

Influence of Distance from Forest Edges on Spontaneous Vegetation Succession Following Small-Scale Gold Mining in the Southeast Peruvian Amazon

 ,
,  , , ,
, , ,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
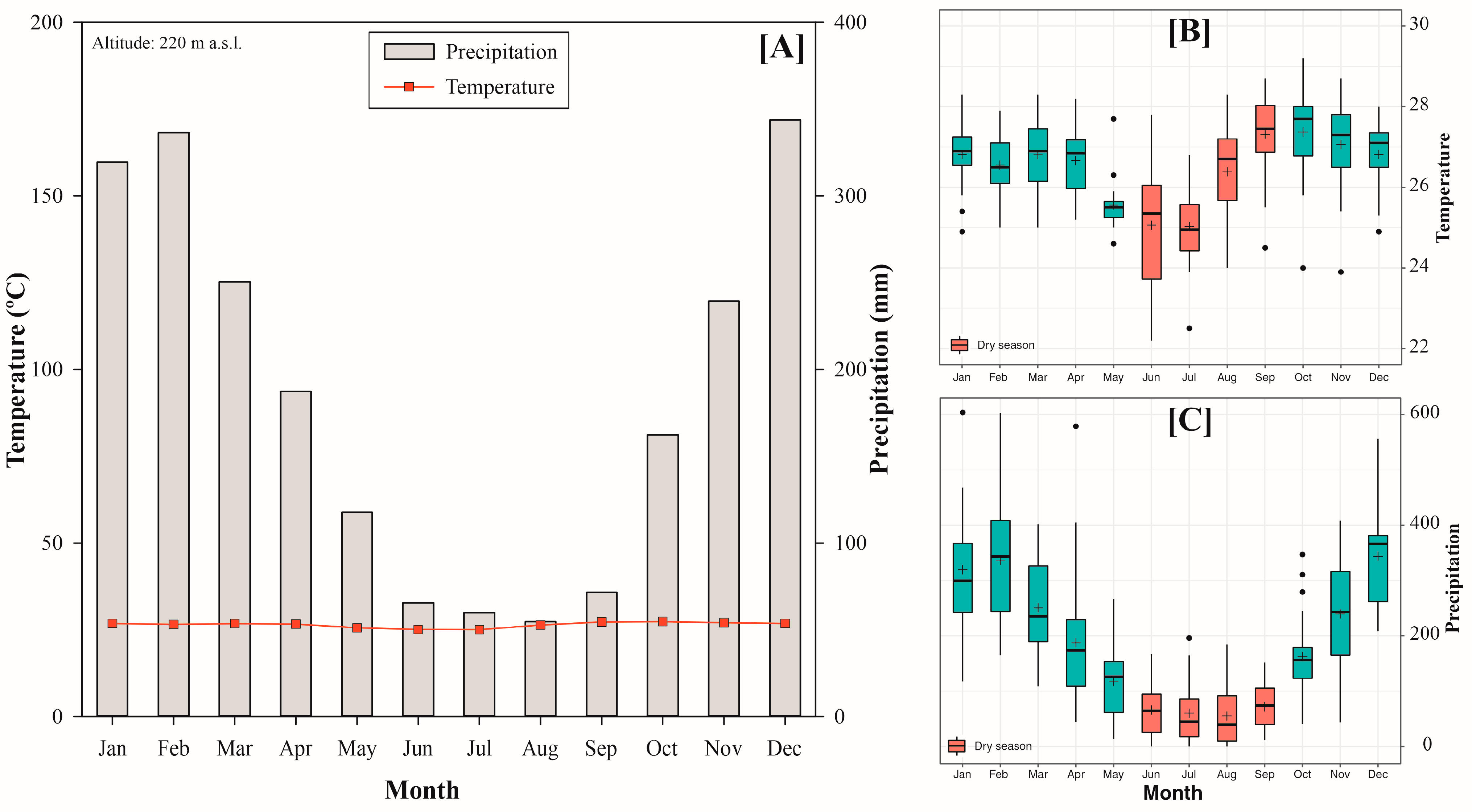
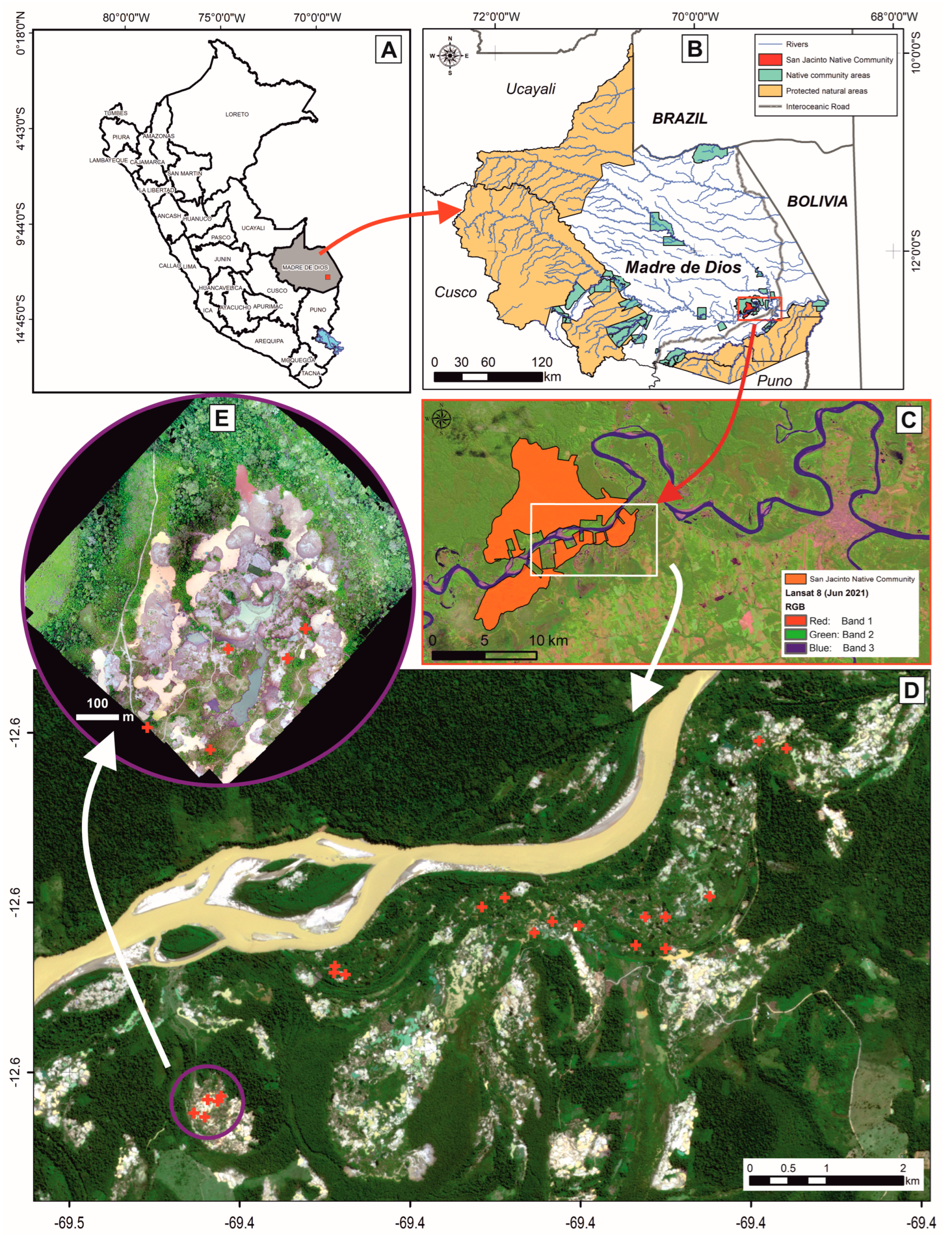
2.1. Study Area

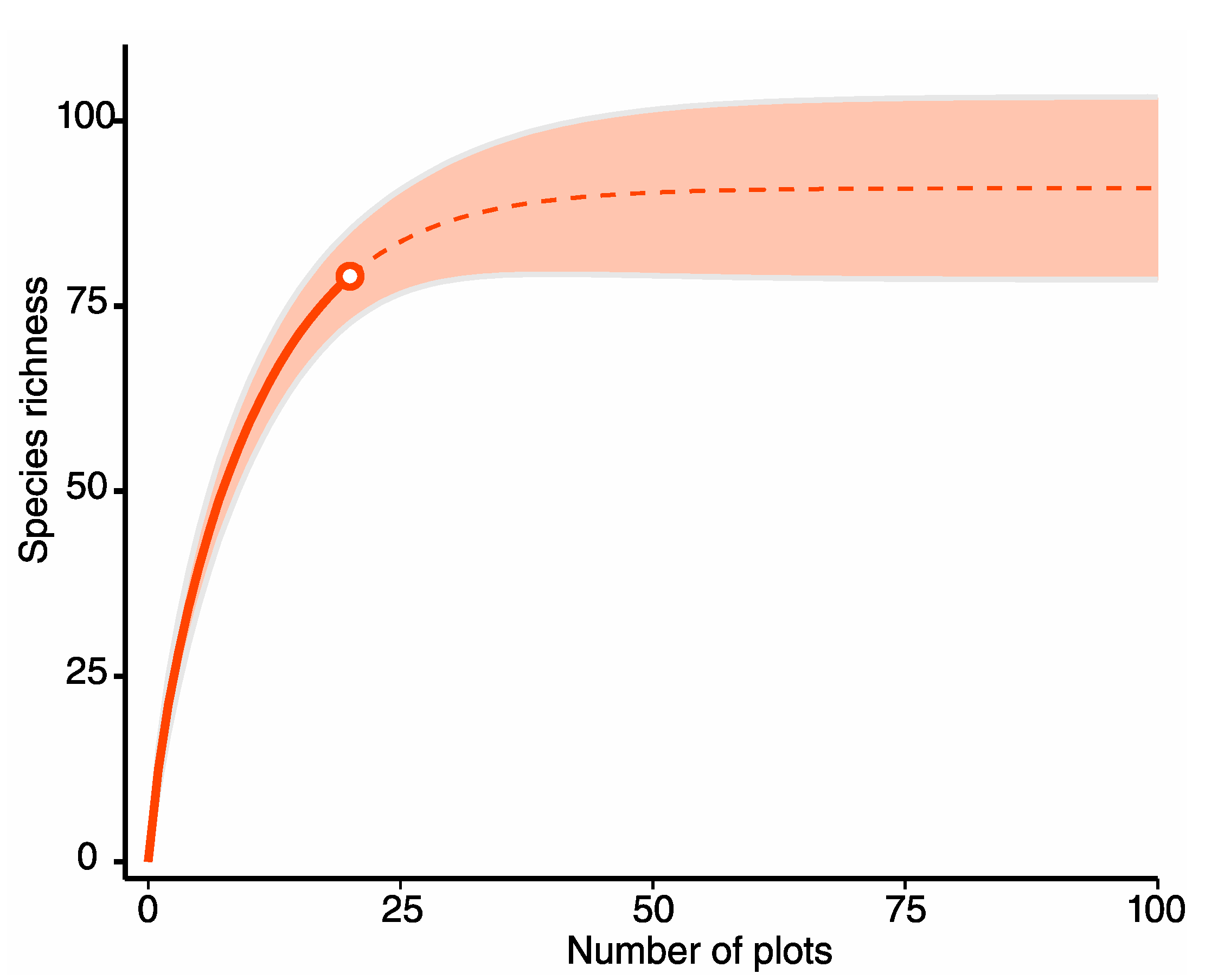
2.2. Sampling and Vegetation Structure
2.3. Species Importance and Diversity Index
2.4. Response of the Species to the Time of Abandonment and Distance to the Forest Edge
2.5. Data Analysis
3. Results
3.1. Structure, Diversity, and Floristic Composition
3.2. Species according to Abandonment Time Categories
3.3. Species Occurrence according to the Gradient of Distance to the Forest Edge and Abandonment Time
3.4. Time and Distance Relationship
3.5. Ordination Analysis of Floristic Composition
3.6. Abandonment Time and Distance to the Forest Edge in the Structuring of Plant Communities
4. Discussion
4.1. Species according to Abandonment Time Category
4.2. Abandonment Time and Distance to the Forest Edge
4.3. Abandonment Time and Distance to the Forest Edge in the Structuring of Forest Communities
4.4. Natural Regeneration Outlook for Forest Recovery
4.5. Limitations and Future Research Directions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cherlet, M.; Hutchinson, C.; Reynolds, J.; Hill, J.; Sommer, S.; von Maltitz, G. World Atlas of Desertification, Mapping Land Degradation and Sustainable Land Management Opportunities, 3rd ed.; Publication Office of the European Union: Luxembourg, 2018. [Google Scholar]
- Timsina, S.; Hardy, N.G.; Woodbury, D.J.; Ashton, M.S.; Cook-Patton, S.C.; Pasternack, R.; Martin, M.P. Tropical surface gold mining: A review of ecological impacts and restoration strategies. Land Degrad. Dev. 2022, 33, 3661–3674. [Google Scholar] [CrossRef]
- Reitschmiedová, E.; Dvorščík, P.; Mudrák, O.; Šimáňová, D.; Frouz, J. Differences in colonization strategies of three common pioneer woody species in post mining heaps. J. Environ. Manag. 2022, 319, 115668. [Google Scholar] [CrossRef]
- Alarcón-Aguirre, G.; Sajami Quispe, E.; Vásquez Zavaleta, T.; Ponce Tejada, L.V.; Ramos Enciso, D.; Rodríguez Achata, L.; Garate-Quispe, J. Vegetation dynamics in lands degraded by gold mining in the southeastern Peruvian Amazon. Trees For. People 2023, 11, 100369. [Google Scholar] [CrossRef]
- Prach, K.; Walker, L.R. Differences between primary and secondary plant succession among biomes of the world. J. Ecol. 2019, 107, 510–516. [Google Scholar] [CrossRef]
- Rocha-Nicoleite, E.; Campos, M.L.; Colombo, G.T.; Overbeck, G.E.; Müller, S.C. Forest restoration after severe degradation by coal mining: Lessons from the first years of monitoring. Braz. J. Bot. 2018, 41, 653–664. [Google Scholar] [CrossRef]
- Leiva, J.D. Appropriate technologies and the geosocial evolution of informal, small-scale gold mining in Madre de Dios, Peru. Extr. Ind. Soc. 2022, 12, 101165. [Google Scholar] [CrossRef]
- Stoll, E.; Roopsind, A.; Maharaj, G.; Velazco, S.; Caughlin, T.T. Detecting gold mining impacts on insect biodiversity in a tropical mining frontier with SmallSat imagery. Remote Sens. Ecol. Conserv. 2022, 8, 379–390. [Google Scholar] [CrossRef]
