
Abundance, Distribution, and Habitat Preference of Syngnathid Species in Sabaudia Lake (Tyrrhenian Sea)


 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
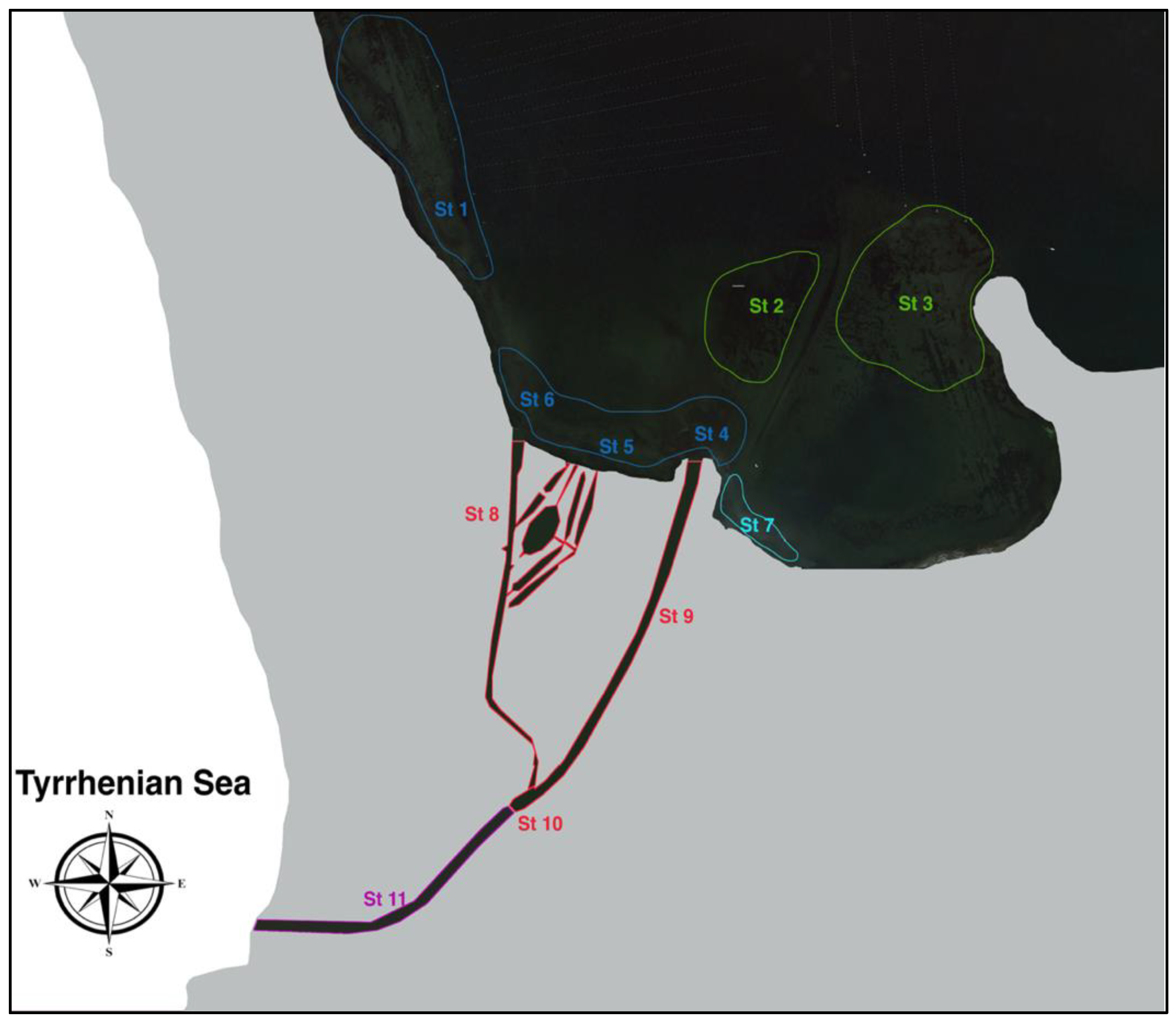
2.1. Sampling Stations and Data Collection
2.2. Data Analysis
3. Results
3.1. Habitat Characterisation
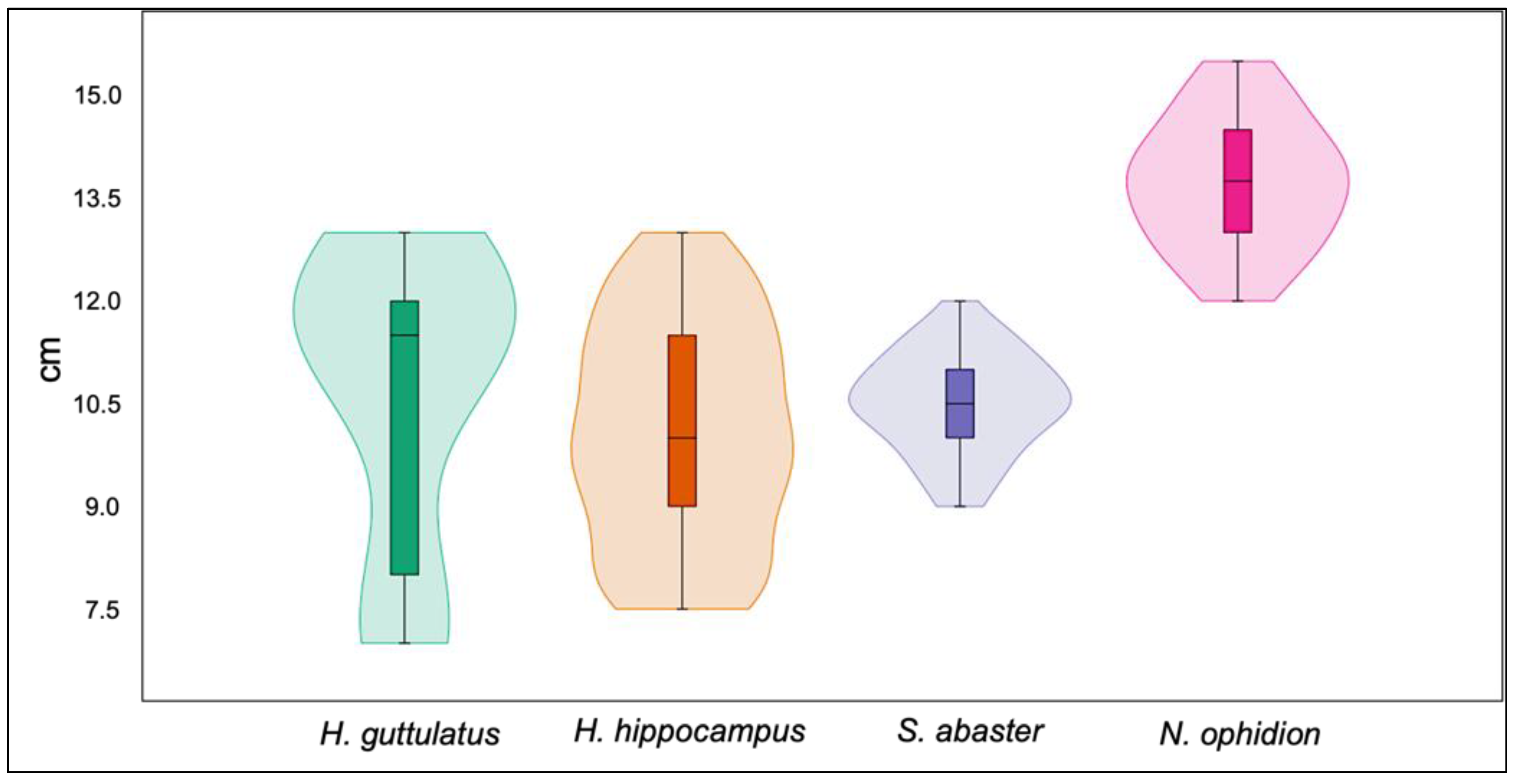
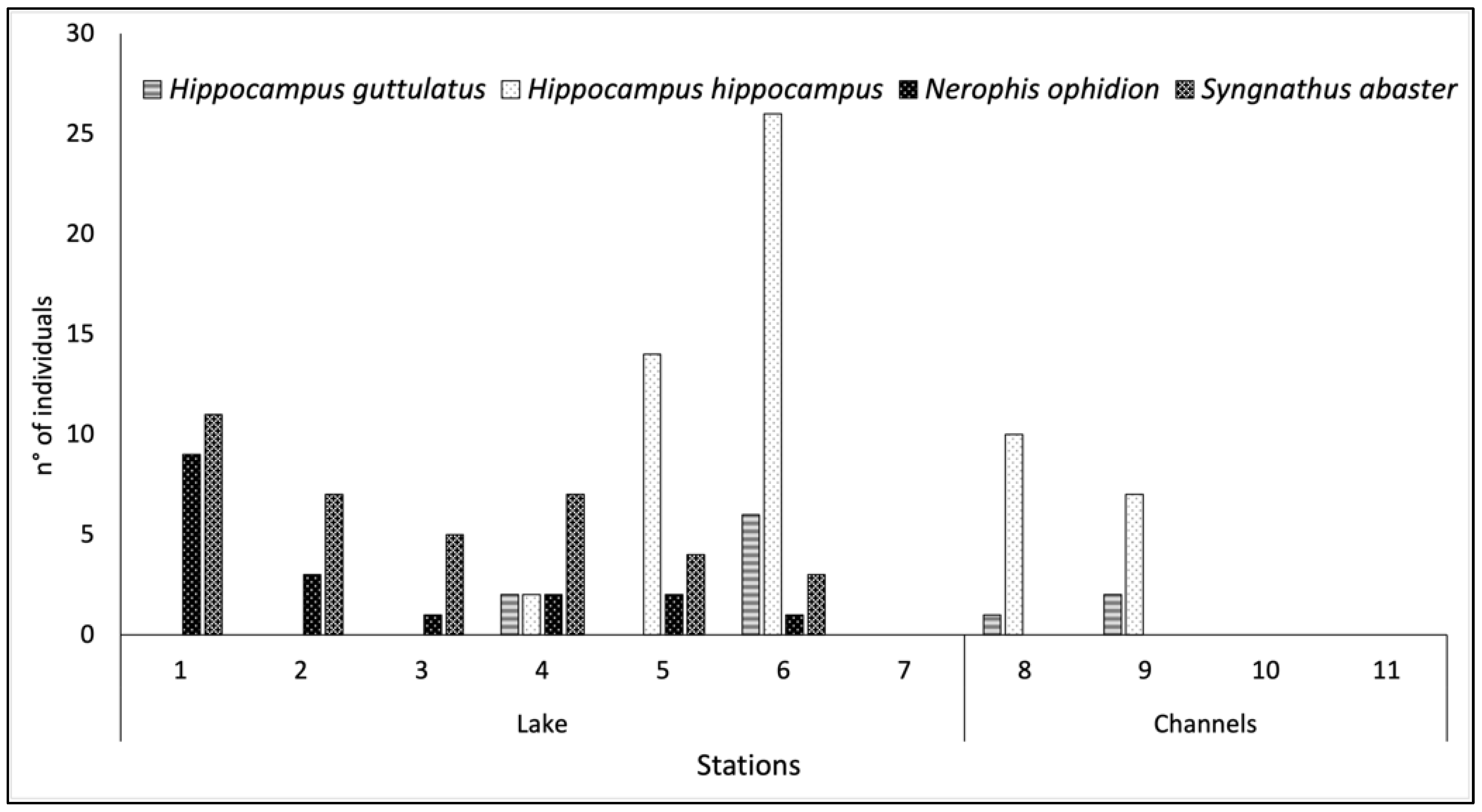
3.2. Syngnathid Population
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Kerr, J.T.; Currie, D.J. Effects of human activity on global extinction risk. Conserv. Biol. 1995, 9, 1528–1538. [Google Scholar] [CrossRef]
- Lotze, H.K.; Lenihan, H.S.; Bourque, B.J.; Bradbury, R.H.; Cooke, R.G.; Kay, M.C.; Kidwell, S.M.; Kirby, M.X.; Peterson, C.H.; Jackson, J.B. Depletion, degradation, and recovery potential of estuaries and coastal seas. Science 2006, 312, 1806–1809. [Google Scholar] [CrossRef] [PubMed]
