

Hydrolithon farinosum and Lithophyllum epiphyticum sp. nov. (Corallinaceae, Corallinales, Rhodophyta), Two Epiphytic Crustose Coralline Algae from the Abrolhos Archipelago, Brazil, Southwestern Atlantic
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
3. Results
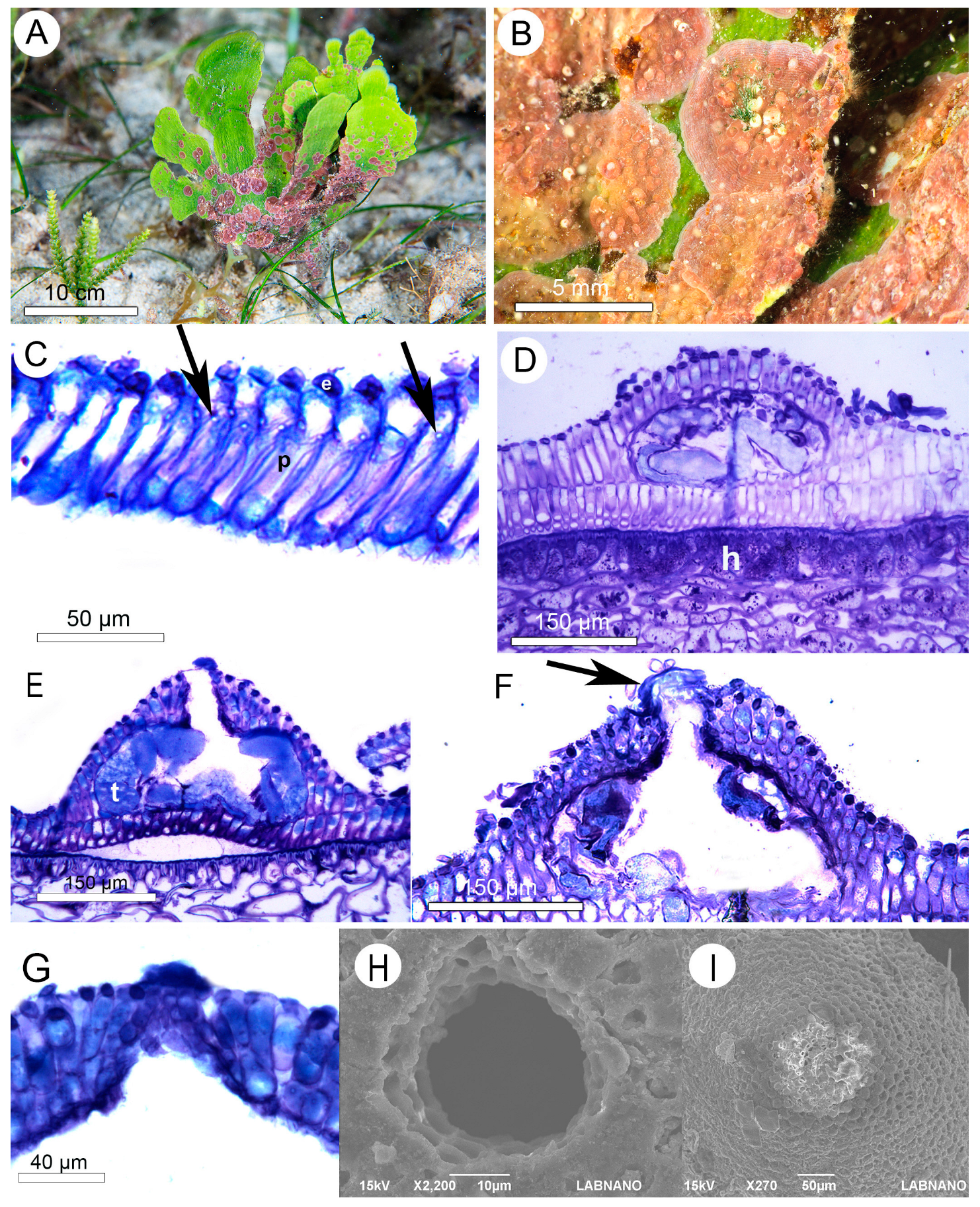
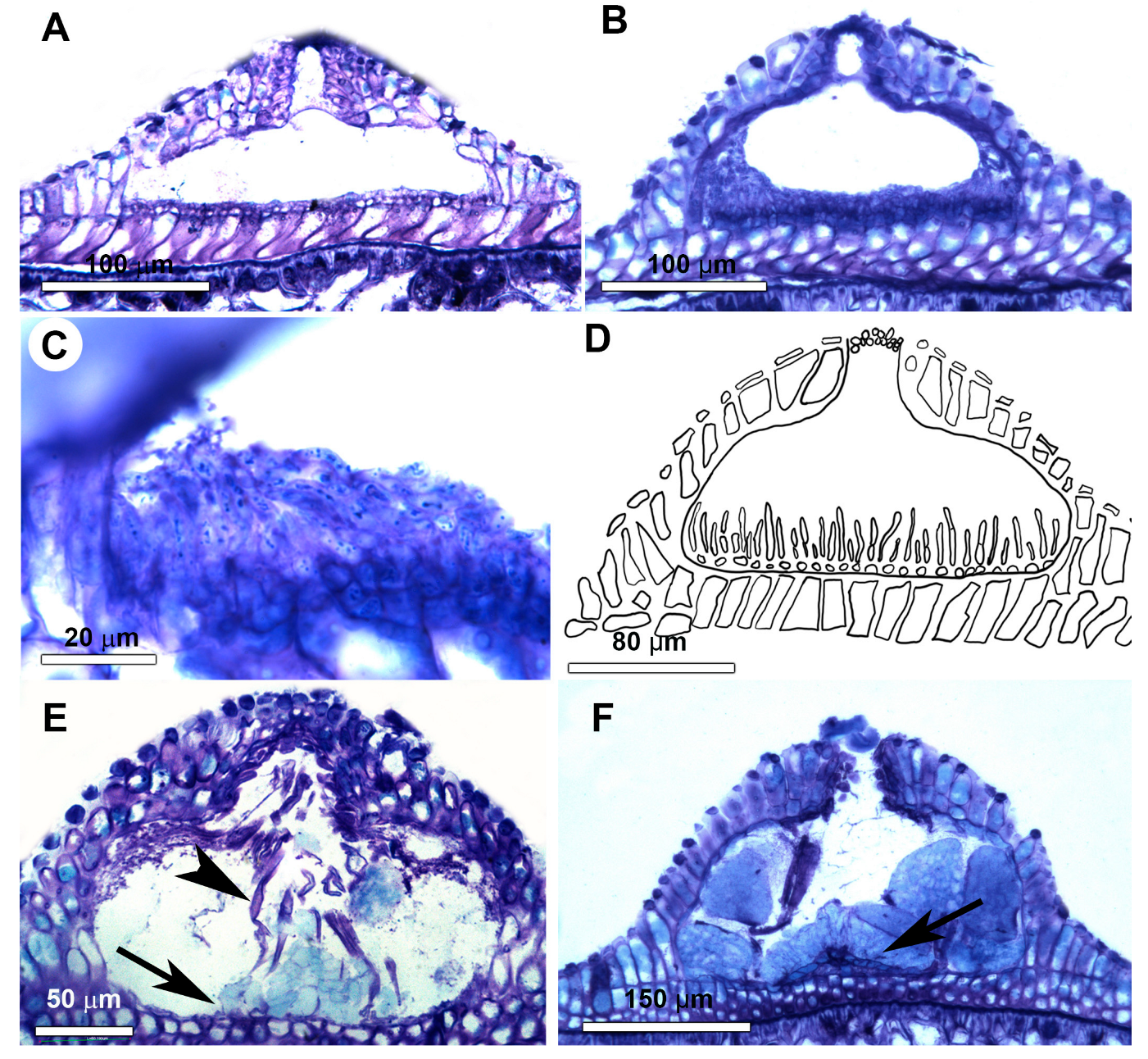
3.1. Morfoanatomical Results
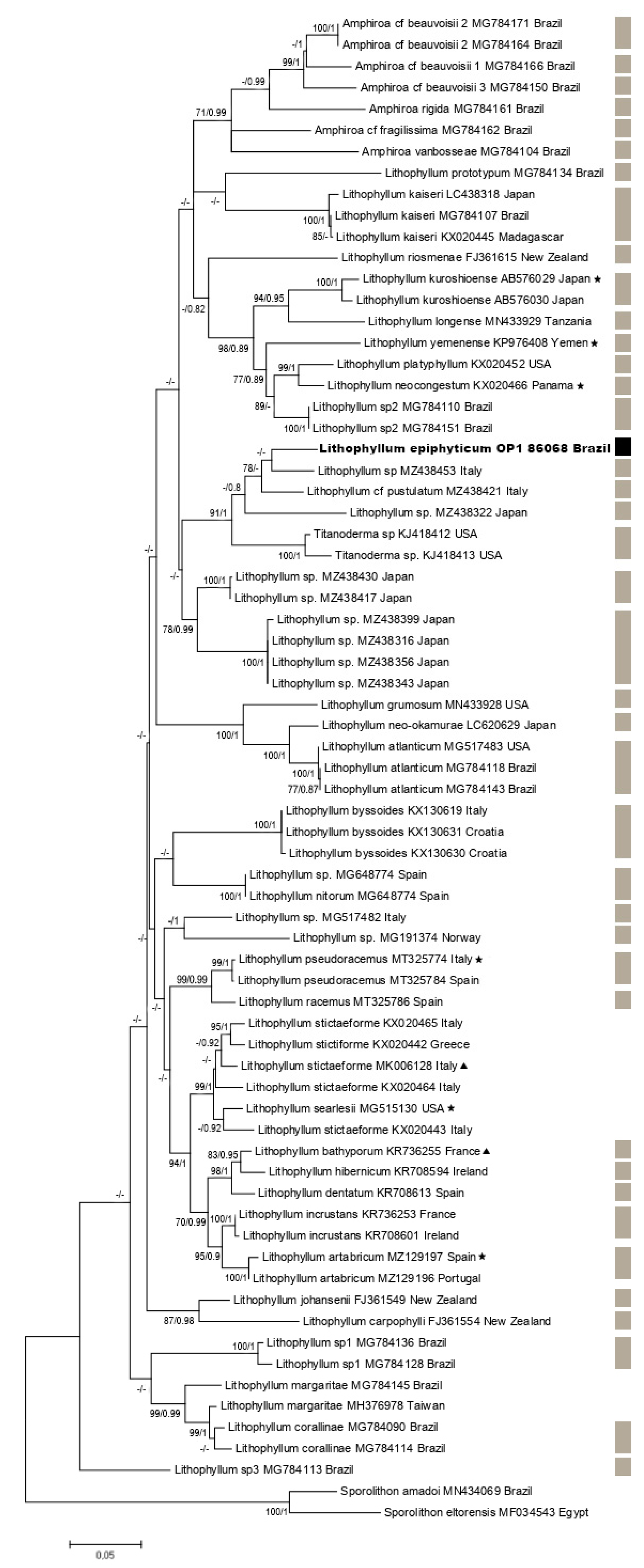
3.2. Molecular Results
4. Discussion
4.1. Ecological and Taxonomical Aspects
4.2. Biogeographic Aspects
4.3. Perspectives Biogeography x Taxonomy
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rosler, A.; Perfectti, F.; Peña, V.; Braga, J.C. Phylogenetic relationships of Corallinaceae (Corallinales, Rhodophyta): Taxonomic implications for reef-building corallines. J. Phycol. 2016, 52, 412–431. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.L.; Sauvage, T.; Schmidt, W.E.; Frederiq, S.; Hughey, J.R.; Gabrielson, P.W. The coralline genera Sporolithon and Heydrichia clarified by sequencing type material of their generitypes and other species. J. Phycol. 2017, 53, 1044–1059. [Google Scholar] [CrossRef]
- Torrano-Silva, B.N.; Vieira, B.R.; Riosmena-Rodríguez, R.; Oliveira, M.C. Guidelines for DNA barcoding of coralline algae, focusing on Lithophylloideae (Corallinales) from Brazil. Bot. Mar. 2018, 61, 127–140. [Google Scholar] [CrossRef]
- Peña, V.; Torres, T.R.D.G. Lithophyllum artabricum V. Peña, sp. Nov.(Corallinales, Rhodophyta): A cryptic species in the Atlantic Iberian Peninsula hitherto assigned to Lithophyllum stictiforme (Areschoug) Hauck. Cryptogam. Algol. 2021, 42, 153–172. [Google Scholar] [CrossRef]
- Nelson, W.A. Calcified macroalgae—Critical to coastal ecosystems and vulnerable to change: A review. Mar. Freshw. Res. 2009, 60, 787–801. [Google Scholar] [CrossRef]
- Diaz-Pulido, G.; Anthony, K.R.N.; Kline, D.I.; Dove, S.; Hoegh-Guldberg, O. Interactions between ocean acidification and warming on the mortality and dissolution of coralline algae. J. Phycol. 2012, 48, 32–39. [Google Scholar] [CrossRef]
- Kamenos, N.A.; Burdett, H.L.; Aloisio, E.; Findlay, H.S.; Martin, S.; Longbone, C.; Dunn, J.; Widdicombe, S.; Calosi, P. Coralline algal structure is more sensitive to rate, rather than the magnitude, of ocean acidification. Glob. Chang. Biol. 2013, 19, 3621–3628. [Google Scholar] [CrossRef] [PubMed]
