

Latitudinal Difference in the Condition Factor of Two Loliginidae Squid (Beka Squid and Indian Squid) in China Seas


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Data Sources
2.2. Marine Environmental Data
2.3. Statistical Analysis and Model Building
3. Result
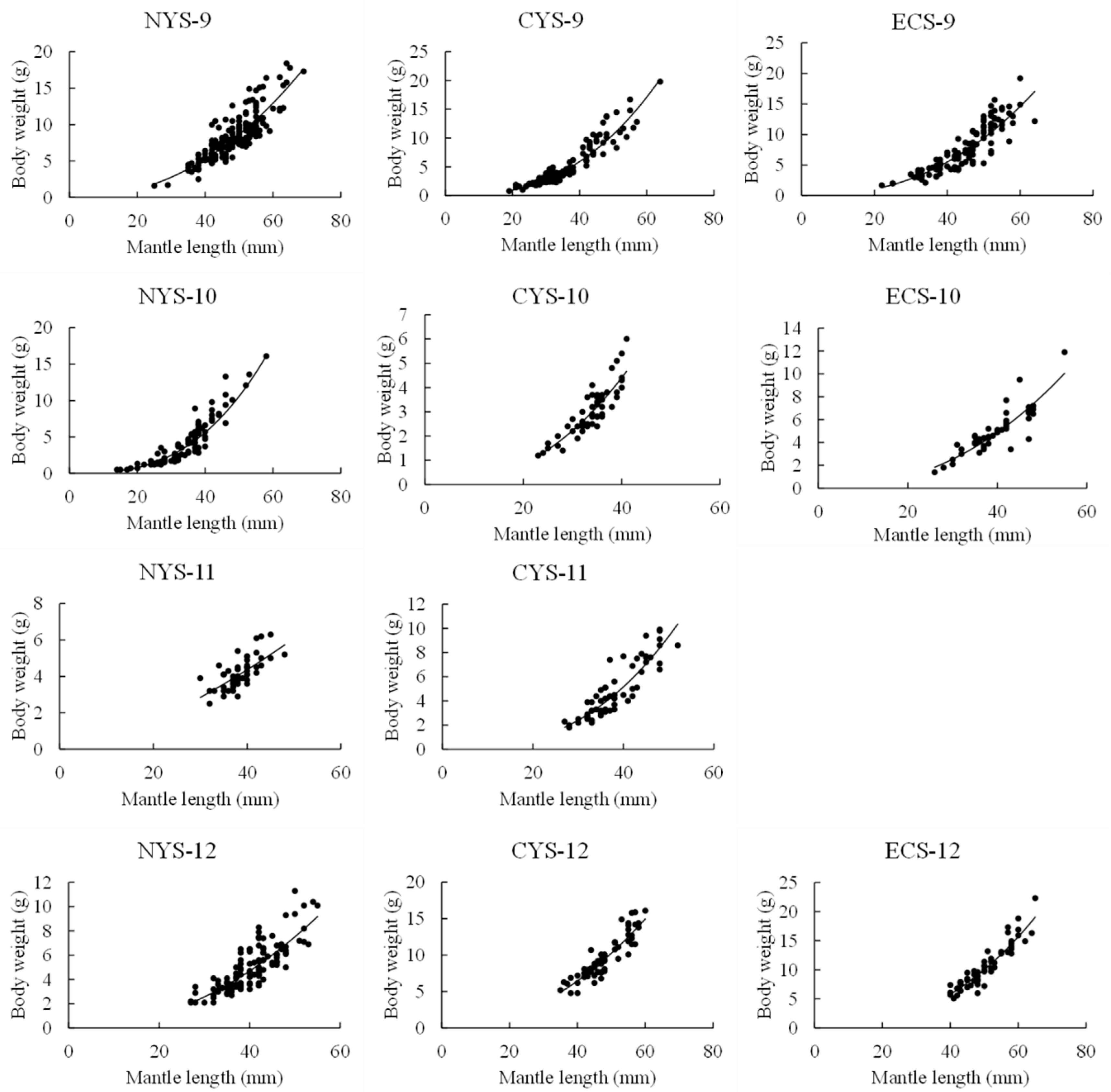
3.1. Mantle Length and Body–Weight Distribution
3.2. Length–Weight Relationship
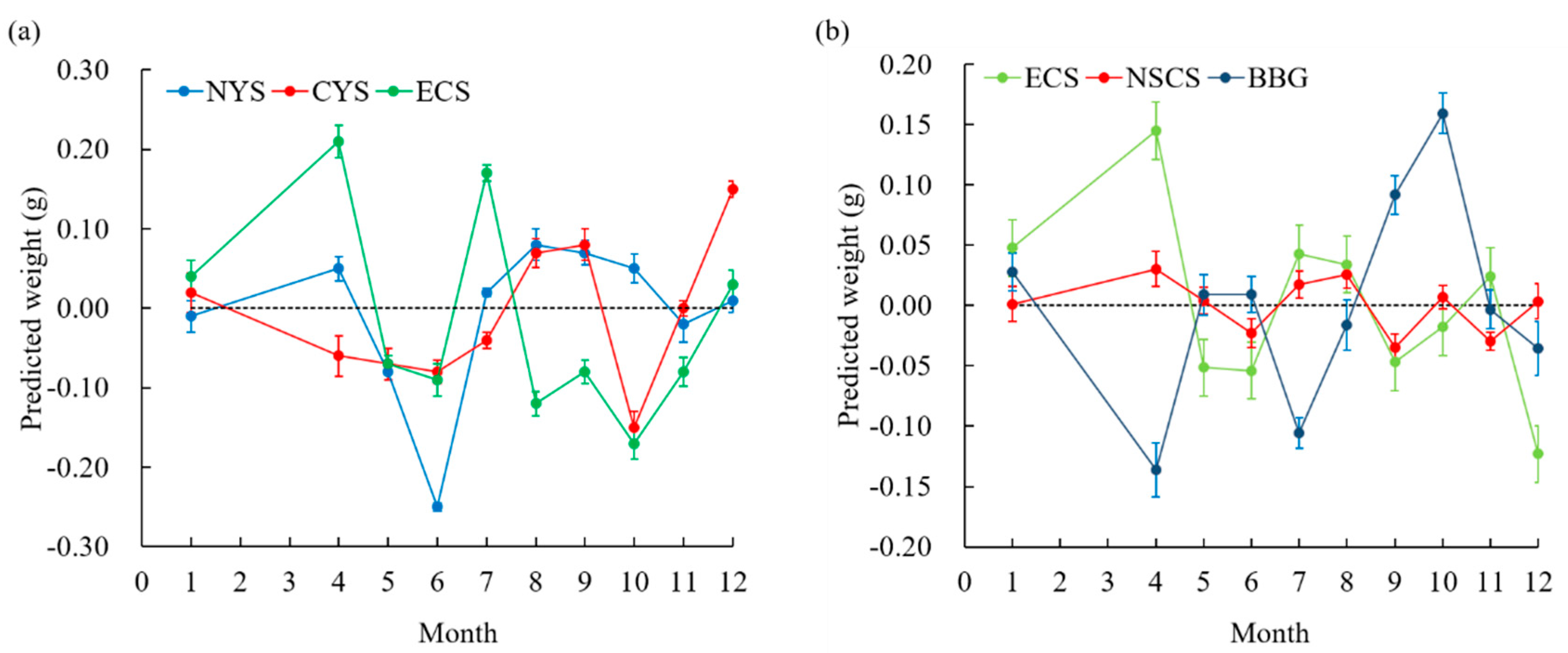
3.3. Variation in Weight Growth across Latitudes for Loligo beka and Uroteuthis duvaucelii
3.4. Temperature Effect on Weight Growth
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Luna, A.; Sánchez, P.; Chicote, C.; Gazo, M. Cephalopods in the diet of Risso’s dolphin (Grampus griseus) from the Mediterranean Sea: A review. Mar. Mammal Sci. 2022, 38, 725–741. [Google Scholar] [CrossRef]
- Arkhipkin, A.I.; Hendrickson, L.C.; Ignaccio, P.; Pierce, G.J.; Roa-Ureta, R.H.; Jran-Paul, R.; Andreas, W. Stock assessment and management of cephalopods: Advances and challenges for short-lived fishery resources. ICES J. Mar. Sci. 2021, 78, 714–730. [Google Scholar] [CrossRef]
- Arkhipkin, A.I.; Rodhouse, P.G.K.; Pierce, G.J.; Sauer, W.; Sakai, M.; Allcock, L.; Arguelles, J.; Bower, J.R.; Castillo, G.; Ceriola, L.; et al. World Squid Fisheries. Rev. Fish. Sci. Aquac. 2015, 23, 92–252. [Google Scholar] [CrossRef]
- Pierce, G.J.; Valavanis, V.D.; Guerra, A.; Jereb, P.; Orsi-Relini, L.; Bellido, J.M.; Katara, I.; Piatkowski, U.; Pereira, J.; Balguerias, E.; et al. A review of cephalopod–environment interactions in European Seas. Hydrobiologia 2008, 612, 49–70. [Google Scholar] [CrossRef]
- Bravo de Laguna, J. Managing an international multispecies fishery: The Saharn trawl fishery for cephalopods. In Marine Invertebrate Fisheries: Their Assessment and Management; Caddy, J.F., Ed.; John Wiley: New York, NY, USA, 1989; Volume 225. [Google Scholar]
- Caddy, J.F.; Rodhouse, P.G. Cephalopod and groundfish landings: Evidence for ecological change in global fisheries? Rev. Fish Biol. Fish. 1998, 8, 431–444. [Google Scholar] [CrossRef]
- Boyle, P.; Rodhouse, P. Cephalopods: Ecology and Fisheries; Blackwell Publishing: Oxford, UK, 2005. [Google Scholar]
- Clarke, M.R. Cephalopods as prey. III Cetaceans. Philos. Trans. R. Soc. B Biol. Sci. 1996, 351, 1053–1065. [Google Scholar]
- Doubleday, Z.A.; Prowse, T.A.; Arkhipkin, A.; Pierce, G.J.; Semmens, J.; Steer, M.; Leporati, S.C.; Lourenco, S.; Quetglas, A.; Sauer, W.; et al. Global proliferation of cephalopods. Curr. Biol. 2016, 26, 406–407. [Google Scholar] [CrossRef]
