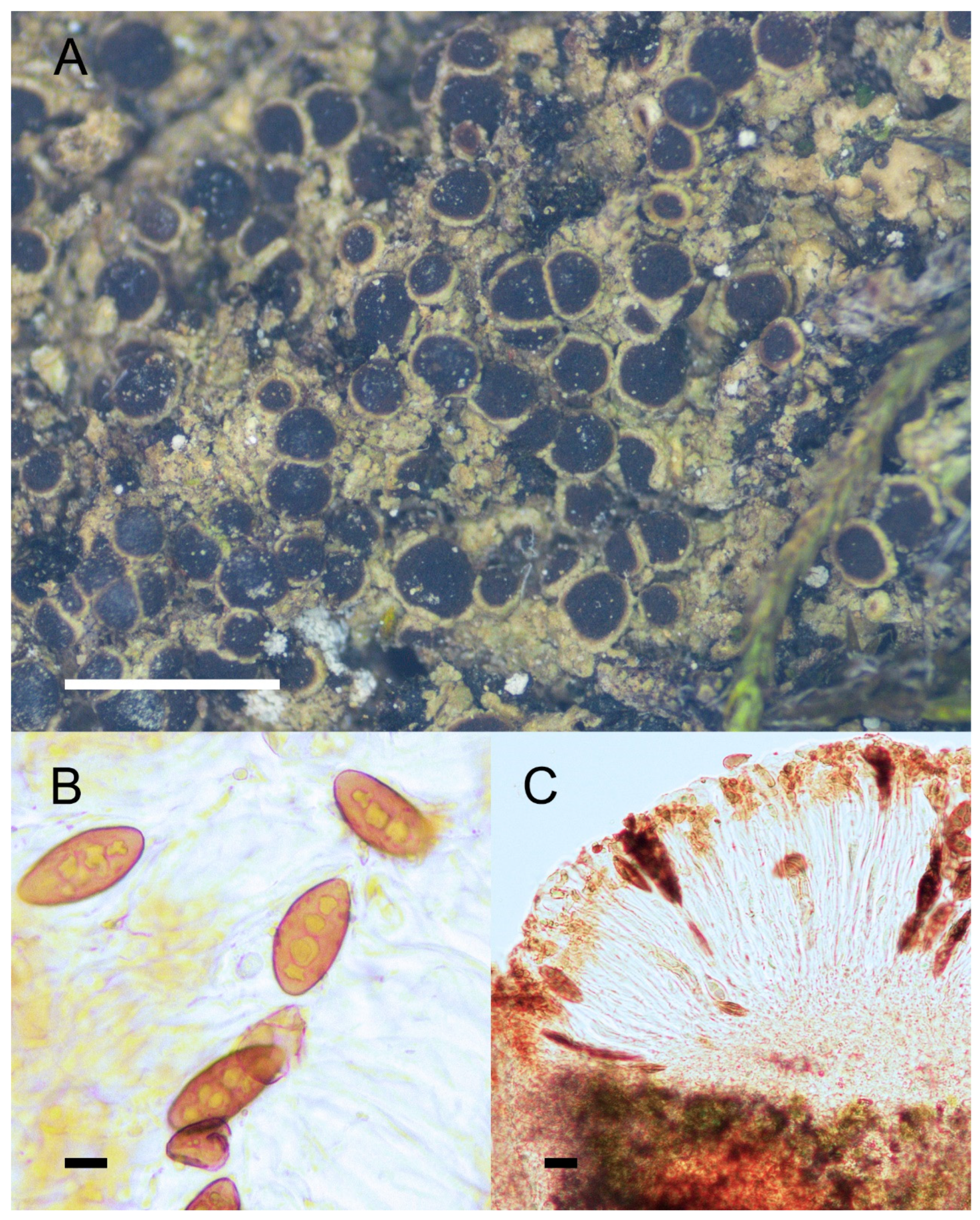
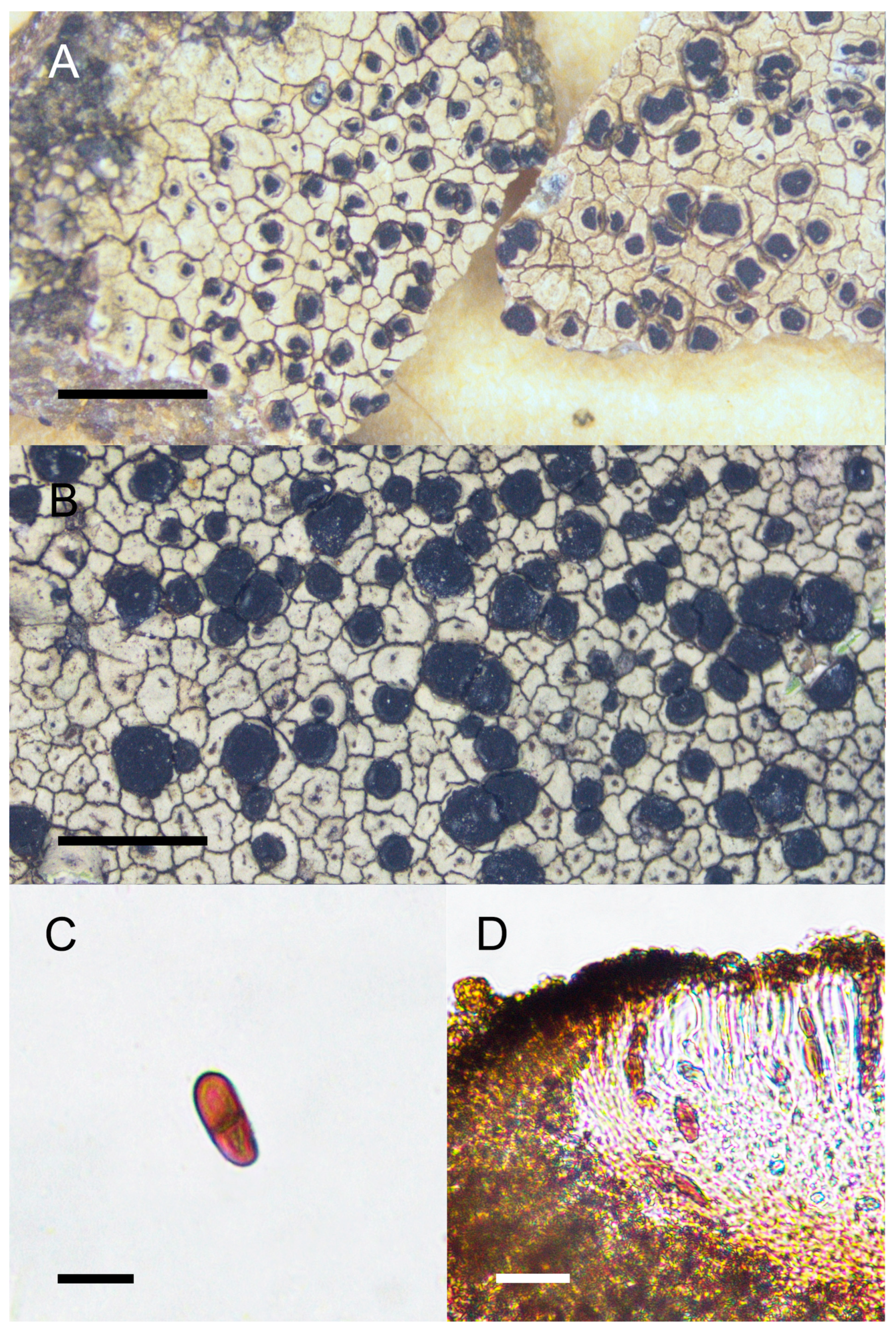
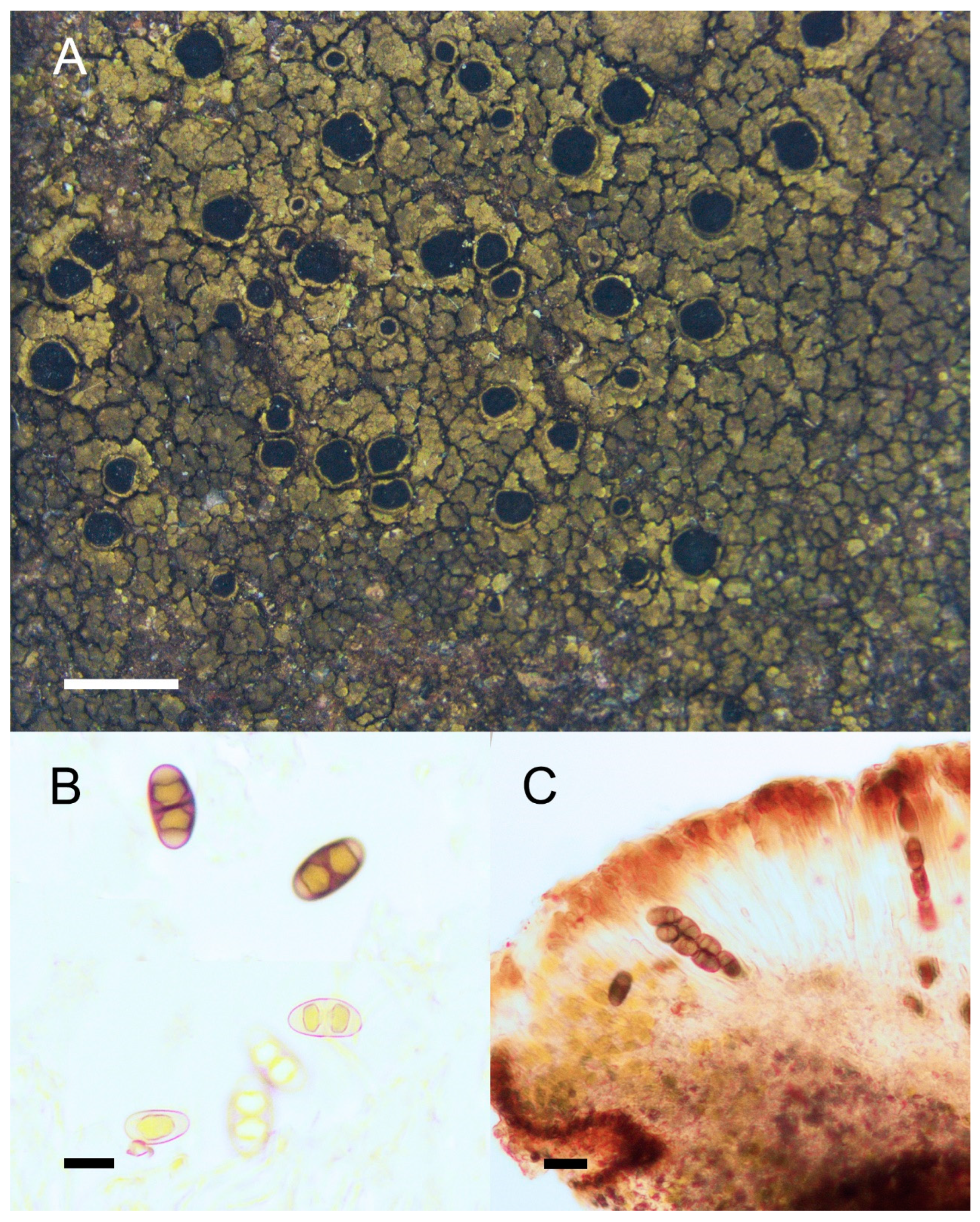
4. Taxonomy
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Giralt, M. The lichen genera Rinodina and Rinodinella (lichenized Ascomycetes, Physciaceae) in the Iberian Peninsula. Bibl. Lichenol. 2001, 79, 1–160. [Google Scholar]
- Mayrhofer, H.; Moberg, R. Rinodina. In Nordic Lichen Flora; Ahti, T., Jørgensen, P.M., Kristinsson, H., Eds.; Nordic Lichen Society: Uddevalla, Sweden, 2002; Volume 2, pp. 41–69. [Google Scholar]
- Giralt, M.; Matzer, M. The corticolous species of the genus Rinodina with biatorine or lecideine apothecia in southern Europe and Macaronesia. Lichenologist 1994, 26, 319–332. Available online: https://www.researchgate.net/publication/231922212 (accessed on 11 April 2023). [CrossRef]
- Giralt, M.; Mayrhofer, H.; Sheard, J.W. The corticolous and lignicolous sorediate, blastidiate and isidiate species of the genus Rinodina in southern Europe. Lichenologist 1995, 27, 3–24. Available online: https://www.researchgate.net/publication/231957857 (accessed on 11 April 2023). [CrossRef]
- Mayrhofer, H. Die saxicolen Arten der Flechtengattungen Rinodina und Rinodinella in der Alten Welt. J. Hattori Bot. Lab. 1984, 55, 327–493. [Google Scholar]
- Sheard, J.W. The Lichen Genus Rinodina (Ach.) Gray (Lecanoromycetidae, Physciaceae) in North America, North of Mexico; NRC Research Press: Ottawa, EO, Canada, 2010; pp. 1–246. [Google Scholar]
- Sheard, J.W. A synopsis and new key to the species of Rinodina (Ach.) Gray (Physciaceae, lichenized Ascomycetes) presently recognized in North America. Herzogia 2018, 31, 395–423. [Google Scholar]
