

Spatiotemporal Variability in Subarctic Lithothamnion glaciale Rhodolith Bed Structural Complexity and Macrofaunal Diversity


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Rhodolith Bed Structure
2.2.1. Bed Composition
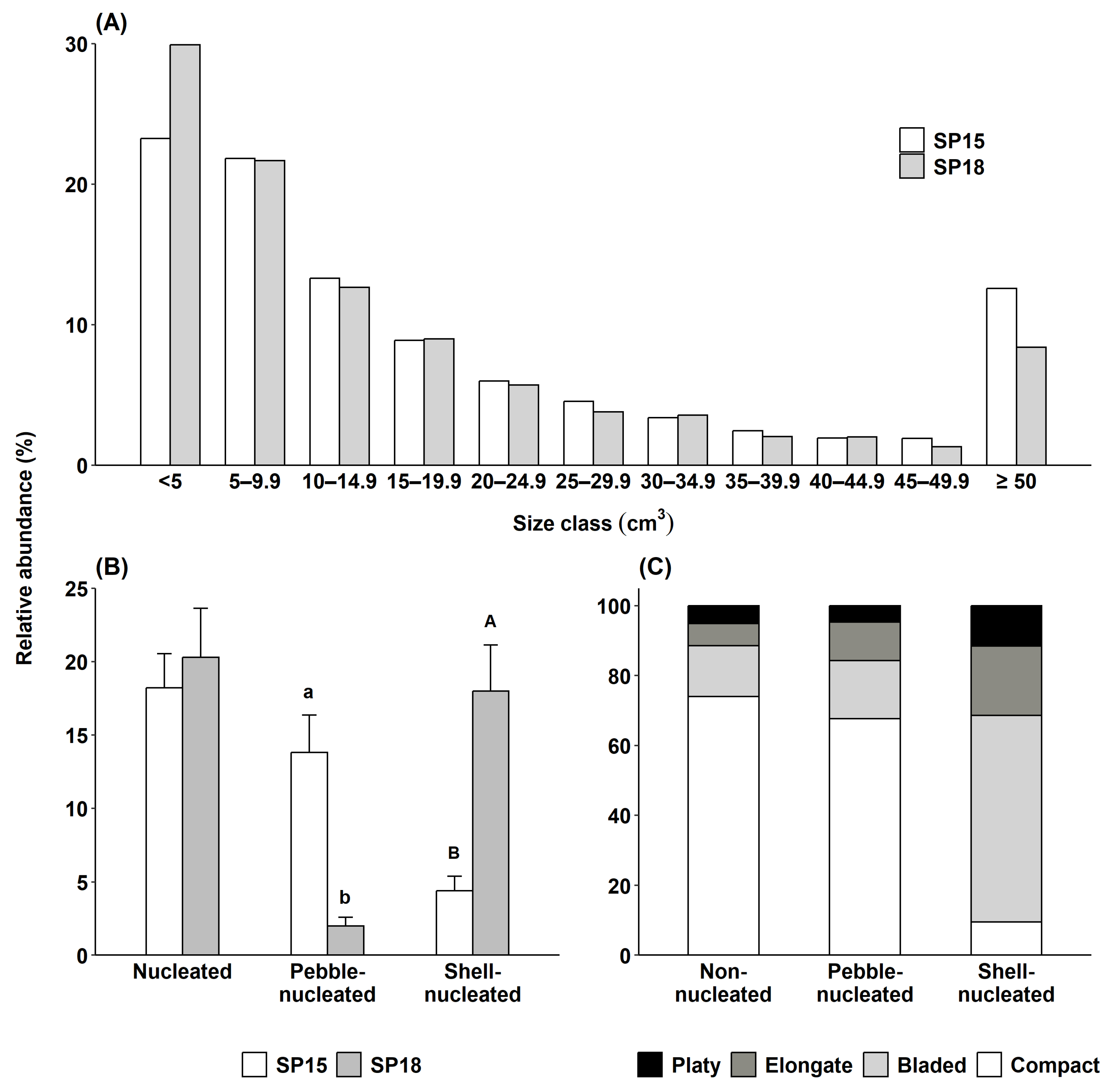
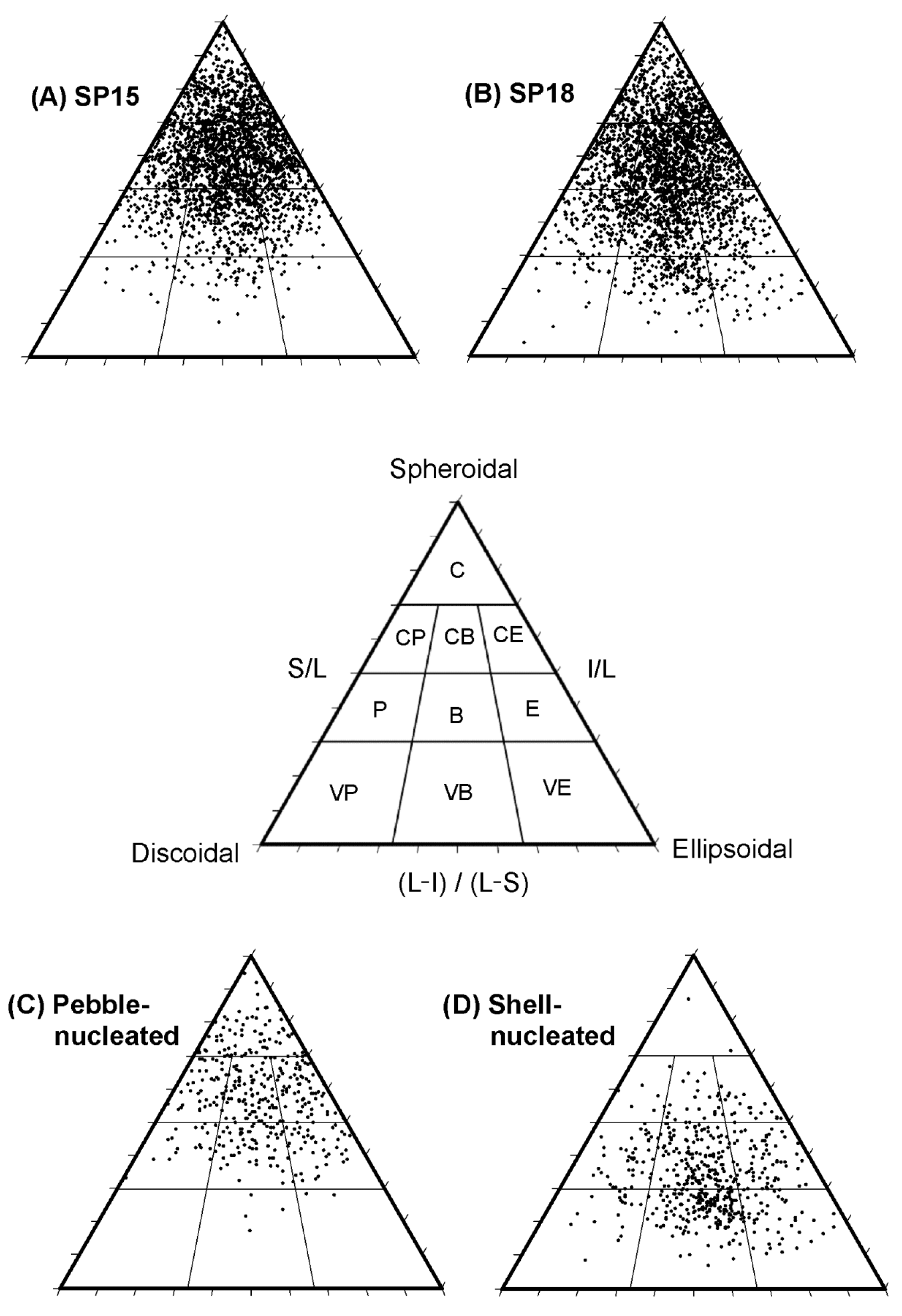
2.2.2. Rhodolith Abundance and Morphology
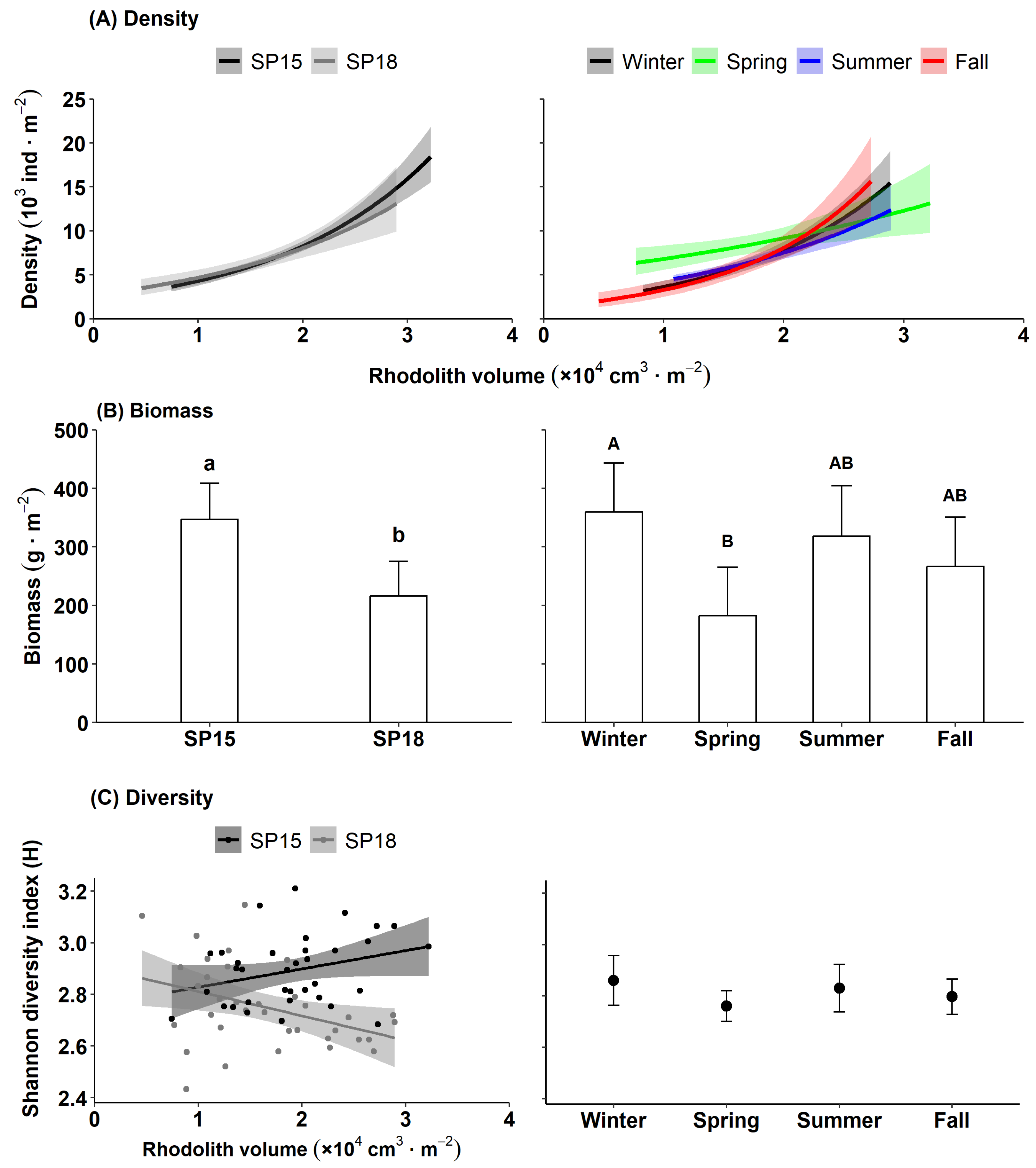
2.3. Macrofaunal Density, Biomass, and Diversity
2.4. Statistical Analysis
2.4.1. Rhodolith Bed Structure
2.4.2. Macrofaunal Density, Biomass, and Diversity
3. Results
3.1. Rhodolith Bed Habitat Structure
3.1.1. Seafloor Composition
3.1.2. Rhodolith Abundance and Morphology
3.2. Rhodolith Macrofauna
4. Discussion
4.1. Rhodolith Bed Structure
4.2. Rhodolith Macrofauna
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McCoy, E.D.; Bell, S.S. Habitat structure: The evolution and diversification of a complex topic. In Habitat Structure; Springer: Dordrecht, The Netherlands, 1991; pp. 3–27. [Google Scholar]
- Byrne, L.B. Habitat Structure: A fundamental concept and framework for urban soil ecology. Urban Ecosyst. 2007, 10, 255–274. [Google Scholar] [CrossRef] [Green Version]
- de Camargo, N.F.; Sano, N.Y.; Vieira, E.M. Forest vertical complexity affects alpha and beta diversity of small mammals. J. Mammal. 2018, 99, 1444–1454. [Google Scholar] [CrossRef]
- Garden, J.G.; Mcalpine, C.A.; Possingham, H.P.; Jones, D.N. Habitat structure is more important than vegetation composition for local-level management of native terrestrial reptile and small mammal species living in urban remnants: A case study from Brisbane, Australia. Austral. Ecol. 2007, 32, 669–685. [Google Scholar] [CrossRef] [PubMed]
- Zimbres, B.; Peres, C.A.; Machado, R.B. Terrestrial mammal responses to habitat structure and quality of remnant riparian forests in an Amazonian cattle-ranching landscape. Biol. Conserv. 2017, 206, 283–292. [Google Scholar] [CrossRef] [Green Version]
- Beck, M.W. Comparison of the measurement and effects of habitat structure on gastropods in rocky intertidal and mangrove habitats. Mar. Ecol. Prog. Ser. 1998, 169, 165–178. [Google Scholar] [CrossRef]