- Mesquita, R.; Massoca, P.; Jakovac, C.C.; Bentos, T.; Williamson, G. Amazon Rain Forest Succession: Stochasticity or Land-Use Legacy? Bioscience 2015, 65, 849–861. [Google Scholar] [CrossRef] [Green Version]
- Tito, R.; Salinas, N.; Cosio, E.G.; Boza Espinoza, T.E.; Muñiz, J.G.; Aragón, S.; Nina, A.; Roman-Cuesta, R.M. Secondary forests in Peru: Differential provision of ecosystem services compared to other post-deforestation forest transitions. Ecol. Soc. 2022, 27, 12. [Google Scholar] [CrossRef]
- Zapata-Carbonell, J.; Bégeot, C.; Carry, N.; Choulet, F.; Delhautal, P.; Gillet, F.; Girardclos, O.; Mouly, A.; Chalot, M. Spontaneous ecological recovery of vegetation in a red gypsum landfill: Betula pendula dominates after 10 years of inactivity. Ecol. Eng. 2019, 132, 31–40. [Google Scholar] [CrossRef]
- Lenart, J.; Schuchová, K.; Kašing, M.; Falteisek, L.; Cimalová, Š.; Bílá, J.; Ličbinská, M.; Kupka, J. The abandoned underground mine as a semi-natural ecosystem: The story of Flaschar’s Mine (Czechia). CATENA 2022, 213, 106178. [Google Scholar] [CrossRef]
- Velásquez-Ramírez, M.; del Castillo Torres, D.; Guerrero Barrantes, J.; Vásquez Bardales, J.; Thomas, E.; Cusi Auca, E.; Chinen Gushiken, M.; Muñoz Diaz, B.; Russo, R.; Corvera Gomringer, R. Soil recovery of alluvial gold mine spoils in the Peruvian Amazon using Stylosanthes guianensis, a promising cover crop. Land Degrad. Dev. 2021, 32, 5143–5153. [Google Scholar] [CrossRef]
- Soto-Benavente, M.; Rodriguez-Achata, L.; Olivera, M.; Arostegui, V.; Colina, C.; Garate, J. Health risks due to the presence of heavy metals in agricultural products cultivated in areas abandoned by gold mining in the Peruvian Amazon. Sci. Agropecu. 2020, 11, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Conrado, D.; Rey-Benayas, J.; Costa, G.; Santos, S. Tree Communities in Three-Year-Old Post-Mining Sites Under Different Forest Restoration Techniques in the Brazilian Amazon. Forests 2020, 11, 527. [Google Scholar] [CrossRef]
- Ribeiro, S.; Cerveira, A.; Soares, P.; Fonseca, T. Natural Regeneration of Maritime Pine: A Review of the Influencing Factors and Proposals for Management. Forests 2022, 13, 386. [Google Scholar] [CrossRef]
- Hariharan, P.; Raman, T.R.S. Active restoration fosters better recovery of tropical rainforest birds than natural regeneration in degraded forest fragments. J. Appl. Ecol. 2022, 59, 274–285. [Google Scholar] [CrossRef]
- Moyo, B.; Ravhuhali, K.E. Abandoned Croplands: Drivers and Secondary Succession Trajectories under Livestock Grazing in Communal Areas of South Africa. Sustainability 2022, 14, 6168. [Google Scholar] [CrossRef]
- Brasil, A.B.; Schwartz, G.; Noronha, N.C.; Gama, M.A.P.; Ferreira, G.C. Natural regeneration for restoration of degraded areas after bauxite mining: A case study in the Eastern Amazon. Ecol. Eng. 2021, 171, 106392. [Google Scholar] [CrossRef]
- Günter, S.; Weber, M.; Erreis, R.; Aguirre, N. Influence of distance to forest edges on natural regeneration of abandoned pastures: A case study in the tropical mountain rain forest of Southern Ecuador. Eur. J. For. Res. 2007, 126, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Cubiña, A.; Aide, T.M. The Effect of Distance from Forest Edge on Seed Rain and Soil Seed Bank in a Tropical Pasture. Biotropica 2001, 33, 260–267. [Google Scholar] [CrossRef]