- Sheard, J.W.; Ezhkin, A.K.; Galanina, I.A.; Himelbrant, D.; Kuznetsova, E.; Shimizu, A.; Stepanchikova, I.; Thor, G.; Tønsberg, T.; Yakovchenko, L.S.; et al. The lichen genus Rinodina (Physciaceae, Caliciales) in north-eastern Asia. Lichenologist 2017, 49, 617–672. Available online: https://www.researchgate.net/publication/321054036 (accessed on 12 April 2023). [CrossRef]
- Helms, G.; Friedl, T.; Rambold, G. Phylogenetic relationships of the Physciaceae inferred from rDNA sequence data and selected phenotypic characters. Mycologia 2003, 95, 1078–1099. Available online: https://www.researchgate.net/publication/49674865 (accessed on 13 April 2023). [CrossRef]
- Nadyeina, O.; Grube, M.; Mayrhofer, H. A contribution to the taxonomy of the genus Rinodina (Physciaceae, lichenized Ascomycotina) using combined ITS and mtSSU rDNA data. Lichenologist 2010, 42, 521–531. Available online: http://europepmc.org/article/PMC/3223597 (accessed on 13 April 2023). [CrossRef]
- Gaya, E.; Högnabba, F.; Holguin, A.; Molnar, K.; Fernández-Brime, S.; Stenroos, S.; Arup, U.; Søchting, U.; den Boom, P.V.; Lücking, R.; et al. Implementing a cumulative supermatrix approach for a comprehensive phylogenetic study of the Teloschistales (Pezizomycotina, Ascomycota). Mol. Phylogenet. Evol. 2012, 63, 374–387. [Google Scholar] [CrossRef]
- Resl, P.; Mayrhofer, H.; Clayden, S.R.; Spribille, T.; Thor, G.; Tønsberg, T.; Sheard, J.W. Morphological, chemical and species delimitation analyses provide new taxonomic insights into two groups of Rinodina. Lichenologist 2016, 48, 469–488. Available online: https://www.researchgate.net/publication/308793339 (accessed on 12 April 2023). [CrossRef]
