



New Material of Karakoromys (Ctenodactylidae, Rodentia) from Late Eocene-Early Oligocene of Ulantatal (Nei Mongol): Taxonomy, Diversity, and Response to Climatic Change


Abstract
:1. Introduction
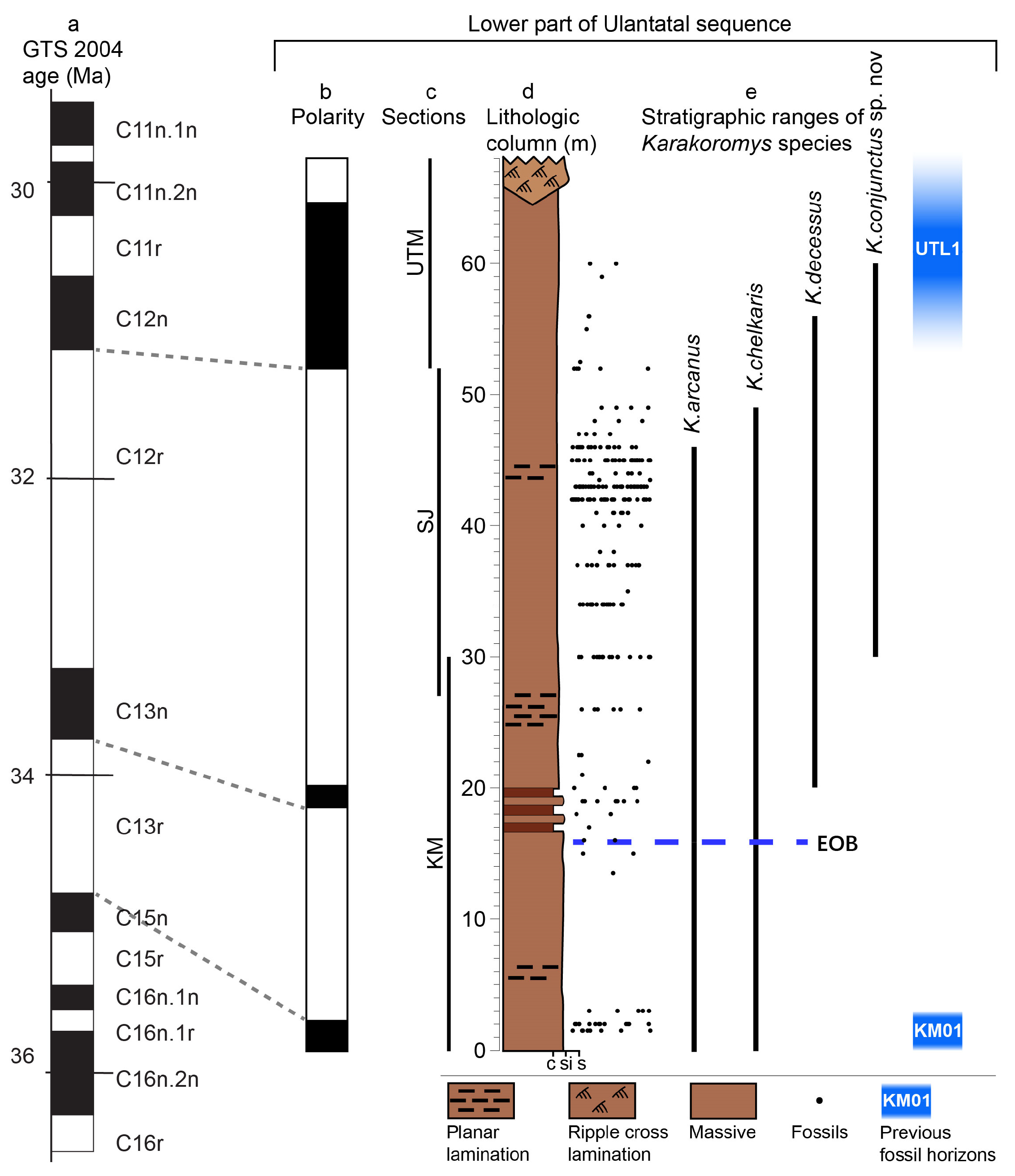
2. Geological Setting
3. Materials and Methods
3.1. Materials
3.2. Determining the Age of Fossils
3.3. Systematic Paleontological Study
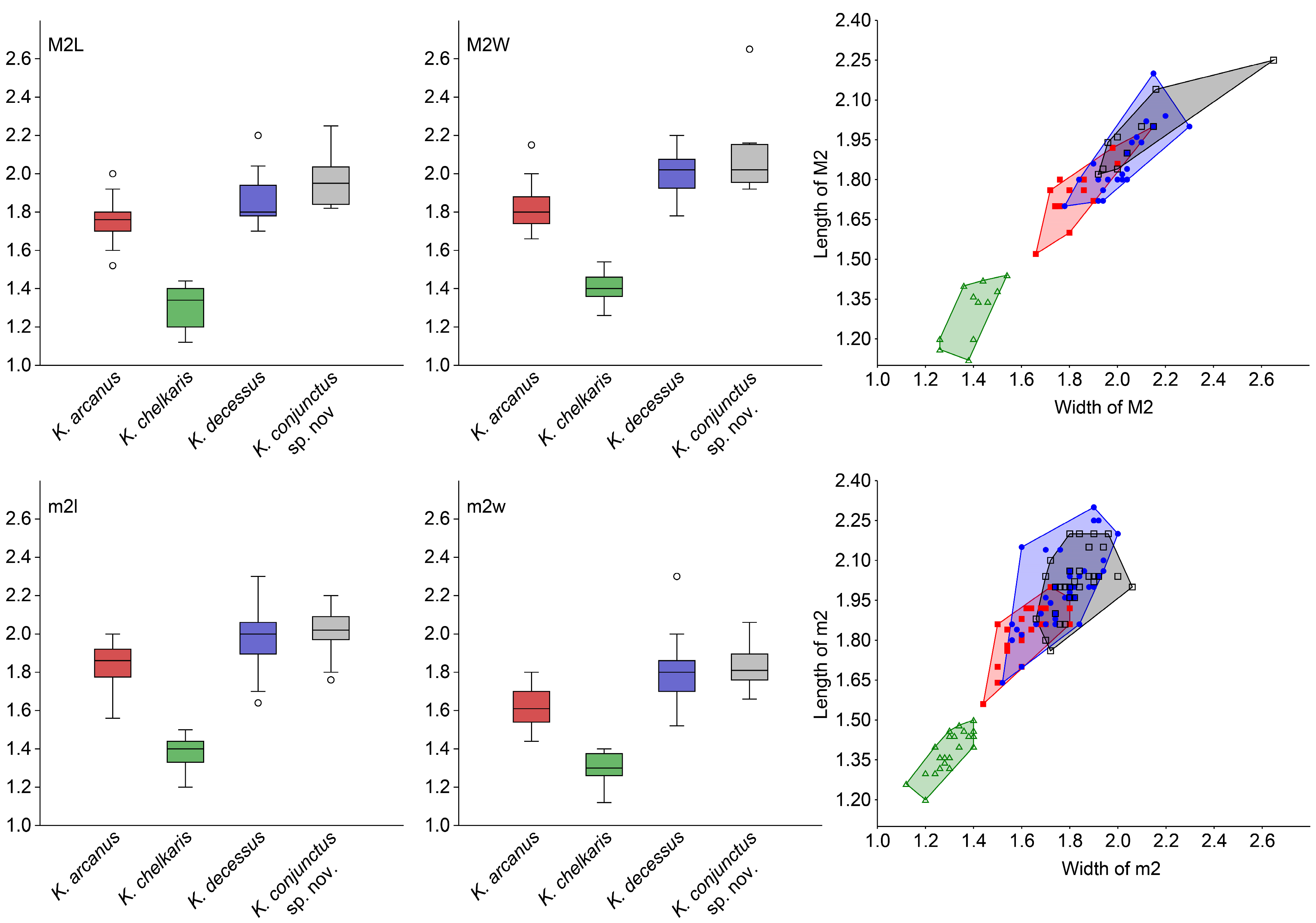