- Noisette, F.; Duong, G.; Six, C.; Davoult, D.; Martin, S. Effects of elevated pCO2 on the metabolism of a temperate rhodolith Lithothamnion corallioides grown under different temperatures. J. Phycol. 2013, 49, 746–757. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.; Hall-Spencer, J.M. Effects of Ocean Warming and Acidification on Rhodolith/Maërl Beds. In Rhodolith/Maerl Beds: A Global Perspective; Riosmena Rodríguez, R., Nelson, W., Aguirre, J., Eds.; Springer International Publishing: Gewerbestrasse, Austria, 2017; Volume 15, pp. 55–85. [Google Scholar] [CrossRef]
- Peña, V.; Harvey, B.P.; Agostini, S.; Porzio, L.; Milazzo, M.; Horta, P.; Le Gall, L.; Hall-Spencer, J.M. Major loss of coralline algal diversity in response to ocean acidification. Glob. Chang. Biol. 2021, 27, 4785–4798. [Google Scholar] [CrossRef]
- McCoy, S.J.; Kamenos, N.A. Coralline algae (Rhodophyta) in a changing world: Integrating ecological, physiological, and geochemical responses to global change. J. Phycol. 2015, 51, 6–24. [Google Scholar] [CrossRef]
- Coletti, G.; Basso, D.; Corselli, C. Coralline algae as depth indicators in the Sommières Basin (early Miocene, Southern France). Geobios 2018, 51, 15–30. [Google Scholar] [CrossRef]
- Žuljević1, A.; Kaleb, S.; Peña, V.; Despalatović, M.; Cvitković, I.; De Clerck, O.; Le Gall, L.; Falace, A.; Vita, F.; Braga, J.C.; et al. First freshwater coralline alga and the role of local features in a major biome transition. Sci. Rep. 2016, 6, 19642. [Google Scholar] [CrossRef]
- Harvey, A.S.; Woelkerling, W.J.; Farr, T.J.; Neill, K.F.; Nelson, W.A. Coralline algae of central New Zealand: An identification guide to common ‘crustose’ species. NIWA Inf. Ser. 2005, 57, 1–145. [Google Scholar]
- Brasileiro, P.S.; Pereira-Filho, G.H.; Bahia, R.G.; Abrantes, D.P.; Guimarães, S.M.P.B.; Moura, R.L.; Francini-Filho, R.B.; Bastos, A.C.; Amado-Filho, G.M. Macroalgal composition and community structure of the largest rhodolith beds in the world. Mar. Biodivers. 2015, 46, 407–420. [Google Scholar] [CrossRef]
- Amado Filho, G.M.; Bahia, R.G.; Pereira Filho, G.H.; Longo, L.L. South Atlantic rhodolith beds: Latitudinal distribution, species composition, structure and ecosystem functions, threats and conservation status. In Rhodolith/Maerl Beds: A Global Perspective; Riosmena Rodríguez, R., Nelson, W., Aguirre, J., Eds.; Springer International Publishing: Gewerbestrasse, Austria, 2017; Volume 15, pp. 299–317. [Google Scholar] [CrossRef]