- Watson, R.; Pauly, D. Systematic distortions in world fisheries catch trends. Nature 2001, 414, 534–536. [Google Scholar] [CrossRef]
- Pang, Y.M.; Tian, Y.J.; Fu, C.H.; Wang, B.; Li, J.C.; Ren, Y.P.; Wan, R. Variability of coastal cephalopods in overexploited China Seas under climate change with implications on fisheries management. Fish. Res. 2018, 208, 22–33. [Google Scholar] [CrossRef]
- Jin, Y.; Chen, X.J. Review on studies of cephalopoda in the coastal waters of China. Mar. Fish. 2017, 39, 696–712. [Google Scholar]
- Pang, Y.M.; Tian, Y.J.; Fu, C.H.; Ren, Y.P.; Wan, R. Growth and distribution of Amphioctopus fangsiao (d’Orbigny, 1839–1841) in Haizhou Bay, Yellow Sea. J. Ocean. Univ. China 2020, 19, 1125–1132. [Google Scholar] [CrossRef]
- Díaz-Santana-Iturrios, M.; Salinas-Zavala, C.A.; Granados-Amores, J.; de la Cruz-Agüero, J.; García-Rodríguez, F.J. Taxonomic considerations of squids of the family Loliginidae (Cephalopoda: Myopsida) supported by morphological, morphometric, and molecular data. Mar. Biodivers. 2019, 49, 2401–2409. [Google Scholar] [CrossRef]
- Perez, J.A.A.; Aguiar, D.C.d.; Santos, J.A.T.d. Gladius and statolith as tools for age and growth studies of the squid Loligo plei (Teuthida: Loliginidae) off Southern Brazil. Braz. Arch. Biol. Technol. 2006, 49, 747–755. [Google Scholar] [CrossRef]
- Ceriola, L.; Jackson, G.D. Growth, hatch size and maturation in a southern population of the loliginid squid Loliolus noctiluca. J. Mar. Biol. Assoc. UK 2010, 90, 755–767. [Google Scholar] [CrossRef]
- Chen, X.J.; Wang, R.G.; Qian, W.G. Important Economic Cephalopod Resources and Fisheries in the Coastal Waters of China; Science Press: Beijing, China, 2013; pp. 64–65. [Google Scholar]
- Guo, J.Z.; Liu, D.; Zhang, C.; Tian, Y.J.; Li, Z.X. Using statolith shape analysis to identify five commercial Loliginidae squid species in Chinese waters. J. Oceanol. Limnol. 2021, 39, 1160–1168. [Google Scholar] [CrossRef]
- Dong, Z.Z. Chinese Fauna, Mollusca, Cephalopods; Science Press: Beijing, China, 1988. [Google Scholar]
- Du, T.F. Resources Assessment for Cephalopod in Offshore Water of China and Classification of Genus Level of Squids; Shanghai Ocean University: Shanghai, China, 2016. [Google Scholar]
- Jereb, P.; Roper, C.F.E. Cephalopods of the world. An Annotated and Illustrated Catalogue of Cephalopod Species Known to Date. Volume 2. Myopsid and Oegopsid Squids; Food and Agriculture Organization of the United Nations: Rome, Italy, 2010. [Google Scholar]
- Sabrah, M.M.; El-Sayed, A.Y.; El-Ganiny, A.A. Fishery and population characteristics of the Indian squids Loligo duvauceli Orbigny, 1848 from trawl survey along the north-west Red Sea. Egypt. J. Aquat. Res. 2015, 41, 279–285. [Google Scholar] [CrossRef]