- Crain, C.M.; Kroeker, K.; Halpern, B.S. Interactive and cumulative effects of multiple human stressors in marine systems. Ecol. Let. 2008, 11, 1304–1315. [Google Scholar] [CrossRef] [PubMed]
- Crain, C.M.; Halpern, B.S.; Beck, M.W.; Kappel, C.V. Understanding and managing human threats to the coastal marine environment. Ann. NY Acad. Sci. 2009, 1162, 39–62. [Google Scholar] [CrossRef] [PubMed]
- Pierri, C.; Lazic, T.; Gristina, M.; Corriero, G.; Sinopoli, M. Large-scale distribution of the European seahorses (Hippocampus Rafinesque, 1810): A systematic review. Biology 2022, 11, 325. [Google Scholar] [CrossRef] [PubMed]
- Lim, A.; Chee, O.; Chong, V.; Wong, C.; Choo, C.K. Diversity, habitats and onservation Threats of Syngnathid (Syngnathidae) Fishes in Malaysia. Trop. Zool. 2011, 24, 193–222. [Google Scholar]
- Vincent, A.C.J.; Foster, S.J.; Koldewey, H.J. Conservation and management of seahorses and other Syngnathidae. J. Fish Biol. 2011, 78, 1681–1724. [Google Scholar] [CrossRef] [PubMed]
- Claassens, L.; Hodgson, A.; Short, G.; Harasti, D. Diversity, distribution, ecology and conservation status of the family Syngnathidae in sub-Saharan Africa and adjacent islands. In Oceanography and Marine Biology: An Annual Review, 1st ed.; CRC Press: Boca Raton, FL, USA, 2022; Volume 60, pp. 169–242. ISBN 978-1-00-328860-2. [Google Scholar]
- Olden, J.D.; Hogan, Z.S.; Zanden, M.J.V. Small fish, big fish, red fish, blue fish: Size-biased extinction risk of the world’s freshwater and marine fishes. Global Ecol. Biogeogr. 2007, 16, 694–701. [Google Scholar] [CrossRef]
- Vincent, A.C.J. The International Trade in Seahorses; Species in Danger; Traffic International: Cambridge, UK, 1996; ISBN 9781858500980. [Google Scholar]
- Salin, K.R.; Yohannan, T.M.; Mohanakumaran Nair, C. Fisheries and Trade of Seahorses, Hippocampus spp., in Southern India. Fish. Manag. 2005, 12, 269–273. [Google Scholar] [CrossRef]
- Correia, M.; Paulo, D.; Samara, E.; Koulouri, P.; Mentogiannis, V.; Dounas, C. Field studies of seahorse population density, structure and habitat use in a semi-closed north-eastern Mediterranean marine area (Stratoni, North Aegean Sea). J. Fish Biol. 2020, 97, 314–317. [Google Scholar] [CrossRef]
- Iucn, Red List of Threatened Species. 2022. Available online: http://www.iucnredlist.org/ (accessed on 15 March 2023).