- Barros, T.C.; Elias, F.; Romano, L.L.; Ferreira, J. Natural recovery of plant species diversity in secondary forests in Eastern Amazonia: Contributions to passive forest restoration. Rev. Bras. Bot. 2020, 43, 165–175. [Google Scholar] [CrossRef]
- Mi, X.; Swenson, N.G.; Jia, Q.; Rao, M.; Feng, G.; Ren, H.; Bebber, D.P.; Ma, K. Stochastic assembly in a subtropical forest chronosequence: Evidence from contrasting changes of species, phylogenetic and functional dissimilarity over succession. Sci. Rep. 2016, 6, 32596. [Google Scholar] [CrossRef]
- Quintana, J.R.; Martín-Sanz, J.P.; Valverde-Asenjo, I.; Molina, J.A. Drought differently destabilizes soil structure in a chronosequence of abandoned agricultural lands. CATENA 2023, 222, 106871. [Google Scholar] [CrossRef]
- O’Brien, S.A.; Dehling, D.M.; Tylianakis, J.M. The recovery of functional diversity with restoration. Ecology 2022, 103, e3618. [Google Scholar] [CrossRef] [PubMed]
- Rocha, G.P.E.; Vieira, D.L.M.; Simon, M.F. Fast natural regeneration in abandoned pastures in southern Amazonia. For. Ecol. Manag. 2016, 370, 93–101. [Google Scholar] [CrossRef]
- Li, T.; Yang, H.; Yang, X.; Guo, Z.; Fu, D.; Liu, C.; Li, S.; Pan, Y.; Zhao, Y.; Xu, F.; et al. Community assembly during vegetation succession after metal mining is driven by multiple processes with temporal variation. Ecol. Evol. 2022, 12, e8882. [Google Scholar] [CrossRef] [PubMed]
- van der Sande, M.T.; Powers, J.S.; Kuyper, T.W.; Norden, N.; Salgado-Negret, B.; Silva de Almeida, J.; Bongers, F.; Delgado, D.; Dent, D.H.; Derroire, G.; et al. Soil resistance and recovery during neotropical forest succession. Philos. Trans. R. Soc. B Biol. Sci. 2023, 378, 20210074. [Google Scholar] [CrossRef]
- Matsuo, T.; Martínez-Ramos, M.; Bongers, F.; van der Sande, M.T.; Poorter, L. Forest structure drives changes in light heterogeneity during tropical secondary forest succession. J. Ecol. 2021, 109, 2871–2884. [Google Scholar] [CrossRef]
- Villa, P.M.; Martins, S.V.; de Oliveira Neto, S.N.; Rodrigues, A.C.; Safar, N.V.H.; Monsanto, L.D.; Cancio, N.M.; Ali, A. Woody species diversity as an indicator of the forest recovery after shifting cultivation disturbance in the northern Amazon. Ecol. Indic. 2018, 95, 687–694. [Google Scholar] [CrossRef]
- Martins, W.B.R.; de Matos Rodrigues, J.I.; de Oliveira, V.P.; Ribeiro, S.S.; dos Santos Barros, W.; Schwartz, G. Mining in the Amazon: Importance, impacts, and challenges to restore degraded ecosystems. Are we on the right way? Ecol. Eng. 2022, 174, 106468. [Google Scholar] [CrossRef]
- Velasquez-Ramirez, M.; Corvera, R.; Guerrero, J.; Thomas, E.; Russo, R.; Cusi, E.; Becerra, E.; Nascimiento, P.; Muñoz, A.; Vasquez, J.; et al. Guía Práctica: Recuperación de la Salud del Suelo para la Restauración de Áreas Degradadas por la Minería Aurífera Aluvial en la Amazonia Peruana, 1st ed.; Instituto de Investigaciones de la Amazonia Peruana (IIAP): Iquitos, Peru, 2022; ISBN 9786120065365. [Google Scholar]