- Sun, M.S.; Xu, Y.W.; Li, J.J.; Huang, Z.R.; Chen, Z.Z.; Yang, C.P.; Liu, W.D.; Zhang, H. Dominant species of cephalopods and their niche characteristics in offshore of northern South China Sea. Chin. J. Appl. Ecol. 2020, 31, 2793–2803. [Google Scholar]
- Jin, Y.; Jin, X.; Gorfine, H.; Wu, Q.; Shan, X. Modeling the oceanographic impacts on the spatial distribution of common cephalopods during autumn in the Yellow Sea. Front. Mar. Sci. 2020, 7, 432. [Google Scholar] [CrossRef]
- Pecl, G.T.; Jackson, G.D. The potential impacts of climate change on inshore squid: Biology, ecology and fisheries. Rev. Fish Biol. Fish. 2008, 18, 373–385. [Google Scholar] [CrossRef]
- Wang, K.-Y.; Lee, K.-T.; Liao, C.-H. Age, growth and maturation of Swordtip Squid (Photololigo edulis) in the Southern East China Sea. J. Mar. Sci. Technol. 2010, 18, 99–105. [Google Scholar] [CrossRef]
- Wang, K.Y.; Chang, K.Y.; Liao, C.H.; Lee, M.A.; Lee, K.T. Growth strategies of the Swordtip Squid, Uroteuthis edulis, in response to environmental changes in the Southern East China Sea—A cohort analysis. Bull. Mar. Sci. 2013, 89, 677–698. [Google Scholar] [CrossRef]
- Jin, Y.; Li, N.; Chen, X.; Liu, B.; Li, J. Comparative age and growth of Uroteuthis chinensis and Uroteuthis edulis from China Seas based on statolith. Aquac. Fish. 2019, 4, 166–172. [Google Scholar] [CrossRef]
- Pang, Y.M.; Chen, C.-S.; Iwata, Y. Variations in female swordtip squid Uroteuthis edulis life history traits between southern Japan and northern Taiwan (Northwestern Pacific). Fish. Sci. 2020, 86, 1005–1017. [Google Scholar] [CrossRef]
- Wang, D.L.; Yao, L.; Yu, J.; Chen, P. The role of environmental factors on the fishery catch of the squid Uroteuthis chinensis in the Pearl River Estuary, China. J. Mar. Sci. Eng. 2021, 9, 131. [Google Scholar] [CrossRef]
- Wu, C.; Liu, W.; Chen, M. Complete mitochondrial genome of the Loligo beka. Mitochondrial DNA Part A 2016, 27, 4278–4279. [Google Scholar]
- Islam, R.; Hajisamae, S.; Pradit, S.; Perngmak, P.; Paul, M. Feeding habits of two sympatric loliginid squids, Uroteuthis (Photololigo) chinensis (Gray, 1849) and Uroteuthis (Photololigo) duvaucelii (d’Orbigny, 1835), in the lower part of the South China Sea. Molluscan Res. 2018, 38, 155–162. [Google Scholar] [CrossRef]
- LeCren, E.D. The length-weight relationship and seasonal cycle in gonad weight and condition in the Perch (Perca fluviatilis). J. Anim. Ecol. 1951, 20, 201–219. [Google Scholar]