- Wen, J. Evolution of eastern Asian and eastern North American disjunct distributions of flowering plants. Annu. Rev. Eco. Evol. Syst. 1999, 30, 421–455. [Google Scholar] [CrossRef]
- Xiang, Q.-Y.; Soltis, D.E.; Soltis, P.S. The eastern Asian and eastern and western North American disjunction: Congruent phylogenetic patterns in seven diverse genera. Mol. Phylogenet. Evol. 1998, 10, 178–190. [Google Scholar] [CrossRef]
- Qian, H. Floristic relationships between eastern Asia and North America: Test of Gray’s hypothesis. Am. Nat. 2002, 160, 317–332. Available online: https://www.researchgate.net/publication/23174933 (accessed on 12 April 2023). [CrossRef]
- Ding, W.; Ree, R.; Spicer, R.; Xing, Y. Ancient orogenic and monsoon-driven assembly of the world’s richest temperate alpine flora. Science 2020, 369, 578–581. [Google Scholar] [CrossRef] [PubMed]
- Jatta, A. Licheni Cinesi raccolti allo Shen-si negli anni 1894-1898 dal. rev. Padre Missionario, G. Giraldi. Nuovo G. Bot. Ital. 1902, 9, 477. Available online: https://bibdigital.rjb.csic.es/viewer/12810/?offset=#page=490&viewer=picture&o=bookmark&n=0&q= (accessed on 11 April 2023).
- Paulson, R. Lichens of Mount Everest. Bot. J. Linn. Soc. Lond. 1925, 63, 189–193. [Google Scholar]
- Zahlbruckner, A. Lichenes (Übersicht über sämtliche bisher aus China bekannten Flechten). In Symbolae Sinicae: Botanische Ergebnisse der Expedition der Akademie der Wissenschaften in Wien Nach Südwest-China, 1914–1918; Handel-Mazzetti, H.F., Brotherus, V.F., Eds.; Springer: Wien, Austria, 1930; Volume 3, pp. 226–234. Available online: https://www.biodiversitylibrary.org/item/15292#page/1/mode/1up (accessed on 12 April 2023).
