


A New Species of Vampirolepis (Cestoda: Cyclophyllidea: Hymenolepididae) from the Bat Artibeus lituratus (Chiroptera: Phyllostomidae) in the Amazon Rainforest, Brazil
 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Study Area
2.2. Parasite Collection
2.3. Integrative Taxonomy
2.3.1. Morphological Studies
2.3.2. Histological Analyses
2.3.3. Molecular and Phylogenetic Analyses
3. Results
3.1. Bat Capture
3.2. Vampirolepis spp.
3.3. Vampirolepis dalvae n. sp.
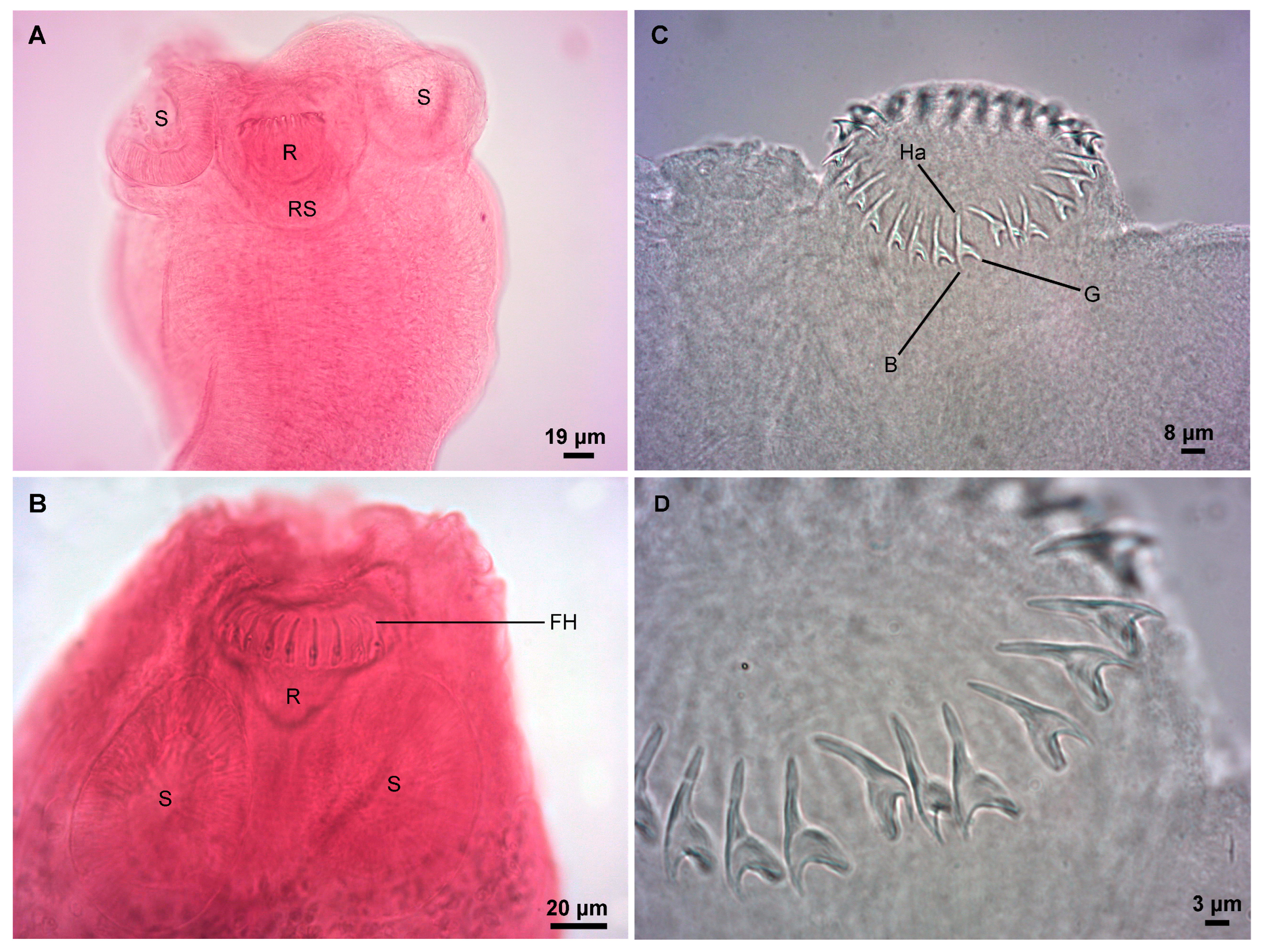
Description
3.4. Molecular and Phylogenetic Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Embrapa. Contando Ciência na Web—Bioma Amazonia. 2023. Available online: https://www.embrapa.br/contando-ciencia/bioma-amazonia (accessed on 12 January 2023).