- Buhl-Mortensen, L.; Buhl-Mortensen, P.; Dolan, M.F.J.; Dannheim, J.; Bellec, V.; Holte, B. Habitat complexity and bottom fauna composition at different scales on the continental shelf and slope of northern Norway. Hydrobiologia 2012, 685, 191–219. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, L.R.S.; Loiola, M.; Barros, F. Manipulating habitat complexity to understand its influence on benthic macrofauna. J. Exp. Mar. Biol. Ecol. 2017, 489, 48–57. [Google Scholar] [CrossRef]
- Preston, F.W. Time and space and the variation of species. Ecology 1960, 41, 612–627. [Google Scholar] [CrossRef]
- MacArthur, R.H.; Wilson, E.O. (Eds.) The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967. [Google Scholar]
- Connor, E.F.; McCoy, E.D. The statistics and biology of the species-area relationship. Am. Nat. 1979, 113, 791–833. [Google Scholar] [CrossRef]
- Dean, R.L.; Connell, J.H. Marine invertebrates in an algal succession. III. Mechanisms linking habitat complexity with diversity. J. Exp. Mar. Biol. Ecol. 1987, 109, 249–273. [Google Scholar] [CrossRef]
- Hixon, M.A.; Menge, B.A. Species diversity: Prey refuges modify the interactive effects of predation and competition. Theor. Popul. Biol. 1991, 39, 178–200. [Google Scholar] [CrossRef]
- Myhre, L.C.; Forsgren, E.; Amundsen, T. Effects of habitat complexity on mating behavior and mating success in a marine fish. Behav. Ecol. 2013, 24, 553–563. [Google Scholar] [CrossRef]
- Bell, S.S.; McCoy, E.D.; Mushinsky, H.R. (Eds.) Habitat Structure: The Physical Arrangement of Objects in Space; Population and Community Biology Series; Springer: Dordrecht, The Netherlands, 1991; Volume 8. [Google Scholar]
- Jones, C.G.; Lawton, J.H.; Shachak, M. Organisms as ecosystem engineers. Oikos 1994, 69, 373. [Google Scholar] [CrossRef]
- Buhl-Mortensen, L.; Vanreusel, A.; Gooday, A.J.; Levin, L.A.; Priede, I.G.; Buhl-Mortensen, P.; Gheerardyn, H.; King, N.J.; Raes, M. Biological structures as a source of habitat heterogeneity and biodiversity on the deep ocean margins. Mar. Ecol. 2010, 31, 21–50. [Google Scholar] [CrossRef]
- Coggan, N.V.; Hayward, M.W.; Gibb, H. A global database and “state of the field” review of research into ecosystem engineering by land animals. J. Anim. Ecol. 2018, 87, 974–994. [Google Scholar] [CrossRef] [PubMed]