- Ferreri, G.A.B. Length—Weight relationships and condition factors of the Humboldt Squid (Dosidicus gigas) from the Gulf of california and the Pacific Ocean. J. Shellfish. Res. 2014, 33, 769–780. [Google Scholar] [CrossRef]
- McBride, R.S. The continuing role of life history parameters to identify stock structure. In Stock Identification Methods: Applications in Fisheries Science, 2nd ed.; Elsevier: London, UK, 2014. [Google Scholar]
- Goicochea-Vigo, C.; Morales-Bojórquez, E.; Zepeda-Benitez, V.Y.; Hidalgo-de-la-Toba, J.Á.; Aguirre-Villaseñor, H.; Mostacero-Koc, J.; Atoche-Suclupe, D. Age and growth estimates of the jumbo flying squid (Dosidicus gigas) off Peru. Aquat. Living Resour. 2019, 32, 7. [Google Scholar] [CrossRef]
- Barrios, A.; Ernande, B.; Mahé, K.; Trenkel, V.; Rochet, M.J. Utility of mixed effects models to inform the stock structure of whiting in the Northeast Atlantic Ocean. Fish. Res. 2017, 190, 132–139. [Google Scholar] [CrossRef]
- Chen, X.J.; Li, J.; Liu, B.; Chen, Y.; Li, G.; Fang, Z.; Tian, S. Age, growth and population structure of jumbo flying squid, Dosidicus gigas, off the Costa Rica Dome. J. Mar. Biol. Assoc. UK 2012, 93, 567–573. [Google Scholar] [CrossRef]
- Arkhipkin, A.I. Diversity in growth and longevity in short-lived animals: Squid of the suborder Oegopsina. Mar. Freshw. Res. 2004, 55, 341–355. [Google Scholar] [CrossRef]
- Lipiński, M.R.; Underhill, L.G. Sexual maturation in squid: Quantum or continuum? S. Afr. J. Mar. Sci. 1995, 15, 207–223. [Google Scholar] [CrossRef]
- Froese, R. Cube law, condition factor and weight-length relationships: History, meta-analysis and recommendations. J. Appl. Ichthyol. 2006, 22, 241–253. [Google Scholar] [CrossRef]
- Ma, Q.Y.; Jiao, Y.; Ren, Y.P. Linear mixed-effects models to describe length-weight relationships for yellow croaker (Larimichthys Polyactis) along the north coast of China. PLoS ONE 2017, 12, e0171811. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Multimodel inference: Understanding AIC and BIC in model selection. Sociol. Methods Res. 2004, 33, 261–304. [Google Scholar] [CrossRef]
- Nakagawa, S.; Schielzeth, H. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Semmens, J.M.; Pecl, G.T.; Villanueva, R.; Jouffre, D.; Sobrino, I.; Wood, J.B.; Rigby, P.R. Understanding octopus growth: Patterns, variability and physiology. Mar. Freshw. Res. 2004, 55, 367–377. [Google Scholar] [CrossRef]
- Morrongiello, J.R.; Thresher, R.E.; Smith, D.C. Aquatic biochronologies and climate change. Nat. Clim. Chang. 2012, 2, 849–857. [Google Scholar] [CrossRef]
- Begg, G.A.; Friedland, K.D.; Pearce, J.B. Stock identification and its role in stock assessment and fisheries management: An overview. Fish. Res. 1999, 43, 1–8. [Google Scholar] [CrossRef]