- Jesionek, M.B.; Bahia, R.G.; Hernández-Kantún, J.J.; Adey, W.H.; Yoneshigue-Valentin, Y.; Longo, L.L.; Amado-Filho, G.M. A taxonomic account of non-geniculate coralline algae (Corallinophycidae, Rhodophyta) from shallow reefs of the Abrolhos Bank, Brazil. Algae 2016, 31, 317–340. [Google Scholar] [CrossRef]
- Moura, R.L.; Secchin, N.A.; Amado-Filho, G.M.; Francini-Filho, R.B.; Freitas, M.O.; Minte-Vera, C.V.; Teixeira, J.B.; Thompson, F.L.; Dutra, G.F.; Sumida, P.Y.G.; et al. Spatial patterns of benthic megahabits and conservation planning in the Abrolhos Bank. Cont. Shelf Res. 2013, 70, 109–117. [Google Scholar] [CrossRef]
- Harvey, A.S.; Harvey, R.M.; Merton, E. The distribution, significance and vulnerability of Australian rhodolith beds: A review. Mar. Freshw. Res. 2016, 68, 411–428. [Google Scholar] [CrossRef]
- Amado-Filho, G.M.; Bahia, R.G.; Mariath, R.; Jesionek, M.B.; Moura, R.L.; Bastos, A.C.; Pereira-Filho, G.H.; Francini-Filho, R.B. Spatial and temporal dynamics of the abundance of crustose calcareous algae on the southernmost coral reefs of the western Atlantic (Abrolhos Bank, Brazil). Algae 2018, 33, 85–99. [Google Scholar] [CrossRef]
- Leão, L.A.S.; Bahia, R.G.; Jesionek, M.B.; Adey, W.H.; Johnson, G.; Salgado, L.T.; Pereira, R.C. Sporolithon franciscanum sp. nov. (Sporolithales, Rhodophyta), a New Rhodolith-Forming Species from Northeast Brazil. Diversity 2020, 12, 199. [Google Scholar] [CrossRef]
- Tâmega, F.T.; Riosmena-Rodriguez, R.; Mariath, R.; Figueiredo, M. Nongeniculate coralline red algae (Rhodophyta: Corallinales) in coral reefs from Northeastern Brazil and a description of Neogoniolithon atlanticum sp. nov. Phytotaxa 2014, 190, 277–298. [Google Scholar] [CrossRef]
- Figueiredo, M.D.O.; Steneck, R.S. Floristic and ecological studies of crustose coralline algae on Brazil’s Abrolhos reefs. In Proceedings of the 9th International Coral Reef Symposium 2002, Bali, Indonesia, 23–27 October 2000; Volume 1, pp. 493–498. [Google Scholar]
- Paula, A.F.; Figueiredo, M.A.O.; Creed, J.C. Structure of the macroalgal community associated with the seagrass Halodule wrightii Ascherson in the Abrolhos Marine National Park, Brazil. Bot. Mar. 2003, 46, 413–424. [Google Scholar] [CrossRef]