- de Jesus Silva, C.V.; dos Santos, J.R.; Galvao, L.S.; da Silva, R.D.; Moura, Y.M. Floristic and structure of an Amazonian primary forest and a chronosequence of secondary succession. Acta Amaz. 2016, 46, 133–150. [Google Scholar] [CrossRef] [Green Version]
- Velásquez-Ramírez, M.; Vega Ruiz, C.; Gomringer, R.; Pillaca, M.; Thomas, E.; Stewart, P.M.; Gamarra Miranda, L.A.; Dañobeytia, F.R.; Guerrero Barrantes, J.A.; Gushiken, M.C.; et al. Mercury in soils impacted by alluvial gold mining in the Peruvian Amazon. J. Environ. Manag. 2021, 288, 112364. [Google Scholar] [CrossRef] [PubMed]
- Caballero, J.; Messinger, M.; Román-Dañobeytia, F.; Ascorra, C.; Fernandez, L.; Silman, M. Deforestation and Forest Degradation Due to Gold Mining in the Peruvian Amazon: A 34-Year Perspective. Remote Sens. 2018, 10, 1903. [Google Scholar] [CrossRef] [Green Version]
- Sáenz, G.; Finegan, B. Monitoreo de la Regeneración Natural con Fines de Manejo Forestal. Manejo Forestal Tropical No 15; CATIE: Turrialba, Costa Rica, 2000. [Google Scholar]
- Lozada, J. Consideraciones metodológicas sobre los estudios de comunidades forestales. Rev. For. Venez. 2010, 54, 77–88. [Google Scholar]
- Sugianto, S.; Muslih, A.M.; Ar-Rasyid, U.H.; Anhar, A. Vegetation analysis of rawa singkil wildlife reserve in Rantau Gedang Village, Singkil Distric, Aceh Singkil Regency, Aceh Province. IOP Conf. Ser. Earth Environ. Sci. 2021, 667, 012068. [Google Scholar] [CrossRef]
- Zhao, J.; Chen, J.; Chen, C.; Lu, S.; Song, C.; Liu, S.; Li, J.; Zhuang, C. Is it sufficient? Assessment of two sampling methods for urban plant species richness investigations. Urban For. Urban Green. 2023, 79, 127824. [Google Scholar] [CrossRef]
- Oksanen, J.; Minchin, P.R. Continuum theory revisited: What shape are species responses along ecological gradients? Ecol. Model. 2002, 157, 119–129. [Google Scholar] [CrossRef]
- Huisman, J.; Olff, H.; Fresco, L.F.M. A hierarchical set of models for species response analysis. J. Veg. Sci. 1993, 4, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Ahmadi, K.; Alavi, S.J.; Hosseini, S.M. Modeling response curves of European yew (Taxus baccata L.) using HOF models along the environmental gradient in north of Iran. Acta Ecol. Sin. 2022, 42, 383–391. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Dexter, E.; Rollwagen-Bollens, G.; Bollens, S.M. The trouble with stress: A flexible method for the evaluation of nonmetric multidimensional scaling. Limnol. Oceanogr. Methods 2018, 16, 434–443. [Google Scholar] [CrossRef]
- Lai, J.; Zou, Y.; Zhang, J.; Peres-Neto, P.R. Generalizing hierarchical and variation partitioning in multiple regression and canonical analyses using the rdacca.hp R package. Methods Ecol. Evol. 2022, 13, 782–788. [Google Scholar] [CrossRef]
- Dueñas, H.; Peña, J.; Báez, S.; Bejar, N.; Berrocal, S.; Garate, J. Influencia de la conservación de bosques en la diversidad y composición florística arbórea en el sureste de la Amazonía peruana. Cienc. Amaz. 2019, 7, 111–125. [Google Scholar] [CrossRef]
- Jakovac, C.C.; Peña-Claros, M.; Kuyper, T.W.; Bongers, F. Loss of secondary-forest resilience by land-use intensification in the Amazon. J. Ecol. 2015, 103, 67–77. [Google Scholar] [CrossRef]