- Magnusson, A.H. Lichens from Central Asia, I. In Reports Scientific Expedition North-West Provinces of China (the Sino-Swedish Expedition) 13, XI, Botany, 1; Hedin, S., Ed.; Aktiebolaget Thule: Stockholm, Sweden, 1940; pp. 145–156. [Google Scholar]
- Magnusson, A.H. Lichens from Central Asia II. In Reports Scientific Expedition North-West Provinces of China (the Sino-Swedish Expedition) 22, XI, Botany, 2; Hedin, S., Ed.; Aktiebolaget Thule: Stockholm, Sweden, 1944; pp. 55–57. [Google Scholar]
- Moreau, F.; Moreau, M.F. Lichens de Chine. Rev. Bryol. Et Lichenol. 1951, 20, 183–199. [Google Scholar]
- Wang Yang, J.-R.; Lai, M.-J. A checklist of the lichens of Taiwan. Taiwania 1973, 18, 83–104. [Google Scholar]
- Wu, J.-N.; Xiang, T.A. Primary study on lichens from Yuntai Mountain, Lianyungang, Jiangsu Province. J. Nanjing Norm. Univ.-Nat. Sci. 1981, 3, 1–11. (In Chinese) [Google Scholar]
- Xu, B.S. Cryptogamic Flora of the Yangtze Delta and Adjacent Regions; Shanghai Scientific and Technical Publishers: Shanghai, China, 1989; pp. 254–255. (In Chinese) [Google Scholar]
- Abbas, A.; Jiang, Y.C.; Wu, J.N. Lichens from the Tianshan Mountains, Xinjiang, China. J. Nanjing Norm. Univ.-Nat. Sci. 1993, 16, 74–82. (In Chinese) [Google Scholar]
- Abbas, A.; Wu, J.N. Lichens of Xinjiang; Sci-Tec & Hygiene Publishing House of Xinjiang: Urumqi, China, 1998; pp. 1–178. (In Chinese) [Google Scholar]
- Aptroot, A.; Seaward, M.R.D. Annotated checklist of Hongkong Lichens. Trop. Bryol. 1999, 17, 57–101. Available online: https://www.researchgate.net/publication/265989726 (accessed on 10 April 2023).
- Aptroot, A.; Sparrius, L.B. New microlichens from Taiwan. Fungal Divers. 2003, 14, 1–50. Available online: https://www.researchgate.net/publication/216829813 (accessed on 10 April 2023).
- Obermayer, W.; Blaha, J.; Mayrhofer, H. Buellia centralis and chemotypes of Dimelaena oreina in Tibet and other Central-Asian regions. Symb. Bot. Ups. 2004, 34, 327–342. Available online: https://www.researchgate.net/publication/242552311 (accessed on 12 April 2023).
- Aptroot, A.; Saipunkaew, W.; Sipman, H.J.M.; Sparrius, L.B.; Wolseley, P.A. New lichens from Thailand, mainly microlichens from Chiang Mai. Fungal Divers. 2007, 24, 75–134. Available online: https://www.researchgate.net/publication/228515656 (accessed on 10 April 2023).
- Li, Y.; Chen, C.; Zhao, Z. A primary study on lichens from Mount Yi. J. Fungal Res. 2008, 6, 70–73. (In Chinese) [Google Scholar]
- Liu, M.; Wei, J.-C. Lichen diversity in Shapotou region of Tengger Desert, China. Mycosystema 2013, 32, 42–50. (In Chinese) [Google Scholar]
- Joshi, S.; Jayalal, U.; Oh, S.-O.; Li, X.-R.; Jia, R.-L.; Hur, J.-S. New records of lichens from Shapotou area in Ningxia of Northwest China. Mycosystema 2014, 33, 167–173. Available online: https://www.researchgate.net/publication/263655180 (accessed on 11 April 2023).
- Kondratyuk, S.Y.; Lökös, L.; Halda, J.P.; Moniri, M.H.; Farkas, E.; Park, J.S.; Lee, B.G.; Oh, S.-O.; Hur, J.-S. New and noteworthy lichen-forming and lichenicolous fungi: 5. Acta Bot. Hung. 2016, 58, 319–396. [Google Scholar] [CrossRef]
- Ren, Q.; Zheng, X. Notes on the genus Rinodina in mainland China. J. Liaocheng Univ. (Nat. Sci.) 2020, 33, 85–97. (In Chinese) [Google Scholar]
- Tursun, A.; Shahidin, H.; Tumur, A. Taxonomic study on Rinodina in Xinjiang, China. Acta Bot. Boreali-Occident. Sin. 2020, 40, 1978–1988. (In Chinese) [Google Scholar]
- Wei, J.-C. The Enumeration of Lichenized Fungi in China; China Forestry Publishing House: Beijing, China, 2020; pp. 450–454. [Google Scholar]
- Culberson, C.F. Improved conditions and new data for identification of lichen products by standardized thin-layer chromatographic method. J. Chromatogr. A 1972, 72, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Orange, A.; James, P.W.; White, F.J. Microchemical Methods for the Identification of Lichens; British Lichen Society: London, UK, 2001; pp. 1–101. [Google Scholar]
- Zoller, S.; Scheidegger, C.; Sperisen, C. PCR primers for the amplification of mitochondrial small subunit ribosomal DNA of lichen-forming ascomycetes. Lichenologist 1999, 31, 511–516. Available online: https://www.researchgate.net/publication/229071472 (accessed on 12 April 2023). [CrossRef]
- Gardes, M.M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application for the identification of mycorrhizae and rust. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. Available online: https://www.researchgate.net/publication/223058289 (accessed on 12 April 2023).