- Maneveldt, G.W.; Van der Merwe, E. Heydrichia cerasina sp. nov. (Sporolithales, Corallinophycidae, Rhodophyta) from the southernmost tip of Africa. Phycologia 2012, 51, 11–21. [Google Scholar] [CrossRef]
- Adey, W.H.; Adey, P.J. Studies on the biosystematics and ecology of the epilithic crustose Corallinaceae of the British Isles. Br. Phycol. J. 1973, 8, 343–407. [Google Scholar] [CrossRef]
- Chamberlain, Y.M. The genus Leptophytum (Rhodophyta, Corallinales) in the British Isles with descriptions of Leptophytum bornetii, L. elatum sp. nov., and L. laevae. Br. Phycol. J. 1990, 25, 179–199. [Google Scholar] [CrossRef]
- Woelkerling, W.J.; Irvine, L.M.; Harvey, A.S. Growthforms in non-geniculate coralline red algae (Corallinales, Rhodophyta). Aust. Syst. Bot. 1993, 6, 277–293. [Google Scholar] [CrossRef]
- Bahia, R.G.; Abrantes, D.P.; Brasileiro, P.S.; Pereira-Filho, G.H.; Amado-Filho, G.M. Rhodolith bed structure along a depth gradient on the northern coast of Bahia State, Brazil. Braz. J. Oceanogr. 2010, 58, 323–337. [Google Scholar] [CrossRef]
- Jesionek, M.B.; Bahia, R.G.; Lyra, M.B.; Leão, L.A.; Oliveira, M.C.; Amado-Filho, G.M. Newly discovered coralline algae in Southeast Brazil: Tectolithon fluminense gen. et sp. nov. and Crustaphytum atlanticum sp. nov.(Hapalidiales, Rhodophyta). Phycologia 2020, 59, 101–115. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. In Nucleic Acids Symposium Series; Oxford University Press: Oxford, UK, 1999; Volume 41, pp. 95–98. [Google Scholar]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Felsenstein, J. Phylogenies and the Comparative Method. Am. Nat. 1985, 125, 1–15. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. Mrbayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Barrón, C.; Duarte, C.M.; Frankignoulle, M.; Borges, A.V. Organic carbon metabolism and carbonate dynamics in a Mediterranean seagrass (Posidonia oceanica), meadow. Estuaries Coasts 2006, 29, 417–426. [Google Scholar] [CrossRef]
- Walker, D.I.; Woelkerling, W.J. Quantative study of sediment contribution by epiphytic coralline red algae in seagrass meadows in Shark Bay, Western Australia. Mar. Ecol. Prog. Ser. 1988, 43, 71–77. [Google Scholar] [CrossRef]
- James, P.J.; Bone, Y. Neritic Carbonate Sediments in a Temperate Realm: Southern Australia; Springer: London, UK, 2011; 254p, ISBN 978-90-4811-9288-5. [Google Scholar]