- Curtis, J.M.R.; Vincent, A.C.J. Life history of an unusual marine fish: Survival, growth and movement patterns of Hippocampus guttulatus Cuvier 1829. J. Fish Biol. 2006, 68, 707–733. [Google Scholar] [CrossRef]
- Correia, M.; Caldwell, I.R.; Koldewey, H.J.; Andrade, J.P.; Palma, J. Seahorse (Hippocampinae) population fluctuations in the ria Formosa lagoon, south Portugal. J. Fish Biol. 2015, 87, 679–690. [Google Scholar] [CrossRef]
- Lazic, T.; Pierri, C.; Gristina, M.; Carlucci, R.; Cardone, F.; Colangelo, P.; Desiderato, A.; Mercurio, M.; Bertrandino, M.S.; Longo, C.; et al. Distribution and habitat preferences of Hippocampus Species along the Apulian Coast. Aquat. Conserv. 2018, 28, 1317–1328. [Google Scholar] [CrossRef]
- Pierri, C.; Lazic, T.; Corriero, G.; Cardone, F.; Onen Tarantini, S.; Desiderato, A.; Mirto, S.; Gristina, M. Site fidelity of Hippocampus Guttulatus Cuvier, 1829 at Mar Piccolo of Taranto (Southern Italy; Ionian Sea). Environ. Biol. Fish. 2020, 103, 1105–1118. [Google Scholar] [CrossRef]
- Campolmi, M.; Franzoi, P.; Mazzola, A. Observations on pipefish (Syngnathidae) biology in the Stagnone Lagoon (West Sicily). Publicaciones Espec. Inst. Esp. Oceanogr. 1996, 21, 205–209. [Google Scholar]
- Franzoi, P.; Maccagnani, R.; Rossi, R.; Ceccherelli, V. Life-Cycles and Feeding-Habits of Syngnathus taenionotus and S. Abaster (Pisces, Syngnathidae) in a Brackish Bay of the Po River Delta (Adriatic Sea). Mar. Ecol. Prog. Ser. 1993, 97, 71–81. [Google Scholar] [CrossRef]
- Sanna, D.; Biagi, F.; Ben Alaya, H.; Maltagliati, F.; Addis, A.; Romero, A.; De Juan, J.; Jean-pierre, Q.; Castelli, A.; Franzoi, P.; et al. Mitochondrial DNA Variability of the Pipefish Syngnathus Abaster. J. fish Biol. 2013, 82, 856–876. [Google Scholar] [CrossRef]
- Scapin, L.; Cavraro, F.; Malavasi, S.; Riccato, F.; Zucchetta, M.; Franzoi, P. Linking Pipefishes and Seahorses to Seagrass Meadows in the Venice Lagoon: Implications for Conservation. Aquat. Conserv. 2018, 28, 282–295. [Google Scholar] [CrossRef]
- Gürkan, Ş. Investigations on the Ecomorphologic Characteristics of the Pipefish (Familia: Syngnathidae) Distributing in the Çamalti Lagoon (İzmir Bay). Ph.D. Thesis, Ege University, Bornova, Turkey, 2004. Volume 215. [Google Scholar]
- Gürkan, S.; Taskavak, E. Length-Weigth Relationships for Syngnathid Fishes of the Aegean Sea, Turkey. Belg. J. Zool. 2007, 137, 227–242. [Google Scholar]
- Amor, M.M.; Salem, M.; Reynaud, C.; Capapé, C. Length–Weight Relationships in Syngnathid Species from Tunisian Waters (Central Mediterranean). Mar. Biodivers. Rec. 2011, 4, e44. [Google Scholar] [CrossRef]
- Planas, M.; Piñeiro-Corbeira, C.; Bouza, C.; Castejon, I.; Vera, M.; Regueira, M.; Ochoa, V.; Bárbara, I.; Terrados, J.; Chamorro Valverde, A.; et al. A Multidisciplinary Approach to Identify Priority Areas for the Monitoring of a Vulnerable Family of Fishes in Spanish Marine National Parks. BMC Evol. Biol. 2021, 21, 77–92. [Google Scholar] [CrossRef] [PubMed]
- Franzoi, P.; Franco, A.; Torricelli, P. Fish Assemblage Diversity and Dynamics in the Venice Lagoon. Rend. Fis. Acc. Lincei 2010, 21, 269–281. [Google Scholar] [CrossRef]