3.4. Statistics
3.5. Diversity Analysis
3.6. Institutional Abbreviations
4. Systematic Paleontology
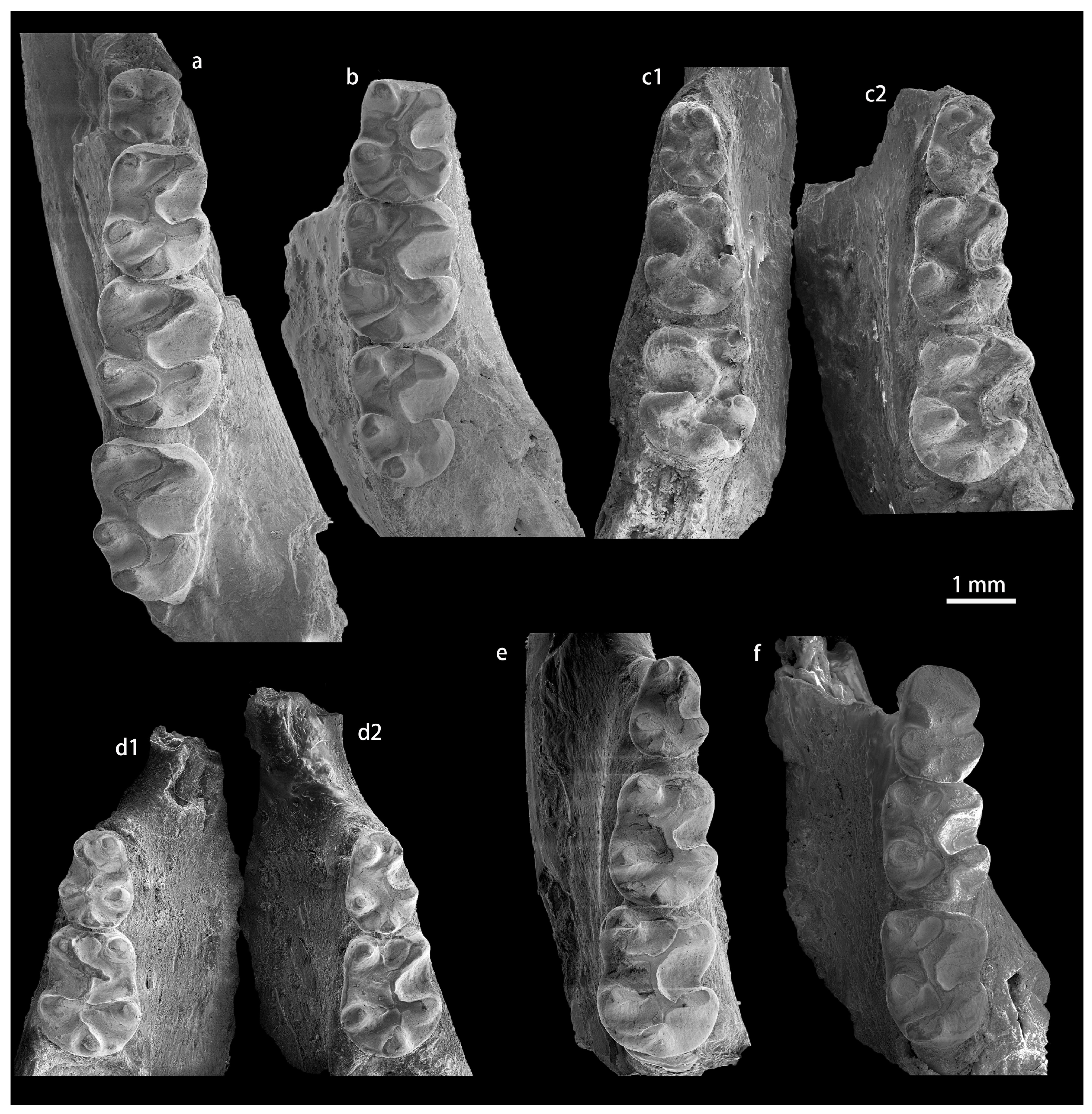
4.1. Karakoromys decessus Matthew et Granger, 1923 [18]
(Figure A1, Figure A2, Figure A3 and Figure A4, Table S1)
4.2. Karakoromys arcanus (Shevyreva, 1971) [53]
(Figure A5 and Figure A6, Table S1)
4.3. Karakoromys chelkaris (Shevyreva, 1971) [53]
(Figure A7 and Figure A8, Table S1)