- Morcom, N.F.; Woelkerling, W.J. A critical interpretation of coralline-coralline (Corallinales, Rhodophyta) and coralline-other plant interactions. Cryptogam. Algol. 2000, 21, 1–31. [Google Scholar] [CrossRef]
- Martin, S.; Rodolfo-Metalpa, R.; Ransome, E.; Rowley, S.; Buia, M.C.; Gattuso, J.P.; Hall-Spencer, J. Effects of naturally acidified seawater on seagrass calcareous epibionts. Biol. Lett. 2008, 4, 689–692. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, R.T.; Rocha, G.M.; Karez, C.S.; Bahia, R.G.; Pereira, R.C.; Bastos, A.C.; Salgado, L.T. Global assessment of coralline algae mineralogy points to high vulnerability of Southwestern Atlantic reefs and rhodolith beds to ocean acidification. Sci. Rep. 2022, 12, 9589. [Google Scholar] [CrossRef]
- Taylor, W.R. Marine Algae of the Eastern Tropical and Subtropical Coasts of the Americas; University of Michigan Press: Ann Arbor, MI, USA, 1960; 870p. [Google Scholar]
- Jones, P.L.; Woelkerling, W.J. An analysis of trichocyte and spore germination attributes as taxonomic characters in the Pneophyllum–Fosliella complex (Corallinaceae, Rhodophyta). Phycologia 1984, 23, 183–194. [Google Scholar] [CrossRef]
- Chamberlain, Y.M. Trichocyte occurrence and phenology in four species of Pneophyllum (Rhodophyta, Corallinaceae) from the British Isles. Br. Phycol. J. 1985, 20, 375–379. [Google Scholar] [CrossRef]
- Harvey, A.; Phillips, L.E.; Woelkerling, W.J.; Millar, A.J.K. The Corallinaceae, subfamily Mastophoroideae (Corallinales, Rhodophyta) in south-eastern Australia. Aust. Syst. Bot. 2006, 19, 387–429. [Google Scholar] [CrossRef]
- Bahia, R.G. Algas Coralináceas Formadoras de Rodolitos da Plataforma Continental Tropical e Ilhas Oceânicas do Brasil: Levantamento Florístico e Taxonomia. Ph.D. Thesis, Escola Nacional de Botânica Tropical, Rio de Janeiro, Brazil, 2014; p. 221. [Google Scholar]
- Pezzolesi, L.; Falace, A.; Kaleb, S.; Hernandez-Kantun, J.J.; Cerrano, C.; Rindi, F. Genetic and morphological variation in an ecosystem engineer, Lithophyllum byssoides (Corallinales, Rhodophyta). J. Phycol. 2017, 53, 146–160. [Google Scholar] [CrossRef]
- Caragnano, A.; Rodondi, G.; Basso, D.; Peña, V.; Le Gall, L.; Rindi, F. Circumscription of Lithophyllum racemus (Corallinales, Rhodophyta) from the western Mediterranean Sea reveals the species Lithophyllum pseudoracemus sp. nov. Phycologia 2020, 59, 584–597. [Google Scholar] [CrossRef]