- Goosem, M.; Paz, C.; Fensham, R.; Preece, N.; Goosem, S.; Laurance, S.G.W. Forest age and isolation affect the rate of recovery of plant species diversity and community composition in secondary rain forests in tropical Australia. J. Veg. Sci. 2016, 27, 504–514. [Google Scholar] [CrossRef]
- Banin, L.F.; Raine, E.H.; Rowland, L.M.; Chazdon, R.L.; Smith, S.W.; Rahman, N.E.B.; Butler, A.; Philipson, C.; Applegate, G.G.; Axelsson, E.P.; et al. The road to recovery: A synthesis of outcomes from ecosystem restoration in tropical and sub-tropical Asian forests. Philos. Trans. R. Soc. B Biol. Sci. 2023, 378, 20210090. [Google Scholar] [CrossRef]
- San-José, M.; Werden, L.; Joyce, F.H.; Reid, J.L.; Holl, K.D.; Zahawi, R.A. Effects of landscape structure on restoration success in tropical premontane forest. Sci. Rep. 2022, 12, 13452. [Google Scholar] [CrossRef] [PubMed]
- van Breugel, M.; Craven, D.; Lai, H.R.; Baillon, M.; Turner, B.L.; Hall, J.S. Soil nutrients and dispersal limitation shape compositional variation in secondary tropical forests across multiple scales. J. Ecol. 2019, 107, 566–581. [Google Scholar] [CrossRef]
- Rozendaal, D.M.A.; Bongers, F.; Aide, T.M.; Alvarez-Dávila, E.; Ascarrunz, N.; Balvanera, P.; Becknell, J.M.; Bentos, T.V.; Brancalion, P.H.S.; Cabral, G.A.L.; et al. Biodiversity recovery of Neotropical secondary forests. Sci. Adv. 2019, 5, eaau3114. [Google Scholar] [CrossRef] [Green Version]
- Garate-Quispe, J.S.; Canahuire-Robles, R.; Surco-Huacachi, O.; Alarcón-Aguirre, G. Desarrollo estructural y composición florística arbórea en áreas afectadas por minería aurífera en la Amazonía peruana: A 20 años de su reforestación. Rev. Mex. Biodivers. 2021, 92, e923437. [Google Scholar] [CrossRef]
- Fagundes, M.V.; Oliveira, R.S.; Fonseca, C.R.; Ganade, G. Nurse-target functional match explains plant facilitation strength. Flora Morphol. Distrib. Funct. Ecol. Plants 2022, 292, 152061. [Google Scholar] [CrossRef]
- Astudillo-Sánchez, E.; Pérez, J.; Troccoli, L.; Aponte, H. Composición, estructura y diversidad vegetal de la Reserva Ecológica Comunal Loma Alta, Santa Elena, Ecuador. Rev. Mex. Biodivers. 2019, 90, e902871. [Google Scholar] [CrossRef]
- Bonilla-Mata, R.; Acosta-Vargas, L.G. Dynamic and growth of the forests of the Isla del Coco National Park, Costa Rica. Rev. Biol. Trop. 2020, 68, S89–S102. [Google Scholar] [CrossRef]
- Vacek, Z.; Cukor, J.; Vacek, S.; Podrázský, V.; Linda, R.; Kovařík, J. Forest biodiversity and production potential of post-mining landscape: Opting for afforestation or leaving it to spontaneous development? For. J. 2018, 64, 116–126. [Google Scholar] [CrossRef]
- Villa, P.M.; Martins, S.V.; Diniz, É.S.; Nolasco de Oliveira Neto, S.; Neri, A.V.; Pinto-Junior, H.; Nunes, J.A.; Bueno, M.L.; Ali, A. Taxonomic and functional beta diversity of woody communities along Amazon forest succession: The relative importance of stand age, soil properties and spatial factor. For. Ecol. Manag. 2021, 482, 118885. [Google Scholar] [CrossRef]
- König, L.A.; Medina-Vega, J.A.; Longo, R.M.; Zuidema, P.A.; Jakovac, C.C. Restoration success in former Amazonian mines is driven by soil amendment and forest proximity. Philos. Trans. R. Soc. B Biol. Sci. 2023, 378, 20210086. [Google Scholar] [CrossRef]