4.4. Karakoromys conjunctus sp. nov.
(Figure A9 and Figure A10, Table S1)
5. Discussion
5.1. Stratigraphic Distribution and Biochronological Significance of Karakoromys

5.2. Karakoromys Diversity in Response to Climatic Change
5.2.1. The Diversity of Karakoromys and Related Taxa

5.2.2. The Possible Relationship between Karakoromys Diversity and Climatic Change
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A










References
- Zachos, J.; Pagani, M.; Sloan, L.; Thomas, E.; Billups, K. Trends, rhythms, and aberrations in global climate 65 Ma to present. Science 2001, 292, 686–693. [Google Scholar] [CrossRef]
- Liu, Z.; Pagani, M.; Zinniker, D.; DeConto, R.; Huber, M.; Brinkhuis, H.; Shah, S.R.; Leckie, R.M.; Pearson, A. Global cooling during the Eocene-Oligocene climate transition. Science 2009, 323, 1187–1190. [Google Scholar] [CrossRef] [Green Version]
- Hren, M.T.; Sheldon, N.D.; Grimes, S.T.; Collinson, M.E.; Hooker, J.J.; Bugler, M.; Lohmann, K.C. Terrestrial cooling in Northern Europe during the eocene-oligocene transition. Proc. Natl. Acad. Sci. USA 2013, 110, 7562–7567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colwyn, D.A.; Hren, M.T. An abrupt decrease in Southern Hemisphere terrestrial temperature during the Eocene–Oligocene transition. Earth Planet. Sci. Lett. 2019, 512, 227–235. [Google Scholar] [CrossRef]
- Westerhold, T.; Marwan, N.; Drury, A.J.; Liebrand, D.; Agnini, C.; Anagnostou, E.; Barnet, J.S.; Bohaty, S.M.; De Vleeschouwer, D.; Florindo, F. An astronomically dated record of Earth’s climate and its predictability over the last 66 million years. Science 2020, 369, 1383–1387. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.G.; Browning, J.V.; Schmelz, W.J.; Kopp, R.E.; Mountain, G.S.; Wright, J.D. Cenozoic sea-level and cryospheric evolution from deep-sea geochemical and continental margin records. Sci. Adv. 2020, 6, eaaz1346. [Google Scholar] [CrossRef]
- Hutchinson, D.K.; Coxall, H.K.; Lunt, D.J.; Steinthorsdottir, M.; de Boer, A.M.; Baatsen, M.; von der Heydt, A.; Huber, M.; Kennedy-Asser, A.T.; Kunzmann, L.; et al. The Eocene-Oligocene transition: A review of marine and terrestrial proxy data, models and model–data comparisons. Clim. Past 2021, 17, 269–315. [Google Scholar] [CrossRef]
- Berggren, W.A.; Prothero, D.R. Eocene-Oligocene Climatic and Biotic Evolution: An Overview. In Eocene-Oligocene Climatic and Biotic Evolution; Berggren, W.A., Prothero, D.R., Eds.; Princeton University Press: Princeton, NJ, USA, 1992; pp. 1–28. [Google Scholar]
- Dashzeveg, D. Asynchronism of the main mammalian faunal events near the Eocene-Oligocene boundary. Tert. Res. 1993, 14, 141–149. [Google Scholar]
- Prothero, D.R.; Heaton, T.H. Faunal stability during the early Oligocene climatic crash. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1996, 127, 257–283. [Google Scholar] [CrossRef]
- Meng, J.; McKenna, M.C. Faunal turnovers of Palaeogene mammals from the Mongolian Plateau. Nature 1998, 394, 364–367. [Google Scholar] [CrossRef]
- Hooker, J.J.; Collinson, M.E.; Sille, N.P. Eocene-Oligocene mammalian faunal turnover in the Hampshire Basin, UK: Calibration to the global time scale and the major cooling event. J. Geol. Soc. 2004, 161, 161–172. [Google Scholar] [CrossRef]
- Sun, J.; Ni, X.; Bi, S.; Wu, W.; Ye, J.; Meng, J.; Windley, B.F. Synchronous turnover of flora, fauna, and climate at the Eocene-Oligocene Boundary in Asia. Sci. Rep. 2014, 4, 7463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B. The Mid-Tertiary Ctenodactylidae (Rodentia, Mammalia) of Eastern and Central Asia. Bull. Am. Mus. Nat. Hist. 1997, 234, 1–88. [Google Scholar]
- Dashzėvėg, D.; Meng, J. New Eocene Ctenodactyloid Rodents from the Eastern Gobi Desert of Mongolia and a Phylogenetic Analysis of Ctenodactyloids Based on Dental Features. Am. Mus. Novit. 1998, 3246, 1–20. [Google Scholar]
- Vianey-Liaud, M.; Gomes Rodrigues, H.; Marivaux, L. A new Oligocene Ctenodactylinae (Rodentia: Mammalia) from Ulantatal (nei Mongol): New insight on the phylogenetic origins of the modern Ctenodactylidae. Zool. J. Linn. Soc. 2010, 160, 531–550. [Google Scholar] [CrossRef]