- Woelkerling, W.J.; Campbell, S.J. An account of southern Australian species of Lithophyllum (Corallinaceae, Rhodophyta). Bull. Br. Mus. (Nat. Hist.) Bot. 1992, 22, 1–107. [Google Scholar]
- Woelkerling, W.J. Subfamily Lithophylloideae. In The Marine Benthic Flora of Southern Australia, Part IIIB.; Gracilariales, Rhodymeniales, Corallinales and Bonnemaisoniales; Womersley, H.B.S.W., Ed.; Australian Biological Resources Study: Canberra, Australia, 1996; pp. 214–237. [Google Scholar] [CrossRef]
- Harvey, A.; Woelkerling, W.J.; Millar, A.J.K. The genus Lithophyllum (Lithophylloideae, Corallinaceae, Rhodophyta) in south-eastern Australia, with the description of L. riosmenae, sp. nov. Aust. Syst. Bot. 2009, 22, 296–317. [Google Scholar] [CrossRef]
- Crespo, T.M.; Bahia, R.G.; Maneveldt, G.W.; Amado-Filho, G.M. Floristic composition of crustose coralline algae from the St. Peter and St. Paul Archipelago, a summit of the Mid-Atlantic Ridge. Phytotaxa 2014, 190, 17. [Google Scholar] [CrossRef]
- Athanasiadis, A. North Aegean marine algae. III. Structure and development of the encrusting coralline Titanoderma cystoseirae (Rhodophyta, Lithophylloideae). Nord. J. Bot. 1989, 9, 435–441. [Google Scholar] [CrossRef]
- Irvine, L.M.; Chamberlain, Y.M. Seaweeds of the British Isles. In Volume 1 Rhodophyta Part 2B Corallinales, Hildenbrandiales; HMSO: London, UK, 1994; pp. 1–276. [Google Scholar]
- Guiry, M.D.; Guiry, G.M.; AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway 2023. Available online: http://www.algaebase.org (accessed on 25 July 2023).
- Penrose, D.; Chamberlain, Y.M. Hydrolithon farinosum (Lamouroux) comb. nov.: Implications for generic concepts in the Mastophoroideae (Corallinaceae, Rhodophyta). Phycologia 1993, 32, 295–303. [Google Scholar] [CrossRef]
- Highsmith, R.C. Floating and algal rafting as potential dispersal mechanisms in brooding invertebrates. Mar. Ecol. Prog. Ser. 1985, 25, 169–179. [Google Scholar] [CrossRef]
- Hansen, G.I.; Hanyuda, T.; Kawai, H. Invasion threat of benthic marine algae arriving on Japanese tsunami marine debris in Oregon and Washington, USA. Phycologia 2018, 57, 641–658. [Google Scholar] [CrossRef]
- Nyberg, C.D.; Wallentinus, I. Can species traits be used to predict marine macroalgal introductions? Biol. Invasions 2005, 7, 265–279. [Google Scholar] [CrossRef]
- Santelices, B. Patterns of reproduction, dispersal and recruitment in seaweeds. Oceanogr. Mar. Biol. Annu. Rev. 1990, 28, 177–276. [Google Scholar]