- Curd, A.; Pernet, F.; Corporeau, C.; Delisle, L.; Firth, L.B.; Nunes, F.L.D.; Dubois, S.F. Connecting Organic to Mineral: How the physiological state of an ecosystem-engineer is linked to its habitat structure. Ecol. Indic. 2019, 98, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Steneck, R.S.; Graham, M.H.; Bourque, B.J.; Corbett, D.; Erlandson, J.M.; Estes, J.A.; Tegner, M.J. Kelp forest ecosystems: Biodiversity, stability, resilience and future. Environ. Conserv. 2002, 29, 436–459. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, B.K.; Rubach, A.; Hillebrand, H. Biotic habitat complexity controls species diversity and nutrient effects on net biomass production. Ecology 2006, 87, 246–254. [Google Scholar] [CrossRef] [Green Version]
- Hauser, A.; Attrill, M.J.; Cotton, P.A. Effects of habitat complexity on the diversity and abundance of macrofauna colonising artificial kelp holdfasts. Mar. Ecol. Prog. Ser. 2006, 325, 93–100. [Google Scholar] [CrossRef]
- Koivisto, M.E.; Westerbom, M. Habitat structure and complexity as determinants of biodiversity in blue mussel beds on sublittoral rocky shores. Mar. Biol. 2010, 157, 1463–1474. [Google Scholar] [CrossRef]
- Adey, W.H.; Macintyre, I.G. Crustose coralline algae: A re-evaluation in the geological sciences. Geol. Soc. Am. Bull. 1973, 84, 883–904. [Google Scholar] [CrossRef]
- Bosence, D.W. Description and Classification of Rhodoliths (Rhodoids, Rhodolites). In Coated Grains; Peryt, T.M., Ed.; Springer: Berlin/Heidelberg, Germany, 1983; pp. 217–224. [Google Scholar] [CrossRef]
- Foster, M.S. Rhodoliths: Between rocks and soft places. J. Phycol. 2001, 37, 659–667. [Google Scholar] [CrossRef]
- Bosence, D.W.J. Coralline Algal Reef Frameworks. J. Geol. Soc. 1983, 140, 365–376. [Google Scholar] [CrossRef]
- Nelson, W.A. Calcified macroalgae critical to coastal ecosystems and vulnerable to change: A review. Mar. Freshw. Res. 2009, 60, 787–801. [Google Scholar] [CrossRef]
- Woelkerling, W.J.; Irvine, L.M.; Harvey, A.S. Growth-forms in non-geniculate coralline red algae (Coralliinales, Rhodophyta). Aust. Syst. Bot. 1993, 6, 277–293. [Google Scholar] [CrossRef]
- Gagnon, P.; Matheson, K.; Stapleton, M. Variation in rhodolith morphology and biogenic potential of newly discovered rhodolith beds in Newfoundland and Labrador (Canada). Bot. Mar. 2012, 55, 85–99. [Google Scholar] [CrossRef]
- Frantz, B.R.; Kashgarian, M.; Coale, K.H.; Foster, M.S. Growth rate and potential climate record from a rhodolith using 14C accelerator mass spectrometry. Limnol. Oceanogr. 2000, 45, 1773–1777. [Google Scholar] [CrossRef]