- Li, Q.; Meng, J. New Ctenodactyloid Rodents from the Erlian Basin, Nei Mongol, China, and the Phylogenetic Relationships of Eocene Asian Ctenodactyloids. Am. Mus. Novit. 2015, 3828, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Matthew, W.D.; Granger, W. Nine new rodents from the Oligocene of Mongolia. Am. Mus. Novit. 1923, 102, 1–10. [Google Scholar]
- Dashzeveg, D. [New data on the stratigraphy of middle Oligocène sédiments of the M.P.R.] Novyye dannyye o stratigrafii sredneoligotsenovkh otlozheniy MNR. Sovmest. Sov. Mong. Nauch. Issled. Geol. Exped. Tr. 1970, 2, 43. (In Russian) [Google Scholar]
- Kowalski, K. Middle Oligocene rodents from Mongolia. In Results of the Polish-Mongolian Palaeontological Expédition, Part V.; Kielan-Jaworowsk, Z., Ed.; Palaeontologia Polonica; Polish Academy of Sciences: Warsaw, Poland, 1974; Volume 30, pp. 147–178. [Google Scholar]
- Mellett, J.S. The Oligocene Hsanda Gol Formation, Mongolia: A revised faunal list. Am. Mus. Novit. 1968, 2318, 1–16. [Google Scholar]
- Daxner-Höck, G.; Höck, V.; Badamgarav, D.; Furtmüller, G.; Frank, W.; Montag, O.; Schmid, H.P. Cenozoic stratigraphy based on a sediment-basalt association in Central Mongolia as requirement for correlation across Central Asia. In Proceedings of the Biochronologie Mammalienne du Cénozoique en Europe et Domaines Reliés, Montpellier, France, 14 April 1997; pp. 163–176. [Google Scholar]
- Schmidt-Kittler, N.; Vianey-Liaud, M.; Marivaux, L. Oligocene-Miocene Vertebrates from the Valley of Lakes (Central Mongolia): Morphology, phylogenetic and stratigraphic implications 6. The Ctenodactylidae (Rodentia, Mammalia). Ann. Nat. Mus. Wien 2007, 108, 173–215. [Google Scholar]
- Daxner-Höck, G.; Badamgarav, D.; Erbajeva, M. Oligocene stratigraphy based on a sediment-basalt association in Central Mongolia (Taatsiin Gol and Taatsiin Tsagaan Nuur area, Valley of Lakes): Review of a Mongolian-Austrian project. Vertebr. PalAsiat. 2010, 48, 348. [Google Scholar]
- Daxner-Hock, G.; Badamgarav, D.; Barsbold, R.; Bayarmaa, B.; Erbajeva, M.; Gohlich, U.B.; Harzhauser, M.; Hock, E.; Hock, V.; Ichinnorov, N.; et al. Oligocene stratigraphy across the Eocene and Miocene boundaries in the Valley of Lakes (Mongolia). Paleobiodivers. Paleoenviron. 2017, 97, 111–218. [Google Scholar] [CrossRef] [Green Version]
- Oliver, A.; Sanisidro, O.; Baatarjav, B.; Niiden, I.; Daxner-Hock, G. Diversification rates in Ctenodactylidae (Rodentia, Mammalia) from Mongolia. Paleobiodivers. Paleoenviron. 2017, 97, 51–65. [Google Scholar] [CrossRef] [Green Version]
- Höck, V.; Daxner-Höck, G.; Schmid, H.P.; Badamgarav, D.; Frank, W.; Furtmüller, G.; Montag, O.; Barsbold, R.; Khand, Y.; Sodov, J. Oligocene-Miocene sediments, fossils and basalts from the Valley of Lakes (Central Mongolia): An integrated study. Mitt. Österr. Geol. Ges. 1999, 90, 83–125. [Google Scholar]
- Wang, B.; Chang, J.; Meng, X.; Chen, J. Stratigraphy of the Upper and Middle Oligocene of Qianlishan District, Nei Mongol (Inner Mongolia). Vertebr. PalAsiat. 1981, 19, 26–34. [Google Scholar]
- Huang, X. Middle Oligocene Ctenodactylids (Rodentia, Mammalia) of Ulantatal, Nei Mongol. Vertebr. PalAsiat. 1985, 23, 27–38. [Google Scholar]
- Wang, B.; Wang, P. Discovery of Early Medial Oligocene Mammalian Fauna From Kekeamu, Alxa Left Banner, Nei Mongol. Vertebr. PalAsiat. 1991, 29, 64–71. [Google Scholar]
- Vianey-Liaud, M.; Schmidt-Kittler, N.; Marivaux, L. The Ctenodactylidae (Rodentia) from the Oligocene of Ulantatal (Inner Mongolia, China). Palaeovertebrata 2006, 34, 111–206. [Google Scholar]
- Zhang, Z.; Liu, Y.; Wang, L.; Kaakinen, A.; Wang, J.; Mao, F.; Tong, Y. Lithostratigraphic context of Oligocene mammalian faunas from Ulantatal, Nei Mongol, China. Comptes Rendus Palevol 2016, 15, 903–910. [Google Scholar] [CrossRef]
- Wasiljeff, J.; Kaakinen, A.; Salminen, J.M.; Zhang, Z. Magnetostratigraphic constraints on the fossiliferous Ulantatal sequence in Inner Mongolia, China: Implications for Asian aridification and faunal turnover before the Eocene-Oligocene boundary. Earth Planet. Sci. Lett. 2020, 535, 116125. [Google Scholar] [CrossRef]