- Fearnside, P.M. The intrinsic value of Amazon biodiversity. Biodivers. Conserv. 2021, 30, 1199–1202. [Google Scholar] [CrossRef]
- Verde, R.S.; Oliveira, S.F.; Menezes, A.O.; Gonçalves, F.; Alencar, L.; Silva, T.M.; Calouro, A.M.; Mews, H.A.; Morato, E.F. Bats (Mammalia, Chiroptera) from a bamboo-dominated forest in the southwestern Brazilian Amazon, with the first records of Glyphonycteris sylvestris Thomas, 1896 and Phylloderma stenops Peters, 1865 from Acre state. Check List 2021, 17, 311–321. [Google Scholar] [CrossRef]
- Santos, C.P.; Gibson, D.I. Checklist of the Helminth Parasites of South American Bats. Zootaxa 2015, 3, 471–499. [Google Scholar] [CrossRef] [Green Version]
- Albuquerque, A.C.A.; Moraes, M.F.D.; Silva, A.C.; Lapera, I.M.; Tebaldi, J.H.; Hoppe, E.G.L. Helminth fauna of chiropterans in Amazonia: Biological interactions between parasite and host. Parasitol. Res. 2016, 115, 3229–3237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zortéa, M. Subfamília Stenodermatinae. In Morcegos do Brasil, 1st ed.; Reis, N.R., Peracchi, A.L., Pedro, W.A., Lima, I.P., Eds.; Biblioteca Central da Universidade Estadual de Londrina: Londrina, Brasil, 2007; Volume único, pp. 107–128. [Google Scholar]
- Vaucher, C. Cestodes parasites de chiroptères en Amerique du Sud: Revision de Hymenolepis elongatus (Rêgo, 1962) et description de Hymenolepis phyllostomi n. sp. Rev. Suisse Zool. 1982, 89, 451–459. [Google Scholar] [CrossRef]
- Vaucher, C. Helminthes parasites du Paraguay. X: Hymenolepis dasipteri n. sp. (Cestoda, Hymenolepididae) chez Dasipterus ega argentinus Thomas (Chiroptera, Vespertilionidae). Bull. De La Société Des Sci. Nat. De Neuchâtel 1985, 10, 155–161. [Google Scholar]
- Vaucher, C. Helminthes parasites du Paraguay. XI: Hymenolepididae (Cestoda) parasites de chiroptères Molossidae, avec description de deux espèces nouvelles. Rev. Suisse Zool. 1986, 93, 393–407. [Google Scholar] [CrossRef]
- Vaucher, C. Cestodes parasites de Chiroptères em Amerique du Sud. II: Hymenolepis mazanensis n. sp., chez Saccopteryx bilineata (Temm.) et Rhynchonycteris naso (Wied-Neuwied) (Chiroptera: Emballonuridae) em Amazonie péruvienne. Rev. Suisse Zool. 1986, 93, 817–821. [Google Scholar] [CrossRef]
- Vaucher, C. Revision of the genus Vampirolepis Spasskij, 1954 (Cestoda: Hymenolepididae). Mem. Inst. Oswaldo Cruz 1992, 87, 299–304. [Google Scholar] [CrossRef]
- Zdzitowiecki, K.; Rutkowska, M. The helminthofauna of bats (Chiroptera) from Cuba. II. A review of cestodes with description of four new species and a key to Hymenolepididae of American bats. Acta Parasitol. Pol. 1980, 26, 187–200. [Google Scholar]
- Mendoza, L.; Chavez, J.; Tantaleán, M. Cestodos parasitos de murciélagos de Ica, Peru. Parasitol. día 1997, 21, 20–24. [Google Scholar]
- Sawada, I.; Harada, M. Bat cestodes from Bolivia, South America, with descriptions of six new species. Zool. Sci. 1986, 3, 367–377. [Google Scholar]