- Teed, L.; Bélanger, D.; Gagnon, P.; Edinger, E. Calcium carbonate (CaCO3) production of a subpolar rhodolith bed: Methods of estimation, effect of bioturbators, and global comparisons. Estuar. Coast. Shelf Sci. 2020, 242, 106822. [Google Scholar] [CrossRef]
- Bélanger, D.; Gagnon, P. High growth resilience of subarctic rhodoliths (Lithothamnion glaciale) to ocean warming and chronic low irradiance. Mar. Ecol. Prog. Ser. 2021, 663, 77–97. [Google Scholar] [CrossRef]
- Bélanger, D.; Gagnon, P. Low growth resilience of subarctic rhodoliths (Lithothamnion glaciale) to coastal eutrophication. Mar. Ecol. Prog. Ser. 2020, 642, 117–132. [Google Scholar] [CrossRef]
- Arnold, C.L.; Bélanger, D.; Gagnon, P. Growth resilience of subarctic rhodoliths (Lithothamnion glaciale, Rhodophyta) to chronic low sea temperature and irradiance. J. Phycol. 2022, 58, 251–266. [Google Scholar] [CrossRef] [PubMed]
- Millar, K.R.; Gagnon, P. Mechanisms of stability of rhodolith beds: Sedimentological aspects. Mar. Ecol. Prog. Ser. 2018, 594, 65–83. [Google Scholar] [CrossRef] [Green Version]
- Freiwald, A.; Henrich, R. Reefal coralline algal build-ups within the Arctic Circle: Morphology and sedimentary dynamics under extreme environmental seasonality. Sedimentology 1994, 41, 963–984. [Google Scholar] [CrossRef]
- Ballantine, D.L.; Bowden-Kerby, A.; Aponte, N.E. Cruoriella rhodoliths from shallow-water back reef environments in La Parguera, Puerto Rico (Caribbean Sea). Coral Reefs 2000, 19, 75–81. [Google Scholar] [CrossRef]
- Teichert, S. Hollow rhodoliths increase svalbard’s shelf biodiversity. Sci. Rep. 2014, 4, 6972. [Google Scholar] [CrossRef] [Green Version]
- Hinojosa-Arango, G.; Riosmena-Rodríguez, R. Influence of rhodolith-forming species and growth-form on associated fauna of rhodolith beds in the central-west Gulf of California, México. Mar. Ecol. 2004, 25, 109–127. [Google Scholar] [CrossRef]
- Gabara, S.S.; Hamilton, S.L.; Edwards, M.S.; Steller, D.L. Rhodolith structural loss decreases abundance, diversity, and stability of benthic communities at Santa Catalina Island, CA. Mar. Ecol. Prog. Ser. 2018, 595, 71–88. [Google Scholar] [CrossRef] [Green Version]
- Bracchi, V.A.; Meroni, A.N.; Epis, V.; Basso, D. Mollusk thanatocoenoses unravel the diversity of heterogeneous rhodolith beds (Italy, Tyrrhenian Sea). Diversity 2023, 15, 526. [Google Scholar] [CrossRef]