- Qiu, Z.; Wang, B.; Qiu, Z.; Xie, G.; Xie, J.; Wang, X. Recent advances in study of the Xianshuihe Formation in Lanzhou Basin. In Evidence for Evolution—Eassays in Honor of Prof. Chungchien Young on the Hundredth Anniversary of His Birth; Tong, Y., Zhang, Y., Wu, W., Li, J., Shi, L., Eds.; China Ocean Press: Beijing, China, 1997; pp. 177–192. [Google Scholar]
- Wu, W.; Meng, J.I.N.; Ye, J.I.E.; Ni, X. Propalaeocastor (Rodentia, Mammalia) from the Early Oligocene of Burqin Basin, Xinjiang. Am. Mus. Novit. 2004, 3461, 1–16. [Google Scholar] [CrossRef]
- Jie, Y.; Jin, M.; Wen-Yu, W.; Xi-Jun, N. Late Eocene-Early Oligocene Lithological and Biological Stratigraphy in the Burqin Region of Xinjiang. Vertebr. PalAsiat. 2005, 43, 49–60. [Google Scholar]
- Emry, R.J.; Lucas, S.G.; Tyutkova, L.; Wang, B. The Ergilian-Shandgolian (Eocene-Oligocene) transition in the Zaysan basin, Kazakhstan. Bull. Carnegie Mus. Nat. Hist. 1998, 34, 298–312. [Google Scholar]
- Wasiljeff, J.; Zhang, Z. Stratigraphical significance of Ulantatal sequence (Nei Mongol, China) in refining the latest Eocene and Oligocene terrestrial regional stages. Vertebr. PalAsiat. 2021, 60, 42–53. [Google Scholar] [CrossRef]
- Tang, X.; Feng, Q.; Li, D. Tectonic features and evolution of Bayanhot Basin in western Inner Mongolia. Oil Gas Geol. 1990, 2, 127–135. [Google Scholar]
- Wang, P.; Wang, B. Tertiary system and its fauna of Alxa area, Nei Mongol, China. Northwest Geosci. 1998, 19, 1–37. [Google Scholar]
- Zhao, J.; Zhou, Y.; Wang, K.; Li, X.; Ma, Z.; Ruan, Z.; Wang, D.; Xue, R. Provenance and paleogeography of Carboniferous–Permian strata in the Bayanhot Basin: Constraints from sedimentary records and detrital zircon geochronology. Geosci. Front. 2021, 12, 101088. [Google Scholar] [CrossRef]
- Wasiljeff, J.; Salminen, J.M.; Stenman, J.; Zhang, Z.; Kaakinen, A. Oligocene moisture variations as evidenced by an aeolian dust sequence in Inner Mongolia, China. Sci. Rep. 2022, 12, 5597. [Google Scholar] [CrossRef]
- Wible, J.R.; Wang, Y.; Li, C.; Dawson, M.R. Cranial Anatomy and Relationships of a New Ctenodactyloid (Mammalia, Rodentia) from the Early Eocene of Hubei Province, China. Ann. Carnegie Mus. 2005, 74, 91–150. [Google Scholar] [CrossRef]
- SPSS. IBM SPSS Statistics for Windows, Version 20.0; SPSS Inc. Ltd.: Chicago, IL, USA, 2013. [Google Scholar]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. Available online: https://palaeo-electronica.org/2001_1/past/issue1_01.htm (accessed on 5 April 2023).
- Grimm, E.C. CONISS: A FORTRAN 77 program for stratigraphically constrained cluster analysis by the method of incremental sum of squares. Comput. Geosci. 1987, 13, 13–35. [Google Scholar] [CrossRef]
- Grimm, E.C. Tilia and Tiliagraph; Springfield, Illinois State Museum: Springfield, IL, USA, 1991; pp. 1–101. [Google Scholar]
- Hammer, Ø.; Harper, D.A. Paleontological Data Analysis, 1st ed.; Blackwell Publishing: Oxford, UK, 2006; pp. 1–351. [Google Scholar]
- Bowdich, T.E. An Analysis of the Natural Classifications of Mammalia: For the Use of Students and Travellers; Printed by J. Smith: Paris, France, 1821; pp. 51–61. [Google Scholar]
- Tullberg, T. Ueber das System der Nagethiere: Eine Phylogenetische Studie; Akademische Buchdruckerei: Herborn, Germany, 1899. [Google Scholar]
- Gervais, M. Description ostéologique de l’Anomalurus et remarques sur la classification naturelle des rongeurs. Ann. Sci. Nat. Zool. 1853, 20, 238–246. [Google Scholar]
- Wang, B. The Ctenodactyloidea of Asia. Natl. Sci. Mus. Monogr. 1994, 8, 35–47. [Google Scholar]
- Shevyreva, N. New middle Oligocene rodents of Kazakhstan and Mongolia. Acad. Sci. USSR Trans. Paleontol. Inst. 1971, 130, 70–86. [Google Scholar]
- Shevyreva, N. New rodents (Rodentia, Mammalia) from the lower Oligocene of the Zaisan depression (Eastern Kazakhstan). Paleontol. Zhurnal 1994, 1, 111–126. [Google Scholar]