- Macy, R.W. A key to the species of Hymenolepis found in bats and the description of a new species, H. christensoni, from Myotis lucifugus. Trans. Am. Microsc. Soc. 1931, 50, 344–347. [Google Scholar] [CrossRef]
- Arandas Rêgo, A. Sobre alguns “Vampirolepis” parasitos de quirópteros (Cestoda, Hymenolepididae). Rev. Brasil. Biol. 1962, 22, 129–136. [Google Scholar]
- Sawada, I. Vampirolepis ezoensis sp. nov. (Cestoda: Hymenolepididae) from the Japanese northern bat, Eptesicus nilssoni parvus Kishida, with a list of known species of the genus Vampirolepis Spassky from bats. Jpn. J. Parasitol. 1990, 2, 176–185. [Google Scholar]
- Diesing, C.L. Systema Helminthum; Sumptibus Academiae Caesareae Scientiarum: Vindobonae, Austria, 1850; Volume 1, pp. 1–679. [Google Scholar]
- Nogueira, M.R.; de Fabio, S.P.; Peracchi, A.L. Gastrointestinal helminth parasitism in fruit-eating bats (Chiroptera, Stenodermatinae) from western Amazonian Brazil. Rev. Biol. Trop. 2004, 52, 387–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arandas Rêgo, A. Nota prévia sobre um novo Vampirolepis parasito de quirópteros (Cestoda, Hymenolepididae). Atas Soc. Biol. 1961, 5, 32–34. [Google Scholar]
- Sawada, I. Check-list of new cestode species recorded by Sawada. Nara Sangyo Univ. J. Ind. Econ. 1997, 11, 111–127. [Google Scholar]
- Haukisalmi, V.; Hardman, L.M.; Foronda, P.; Feliu, C.; Laakkonen, J.; Niemimaa, J.; Lehtonen, J.T.; Henttonen, H. Systematic relationships of hymenolepidid cestodes of rodents and shrews inferred from sequences of 28S ribosomal RNA. Zool. Scr. 2010, 39, 631–641. [Google Scholar] [CrossRef]
- Makarikova, T.A.; Gulyaev, V.D.; Tiunov, M.P.; Feng, J. A new species of cestode, Vampirolepis muraiae n. sp. (Cyclophyllidea: Hymenolepididae), from a Chinese bat. Syst. Parasitol. 2012, 82, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Semapi, M. Parque Estadual Chandless. 2020. Available online: http://semapi.acre.gov.br/parque-estadual-chandless (accessed on 14 January 2020).
- Plano de Manejo do Parque Estadual Chandless. I, II, III e IV. In Secretaria de Estado de Meio Ambiente (SEMA) 2010; Governo do Estado do Acre: Rio Branco, Brazil, 2010; pp. 1–77.
- Gardiner, C.H.; Poynton, S.L. Morphological characteristics of Cestodes in tissue section. In An Atlas of Metazoan Parasites in Animal Tissues, 1st ed.; Armed Forces Institute of Pathology: Washington, DC, USA, 1999; pp. 50–55. [Google Scholar]
- Díaz, M.M.; Solari, S.; Aguirre, L.F.; Aguiar, L.M.S.; Barquez, R.M. Chave de Identificação dos Morcegos da Améria do Sul, 2nd ed.; PCMA: Tucumán, Argentina, 2016; pp. 1–160. [Google Scholar]
- López-Baucells, A.; Rocha, R.; Bobrowiec, P.; Bernard, E.; Palmeirim, J.; Meyer, C. Field Guide to Amazonian Bats, 1st ed.; Editora INPA: Manaus, Brazil, 2016; pp. 1–168.