- Kamenos, N.A.; Moore, P.G.; Hall-Spencer, J.M. Attachment of the juvenile queen scallop (Aequipecten opercularis (L.)) to maerl in mesocosm conditions; juvenile habitat selection. J. Exp. Mar. Biol. Ecol. 2004, 306, 139–155. [Google Scholar] [CrossRef]
- Steller, D.L.; Cáceres-Martínez, C. Coralline algal rhodoliths enhance larval settlement and early growth of the Pacific calico scallop Argopecten ventricosus. Mar. Ecol. Prog. Ser. 2009, 396, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Pearce, C.M.; Scheibling, R.E. Induction of metamorphosis of larvae of the green sea urchin, Strongylocentrotus droebachiensis, by coralline red algae. Biol. Bull. 1990, 179, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Heyward, A.J.; Negri, A.P. Natural inducers for coral larval metamorphosis. Coral Reefs 1999, 18, 273–279. [Google Scholar] [CrossRef]
- Whalan, S.; Webster, N.S.; Negri, A.P. Crustose coralline algae and a cnidarian neuropeptide trigger larval settlement in two coral reef sponges. PLoS ONE 2012, 7, e30386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adey, W.H. Distribution of saxicolous crustose corallines in the northwestern North Atlantic. J. Phycol. 1966, 2, 49–54. [Google Scholar] [CrossRef]
- Adey, W.H.; Hayek, L.-A.C. Elucidating marine biogeography with macrophytes: Quantitative analysis of the North Atlantic supports the thermogeographic model and demonstrates a distinct subarctic region in the northwestern Atlantic. Northeast. Nat. 2011, 18, 1–128. [Google Scholar] [CrossRef]
- Hacker Teper, S.; Parrish, C.C.; Gagnon, P. Multiple trophic tracer analyses of subarctic rhodolith (Lithothamnion glaciale) bed trophodynamics uncover bottom-up forcing and benthic-pelagic coupling. Front. Mar. Sci. 2022, 9, 1–20. [Google Scholar] [CrossRef]
- Basso, D. Deep rhodolith distribution in the Pontian Islands, Italy: A model for the paleoecology of a temperate sea. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1998, 137, 173–187. [Google Scholar] [CrossRef]
- Wentworth, C.K. A scale of grade and class terms for clastic sediments. J. Geol. 1922, 30, 377–392. [Google Scholar] [CrossRef]
- Graham, D.J.; Midgley, N.G. Graphical representation of particle shape using triangular diagrams: And excel spreadsheet method. Earth Surf. Proc. Land. 2000, 25, 1473–1477. [Google Scholar] [CrossRef]
- Sneed, E.D.; Folk, R.L. Pebbles in the lower Colorado River, Texas a study in particle morphogenesis. J. Geol. 1958, 66, 114–150. [Google Scholar] [CrossRef]