- Hendy, A.; Hernandez-Acosta, E.; Valério, D.; Ferreira, N.; Reis, C.; Rodrigues, E.; Silva, E.; Andes, J.; Prado, F.; Margarete, V.; et al. Where boundaries become bridges: Mosquito community composition, key vectors, and environmental associations at forest edges in the central Brazilian Amazon. PLoS Negl. Trop. Dis. 2023, 17, e0011296. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Fu, S.; Yan, S.; Ren, C.; Wu, S.; Deng, J.; Li, B.; Han, X.; Yang, G. Responses of plant community to the linkages in plant-soil C:N:P stoichiometry during secondary succession of abandoned farmlands, China. J. Arid Land 2020, 12, 215–226. [Google Scholar] [CrossRef]
- de Paula, F.R.; Ruschel, A.R.; Felizzola, J.F.; Frauendorf, T.C.; de Barros Ferraz, S.F.; Richardson, J.S. Seizing resilience windows to foster passive recovery in the forest-water interface in Amazonian lands. Sci. Total Environ. 2022, 828, 154425. [Google Scholar] [CrossRef] [PubMed]
- Longworth, J.B.; Williamson, G.B. Composition and Diversity of Woody Plants in Tree Plantations Versus Secondary Forests in Costa Rican Lowlands. Trop. Conserv. Sci. 2018, 11, 194008291877329. [Google Scholar] [CrossRef] [Green Version]
- Rocchini, D.; Bacaro, G.; Chirici, G.; Da Re, D.; Feilhauer, H.; Foody, G.M.; Galluzzi, M.; Garzon-Lopez, C.X.; Gillespie, T.W.; He, K.S.; et al. Remotely sensed spatial heterogeneity as an exploratory tool for taxonomic and functional diversity study. Ecol. Indic. 2018, 85, 983–990. [Google Scholar] [CrossRef] [Green Version]
- Román-Dañobeytia, F.; Huayllani, M.; Michi, A.; Ibarra, F.; Loayza-Muro, R.; Vázquez, T.; Rodríguez, L.; García, M. Reforestation with four native tree species after abandoned gold mining in the Peruvian Amazon. Ecol. Eng. 2015, 85, 39–46. [Google Scholar] [CrossRef]
- Purvis, A.; Hector, A. Getting the measure of biodiversity. Nature 2000, 405, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Suding, K.; Higgs, E.; Palmer, M.; Callicott, J.B.; Anderson, C.B.; Baker, M.; Gutrich, J.J.; Hondula, K.L.; LaFevor, M.C.; Larson, B.M.H.; et al. Committing to ecological restoration. Science 2015, 348, 638–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yunanto, T.; Amanah, F.; Siregar, I.Z. The influence of fast-growing species composition on natural regeneration of coal mine reclamation. IOP Conf. Ser. Earth Environ. Sci. 2021, 918, 012048. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N | Abandonment Time | Distance to the Forest Edge | Species Richness | Abundance | Shannon-H | Margalef | α-Fisher |
|---|---|---|---|---|---|---|---|
| 1 | 10 | 60 | 13 | 73 | 1.76 | 2.80 | 4.60 |
| 2 | 3 | 110 | 10 | 65 | 1.77 | 2.16 | 3.30 |
| 3 | 4 | 125 | 11 | 68 | 1.99 | 2.37 | 3.72 |
| 4 | 20 | 40 | 15 | 61 | 2.20 | 3.41 | 6.35 |
| 5 | 23 | 35 | 20 | 66 | 2.41 | 4.54 | 9.76 |
| 6 | 11 | 110 | 21 | 66 | 2.73 | 4.77 | 10.63 |
| 7 | 14 | 70 | 23 | 55 | 2.69 | 5.49 | 14.86 |
| 8 | 5 | 150 | 10 | 28 | 2.01 | 2.70 | 5.57 |
| 9 | 2 | 260 | 8 | 43 | 1.41 | 1.86 | 2.90 |
| 10 | 6 | 250 | 5 | 12 | 1.42 | 1.61 | 3.22 |
| 11 | 7 | 139 | 16 | 50 | 2.16 | 3.83 | 8.14 |
| 12 | 7 | 227 | 9 | 41 | 1.60 | 2.15 | 3.56 |
| 13 | 3 | 560 | 4 | 21 | 1.12 | 0.99 | 1.47 |