- Maneveldt, G.W.; Van der Merwe, E.; Keats, D.W. Updated keys to the non-geniculate coralline red algae (Corallinophycidae, Rhodophyta) of South Africa. South Afr. J. Bot. 2016, 106, 158–164. [Google Scholar] [CrossRef]
- Sissini, M.N.; Oliveira, M.C.; Gabrielson, P.W.; Robinson, N.M.; Okolodkov, Y.B.; Riosmena-Rodríguez, R.; Horta, P.A. Mesophyllum erubescens (Corallinales, Rhodophyta)—So many species in one epithet. Phytotaxa 2014, 190, 299–319. [Google Scholar] [CrossRef]
- Hernandez-Kantun, J.J.; Gabrielson, P.; Hughey, J.R.; Pezzolesi, L.; Rindi, F.; Robinson, N.M.; Peña, V.; Riosmena-Rodriguez, R.; Le Gall, L.; Adey, W. Reassessment of branched Lithophyllum spp. (Corallinales, Rhodophyta) in the Caribbean Sea with global implications. Phycologia 2016, 55, 619–639. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| L. epiphytica 1 | L. irvineanum 2 | L. chamberlainianum (Type Collection) 2 | L. prototypum (Type Collection) 2 | L. pustulatum 2,3,4 | L. cystoseirae 5 | L. laminariae 6 | |
|---|---|---|---|---|---|---|---|
| Distribution 7 | Abrolhos Archipelago (Brazil) | South and Western Australia | South Pacific | Caribbean, Western Atlantic, Arabian Gulf, Indo-Pacific | Mediterranean, Indo-Pacific, Eastern and Western Atlantic, Red Sea, Arabian Gulf, Caribbean, Arctic Ocean | Adriatic Sea, Mediterranean, Eastern Atlantic | Eastern Atlantic |
| Habit | Epiphytic | Epiphytic | Epiphytic | Epilithic/epiphytic/free-living as rhodolith | Epilithic/free-living as rhodolith | Epiphytic | Epiphytic |
| Growth form | Encrusting | Encrusting | Encrusting | Encrusting | Encrusting/warty to lumpy | Encrusting | Encrusting |
| Thalli structured in several applanate branches with swirled margins in surface view | Absent | Present | Absent | Present | Absent | Absent | Absent |
| Basal layer of palisade or sinuate cells | Present | Present | Present | Present | Present | Present | Present |
| Mature thallus thickness (number of cell layers) | <10 | <10 | <10 | <10 | >10 | >10 | >10 |
| Trichocytes | Absent | Absent | Absent | Absent | Absent | Present | Absent |
| Tetra/bisporangial conceptacle chamber diameter (µm) | 240–300 | 180–263 | 145–252 | 330–415 | 420–465/328–556 | 288–475/307–324 | 312–364 |
| Tetra/bisporangial conceptacle chamber height (µm) | 100–140 | 82–125 | 60–135 | 100–190 | 300–320/110–180 | 80–172/80–124 | 104–156 |