- Abbott, R.T. American Seashells; the Marine Mollusca of the Atlantic and Pacific Coasts of North America, 2nd ed.; Van Nostrand Reinhold: New York, NY, USA, 1974; ISBN 0442202288. [Google Scholar]
- Pettibone, M.H. Marine Polychaete Worms of the New England Region: I. Families Aphroditidae through Trochocaetidae; Bulletin of the United States National Museum: Washington, DC, USA, 1963; pp. 1–356. [Google Scholar]
- Fauchald, K. The Polychaete Worms. Definitions and Keys to the Orders, Families and Genera; Science Series; Natural History Museum of Los Angeles County: Los Angeles, CA, USA, 1977; Volume 28, pp. 1–188. [Google Scholar]
- Appy, T.D.; Linkletter, L.E.; Dadswell, M.J. Annelida: Polychaeta: A Guide to the Marine Flora and Fauna of the Bay of Fundy; Canada Fisheries and Marine Service Technical Report; Government of Canada: St. Andrews, NB, Canada, 1980; p. 920. [Google Scholar]
- Pocklington, P. Polychaetes of Eastern Canada: An Illustrated Key to Polychaetes of Eastern Canada including the Eastern Arctic; Scientific Monograph Funded by Ocean Dumping Control Act Research Fund; National Museums of Canada & Department of Fisheries and Oceans: Mont-Joli, QC, Canada, 1989. [Google Scholar]
- Gosner, K.L. Atlantic Seashore: A Field Guide to Sponges, Jellyfish, Sea Urchins, and More; Houghton Mifflin Harcourt: Boston, MA, USA, 2014; ISBN 0544530853. [Google Scholar]
- Pollock, L.W. A Practical Guide to the Marine Animals of Northeastern North America; Rutgers University Press: New Brunswick, MA, USA, 1998; ISBN 0813523990. [Google Scholar]
- Abbott, R.T.; Morris, P.A. Shells of the Atlantic and Gulf Coasts and the West Indies, 4th ed.; Houghton Mifflin Harcourt: Boston, MA, USA, 2001; ISBN 0442202288. [Google Scholar]
- Squires, H.J. Decapod Crustacea of the Atlantic Coast of Canada (Canadian Bulletin Fisheries Aquatic Sciences); Department of Fisheries and Oceans Canada: Ottawa, ON, Canada, 1990; Volume 221. [Google Scholar]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press: New York, NY, USA, 2002; ISBN 0521009766. [Google Scholar]
- Snedecor, G.; Cochran, W. Statistical Methods, 8th ed.; Iowa State University Press: Ames, IA, USA, 1994; ISBN 978-0-813-81561-9. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.r-project.org/ (accessed on 14 February 2022).