- Wang, B.; Qiu, Z. Discovery of early Oligocene mammalian fossils from Danghe area, Gansu, China. Vertebr. PalAsiat. 2004, 42, 130–143. [Google Scholar]
- Li, Q.; Wang, X.; Xie, G.; Yin, A. Oligocene-Miocene mammalian fossils from Hongyazi Basin and its bearing on tectonics of Danghe Nanshan in Northern Tibetan Plateau. PLoS ONE 2013, 8, e82816. [Google Scholar] [CrossRef] [Green Version]
- Wood, A.E. The evolution of the rodent family Ctenodactylidae. J. Palaeontol. Soc. India 1977, 20, 120–137. [Google Scholar]
- Gradstein, F.M.; Ogg, J.G.; Smith, A.G. A Geologic Time Scale 2004; Cambridge University Press: Cambridge, UK, 2004. [Google Scholar]
- Sun, R. Principles of Animal Ecology (Translated from Chinese); Beijing Normal University Publishing Group: Bejing, China, 2006; p. 386. [Google Scholar]
- Hadly, E.A. Influence of late-Holocene climate on northern Rocky Mountain mammals. Quat. Res. 1996, 46, 298–310. [Google Scholar] [CrossRef]
- Wagner, P.J.; Kosnik, M.A.; Lidgard, S. Abundance distributions imply elevated complexity of post-Paleozoic marine ecosystems. Science 2006, 314, 1289–1292. [Google Scholar] [CrossRef] [Green Version]
- Canales, M.L.; Henriques, M.H. Foraminifera from the Aalenian and the Bajocian GSSP (Middle Jurassic) of Murtinheira section (Cabo Mondego, West Portugal): Biostratigraphy and paleoenvironmental implications. Mar. Micropaleontol. 2008, 67, 155–179. [Google Scholar] [CrossRef] [Green Version]
- Western, D.; Behrensmeyer, A.K. Bone assemblages track animal community structure over 40 years in an African savanna ecosystem. Science 2009, 324, 1061–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elshanawany, R.; Mohamed, Y.; Ibrahim, M.I.A. Paleontological Records of Holocene Environmental Changes offshore, Egypt. J. Ocean Univ. China 2019, 18, 1382–1394. [Google Scholar] [CrossRef]
- Cole, S.R.; Wright, D.F.; Ausich, W.I.; Koniecki, J.M. Paleocommunity composition, relative abundance, and new camerate crinoids from the Brechin Lagerstätte (Upper Ordovician). J. Paleontol. 2020, 94, 1103–1123. [Google Scholar] [CrossRef]
- Miller, K.G.; Wright, J.D.; Fairbanks, R.G. Unlocking the Ice House: Oligocene-Miocene oxygen isotopes, eustasy, and margin erosion. J. Geophys. Res. Solid Earth 1991, 96, 6829–6848. [Google Scholar] [CrossRef]
- Coxall, H.K.; Wilson, P.A.; Pälike, H.; Lear, C.H.; Backman, J. Rapid stepwise onset of Antarctic glaciation and deeper calcite compensation in the Pacific Ocean. Nature 2005, 433, 53–57. [Google Scholar] [CrossRef]
- Harzhauser, M.; Daxner-Hock, G.; Lopez-Guerrero, P.; Maridet, O.; Oliver, A.; Piller, W.E.; Richoz, S.; Erbajeva, M.A.; Neubauer, T.A.; Gohlich, U.B. Stepwise onset of the Icehouse world and its impact on Oligo-Miocene Central Asian mammals. Sci. Rep. 2016, 6, 36169. [Google Scholar] [CrossRef] [Green Version]
- Richoz, S.; Baldermann, A.; Frauwallner, A.; Harzhauser, M.; Daxner-Hock, G.; Klammer, D.; Piller, W.E. Geochemistry and mineralogy of the Oligo-Miocene sediments of the Valley of Lakes, Mongolia. Paleobiodivers. Paleoenviron. 2017, 97, 233–258. [Google Scholar] [CrossRef] [Green Version]
- Hopkins, S.S.B. Estimation of body size in fossil mammals. In Methods in Paleoecology: Reconstructing Cenozoic Terrestrial Environments and Ecological Communities; Croft, D.A., Su, D.F., Simpson, S.W., Eds.; Springer: Cham, Switzerland, 2018; pp. 7–22. [Google Scholar]
- Barnosky, A.D. Distinguishing the effects of the Red Queen and Court Jester on Miocene mammal evolution in the northern Rocky Mountains. J. Vertebr. Paleontol. 2001, 21, 172–185. [Google Scholar] [CrossRef]
- Žliobaitė, I.; Fortelius, M.; Stenseth, N.C. Reconciling taxon senescence with the Red Queen’s hypothesis. Nature 2017, 552, 92–95. [Google Scholar] [CrossRef] [Green Version]
- Ladant, J.-B.; Donnadieu, Y.; Lefebvre, V.; Dumas, C. The respective role of atmospheric carbon dioxide and orbital parameters on ice sheet evolution at the Eocene-Oligocene transition. Paleoceanography 2014, 29, 810–823. [Google Scholar] [CrossRef]