| 14 | 6 | 120 | 15 | 73 | 1.74 | 3.26 | 5.72 |
| 15 | 4 | 318 | 8 | 19 | 1.78 | 2.38 | 5.21 |
| 16 | 4 | 195 | 12 | 48 | 1.74 | 2.84 | 5.14 |
| 17 | 5 | 175 | 11 | 45 | 1.69 | 2.63 | 4.64 |
| 18 | 5 | 171 | 11 | 42 | 1.59 | 2.68 | 4.85 |
| 19 | 8 | 250 | 13 | 20 | 2.32 | 4.01 | 16.10 |
| 20 | 6 | 150 | 12 | 35 | 2.11 | 3.09 | 6.45 |
| Reference values for the old-growth forest in Madre de Dios [46] | 82.7 ± 13 (61–125) | 215 ± 51 (129–323) | 3.9 ± 0.4 (2.4–4.5) | - | 53.3 ± 16 (24.5–90.8) | ||
| Reference values for early secondary forests (5 years after abandonment) in the Amazon [47] | 41.7 ± 10.4 (26–70) | - | 2.8 ± 0.4 (1.8–3.7 | - | - | ||
| Family | Species Richness | Abundance | Relative Abundance (%) | Basal Area (m2) | Relative Dominance (%) | Importance Index (%) |
|---|---|---|---|---|---|---|
| Fabaceae | 20 | 141 | 15.15 | 0.7369 | 28.76 | 21.95 |
| Malvaceae | 8 | 155 | 16.65 | 0.5498 | 21.46 | 19.05 |
| Urticaceae | 3 | 161 | 17.29 | 0.5139 | 20.05 | 18.67 |
| Asteraceae | 4 | 165 | 17.72 | 0.0658 | 2.57 | 10.15 |
| Piperaceae | 1 | 78 | 8.38 | 0.0644 | 2.51 | 5.45 |
| Moraceae | 6 | 15 | 1.61 | 0.2121 | 8.28 | 4.94 |
| Euphorbiaceae | 5 | 22 | 2.36 | 0.1316 | 5.14 | 3.75 |
| Verbenaceae | 2 | 60 | 6.44 | 0.0225 | 0.88 | 3.66 |
| Cannabaceae | 4 | 17 | 1.83 | 0.0530 | 2.07 | 1.95 |
| Rubiaceae | 2 | 16 | 1.72 | 0.0552 | 2.16 | 1.94 |
| Myrtaceae | 1 | 13 | 1.40 | 0.0134 | 0.52 | 0.96 |
| Hypericaceae | 2 | 14 | 1.50 | 0.0086 | 0.33 | 0.92 |
| Muntingiaceae | 1 | 10 | 1.07 | 0.0119 | 0.46 | 0.77 |
| Bixaceae | 1 | 8 | 0.86 | 0.0121 | 0.47 | 0.67 |
| Primulaceae | 1 | 3 | 0.32 | 0.0240 | 0.93 | 0.63 |
| Salicaceae | 3 | 8 | 0.86 | 0.0075 | 0.29 | 0.58 |
| Caryophyllaceae | 1 | 4 | 0.43 | 0.0179 | 0.70 | 0.56 |
| Meliaceae | 1 | 1 | 0.11 | 0.0219 | 0.85 | 0.48 |
| Poaceae | 1 | 8 | 0.86 | 0.0025 | 0.10 | 0.48 |
| Solanaceae | 2 | 6 | 0.64 | 0.0037 | 0.14 | 0.39 |
| Melastomataceae | 4 | 6 | 0.64 | 0.0034 | 0.13 | 0.39 |
| Polygonaceae | 1 | 4 | 0.43 | 0.0082 | 0.32 | 0.38 |
| Peraceae | 1 | 4 | 0.43 | 0.0046 | 0.18 | 0.30 |
| Rutaceae | 1 | 2 | 0.21 | 0.0099 | 0.39 | 0.30 |
| Malpighiaceae | 1 | 4 | 0.43 | 0.0034 | 0.13 | 0.28 |
| Boraginaceae | 4 | 2 | 0.21 | 0.0007 | 0.03 | 0.12 |
| Araliaceae | 1 | 1 | 0.11 | 0.0021 | 0.08 | 0.10 |
| Phyllanthaceae | 1 | 1 | 0.11 | 0.0007 | 0.03 | 0.07 |
| Simaroubaceae | 1 | 1 | 0.11 | 0.0006 | 0.02 | 0.06 |
| Gentianaceae | 1 | 1 | 0.11 | 0.0002 | 0.01 | 0.06 |
| Basal Area | Species Richness | Individuals | Diversity Index | |||
|---|---|---|---|---|---|---|
| Shannon | Margalef | α-Fisher | ||||
| (A) All individuals | ||||||
| Abandonment time | 0.48 * | 0.76 *** | 0.31 ns | 0.67 ** | 0.75 *** | 0.71 *** |
| Distance to the forest edge | −0.58 ** | −0.77 *** | −0.81 *** | −0.63 ** | −0.63 ** | −0.45 * |
| (B) Seedling category | ||||||
| Abandonment time | 0.48 * | 0.52 * | 0.09 ns | 0.42 ns | 0.56 * | 0.43 ns |
| Distance to the forest edge | −0.58 ** | −0.86 *** | −0.63 *** | −0.80 *** | −0.77 *** | −0.69 *** |
| (C) Sapling category | ||||||
| Abandonment time | 0.49 * | 0.61 ** | 0.30 ns | 0.58 * | 0.61 ** | 0.41 ns |
| Distance to the forest edge | −0.46 ns | −0.54 * | −0.51a * | −0.33a (ns) | −0.30a (ns) | −0.20 ns |