- Amato, J.F.R.; Amato, S.B. Técnicas gerais para coleta e preparação de helmintos endoparasitos de aves. In Ornitologia e Conservação: Ciência Aplicada, Técnicas de Pesquisa e Levantamento; Universidade Federal do Rio Grande do Sul: Rio Grande do Sul, Brazil, 2010; pp. 1–25. [Google Scholar]
- Khalil, L.F.; Jones, A.; Bray, R.A. Keys to the Cestoda Parasites of Vertebrates; CAB International: Wallingford, UK, 2006; pp. 1–751. [Google Scholar]
- Survana, S.K.; Layton, C.; Bancroft, J.D. Bancroft´s Theory and Practice of Histological Techniques, 8th ed.; Elsevier Limited: Beijing, China, 2019; pp. 1–537. [Google Scholar]
- Dorris, M.; Blaxter, M. The small subunit ribosomal RNA sequence of Strongyloides stercoralis. Int. J. Parasitol. 2000, 30, 939–941. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximumlikelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefort, V.; Longueville, J.E.; Gascuel, O. SMS: Smart model selection in PhyML. Mol. Biol. Evol. 2017, 34, 2422–2424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anisimova, M.; Gascuel, O. Approximate likelihood-ratio test for branches: A fast, accurate, and powerful alternative. Syst. Biol. 2006, 55, 539–552. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van derMark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes version 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Czaplinski, B.; Vaucher, C. Family Hymenolepididae Ariola, 1899. In Keys to the Cestode Parasites of Vertebrates; Khalil, L.F., Jones, A., Bray, R.A., Eds.; CAB International: Wallingford, UK, 1994; pp. 595–663. [Google Scholar]
- Vaucher, C. Les cestodes parasites des Soricidae dÉurope. Étude anatomique revision taxonomique et biologie. Rev. Suisse Zool. 1971, 78, 1–113. [Google Scholar] [CrossRef] [Green Version]
- Georgiev, B.B.; Bray, R.A.; Littlewood, D.T.J. Small mammals as definitive hosts of cestodes. In Micromammals and Macroparasites. from Evolutionary Ecology to Management; Morand, S., Krasnov, B.R., Poulin, R., Eds.; Springer: Tokyo, Japan, 2006; pp. 23–62. [Google Scholar]
- Mariaux, J.; Tkach, V.V.; Vasileva, G.P.; Waeschenbach, A.; Beveridge, I.; Dimitrova, Y.D.; Haukisalmi, V.; Greiman, S.E.; Littlewood, D.T.J.; Makarikov, A.A.; et al. Cyclophyllidea van Beneden in Braun, 1900. In Planetary Biodiversity Inventory (2008–2017): Tapeworms from Vertebrate Bowels of the Earth; Caira, J.N., Jensen, K., Eds.; University of Kansas: Lawrence, KS, USA; Natural History Museum: Lawrence, KS, USA, 2017; pp. 77–148. [Google Scholar]
- Spassky, A.A. Classification of the hymenolepidids of mammals. Tr. Gel’mintol. Lab. 1954, 7, 121–167. [Google Scholar]