- Ao, H.; Dupont-Nivet, G.; Rohling, E.J.; Zhang, P.; Ladant, J.B.; Roberts, A.P.; Licht, A.; Liu, Q.; Liu, Z.; Dekkers, M.J.; et al. Orbital climate variability on the northeastern Tibetan Plateau across the Eocene-Oligocene transition. Nat. Commun. 2020, 11, 5249. [Google Scholar] [CrossRef] [PubMed]
- Gomes Rodrigues, H.; Marivaux, L.; Vianey-Liaud, M. Expansion of open landscapes in Northern China during the Oligocene induced by dramatic climate changes: Paleoecological evidence. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2012, 358–360, 62–71. [Google Scholar] [CrossRef]
- Sousa, F.V. Biogeography and Phylogenetic Position of a Sahara-Sahel Mountain Endemic, Felovia Vae (Ctenodactylidae). Master’s Dissertation, University of Porto, Porto, Portugal, 2015. [Google Scholar]
- Xu, R.; Li, Q. New skulls of ctenodactyloids from the Early Oligocene of Ulantatal, Nei Mongol, China. Vertebr. PalAsiat. 2020, 58, 305–327. [Google Scholar]
- Antoine, P.-O.; Yans, J.; Castillo, A.A.; Stutz, N.; Abello, M.A.; Adnet, S.; Custódio, M.A.; Benites-Palomino, A.; Billet, G.; Boivin, M.; et al. Biotic community and landscape changes around the Eocene–Oligocene transition at Shapaja, Peruvian Amazonia: Regional or global drivers? Glob. Planet. Chang. 2021, 202, 103512. [Google Scholar] [CrossRef]
- Butler, P. Homologies of molar cusps and crests, and their bearing on assessments of rodent phylogeny. In Evolutionary Relationships among Rodents: A Multidisciplinary Analysis; Springer: New York, NY, USA, 1985; pp. 381–401. [Google Scholar]
- Ferrusquia-Villafranca, I.; Flynn, L.J.; Ruiz-Gonzalez, J.E.; Torres-Hernandez, J.R.; Martinez-Hernandez, E. New Eocene rodents from northwestern Oaxaca, southeastern Mexico, and their paleobiological significance. J. Vertebr. Paleontol. 2018, 38, e1514615. [Google Scholar] [CrossRef]
- Angelone, C.; Zhang, Z. Climate change and evolution of early lagomorphs (Mammalia): A study perspective based on new materials of Ordolagus from Nei Mongol (northern China). Vertebr. PalAsiat. 2021, 59, 138. [Google Scholar]
- Lukens, W.E.; Stinchcomb, G.E.; Nordt, L.C.; Kahle, D.J.; Driese, S.G.; Tubbs, J.D. Recursive partitioning improves paleosol proxies for rainfall. Am. J. Sci. 2019, 319, 819–845. [Google Scholar] [CrossRef]
- Fischer, R.; Turner, N.C. Plant productivity in the arid and semiarid zones. Annu. Rev. Plant Physiol. 1978, 29, 277–317. [Google Scholar] [CrossRef]
- Zheng, S.; Shangguan, Z. Spatial patterns of foliar stable carbon isotope compositions of C3 plant species in the Loess Plateau of China. Ecol. Res. 2006, 22, 342–353. [Google Scholar] [CrossRef]
- Zhao, X.; Tan, K.; Zhao, S.; Fang, J. Changing climate affects vegetation growth in the arid region of the northwestern China. J. Arid Environ. 2011, 75, 946–952. [Google Scholar] [CrossRef]
- Peng, J.; Dong, W.; Yuan, W.; Zhang, Y. Responses of grassland and forest to temperature and precipitation changes in Northeast China. Adv. Atmos. Sci. 2012, 29, 1063–1077. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | K. decessus | K. arcanus | K. chelkaris | K. conjunctus sp. nov. | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | Range | M | SD | N | Range | M | SD | N | Range | M | SD | N | Range | M | SD | ||
| DP3 | L | 3 | 0.40–0.44 | 0.43 | 0.019 | 0 | / | / | / | 0 | / | / | / | 2 | / | 0.44 | 0.00 |
| DP3 | W | 3 | 0.48–0.60 | 0.53 | 0.052 | 0 | / | / | / | 0 | / | / | / | 2 | 0.54–0.56 | 0.55 | 0.010 |
| DP4 | L | 6 | 1.30–1.50 | 1.40 | 0.069 | 3 | 1.10–1.46 | 1.32 | 0.157 | 0 | / | / | / | 4 | 1.32–1.48 | 1.40 | 0.057 |
| DP4 | W | 6 | 1.38–1.50 | 1.44 | 0.040 | 3 | 1.20–1.54 | 1.42 | 0.156 | 0 | / | / | / | 4 | 1.38–1.60 | 1.47 | 0.082 |
| P4 | L | 16 | 0.96–1.36 | 1.22 | 0.091 | 7 | 0.86–1.28 | 1.08 | 0.130 | 9 | 0.60–0.98 | 0.83 | 0.099 | 3 | 1.20–1.24 | 1.21 | 0.019 |
| P4 | W | 16 | 1.42–1.72 | 1.59 | 0.091 | 7 | 1.32–1.66 | 1.50 | 0.113 | 9 | 0.92–1.20 | 1.03 | 0.082 | 3 | 1.52–1.54 | 1.53 | 0.009 |
| M1 | L | 28 | 1.56–1.90 | 1.72 | 0.095 | 9 | 1.40–1.84 | 1.60 | 0.140 | 10 | 0.94–1.30 | 1.14 | 0.117 | 9 | 1.60–1.90 | 1.74 | 0.085 |