- Malavasi, S.; Franco, A.; Riccato, F.; Valerio, C.; Torricelli, P.; Franzoi, P. Habitat Selection and Spatial Segregation in Three Pipefish Species. Estuar. Coast. Shelf S. 2007, 75, 143–150. [Google Scholar] [CrossRef]
- Hare, M.P.; Nunney, L.; Schwartz, M.K.; Ruzzante, D.E.; Burford, M.; Waples, R.S.; Ruegg, K.; Palstra, F. Understanding and Estimating Effective Population Size for Practical Application in Marine Species Management: Applying Effective Population Size Estimates to Marine Species Management. Conserv. Biol. 2011, 25, 438–449. [Google Scholar] [CrossRef] [PubMed]
- Macali, A.; Conde, A.; Smriglio, C.; Mariottini, P.; Crocetta, F. The Evolution of the Molluscan Biota of Sabaudia Lake: A Matter of Human History. Sci. Mar. 2013, 77, 649–662. [Google Scholar] [CrossRef]
- Zucchetta, M.; Capoccioni, F.; Franzoi, P.; Ciccotti, E.; Leone, C. Fish Response to Multiple Anthropogenic Stressors in Mediterranean Coastal Lagoons: A Comparative Study of the Role of Different Management Strategies. Water 2021, 13, 130. [Google Scholar] [CrossRef]
- Millet, B.; Guelorget, O. Spatial and Seasonal Variability in the Relationships between Benthic Communities and Physical Environment in a Lagoon Ecosystem. Mar. Ecol. Prog. Ser. 1994, 108, 161–174. [Google Scholar] [CrossRef]
- Perdicaro, R. Osservazioni sulle condizioni idrologiche del Lago di Sabaudia o di Paola nel biennio 1982-1983. Quad. Ist.Idrobiol. Aquacolt. Brunelli 1983, 3, 59–72. [Google Scholar]
- Perdicaro, R. Condizioni chimico-fisiche del Lago di Sabaudia o di Paola dal 1933 al 1984. Quad. Ist. Idrobiol. Aquacolt.Brunelli 1984, 4, 71–139. [Google Scholar]
- Guélorget, O.; Perthuisot, J.P. Indicateurs Biologiques et Diagnose Écologique Dans Le Domaine Paralique. Bull. Ecol. 1984, 15, 67–76. [Google Scholar]
- Guélorget, O.; Perthuisot, J.P. Paralic Ecosystems Biological Organization and Functionning. Vie et Milieu 1992, 42, 215–251. [Google Scholar]
- Samoilys, M. Underwater Visual Census Surveys. Manual for Assessing Fish Stocks on Pacific Coral Reefs; Training Series QE97009; Queensland Department of Primary Industries: Brisbane, Australia, 1997; pp. 16–29. [Google Scholar]
- Gürkan, S.; Taylan, B. The first observations of oocyte diameters for the pipefish from the southern Black Sea. J. Black Sea/Mediterr. Environ. 2013, 19, 206–216. [Google Scholar]
- Curtis, J.; Moreau, M.-A.; Marsden, D.; Bell, E.; Martin-Smith, K.; Samoilys, M.; Vincent, A. Underwater Visual Census for Seahorse Population Assessments; Project Seahorse Technical Report No. 8; University of British Columbia: Vancouver, BC, Canada, 2004. [Google Scholar]
- Curtis, J.; Vincent, A. Distribution of Sympatric Seahorse Species along a Gradient of Habitat Complexity in a Seagrass-Dominated Community. Mar. Ecol. Prog. Ser. 2005, 291, 81–91. [Google Scholar] [CrossRef]
- Perera, N.; Dahanayaka, D.D.G.L.; Udagedara, S. Habitat Preference and Population Structure of Two Data Deficient Seahorse (Syngnathidae) Species. OUSL Jnl 2017, 12, 75. [Google Scholar] [CrossRef]
- Fagan, W.F. Connectivity, fragmentation, and extinction risk in dendritic metapopulations. Ecology 2002, 83, 3243–3249. [Google Scholar] [CrossRef]
- Hernández-Urcera, J.; Murillo, F.J.; Regueira, M.; Cabanellas-Reboredo, M.; Planas, M. Preferential Habitats Prediction in Syngnathids Using Species Distribution Models. Mar. Environ. Res. 2021, 172, 105488. [Google Scholar] [CrossRef] [PubMed]