- Makarikova, T.A.; Makarikov, A.A. Sawadalepis prima n. g., sp. (Cestoda: Cyclophyllidea) from the Schreiber´s bent-winged bat Miniopterus schreibersii Kuhl (Chiroptera: Vespertilionidae) from China. Syst. Parasitol. 2013, 86, 59–68. [Google Scholar] [CrossRef]
- Sapssky, A.A. On the systematic position of the hymenolepidids (Cestoda) from Australian Marsupialia. Parazitologiya 1994, 28, 66–69. [Google Scholar]
- Vaucher, C.; Beveridge, I. New species of Potorolepis Spassky (Cestoda: Hymenolepididae) parasitic in Dasyurid marsupials from New Guinea. Trans. R. Soc. South Aust. 1997, 121, 95–102. [Google Scholar]
- Greiman, S.E.; Tkach, V.V. Description and phylogenetic relationships of Rodentolepis gnoskei n. sp. (Cyclophyllidea: Hymenolepididae) from a shrew Suncus varilla minor in Malawi. Parasitol. Int. 2012, 61, 343–350. [Google Scholar] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Number of Rostellar Hooks | Host(s) | Locality | Reference(s) |
|---|---|---|---|---|
| Vampirolepis artibei (Rutkowska & Zdzitowiecki, 1980) | 20–23 (n = 14) | Glossophaga soricina | Peru | [4,12,13,17] |
| Vampirolepis bihamata (Sawada & Harada, 1986) | 88–90 (n = 2) | Micronycteris minuta | Bolivia | [4,14,17,21] |
| Vampirolepis christensoni (Macy, 1931) | 36–38 (n = 14) | Nyctinomops laticaudatus, Molossidae gen. sp. and Chiroptera gen. sp. | Brazil, Bolivia and Paraguay | [4,12,15,16,17] |
| Vampirolepis crassihamata (Sawada & Harada, 1986) | 22 (n = 1) | Molossus molossus | Bolivia | [4,14,17,21] |
| Vampirolepis decipiens (Diesing, 1850) | 44 (n = 9) | Cynomops abrasus brachymeles, Eumops patagonicus (as Eumops bonariensis beckeri), Eumops glaucinus, E. perotis, Molossops temminckii and Pteronotus parnellii rubiginosus (as Chilonycteris rubiginosa) | Brazil, Argentina and Paraguay | [4,9,12,16,17,18] |
| Vampirolepis elongatus (Rêgo, 1962) | 32 (n = 20) | Molossus rufus, Glossophaga soricina, Artibeus obscurus (as A. fuliginosus), A. planirostris, Artibeus lituratus, Pygoderma bilabiatum, Phyllostomus hastatus, Platyrrhinus helleri and Chiroptera gen. sp. | Brazil, Argentina, Paraguay, Bolivia and Peru | [4,7,12,16,17,19] |
| Vampirolepis guarany (Rêgo, 1961) | 24–26 (n = 13) | Molossus crassicaudatus, Molossus sp. and Chiroptera sp. | Paraguay and Brazil | [9,16,17,20] |
| Vampirolepis longisccata (Sawada & Harada, 1986) | 36–38 (n = 2) | Molossus molossus | Bolivia | [14,17,21] |
| Vampirolepis temmincki (Vaucher, 1986) | 29–34 (n = 13) | Molossus temminckii | Paraguay | [4,9,17] |
| Vampirolepis promopsis (Vaucher, 1986) | 45 (n = 1) | Promops centralis | Paraguay | [4,9,17] |
| Vampirolepis phyllostomi (Vaucher, 1982) | 42–52 (n = 8) | Phyllostomus hastatus and Eumops bonariensis beckeri | Peru, Bolivia and Paraguay | [4,9,14,17] |
| Vampirolepis santacruzensis (Sawada & Harada, 1986) | 23 (n = 1) | Molossus molossus | Bolivia | [4,14,17,21] |