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002; ISBN 0-387-95457-0. Available online: https://www.stats.ox.ac.uk/pub/MASS4/ (accessed on 25 December 2022).
- Oksanen, J.; Simpson, G.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.; O’hara, R.; Solymos, P.; Stevens, M.; Szoecs, E.; et al. Vegan: Community Ecology Package, Version 2.6-4. 2022. Available online: https://CRAN.R-project.org/package=vegan (accessed on 1 January 2023).
- Piller, W.E.; Rasser, M. Rhodolith formation induced by reef erosion in the Red Sea, Egypt. Coral Reefs 1996, 15, 191–198. [Google Scholar] [CrossRef]
- Del Río, J.; Ramos, D.A.; Sánchez-Tocino, L.; Peñas, J.; Braga, J.C. The Punta de La Mona rhodolith bed: Shallow-water Mediterranean rhodoliths (Almuñecar, Granada, southern Spain). Front. Earth Sci. 2022, 10, 884685. [Google Scholar] [CrossRef]
- Teichert, S.; Woelkerling, W.; Rüggeberg, A.; Wisshak, M.; Piepenburg, D.; Meyerhöfer, M.; Form, A.; Freiwald, A. Arctic rhodolith beds and their environmental controls (Spitsbergen, Norway). Facies 2014, 60, 15–37. [Google Scholar] [CrossRef]
- Teichert, S.; Woelkerling, W.; Rüggeberg, A.; Wisshak, M.; Piepenburg, D.; Meyerhöfer, M.; Form, A.; Büdenbender, J.; Freiwald, A. Rhodolith beds (Corallinales, Rhodophyta) and their physical and biological environment at 80 ° 31′ N in Nordkappbukta (Nordaustlandet, Svalbard Archipelago, Norway). Phycologia 2012, 51, 371–390. [Google Scholar] [CrossRef]
- Basso, D.; Nalin, R.; Nelson, C.S. Shallow-water Sporolithon rhodoliths from North Island (New Zealand). Palaios 2009, 24, 92–103. [Google Scholar] [CrossRef]
- Bahia, R.G.; Abrantes, D.P.; Brasileiro, P.S.; Pereira Filho, G.H.; Amado Filho, G.M. Rhodolith bed structure along a depth gradient on the northern coast of Bahia state, Brazil. Braz. J. Oceanogr. 2010, 58, 323–337. [Google Scholar] [CrossRef]
- Amado-Filho, G.M.; Maneveldt, G.; Manso, R.C.C.; Marins-Rosa, B.V.; Pacheco, M.R.; Guimarães, S. Structure of rhodolith beds from 4 to 55 meters deep along the southern coast of Espírito Santo State, Brazil. Cienc. Mar. 2007, 33, 399–410. [Google Scholar] [CrossRef] [Green Version]
- Marrack, E.C. The relationship between water motion and living rhodolith beds in the southwestern Gulf of California, Mexico. Palaios 1999, 14, 159–171. [Google Scholar] [CrossRef]
- James, D.W. Diet, movement, and covering behavior of the sea urchin Toxopneustes roseus in rhodolith beds in the Gulf of California, México. Mar. Biol. 2000, 137, 913–923. [Google Scholar] [CrossRef]
- Pereira-Filho, G.H.; Veras, P.C.; Francini-Filho, R.B.; Moura, R.L.; Pinheiro, H.T.; Gibran, F.Z.; Matheus, Z.; Neves, L.M.; Amado-Filho, G.M. Effects of the sand tilefish Malacanthus plumieri on the structure and dynamics of a rhodolith bed in the Fernando de Noronha Archipelago, tropical West Atlantic. Mar. Ecol. Prog. Ser. 2015, 541, 65–73. [Google Scholar] [CrossRef]
- Kamenos, N.A.; Law, A. Temperature controls on coralline algal skeletal growth. J. Phycol. 2010, 46, 331–335. [Google Scholar] [CrossRef]
- Halfar, J.; Zack, T.; Kronz, A.; Zachos, J.C. Growth and high-resolution paleoenvironmental signals of rhodoliths (coralline red algae): A new biogenic archive. J. Geophys. Res. Oceans 2000, 105, 22107–22116. [Google Scholar] [CrossRef]