- Hassouna, N.; Mithot, B.; Bachellerie, J.P. The complete nucleotide sequence of mouse 28S rRNA gene. Implications for the process of size increase of the large subunit rRNA in higher eukaryotes. Nucleic Acids Res. 1984, 12, 3563–3583. [Google Scholar] [CrossRef] [PubMed]
- Rehner, S.A.; Samuels, G.J. Taxonomy and phylogeny of Gliocladium analysed from nuclear large subunit ribosomal DNA sequences. Mycol. Res. 1994, 98, 625–634. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. Available online: http://europepmc.org/article/PMC/213247 (accessed on 12 April 2023). [CrossRef]
- Zhao, X.; Zhang, L.L.; Zhao, Z.T.; Wang, W.C.; Leavitt, S.D.; Lumbsch, H.T. Molecular phylogeny of the lichen genus Lecidella focusing on species from mainland China. PLoS ONE 2015, 10, e0139405. Available online: https://www.researchgate.net/publication/283181184 (accessed on 12 April 2023). [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3603318/pdf/mst010.pdf (accessed on 11 April 2023). [CrossRef]
- Chernomor, O.A.; von Haeseler, A.; Minh, B.Q. Terrace aware data structure for phylogenomic inference from supermatrices. Syst. Biol. 2016, 65, 997–1008. Available online: http://europepmc.org/backend/ptpmcrender.fcgi?accid=PMC5066062&blobtype=pdf (accessed on 10 April 2023). [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5453245/pdf/emss-72237.pdf (accessed on 11 April 2023). [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. Available online: http://europepmc.org/backend/ptpmcrender.fcgi?accid=PMC4271533&blobtype=pdf (accessed on 11 April 2023). [CrossRef] [PubMed]
- Trifinopoulos, J.; Nguyen, L.-T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. Available online: http://europepmc.org/article/PMC/4987875 (accessed on 12 April 2023).
- Zhou, X.; Shen, X.-X.; Hittinger, C.T.; Rokas, A. Evaluating fast maximum likelihood-based phylogenetic programs using empirical phylogenomic data sets. Mol. Biol. Evol. 2017, 35, 486–530. Available online: https://academic.oup.com/mbe/article/35/2/486/4644721 (accessed on 12 April 2023). [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3329765/pdf/sys029.pdf (accessed on 12 April 2023). [CrossRef] [PubMed]
- Rambaut, A. FigTree v. 1.4.2. Institute of Evolutionary Biology, University of Edinburgh. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 12 April 2023).
- Prieto, M.; Wedin, M. Phylogeny, taxonomy and diversification events in the Caliciaceae. Fungal Divers. 2017, 82, 221–238. Available online: https://link.springer.com/article/10.1007/s13225-016-0372-y (accessed on 16 April 2023). [CrossRef]
- Tibell, L. Tholurna dissimilis and generic delimitations in Caliciaceae inferred from nuclear ITS and LSU rDNA phylogenies (Lecanorales, lichenized ascomycetes). Mycol. Res. 2003, 107, 1403–1418. [Google Scholar] [CrossRef]
- Ohmura, Y.; Sugimoto, M.; Yakovchenko, L.; Davydov, E.A. Additional species and ITS rDNA data for the lichen mycota of the Imperial Palace grounds, Tokyo, Japan. Bull. Natl. Mus. Nat. Sci. 2022, 48, 1–16. Available online: https://www.researchgate.net/publication/358952425 (accessed on 16 April 2023).
- Lohtander, K.; Ahti, T.; Stenroos, S.; Urbanavichus, G. Is Anaptychia monophyletic? A phylogenetic study based on nuclear and mitochondrial genes. Ann. Bot. Fenn. 2008, 45, 55–60. Available online: https://www.researchgate.net/publication/232273420 (accessed on 16 April 2023). [CrossRef]
- Molina, M.C.; Divakar, P.K.; González, N. Success in the isolation and axenic culture of Anaptychia ciliaris (Physciaceae, Lecanoromycetes) mycobiont. Mycoscience 2015, 56, 351–358. Available online: https://www.researchgate.net/publication/272891236 (accessed on 16 April 2023). [CrossRef]
- Wedin, M.; Baloch, E.; Grube, M. Parsimony analyses of mtSSU and nITS rDNA sequences reveal the natural relationships of the lichen families Physciaceae and Caliciaceae. Taxon 2002, 51, 655–660. Available online: https://nhm2.uio.no/botanisk/lav/RLL/PDF20/R24173.pdf (accessed on 16 April 2023). [CrossRef]
- Crespo, A.; Blanco, O.; Llimona, X.; Ferencová, Z.L.; Hawksworth, D.L. Coscinocladium, an overlooked endemic and monotypic Mediterranean lichen genus of Physciaceae, reinstated by molecular phylogenetic analysis. Taxon 2004, 53, 405–414. [Google Scholar] [CrossRef]
- Aptroot, A.; Maphangwa, K.; Zedda, L.; Tekere, M.; Alvarado, P.; Sipman, H. The phylogenetic position of Culbersonia is in the Caliciaceae (lichenized ascomycetes). Lichenologist 2019, 51, 187–191. [Google Scholar] [CrossRef]