- Reiss, H.; Hoarau, G.; Dickey-Collas, M.; Wolff, W.J. Genetic population structure of marine fish: Mismatch between biological and fisheries management units. Fish Fish. 2009, 10, 361–395. [Google Scholar] [CrossRef]
- Sukramongkol, N.; Tsuchiya, K.; Tokai, T.; Segawa, S. Fishery biology of Loligo edulis in Moroiso Bay, Kanagawa Prefecture, Japan. La Mer 2006, 44, 131–143. [Google Scholar]
- Mulyono, M.; Nuraini, A.; Dewi, I.J.; Kritiani, M.G.; Syamsudin, S. Biology aspects and length-weight relationship of squid Loligo chinensis in the waters of Lamongan Regency, East Java Province, Indonesia. AACL Bioflux 2017, 10, 1221–1225. [Google Scholar]
- Qiu, X.Y. Biological characteristics and stock changes of Loligo japonica in the Yellow Sea. Mar. Fish. Res. 1986, 7, 109–120. [Google Scholar]
- Wu, Y.Q.; Zhang, B.L. Ecology of the economic invertebrates in the Bohai Sea. Mar. Sci. 1990, 14, 48–52. [Google Scholar]
- Yunrong, Y.; Yuyuan, L.; Shengyun, Y.; Guirong, W.; Yajin, T.; Qibin, F.; Huosheng, L. Biological characteristics and spatial—Temporal distribution of Mitre Squid, Uroteuthis Chinensis, in the Beibu Gulf, South China Sea. J. Shellfish. Res. 2013, 32, 835–844. [Google Scholar]
- Li, N.; Fang, Z.; Chen, X.J. Study on microstructure and growth characteristics of Uroteuthis edulis statolith in East China Sea. South China Fish. Sci. 2020, 16, 21–31. [Google Scholar]
- Forsythe, J.W. Accounting for the effect of temperature on squid growth in nature: From hypothesis to practice. Mar. Freshw. Res. 2004, 55, 331–339. [Google Scholar] [CrossRef]
- Sasikumar, G.; Mohamed, K.S.; Mini, K.G.; Sajikumar, K.K. Effect of tropical monsoon on fishery abundance of Indian squid (Uroteuthis (Photololigo) duvaucelii). J. Nat. Hist. 2018, 52, 751–766. [Google Scholar] [CrossRef]
- Quetglas, A.; Ordines, F.; Valls, M. What drives seasonal fluctuations of body condition in a semelparous income breeder octopus? Acta Oecol. 2011, 37, 476–483. [Google Scholar] [CrossRef]
- Fogarty, M.J.; Sissenwine, M.P.; Cohen, E.B. Recruitment variability and the dynamics of exploited marine populations. Trends Ecol. Evol. 1991, 6, 241–246. [Google Scholar] [CrossRef]
- Reed, T.E.; Waples, R.S.; Schindler, D.E.; Hard, J.J.; Kinnison, M.T. Phenotypic plasticity and population viability: The importance of environmental predictability. Proc. R. Soc. B Biol. Sci. 2010, 277, 3391–3400. [Google Scholar] [CrossRef]