| Vampirolepis mazanensis (Vaucher, 1986) | 37–40 (n = 4) | Saccopteryx bilineata and Rhynchonycteris naso | Peru | [10,17] |
| Vampirolepis pandoensis (Sawada & Harada, 1986) | 41 (n = 1) | Eptesicus furinalis | Bolivia | [4,14,17,21] |
| Structural Measurements (mm) | Vampirolepis dalvae n. sp. | Vampirolepis artibei (Rutkowska & Zdzitowiecki, 1980) | Vampirolepis crassihamata (Harada & Sawada, 1986) | Vampirolepis guarany (Rêgo, 1962) | Vampirolepis santacruzensis (Harada & Sawada, 1986) |
|---|---|---|---|---|---|
| Length | 40.81 (n = 1) | 40 (n = 1) | 32 × 0.5 (#) (n = 1) | 30.15−36.83 (n = 13) | 65 × 1.2 (#) (n = 1) |
| Neck (L × W) | - | - | - | - | 0.9 × 0.4 (n = 1) |
| Scolex (L × W) | 0.363 × 0.342 (0.236–0.47 × 0.281–0.4) (n = 10) | 0.211 × 0.276 (n = 14) | 0.560 × 0.476 (n = 1) | 0.415 × 0.581 (n = 13) | 0.420 × 0.476 (n = 1) |
| Rostellar sac (L × W) | 0.134 × 0.099 (0.097–0.155 × 0.069–0.123) (n = 5) | (0.096–0.125) (W) (n = 14) | 0.245 (*) (n = 1) | 0.363 × 0.231 (n = 13) | 0.315 × 0.345 (n = 1) |
| Rostellum (L × W) | 0.069 × 0.073 (0.057–0.089 × 0.052–0.111) (n = 10) | (0.074–0.089 × 0.077–0.083) (n = 14) | 0.189 × 0.161 (n = 1) | 0.209 × 0.182 (n = 13) | 0.231 × 0.175 (n = 1) |
| Suckers (L × W) | 0.108 × 0.106 (0.078–0.144 × 0.086–0.128) (n = 10; ns = 36) | (0.089–0.104) (*) (n = 14) | (0.133–0.140 × 0.154) (n = 1) | 0.126 (*) (n = 13) | (0.147–0.161 × 0.126–0.161) (n = 1) |
| Mature proglottids (L × W) | 0.086 × 0.340 (0.061–0.105 × 0.247–0.413) (n = 1; ns = 12) | (0.089–0.126 × 0.4–0.43) (n = 1) | - | 0.174 × 2.53 (n = 13) | - |
| Gravid proglottids (L × W) | 0.178 × 0.476 (0.135–0.267 × 0.333–0.547) (n = 2; ns = 8) | (0.148–0.193 × 0.34–0.445) (n = 1) | - | - | - |
| Testes (L × W) | 0.02 × 0.021 (0.018–0.023 × 0.018–0.026) (n = 1; ns = 11) | (0.042–0.053 × 0.055–0.072) (n = 1) | (0.084–0.091 × 0.067–0.084) (n = 1) | 0.191 × 0.104 (n = 13) | (0.105–0.119 × 0.070–0.084) (n = 1) |
| Testes arrangement | Transversal row | Transversal row | Transversal row | Transversal row | Transversal row |
| Cirrus sac (L × W) | 0.084 × 0.028 (0.077–0.095 × 0.027–0.030) (n = 1; ns = 3) | (0.096–0.103 × 0.029–0.048) (n = 1) | (0.140–0.175 × 0.028) (n = 1) | 0.182 × 0.043 (n = 13) | (0.154–0.175 × 0.035) (n = 1) |
| Cirrus | 0.032 (*) (n = 1; ns = 1) | 0.024 (L)/0.06 (*) (n = 1) | - | - | - |
| External seminal vesicle (L × W) | 0.027 × 0.025 (0.018–0.034 × 0.018–0.031) (n = 1; ns = 4) | (0.054–0.072 × 0.030–0.038) (n = 1) | (0.091–0.098 × 0.028) (n = 1) | (0.070–0.084 × 0.028–0.053) (n = 1) | |
| Internal seminal vesicle (L × W) | 0.027 × 0.030 (0.018–0.039 × 0.026–0.034) (n = 1; ns = 4) | - | 0.070–0.105 × 0.028 (n = 1) | - | (0.070–0.084 × 0.028–0.053) (n = 1) |