- Schmull, M.; Miadlikowska, J.; Pelzer, M.; Stocker-Worgotter, E.; Hofstetter, V.; Fraker, E.; Hodkinson, B.P.; Reeb, V.; Kukwa, M.; Lumbsch, H.T.; et al. Phylogenetic affiliations of members of the heterogeneous lichen-forming fungi of the genus Lecidea sensu zahlbruckner (lecanoromycetes, ascomycota). Mycologia 2011, 103, 983–1003. Available online: https://www.researchgate.net/publication/51192614 (accessed on 16 April 2023). [CrossRef] [PubMed]
- Miadlikowska, J.; Kauff, F.; Hofstetter, V.; Fraker, E.; Grube, M.; Hafellner, J.; Reeb, V.; Hodkinson, B.P.; Kukwa, M.; Lücking, R.; et al. New insights into classification and evolution of the Lecanoromycetes (Pezizomycotina, Ascomycota) from phylogenetic analyses of three ribosomal RNA- and two protein-coding genes. Mycologia 2006, 98, 1088–1103. Available online: https://www.researchgate.net/publication/6344409 (accessed on 16 April 2023). [CrossRef] [PubMed]
- Grube, M.; Arup, U. Molecular and morphological evolution in the Physciaceae (Lecanorales, lichenized Ascomycotina), with special emphasis on the genus Rinodina. Lichenologist 2001, 33, 63–72. [Google Scholar] [CrossRef]
- Liu, D.; Hur, J.-S. Revision of the lichen genus Phaeophyscia and allied atranorin absent taxa (Physciaceae) in South Korea. Microorganisms 2019, 7, 242. Available online: https://www.mdpi.com/2076-2607/7/8/242 (accessed on 16 April 2023). [CrossRef]
- Kondratyuk, S.Y.; Lőkös, L.; Kim, J.A.; Jeong, M.-H.; Kondratiuk, A.; Oh, S.-O.; Hur, J.-S. Kashiwadia gen. nov. (Physciaceae, lichen-forming Ascomycota), proved by phylogenetic analysis of the Eastern Asian Physciaceae. Acta Bot. Hung. 2014, 56, 369–378. Available online: https://www.researchgate.net/publication/277924093 (accessed on 16 April 2023). [CrossRef]
- Lohtander, K.; Källersjö, M.; Moberg, R.; Tehler, A. The family Physciaceae in Fennoscandia: Phylogeny inferred from ITS sequences. Mycologia 2000, 92, 728–735. Available online: https://www.researchgate.net/publication/232273357 (accessed on 16 April 2023). [CrossRef]
- Kondratyuk, S.Y.; Lőkös, L.; Kim, J.A.; Jeong, M.-H.; Zarei-Darki, B.; Hur, J.-S. Oxnerella safavidiorum gen. et spec. nov. (Lecanoromycetidae, Ascomycota) from Iran (Asia) proved by phylogenetic analysis. Acta Bot. Hung. 2014, 56, 379–398. Available online: https://go.gale.com/ps/i.do?p=AONE&u=googlescholar&id=GALE|A458953398&v=2.1&it=r&asid=e883ca1e (accessed on 16 April 2023). [CrossRef]
- Marthinsen, G.; Rui, S.; Timdal, E. OLICH: A reference library of DNA barcodes for Nordic lichens. Biodivers. Data J. 2019, 7, e36252. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6711938/ (accessed on 16 April 2023). [CrossRef]
- Zhong, Q.; Zhang, Y.; Wang, X.; Timdal, E.; Gong, H.; Wang, Z.; Wang, L. Phaeorrhiza (Physciaceae), a new lichen genus record to China. Phytotaxa 2021, 510, 228–238. [Google Scholar] [CrossRef]
- Yang, M.-X.; Wang, X.-Y.; Liu, D.; Zhang, Y.-Y.; Li, L.-J.; Yin, A.-C.; Scheidegger, C.; Wang, L.-S. New species and records of Pyxine (Caliciaceae) in China. MycoKeys 2019, 45, 93–109. Available online: https://mycokeys.pensoft.net/article/29374/ (accessed on 16 April 2023). [CrossRef] [PubMed]
- Sipman, H.; Aptroot, A. Ikaeria serusiauxii, a new Caloplaca-like lichen from Macaronesia and mainland Portugal, with a lichen checklist for Porto Santo. Plant Fungal Syst. 2020, 65, 120–130. Available online: https://www.semanticscholar.org/paper/02f5c28f8a50f47d97994584d5283511e0c4c6d8 (accessed on 16 April 2023). [CrossRef]
- Kaschik, M. Taxonomic Studies on Saxicolous Species of the Genus Rinodina (Lichenized Ascomycetes, Physciaceae) in the Southern Hemisphere with Emphasis in Australia and New Zealand; Bibliotheca Lichenologica: Stuttgart, Germany, 2006; pp. 1–162. [Google Scholar]
- Lee, B.G.; Hur, J.-S. Two new Rinodina lichens from South Korea, with an updated key to the species of Rinodina in the far eastern Asia. Mycokeys 2022, 87, 159–182. Available online: https://europepmc.org/article/PMC/PMC8891229 (accessed on 16 April 2023). [CrossRef] [PubMed]
- Bhattacharya, D.; Friedl, T.; Helms, G. Vertical evolution and intragenic spread of lichen-fungal group I introns. J. Mol. Evol. 2002, 55, 74–84. [Google Scholar] [CrossRef]
- Abbas, A.; Wu, J.N. The lichens from Tielimaiti Pass Kuqa County, Xinjiang, China. Arid Zone Res. 1994, 11, 19–23. (In Chinese) [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Locality | Voucher Specimens | GenBank Numbers | References | ||
|---|---|---|---|---|---|---|
| nrITS | mtSSU | nrLSU | ||||
| Acolium karelicum | unknown | UPS Hermansson 16472 | KX512897 | na | KX512879 | [56] |
| Acroscyphus sphaerophoroides 1 | unknown | UPS Obermayer 6077 | KX512898 | KX512984 | na | [56] |
| A. sphaerophoroides 2 | unknown | TNS Shimizu | AY450562 | na | na | [57] |
| Allocalicium adaequatum | UPS Spribille 14143 | KX512906 | KX512986 | KX512859 | [56] | |
| Amandinea punctata 1 | Japan, Tokyo | TNS YO6843 | LC669587 | na | na | [58] |