| M1 | W | 28 | 1.56–2.06 | 1.84 | 0.108 | 9 | 1.40–1.90 | 1.66 | 0.165 | 10 | 1.14–1.36 | 1.25 | 0.066 | 9 | 1.70–1.94 | 1.84 | 0.078 |
| M2 | L | 25 | 1.70–2.20 | 1.86 | 0.124 | 17 | 1.52–2.00 | 1.75 | 0.107 | 11 | 1.12–1.44 | 1.31 | 0.108 | 10 | 1.82–2.25 | 1.97 | 0.131 |
| M2 | W | 25 | 1.78–2.30 | 2.02 | 0.116 | 17 | 1.66–2.15 | 1.82 | 0.121 | 11 | 1.26–1.54 | 1.40 | 0.083 | 10 | 1.92–2.65 | 2.09 | 0.202 |
| M3 | L | 13 | 1.56–1.96 | 1.74 | 0.118 | 10 | 1.40–1.90 | 1.70 | 0.163 | 10 | 1.20–1.44 | 1.33 | 0.073 | 8 | 1.76–1.96 | 1.89 | 0.071 |
| M3 | W | 13 | 1.70–2.06 | 1.85 | 0.090 | 10 | 1.46–2.04 | 1.75 | 0.203 | 10 | 1.30–1.50 | 1.38 | 0.060 | 8 | 1.94–2.00 | 1.97 | 0.021 |
| dp4 | L | 6 | 1.34–1.46 | 1.40 | 0.045 | 3 | 1.30–1.40 | 1.34 | 0.043 | 1 | / | 1.08 | / | 2 | 1.30–1.52 | 1.41 | 0.110 |
| dp4 | W | 6 | 0.96–1.20 | 1.08 | 0.071 | 3 | 1.00–1.06 | 1.03 | 0.025 | 1 | / | 0.76 | / | 2 | 0.94–1.08 | 1.01 | 0.070 |
| p4 | L | 18 | 1.16–1.40 | 1.25 | 0.063 | 6 | 1.00–1.20 | 1.16 | 0.073 | 12 | 0.78–0.96 | 0.88 | 0.051 | 9 | 1.16–1.36 | 1.24 | 0.066 |
| p4 | W | 18 | 0.94–1.26 | 1.10 | 0.090 | 6 | 0.86–1.16 | 1.02 | 0.097 | 12 | 0.72–0.90 | 0.80 | 0.053 | 9 | 0.96–1.10 | 1.04 | 0.049 |
| m1 | L | 33 | 1.60–1.90 | 1.75 | 0.071 | 25 | 1.44–1.84 | 1.65 | 0.108 | 22 | 1.16–1.40 | 1.27 | 0.067 | 24 | 1.60–1.70 | 1.79 | 0.098 |
| m1 | W | 33 | 1.24–1.92 | 1.52 | 0.112 | 25 | 1.24–1.60 | 1.36 | 0.080 | 22 | 0.98–1.24 | 1.10 | 0.070 | 24 | 1.44–1.56 | 1.55 | 0.077 |
| m2 | L | 42 | 1.64–2.30 | 1.99 | 0.140 | 22 | 1.56–2.00 | 1.83 | 0.105 | 24 | 1.20–1.50 | 1.39 | 0.074 | 32 | 1.76–2.20 | 2.02 | 0.112 |
| m2 | W | 42 | 1.52–2.00 | 1.77 | 0.118 | 22 | 1.44–1.80 | 1.62 | 0.101 | 24 | 1.12–1.40 | 1.30 | 0.073 | 32 | 1.66–2.06 | 1.83 | 0.091 |
| m3 | L | 25 | 1.80–2.25 | 2.04 | 0.129 | 11 | 1.66–2.12 | 1.91 | 0.149 | 18 | 1.30–1.54 | 1.42 | 0.076 | 22 | 1.90–2.40 | 2.10 | 0.112 |
| m3 | W | 25 | 1.46–1.80 | 1.63 | 0.102 | 11 | 1.30–1.70 | 1.49 | 0.131 | 18 | 1.04–1.32 | 1.19 | 0.069 | 22 | 1.50–1.92 | 1.66 | 0.092 |
| SS | MS | df | F | p | ||
|---|---|---|---|---|---|---|
| M2L | Between Groups | 3.576 | 1.192 | 3 | 84.447 | <0.001 |
| Within Groups | 0.889 | 0.014 | 63 | |||
| M2W | Between Groups | 3.480 | 1.160 | 3 | 64.559 | <0.001 |
| Within Groups | 1.060 | 0.018 | 59 | |||
| m2l | Between Groups | 7.206 | 2.402 | 3 | 179.626 | <0.001 |
| Within Groups | 1.631 | 0.013 | 122 | |||
| m2w | Between Groups | 4.489 | 1.496 | 3 | 145.565 | <0.001 |
| Within Groups | 1.193 | 0.010 | 116 |
| K. arcanus | K. chelkaris | K. decessus | K. conjunctus sp. nov. | |
|---|---|---|---|---|
| K. arcanus | M2L, M2W, m2l, m2w | M2L, M2W, m2l, m2w | M2L, M2W, m2l, m2w | |
| K. chelkaris | M2L, M2W, m2l, m2w | M2L, M2W, m2l, m2w | M2L, M2W, m2l, m2w | |
| K. decessus | M2L, M2W, m2l, m2w | M2L, M2W, m2l, m2w | M2L, M2W, m2l, m2w | |
| K. conjunctus sp. nov. | M2L, M2W, m2l, m2w | M2L, M2W, m2l, m2w | M2L, M2W, m2l, m2w |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, R.; Zhang, Z.; Li, Q.; Wang, B. New Material of Karakoromys (Ctenodactylidae, Rodentia) from Late Eocene-Early Oligocene of Ulantatal (Nei Mongol): Taxonomy, Diversity, and Response to Climatic Change. Diversity 2023, 15, 744. https://doi.org/10.3390/d15060744
Xu R, Zhang Z, Li Q, Wang B. New Material of Karakoromys (Ctenodactylidae, Rodentia) from Late Eocene-Early Oligocene of Ulantatal (Nei Mongol): Taxonomy, Diversity, and Response to Climatic Change. Diversity. 2023; 15(6):744. https://doi.org/10.3390/d15060744
Chicago/Turabian StyleXu, Rancheng, Zhaoqun Zhang, Qian Li, and Bian Wang. 2023. "New Material of Karakoromys (Ctenodactylidae, Rodentia) from Late Eocene-Early Oligocene of Ulantatal (Nei Mongol): Taxonomy, Diversity, and Response to Climatic Change" Diversity 15, no. 6: 744. https://doi.org/10.3390/d15060744
APA StyleXu, R., Zhang, Z., Li, Q., & Wang, B. (2023). New Material of Karakoromys (Ctenodactylidae, Rodentia) from Late Eocene-Early Oligocene of Ulantatal (Nei Mongol): Taxonomy, Diversity, and Response to Climatic Change. Diversity, 15(6), 744. https://doi.org/10.3390/d15060744