| Pore canals of tetra/bisporangial conceptacles occluded by cells | Absent/present | Present | Present | Absent | Absent | Present | Absent |
| Number of cell layers in tetra/bisporangial roof filaments | 2–4 | 2–4 | 2–5 | 2–3 | 2–4 | 3–6 | ND |
| Position of tetra/bisporangial chamber floor below thallus surface (number of cells) | 1 | 1–3 | 1 | 1 | 1–5 | ND | 6-+ |
| Conceptacle position in the thallus | Superficial | Superficial/immersed | Superficial/immersed | Superficial/immersed | Superficial/immersed | Superficial/immersed | superficial/immersed |
| Columella | Present | Present | Present | Present | Present | Present | Present |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyra, M.B.; Bahia, R.G.; Jesionek, M.B.; Carvalho, R.T.; Moraes, F.C.; Harvey, A.S.; Pereira, R.C.; Salgueiro, F.; Salgado, L.T. Hydrolithon farinosum and Lithophyllum epiphyticum sp. nov. (Corallinaceae, Corallinales, Rhodophyta), Two Epiphytic Crustose Coralline Algae from the Abrolhos Archipelago, Brazil, Southwestern Atlantic. Diversity 2023, 15, 1013. https://doi.org/10.3390/d15091013
Lyra MB, Bahia RG, Jesionek MB, Carvalho RT, Moraes FC, Harvey AS, Pereira RC, Salgueiro F, Salgado LT. Hydrolithon farinosum and Lithophyllum epiphyticum sp. nov. (Corallinaceae, Corallinales, Rhodophyta), Two Epiphytic Crustose Coralline Algae from the Abrolhos Archipelago, Brazil, Southwestern Atlantic. Diversity. 2023; 15(9):1013. https://doi.org/10.3390/d15091013
Chicago/Turabian StyleLyra, Manoela B., Ricardo G. Bahia, Michel B. Jesionek, Rodrigo T. Carvalho, Fernando C. Moraes, Adele S. Harvey, Renato C. Pereira, Fabiano Salgueiro, and Leonardo T. Salgado. 2023. "Hydrolithon farinosum and Lithophyllum epiphyticum sp. nov. (Corallinaceae, Corallinales, Rhodophyta), Two Epiphytic Crustose Coralline Algae from the Abrolhos Archipelago, Brazil, Southwestern Atlantic" Diversity 15, no. 9: 1013. https://doi.org/10.3390/d15091013
APA StyleLyra, M. B., Bahia, R. G., Jesionek, M. B., Carvalho, R. T., Moraes, F. C., Harvey, A. S., Pereira, R. C., Salgueiro, F., & Salgado, L. T. (2023). Hydrolithon farinosum and Lithophyllum epiphyticum sp. nov. (Corallinaceae, Corallinales, Rhodophyta), Two Epiphytic Crustose Coralline Algae from the Abrolhos Archipelago, Brazil, Southwestern Atlantic. Diversity, 15(9), 1013. https://doi.org/10.3390/d15091013