- Green, C.P.; Robertson, S.G.; Hamer, P.A.; Virtue, P.; Jackson, G.D.; Moltschaniwskyj, N.A. Combining statolith element composition and Fourier shape data allows discrimination of spatial and temporal stock structure of arrow squid (Nototodarus gouldi). Can. J. Fish. Aquat. Sci. 2015, 72, 1609–1618. [Google Scholar] [CrossRef]
- Agus, B.; Mereu, M.; Cannas, R.; Cau, A.; Coluccia, E.; Follesa, M.C.; Cuccu, D. Age determination of Loligo vulgaris and Loligo forbesii using eye lens analysis. Zoomorphology 2018, 137, 63–70. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Region | Stations | Numbers | Year | Month | Body Weight/g | Mantle Length/mm | ||
|---|---|---|---|---|---|---|---|---|---|
| Range | Mean | Range | Mean | ||||||
| Loligo beka | NYS | DL | 660 | 2019 | 9–12 | 0.5–20.6 | 6.6 ± 2.9 | 14–78 | 44.6 ± 8.0 |
| 2020 | 1, 4–9 | ||||||||
| YT | 103 | 2019 | 6, 7 | 2.4–40.8 | 13.5 ± 8.1 | 42–83 | 61.2 ± 10.7 | ||
| SD | 170 | 2019 | 9, 10, 12 | 1.2–18.4 | 5.6 ± 3.9 | 20–65 | 39.9 ± 9.4 | ||
| CYS | QD | 659 | 2019 | 6–12 | 0.8–41.3 | 8.6 ± 5.3 | 19–87 | 47.1 ± 12.5 | |
| 2020 | 1, 4, 8, 9 | ||||||||
| LYG | 209 | 2019 | 6, 7 | 2.6–24.3 | 9.8 ± 4.2 | 32–72 | 51.1 ± 7.2 | ||
| 2020 | 7 | ||||||||
| SYS | NT | 60 | 2020 | 5 | 8.7–43.1 | 23.7 ± 8.8 | 45–83 | 61.4 ± 9.0 | |
| ECS | NB | 240 | 2019 | 8–10 | 1.4–29.1 | 7.6 ± 3.8 | 22–68 | 45.8 ± 8.7 | |
| 2020 | 4 | ||||||||
| ND | 386 | 2019 | 12 | 2.4–31.3 | 9.2 ± 3.8 | 27–83 | 47.6 ± 7.2 | ||
| 2020 | 1, 5–9 | ||||||||
| ZZ | 50 | 2019 | 8 | 3.5–12.2 | 7.3 ± 2.2 | 38–58 | 46.6 ± 4.5 | ||
| NSCS | ST | 2 | 2019 | 8 | 6.1–12.6 | 9.4 ± 3.3 | 42–55 | 48.5 ± 6.5 | |
| Uroteuthis duvaucelii | ECS | NB | 307 | 2019 | 11 | 3.3–66.7 | 25.2 ± 13.7 | 37–118 | 79.2 ± 18.5 |
| 2020 | 1, 9–11 | ||||||||
| ND | 293 | 2019 | 11, 12 | 2.1–51.5 | 20.7 ± 11.1 | 28–124 | 77.7 ± 25.3 | ||
| 2020 | 8, 9, 11 | ||||||||
| ZZ | 764 | 2019 | 8–12 | 1.5–57.5 | 15.5 ± 10.3 | 28–132 | 69.0 ± 19.3 | ||
| 2020 | 1, 4–8 | ||||||||
| NSCS | ST | 840 | 2019 | 8, 10–12 | 2.4–63.1 | 12.4 ± 7.3 | 34–121 | 61.6 ± 11.6 | |
| 2020 | 1, 4–11 | ||||||||
| YJ | 411 | 2020 | 5–11 | 12.9–82.0 | 33.4 ± 8.8 | 65–138 | 99.1 ± 10.7 | ||
| BBG | BH | 778 | 2019 | 7, 9, 12 | 3.6–138.0 | 16.3 ± 9.38 | 40–170 | 69.1 ± 14.3 | |
| 2020 | 1, 4–11 | ||||||||
| HK | 108 | 2019 | 7, 8, 10 | 16.4–92.9 | 38.9 ± 16.9 | 68–145 | 96.2 ± 17.2 | ||
| DZ | 100 | 2020 | 6 | 18.6–160.9 | 80.0 ± 38.0 | 60–215 | 143.5 ± 41.9 | ||
| LG | 50 | 2020 | 5 | 27.6–125.7 | 67.7 ± 20.3 | 99–171 | 120.5 ± 13.2 | ||
| Model Factors | Loligo beka | Uroteuthis duvaucelii | ||||
|---|---|---|---|---|---|---|
| df | AICc | R2 | df | AICc | R2 | |
| ln(BW) = ln(a) + b × ln(ML) | 3 | −290.08 | 0.84 | 3 | −1657 | 0.92 |