| A. punctata 2 | Japan, Tokyo | TNS YO6855 | LC669588 | na | na | [58] |
| Anaptychia bryorum | Bhutan, Thimphu | Soehting 8378 | EF582777 | EF582825 | na | [59] |
| A. ciliaris | Spain | MAF-Lich 17758 | KC559095 | na | na | [60] |
| Baculifera remensa | S Prieto | na | KX512962 | KX512881 | [56] | |
| Buellia disciformis 1 | Jämtland, Klaxåsen | Wedin 6155 | AY143392 | AY143401 | na | [61] |
| B. disciformis 2 | Sweden | UPS A. Nordin 4429 | AF540498 | na | na | [9] |
| B. elegans | unknown | S Hansen | KX512901 | KX512993 | na | [56] |
| B. erubescens | unknown | S Wetmore 95879 | KX512902 | KX512969 | KX512874 | [56] |
| B. frigida | unknown | S Westberg | KX512903 | KX512992 | KX512852 | [56] |
| B. polita 1 | China, Sichuan | KUN-L XY20-341 | OP526814 | OP526788 | OP526772 | |
| B. polita 2 | China, Yunnan | KUN-L 15-47741 | OP526792 | OP526774 | OP526763 | |
| B. polita 3 | China, Sichuan | KUN-L 19-62813 | OP526800 | OP526780 | OP526768 | |
| B. polita 4 | China | KUN-L 14-43546 | MN615679 | na | na | |
| B. polita 5 | China | KUN-L 15-48012 | MN615678 | na | na | |
| B. polita 6 | China | KUN-L 13-41328 | MN615677 | na | na | |
| B. setschwana 1 | China, Yunnan | KUN-L 15-48032 | OP526796 | na | na | |
| B. setschwana 2 | China, Yunnan | KUN-L 15-47744 | OP526794 | na | na | |
| B. setschwana 3 | China, Yunnan | KUN-L 15-47743 | OP526793 | OP526775 | OP526764 | |
| B. tesserata | unknown | S Tehler 7323 | KX512904 | na | KX512885 | [56] |
| Calicium abietinum | unknown | UPS Tibell 25061 | KX512905 | KX512971 | KX512872 | [56] |
| C. corynellum | unknown | S Prieto | KX512908 | KX512985 | KX512855 | [56] |
| Coscinocladium gaditanum 1 | Spain | MAF 9855 | AY449720 | AY464073 | na | [62] |
| C. gaditanum 2 | Spain | MAF 9856 | AY449721 | AY464074 | na | [62] |
| Culbersonia nubila 1 | South Africa | ALV14224 | MH121317 | na | MH121319 | [63] |
| C. nubila 2 | South Africa | ALV14225 | MH121318 | na | MH121320 | [63] |
| Dimelaena oreina | unknown | S Lendemer 4193 | KX512922 | KX512976 | KX512867 | [56] |
| Diplotomma alboatrum | unknown | S Prieto 3034 | KX512924 | KX512966 | KX512877 | [56] |
| Heterodermia diademata | Bolivia, Depto. Beni | B K. Bach, M. Kesseler and Portugal 389 | AF540518 | na | na | [9] |
| H. vulgaris | unknown | DUKE AFTOL-320 | HQ650704 | na | DQ883798 | [64,65] |
| Hyperphyscia adglutinata | unknown | BCN-Lich 17031 | AF250795 | GU247189 | na | [66] |
| H. crocata | South Korea | 120413 | MN150490 | na | na | [67] |
| Kashiwadia orientalis 1 | Eastern Asia | Hur 040044 | na | KM397366 | na | [68] |
| K. orientalis 2 | Eastern Asia | Hur 040164 | na | KM397365 | na | [68] |
| Mobergia calculiformis 1 | Mexico | Moberg 10412 | AF224359 | na | na | [69] |
| M. calculiformis 2 | unknown | M231 | AF250796 | na | na | [66] |
| Oxnerella safavidiorum | Iran | KW-L 70300 | KM410153 | KM410156 | na | [70] |
| Phaeophyscia endococcinodes | South Korea | 130163 | MN150503 | na | na | [67] |
| P. sciastra | Norway, Sor-Trondelag | O-L-196352 | MK812372 | na | na | [71] |
| Phaeorrhiza nimbosa | China, Xizang | KUN-L 19-65695 | MW133637 | MW133652 | MW133660 | [72] |
| P. sareptana | China, Gansu | KUN-L 18-59809 | MW133625 | MW133640 | MW133654 | [72] |
| Physcia adscendens | Norway, Vestfold | O-L-198947 | MK812201 | na | na | [71] |
| P. tenella 1 | Gästrikland | S Odelvik and Hellstrom 0827 | KX512932 | KX512974 | KX512869 | [56] |
| P. tenella 2 | Finland | Lohtander 650 | AF224425 | EF582800 | na | [59] |
| Physciella chloantha 1 | Iberian Peninsula | BCN-Lich 15525 | GU247166 | GU247200 | na | |
| P. chloantha 2 | Iberian Peninsula | BCN-Lich 17033 | GU247164 | GU247198 | na | |
| Physconia enteroxantha | Norway, Akershus | O-L-196328 | MK812142 | na | na | [71] |
| P. grisea | Morocco, Imli | Staro192 | LS483208 | na | na | [71] |
| Pseudothelomma ocellatum 1 | unknown | S Tehler 8063 | KX512934 | KX512957 | KX512862 | [56] |
| P. ocellatum 2 | unknown | UPS Hermansson 18662 | KX512935 | KX512952 | KX512891 | [56] |
| Pyxine himalayensis | China, Yunnan | KUN-L 12-36055 | KY611881 | KY751388 | na | [73] |
| P. subcinerea | China, Taiwan | KUN-L 15-49012 | KY611867 | KY751374 | na | [73] |
| Rinodina cana | unknown | Sipman 63008 | MN587029 | na | na | [74] |
| R. conradii | China, Yunnan | KUN-L 13-40531 | OP526791 | OP526773 | na | |
| R. cornutula 1 | China, Yunnan | KUN-L XY20-3571 | OP526815 | OP526789 | na | |
| R. cornutula 2 | China, Yunnan | KUN-L XY20-3572 | OP526816 | OP526790 | na | |
| R. gennarii | Netherlands, Utrecht | B H. Sipman 44435 | AJ544187 | na | na | [9] |
| R. globulans 1 | China, Yunnan | KUN-L 15-47871 | OP526795 | OP526776 | na | |