| (D) Tree category | ||||||
| Abandonment time | 0.45 ns | 0.59 * | 0.55 * | 0.62 ** | 0.59 * | 0.52 * |
| Distance to the forest edge | −0.50 * | −0.36 ns | −0.48 * | −0.33a (ns) | −0.35 ns | −0.37 ns |
| Variable | Predictors | Partial Correlation | Hierarchical Partitioning | p-Value | Model Adjusted R2 | ||
|---|---|---|---|---|---|---|---|
| Independent | Joint | Independent Effect (%) | |||||
| Basal Area | Abandonment time | 0.175 | 0.134 | 0.113 | 40.81 | 0.034 | 0.25 |
| Distance to the forest edge | −0.329 | 0.195 | 59.19 | ||||
| Species richness | Abandonment time | 0.341 | 0.274 | 0.231 | 40.91 | <0.001 | 0.631 |
| Distance to the forest edge | −0.578 | 0.396 | 59.09 | ||||
| Abundance | Abandonment time | −0.378 | 0.099 | 0.039 | 15.47 | <0.001 | 0.642 |
| Distance to the forest edge | −0.764 | 0.543 | 84.53 | ||||
| Shannon diversity index | Abandonment time | 0.38 | 0.27 | 0.193 | 49.89 | 0.001 | 0.488 |
| Distance to the forest edge | −0.382 | 0.272 | 50.11 | ||||
| Margalef diversity index | Abandonment time | 0.421 | 0.297 | 0.206 | 51.29 | <0.001 | 0.53 |
| Distance to the forest edge | −0.391 | 0.282 | 48.71 | ||||
| α-Fisher diversity index | Abandonment time | 0.469 | 0.277 | 0.103 | 72.79 | 0.017 | 0.308 |
| Distance to the forest edge | −0.027 | 0.104 | 27.21 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garate-Quispe, J.; Velásquez Ramírez, M.; Becerra-Lira, E.; Baez-Quispe, S.; Abril-Surichaqui, M.; Rodriguez-Achata, L.; Muñoz-Ushñahua, A.; Nascimento Herbay, P.; Fernandez-Mamani, Y.; Alarcon-Aguirre, G.; et al. Influence of Distance from Forest Edges on Spontaneous Vegetation Succession Following Small-Scale Gold Mining in the Southeast Peruvian Amazon. Diversity 2023, 15, 793. https://doi.org/10.3390/d15060793
Garate-Quispe J, Velásquez Ramírez M, Becerra-Lira E, Baez-Quispe S, Abril-Surichaqui M, Rodriguez-Achata L, Muñoz-Ushñahua A, Nascimento Herbay P, Fernandez-Mamani Y, Alarcon-Aguirre G, et al. Influence of Distance from Forest Edges on Spontaneous Vegetation Succession Following Small-Scale Gold Mining in the Southeast Peruvian Amazon. Diversity. 2023; 15(6):793. https://doi.org/10.3390/d15060793
Chicago/Turabian StyleGarate-Quispe, Jorge, Manuel Velásquez Ramírez, Edwin Becerra-Lira, Sufer Baez-Quispe, Milagro Abril-Surichaqui, Liset Rodriguez-Achata, Adenka Muñoz-Ushñahua, Pedro Nascimento Herbay, Yoni Fernandez-Mamani, Gabriel Alarcon-Aguirre, and et al. 2023. "Influence of Distance from Forest Edges on Spontaneous Vegetation Succession Following Small-Scale Gold Mining in the Southeast Peruvian Amazon" Diversity 15, no. 6: 793. https://doi.org/10.3390/d15060793
APA StyleGarate-Quispe, J., Velásquez Ramírez, M., Becerra-Lira, E., Baez-Quispe, S., Abril-Surichaqui, M., Rodriguez-Achata, L., Muñoz-Ushñahua, A., Nascimento Herbay, P., Fernandez-Mamani, Y., Alarcon-Aguirre, G., Herrera-Machaca, M., Hilares Vargas, L., Corvera Gomringer, R., & del Castillo Torres, D. (2023). Influence of Distance from Forest Edges on Spontaneous Vegetation Succession Following Small-Scale Gold Mining in the Southeast Peruvian Amazon. Diversity, 15(6), 793. https://doi.org/10.3390/d15060793