| ln(BW) = ln(a) + b × ln(ML) + Month | 4 | −403.49 | 0.85 | 4 | −1732 | 0.93 |
| ln(BW) = ln(a) + b × ln(ML) + Sex + Month | 5 | −532.64 | 0.86 | 5 | −1997 | 0.93 |
| ln(BW) = ln(a) + b × ln(ML) + Sex + lnML × Sex + Month | 6 | −528.19 | 0.86 | 6 | −1999 | 0.93 |
| ln(BW) = ln(a) + b × ln(ML) + Sex + lnML × Sex + Region + Month | 7 | −692.25 | 0.87 | 7 | −2068 | 0.94 |
| Areas | Sex | Month | N | a | b | R2 |
|---|---|---|---|---|---|---|
| Northern Yellow Sea | All | All | 933 | 0.0009 | 2.3363 | 0.8537 |
| Male | All | 682 | 0.0014 | 2.2022 | 0.7590 | |
| Female | All | 251 | 0.0010 | 2.3179 | 0.8658 | |
| All | 1 | 60 | 0.0066 | 1.7739 | 0.7634 | |
| All | 4 | 60 | 0.0035 | 1.9839 | 0.8581 | |
| All | 5 | 60 | 0.0147 | 1.5506 | 0.5202 | |
| All | 6 | 92 | 0.0013 | 2.1542 | 0.4696 | |
| All | 7 | 131 | 0.0004 | 2.5229 | 0.8908 | |
| All | 8 | 60 | 0.0032 | 2.0188 | 0.7813 | |
| All | 9 | 190 | 0.0015 | 2.2192 | 0.7165 | |
| All | 10 | 100 | 0.0002 | 2.8383 | 0.8708 | |
| All | 11 | 60 | 0.0177 | 1.4937 | 0.4864 | |
| All | 12 | 120 | 0.0020 | 2.1009 | 0.6901 | |
| Central Yellow Sea | All | All | 868 | 0.0010 | 2.3269 | 0.8166 |
| Male | All | 608 | 0.0011 | 2.2920 | 0.7748 | |
| Female | All | 260 | 0.0014 | 2.2571 | 0.7846 | |
| All | 1 | 60 | 0.0063 | 1.8498 | 0.5441 | |
| All | 4 | 60 | 0.0006 | 2.4257 | 0.6965 | |
| All | 6 | 114 | 0.0013 | 2.2362 | 0.7311 | |
| All | 7 | 204 | 0.0009 | 2.3381 | 0.8297 | |
| All | 8 | 130 | 0.0002 | 2.7131 | 0.7609 | |
| All | 9 | 130 | 0.0004 | 2.6210 | 0.8910 | |
| All | 10 | 50 | 0.0007 | 2.3745 | 0.7791 | |
| All | 11 | 60 | 0.0003 | 2.6568 | 0.7957 | |
| All | 12 | 60 | 0.0031 | 2.0699 | 0.8244 | |
| East China Sea | All | All | 676 | 0.0012 | 2.2957 | 0.7306 |
| Male | All | 439 | 0.0025 | 2.0792 | 0.6619 | |
| Female | All | 237 | 0.0018 | 2.2003 | 0.6786 | |
| All | 1 | 60 | 0.0013 | 2.2874 | 0.9103 | |
| All | 4 | 60 | 0.0138 | 1.6969 | 0.5453 | |
| All | 5 | 60 | 0.0012 | 2.2890 | 0.6969 | |
| All | 6 | 60 | 0.0105 | 1.7204 | 0.6111 | |
| All | 7 | 60 | 0.0042 | 1.9954 | 0.7792 | |
| All | 8 | 146 | 0.0038 | 1.9612 | 0.5448 | |
| All | 9 | 120 | 0.0008 | 2.4053 | 0.7911 | |
| All | 10 | 50 | 0.0011 | 2.2671 | 0.7593 | |
| All | 12 | 60 | 0.0007 | 2.4402 | 0.8686 |
| Areas | Sex | Month | N | a | b | R2 |
|---|---|---|---|---|---|---|
| East China Sea | All | All | 1364 | 0.0010 | 2.2624 | 0.9000 |
| Male | All | 1190 | 0.0009 | 2.2868 | 0.8552 | |
| Female | All | 174 | 0.0165 | 1.6519 | 0.7499 | |
| All | 1 | 180 | 0.0018 | 2.1305 | 0.7200 | |
| All | 4 | 60 | 0.0005 | 2.4799 | 0.8659 | |
| All | 5 | 60 | 0.0008 | 2.2859 | 0.9149 | |