- Steller, D.L.; Riosmena-Rodríguez, R.; Foster, M.S.; Roberts, C.A. Rhodolith bed diversity in the Gulf of California: The importance of rhodolith structure and consequences of disturbance. Aquat. Conserv. 2003, 13, S5–S20. [Google Scholar] [CrossRef]
- Veras, P.C.; Pierozzi, I., Jr.; Lino, J.B.; Amado-Filho, G.M.; Senna, A.R.; Santos, C.S.G.; de Moura, R.L.; Passos, F.D.; Giglio, V.J.; Pereira-Filho, G.H. Drivers of biodiversity associated with rhodolith beds from euphotic and mesophotic zones: Insights for management and conservation. Perspec. Ecol. Conserv. 2020, 18, 37–43. [Google Scholar] [CrossRef]
- Foster, M.S.; McConnico, L.M.; Lundsten, L.; Wadsworth, T.; Kimball, T.; Brooks, L.B.; Medina-López, M.; Riosmena-Rodríguez, R.; Hernández-Carmona, G.; Vásquez-Elizondo, R.M. Diversity and natural history of a Lithothamnion muelleri-Sargassum horridum community in the Gulf of California. Cienc. Mar. 2007, 33, 367–384. [Google Scholar] [CrossRef] [Green Version]
- Himmelman, J.H.; Dumont, C.P.; Gaymer, C.F.; Vallières, C.; Drolet, D. Spawning synchrony and aggregative behaviour of cold-water echinoderms during multi-species mass spawnings. Mar. Ecol. Prog. Ser. 2008, 361, 161–168. [Google Scholar] [CrossRef]
- Doyle, G.M.; Hamel, J.F.; Mercier, A. Small-scale spatial distribution and oogenetic synchrony in brittlestars (Echinodermata: Ophiuroidea). Estuar. Coast. Shelf Sci. 2014, 136, 172–178. [Google Scholar] [CrossRef]
- Guo, L.; Perrie, W.; Long, Z.; Toulany, B.; Sheng, J. The impacts of climate change on the autumn North Atlantic wave climate. Atmos. Ocean 2015, 53, 491–509. [Google Scholar] [CrossRef]
- Working Group II. Climate Change 2022: Impacts, Adaptation and Vulnerability; IPCC Sixth Assessment Report; IPCC: Geneva, Switzerland, 2022. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Survey | Site | Sampling Date (2013) | Temperature °C (±SD) |
|---|---|---|---|
| Winter | SP15 | 14 Mar | −0.1 (0.2) |
| SP18 | 4 Mar | ||
| Spring | SP15 | 17 Jun | 5.5 (1.9) |
| SP18 | 10 Jun | ||
| Summer | SP15 | 17 Sep | 11.7 (1.5) |
| SP18 | 1 Oct | ||
| Fall | SP15 and SP18 | 15 Dec | 4.4 (0.6) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bélanger, D.; Gagnon, P. Spatiotemporal Variability in Subarctic Lithothamnion glaciale Rhodolith Bed Structural Complexity and Macrofaunal Diversity. Diversity 2023, 15, 774. https://doi.org/10.3390/d15060774
Bélanger D, Gagnon P. Spatiotemporal Variability in Subarctic Lithothamnion glaciale Rhodolith Bed Structural Complexity and Macrofaunal Diversity. Diversity. 2023; 15(6):774. https://doi.org/10.3390/d15060774
Chicago/Turabian StyleBélanger, David, and Patrick Gagnon. 2023. "Spatiotemporal Variability in Subarctic Lithothamnion glaciale Rhodolith Bed Structural Complexity and Macrofaunal Diversity" Diversity 15, no. 6: 774. https://doi.org/10.3390/d15060774
APA StyleBélanger, D., & Gagnon, P. (2023). Spatiotemporal Variability in Subarctic Lithothamnion glaciale Rhodolith Bed Structural Complexity and Macrofaunal Diversity. Diversity, 15(6), 774. https://doi.org/10.3390/d15060774