| Seminal receptacle (L × W) | 0.019 × 0.026 (0.015–0.028 × 0.018–0.042) (n = 1; ns = 4) | (0.082–0.144 × 0.048–0.108) (n = 1) | 0.063–0.077 × 0.035 (n = 1) | 0.174 × 0.052 (n = 13) | (0.105–0.154 × 0.084) (n = 1) |
| Ovary (L × W) | 0.039 × 0.087 (0.036–0.039 × 0.076–0.1) (n = 1; ns = 4) | (0.054–0.067 × 0.156–0.175) (n = 1) | - | 0.783 (n = 13) | - |
| Vitelline gland (L × W) | - | (0.040–0.050 × 0.060–0.075) (n = 1) | (0.049 × 0.021–0.028) (n = 1) | 0.348 (n = 13) | (0.140 × 0.077–0.084) (n = 1) |
| Eggs (L × W) | 0.055 × 0.046 0.045–0.066 × 0.042–0.053 (n = 1; ns = 10) | (0.035–0.045) (*) (n = 1) | - | 0.034 (n = 13) | (0.046–0.056 × 0.035–0.046) (n = 1) |
| Onchosphere | 0.037 × 0.030 (0.030–0.050 × 0.025–0.038) (n = 1; ns = 11) | (0.025–0.029 × 0.022–0.027) (n = 1) | - | - | (0.028–0.039 × 0.025–0.028) (n = 1) |
| Onchosphere hooks (L) | - | 0.016 (n = 1) | - | 0.015 (n = 13) | 0.014 (n = 1) |
| Hooks (N) | (21–25) (n = 10) | (20–23) (n = 14) | 22 (n = 1) | (24–26) (n = 13) | 23 (n = 1) |
| Hook blades (L × W) | - | - | - | (0.014–0.017) (n = 13) | - |
| Hooks (L) | 0.020 (0.016–0.030) (n = 10; ns = 43) | (0.019–0.020) (n = 14) | 0.053 (n = 1) | 0.05 (n = 13) | 0.046 (n = 1) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
dos Santos, M.M.; Simões, R.d.O.; D’Andrea, P.S.; Verde, R.d.S.; Maldonado Júnior, A.; Argueta Cartagena, R.I.; Ubiali, D.G.; Luque, J.L. A New Species of Vampirolepis (Cestoda: Cyclophyllidea: Hymenolepididae) from the Bat Artibeus lituratus (Chiroptera: Phyllostomidae) in the Amazon Rainforest, Brazil. Diversity 2023, 15, 791. https://doi.org/10.3390/d15060791
dos Santos MM, Simões RdO, D’Andrea PS, Verde RdS, Maldonado Júnior A, Argueta Cartagena RI, Ubiali DG, Luque JL. A New Species of Vampirolepis (Cestoda: Cyclophyllidea: Hymenolepididae) from the Bat Artibeus lituratus (Chiroptera: Phyllostomidae) in the Amazon Rainforest, Brazil. Diversity. 2023; 15(6):791. https://doi.org/10.3390/d15060791
Chicago/Turabian Styledos Santos, Michele Maria, Raquel de Oliveira Simões, Paulo Sérgio D’Andrea, Rair de Sousa Verde, Arnaldo Maldonado Júnior, Reina Isabel Argueta Cartagena, Daniel Guimarães Ubiali, and José Luis Luque. 2023. "A New Species of Vampirolepis (Cestoda: Cyclophyllidea: Hymenolepididae) from the Bat Artibeus lituratus (Chiroptera: Phyllostomidae) in the Amazon Rainforest, Brazil" Diversity 15, no. 6: 791. https://doi.org/10.3390/d15060791
APA Styledos Santos, M. M., Simões, R. d. O., D’Andrea, P. S., Verde, R. d. S., Maldonado Júnior, A., Argueta Cartagena, R. I., Ubiali, D. G., & Luque, J. L. (2023). A New Species of Vampirolepis (Cestoda: Cyclophyllidea: Hymenolepididae) from the Bat Artibeus lituratus (Chiroptera: Phyllostomidae) in the Amazon Rainforest, Brazil. Diversity, 15(6), 791. https://doi.org/10.3390/d15060791