| All | 6 | 60 | 0.0012 | 2.2054 | 0.8609 | |
| All | 7 | 84 | 0.0031 | 2.0187 | 0.8883 | |
| All | 8 | 150 | 0.0002 | 2.6204 | 0.7504 | |
| All | 9 | 153 | 0.0001 | 2.7474 | 0.9401 | |
| All | 10 | 110 | 0.0001 | 2.9671 | 0.8073 | |
| All | 11 | 367 | 0.0025 | 2.0539 | 0.7686 | |
| All | 12 | 140 | 0.0005 | 2.4164 | 0.9259 | |
| Northern South China Sea | All | All | 1251 | 0.0008 | 2.3301 | 0.9556 |
| Male | All | 1149 | 0.0008 | 2.3167 | 0.9359 | |
| Female | All | 102 | 0.0007 | 2.3547 | 0.9403 | |
| All | 1 | 60 | 0.0002 | 2.6819 | 0.9208 | |
| All | 4 | 60 | 0.0019 | 2.1194 | 0.9120 | |
| All | 5 | 120 | 0.0004 | 2.4956 | 0.9322 | |
| All | 6 | 103 | 0.0004 | 2.4508 | 0.9786 | |
| All | 7 | 120 | 0.0018 | 2.1289 | 0.9553 | |
| All | 8 | 128 | 0.0003 | 2.5403 | 0.9599 | |
| All | 9 | 120 | 0.0032 | 1.9930 | 0.9522 | |
| All | 10 | 170 | 0.0013 | 2.2080 | 0.9686 | |
| All | 11 | 310 | 0.0003 | 2.5018 | 0.8800 | |
| All | 12 | 60 | 0.0011 | 2.2538 | 0.8399 | |
| Beibu Gulf | All | All | 1036 | 0.0012 | 2.2328 | 0.9224 |
| Male | All | 836 | 0.0018 | 2.1354 | 0.9594 | |
| Female | All | 200 | 0.0011 | 2.2969 | 0.8065 | |
| All | 1 | 60 | 0.0275 | 1.4695 | 0.7084 | |
| All | 4 | 60 | 0.0004 | 2.4517 | 0.9161 | |
| All | 5 | 110 | 0.0001 | 2.7653 | 0.8691 | |
| All | 6 | 137 | 0.0037 | 1.9915 | 0.9435 | |
| All | 7 | 191 | 0.0014 | 2.1817 | 0.8318 | |
| All | 8 | 68 | 0.0015 | 2.1724 | 0.9031 | |
| All | 9 | 120 | 0.0019 | 2.1568 | 0.6975 | |
| All | 10 | 110 | 0.0005 | 2.4722 | 0.8936 | |
| All | 11 | 120 | 0.0011 | 2.2393 | 0.7669 | |
| All | 12 | 60 | 0.0023 | 2.0667 | 0.7075 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, J.; Zhang, C.; Li, Z.; Liu, D.; Tian, Y. Latitudinal Difference in the Condition Factor of Two Loliginidae Squid (Beka Squid and Indian Squid) in China Seas. Diversity 2023, 15, 812. https://doi.org/10.3390/d15070812
Guo J, Zhang C, Li Z, Liu D, Tian Y. Latitudinal Difference in the Condition Factor of Two Loliginidae Squid (Beka Squid and Indian Squid) in China Seas. Diversity. 2023; 15(7):812. https://doi.org/10.3390/d15070812
Chicago/Turabian StyleGuo, Jianzhong, Chi Zhang, Zhixin Li, Dan Liu, and Yongjun Tian. 2023. "Latitudinal Difference in the Condition Factor of Two Loliginidae Squid (Beka Squid and Indian Squid) in China Seas" Diversity 15, no. 7: 812. https://doi.org/10.3390/d15070812
APA StyleGuo, J., Zhang, C., Li, Z., Liu, D., & Tian, Y. (2023). Latitudinal Difference in the Condition Factor of Two Loliginidae Squid (Beka Squid and Indian Squid) in China Seas. Diversity, 15(7), 812. https://doi.org/10.3390/d15070812