| R. globulans 2 | China, Yunnan | KUN-L XY20-171 | OP526807 | OP526785 | OP526770 | |
| R. globulans 3 | China, Yunnan | KUN-L XY20-117 | OP526805 | na | na | |
| R. globulans 4 | China, Sichuan | KUN-L XY20-279 | OP526810 | na | na | |
| R. globulans 5 | China, Sichuan | KUN-L XY20-280 | OP526811 | na | na | |
| R. handelii 1 | China, Yunnan | KUN-L XY20-224 | OP526808 | na | na | |
| R. handelii 2 | China, Yunnan | KUN-L XY20-238-1 | OP526809 | na | na | |
| R. mniaroea 1 | USA, Idaho | GZU Spribille 15242 | KX015687 | KX015706 | na | [12] |
| R. mniaroea 2 | USA, Montana | GZU Spribille 20391 | KX015692 | KX015711 | na | [12] |
| R. mniaroea 3 | Norway, Svalbard | TROM_L_565871 | MK812098 | na | na | [71] |
| R. mniaroea 4 | unknown | M249 | AF250811 | na | na | [66] |
| R. mniaroeiza | Canada | GZU V. Wagner 15.07.06/1 | KX015691 | KX015710 | na | [12] |
| R. moziana 1 | Australia, Queensland | GZU H. Mayrhofer 11742 | DQ849306 | na | na | [75] |
| R. moziana 2 | New Zealand, Nelson | GZU M. Lambauer 0214 | DQ849305 | na | na | [75] |
| R. oxydata 1 | Australia, Queensland | GZU H. Mayrhofer 11790 | DQ849311 | na | na | [75] |
| R. oxydata 2 | Australia, Queensland | GZU H. Mayrhofer 11406 | DQ849313 | na | na | [75] |
| R. oxydata 3 | New Zealand, Nelson | GZU M. Lambauer 0206 | DQ849310 | na | na | [75] |
| R. oxydata 4 | Austria, Steiermark | GZU H. Mayrhofer 13.930 | AF540548 | na | na | [9] |
| R. oxydata 5 | Austria, Styria | GZU H. Mayrhofer 15761 | DQ849312 | na | na | [75] |
| R. pluriloculata 1 | China, Yunnan | KUN-L XY20-162 | OP526806 | OP526784 | na | |
| R. pluriloculata 2 | China, Yunnan | KUN-L 20-66417 | OP526802 | OP526782 | na | |
| R. sophodes 1 | Austria, Styria | GZU 000272661 | GU553304 | GU553321 | na | [10] |
| R. sophodes 2 | Austria, Steiermark | GZU P. Bilovitz 968 | AF540550 | na | na | [9] |
| R. orientalis 1 | South Korea | KBA BDNA-L-0000284 | MW832807 | na | na | [76] |
| R. orientalis 2 | South Korea | KBA BDNA-L-0000653 | MW832808 | na | na | [76] |
| R. salicis 1 | South Korea | KBA BDNA-L-0000558 | MW832810 | na | na | [76] |
| R. salicis 2 | South Korea | KBA BDNA-L-0000560 | MW832811 | na | na | [76] |
| R. hengduanensis 1 | China, Sichuan | KUN-L 20-66506 | OP526803 | OP526783 | na | |
| R. hengduanensis 2 | China, Yunnan | KUN-L XY20-44 | OP526804 | na | na | |
| R. hengduanensis 3 | China, Sichuan | KUN-L XY20-290 | OP526813 | OP526787 | na | |
| R. hengduanensis 4 | China, Sichuan | KUN-L XY20-287 | OP526812 | OP526786 | OP526771 | |
| R. zeorina 1 | South Korea | KBA BDNA-L-0000642 | MW832812 | na | na | [76] |
| R. zeorina 2 | South Korea | KBA BDNA-L-0000646 | MW832813 | na | na | [76] |
| Rinodinella controversa 1 | unknown | M281 | AF250814 | na | na | [66] |
| R. controversa 2 | Greece, Kreta | GZU Mayrhofer and Ertl 13.74 | AJ421423 | na | na | [77] |
| Tetramelas pulverulentus | unknown | UPS Nordin 6368 | KX512940 | KX512983 | KX512860 | [56] |
| Texosporium sancti-jacobi | unknown | UPS Rosentreter and De Bolt 6514 | KX512941 | KX512981 | KX512863 | [56] |
| Thelomma mammosum | unknown | UPS Tibell 23775 | KX512942 | KX512954 | KX512888 | [56] |
| T. santessonii | unknown | UPS Nordin 4011 | KX512944 | KX512951 | KX512889 | [56] |
| Tholurna dissimilis | unknown | UPS Wedin 6330 | AY143397 | AY143407 | KX512893 | [56] |
| Tornabea scutellifera 1 | Spain | AFTOL-ID 1061 | JQ301698 | na | DQ973037 | [11,65] |
| T. scutellifera 2 | unknown | UPS Tibell 23833 | KX512946 | KX512970 | KX512873 | [56] |
| Xanthoria aureola | Sweden, Bohuslän | E. Gaya etc. (BCN) | JQ301690 | JQ301526 | JQ301585 | [12] |
| Xanthoria parietina | Sweden, Bohuslän | E. Gaya etc. (BCN) | JQ301691 | JQ301530 | JQ301589 | [12] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, Q.; Ai, M.; Worthy, F.R.; Yin, A.; Jiang, Y.; Wang, L.; Wang, X. Rediscovery of Five Rinodina Species Originally Described from Southwest China and One New Species. Diversity 2023, 15, 705. https://doi.org/10.3390/d15060705
Zhong Q, Ai M, Worthy FR, Yin A, Jiang Y, Wang L, Wang X. Rediscovery of Five Rinodina Species Originally Described from Southwest China and One New Species. Diversity. 2023; 15(6):705. https://doi.org/10.3390/d15060705
Chicago/Turabian StyleZhong, Qiuyi, Min Ai, Fiona Ruth Worthy, Ancheng Yin, Yi Jiang, Lisong Wang, and Xinyu Wang. 2023. "Rediscovery of Five Rinodina Species Originally Described from Southwest China and One New Species" Diversity 15, no. 6: 705. https://doi.org/10.3390/d15060705
APA StyleZhong, Q., Ai, M., Worthy, F. R., Yin, A., Jiang, Y., Wang, L., & Wang, X. (2023). Rediscovery of Five Rinodina Species Originally Described from Southwest China and One New Species. Diversity, 15(6), 705. https://doi.org/10.3390/d15060705