- Rosa, I.L.; Oliveira, T.P.R.; Castro, A.L.C.; Moraes, L.E.D.S.; Xavier, J.H.A.; Nottingham, M.C.; Dias, T.L.P.; Bruto-Costa, L.V.; Araújo, M.E.; Birolo, A.B.; et al. Population Characteristics, Space Use and Habitat Associations of the Seahorse Hippocampus Reidi (Teleostei: Syngnathidae). Neotrop. Ichthyol. 2007, 5, 405–414. [Google Scholar] [CrossRef]
- Nester, G.M.; De Brauwer, M.; Koziol, A.; West, K.M.; DiBattista, J.D.; White, N.E.; Power, M.; Heydenrych, M.J.; Harvey, E.; Bunce, M. Development and Evaluation of Fish eDNA Metabarcoding Assays Facilitate the Detection of Cryptic Seahorse Taxa (Family: Syngnathidae). Environ. DNA 2020, 2, 614–626. [Google Scholar] [CrossRef]
- Vizzini, S.; Mazzola, A. The trophic structure of the pipefish community (Pisces: Syngnathidae) from a western Mediterranean seagrass meadow based on stable isotope analysis. Estuaries 2004, 27, 325–333. [Google Scholar] [CrossRef]
- Amor, M.M.B.; Salem, M.B.; Souissi, J.B.; Capape, C. On the occurrence of the deep-snouted pipefish Syngnathus typhle Linnaeus, 1758 (Osteichthyes: Syngnathidae) in Tunisian waters (Central Mediterranean). Acta Adriat. 2007, 48, 15–23. [Google Scholar]
- Ben Alaya, H.; Sanna, D.; Casu, M.; Biagi, F.; Francalacci, P.; Carcupino, M.; Galzin, R.; Quignard, J.P.; Trabelsi, M. Analysis of meristic and mitochondrial DNA variation in Syngnathus abaster (Teleostea: Syngnathidae) from two western Mediterranean lagoons. Biologia 2011, 66, 1140–1147. [Google Scholar] [CrossRef]
- Hablützel, P.I.; Wilson, A.B. Notes on the occurrence of Syngnathus rostellatus (Teleostei: Syngnathidae) in the Mediterranean. Mar. Biodiv. Rec. 2011, 4, E57. [Google Scholar] [CrossRef]
- Gristina, M.; Cardone, F.; Carlucci, R.; Castellano, L.; Passarelli, S.; Corriero, G. Abundance, distribution and habitat preference of Hippocampus guttulatus and Hippocampus hippocampus in a semi-enclosed central Mediterranean marine area. Mar. Ecol. 2015, 36, 57–66. [Google Scholar] [CrossRef]
- Moser, F.N.; Wilson, A.B. Reproductive Isolation Following Hybrid Speciation in Mediterranean Pipefish (Syngnathus Spp.). Anim. Behav. 2020, 161, 77–87. [Google Scholar] [CrossRef]
- Pierri, C.; Cardone, F.; Corriero, G.; Lazic, T.; Quattrocchi, F.; Alabiso, G.; Gristina, M. Density Decline in a Mediterranean Seahorse Population: Natural Fluctuations or New Emerging Threats? Front. Mar. Sci. 2021, 8, 692068. [Google Scholar] [CrossRef]
- Simal Rodríguez, A.; Grau, A.; Castro-Fernández, J.; Castejón, I.; Terrados, J.; Morales-Nin, B.; Arechavala-Lopez, P. Reproductive Biology of Pipefish Syngnathus typhle and S. abaster (Syngnathidae) from Western Mediterranean Sea. J. Ichthyol. 2021, 61, 608–615. [Google Scholar] [CrossRef]
- Martin-Smith, K.M.; Vincent, A.C.J. Exploitation and Trade of Australian Seahorses, Pipehorses, Sea Dragons and Pipefishes (Family Syngnathidae). Oryx 2006, 40, 141–151. [Google Scholar] [CrossRef]
- Rosa, I.; Sampaio, C. Collaborative Monitoring of the Ornamental Trade of Seahorses and Pipefishes (Teleostei: Syngnathidae) in Brazil: Bahia State as a Case Study. Neotrop. Ichthyol. 2006, 4, 247–252. [Google Scholar] [CrossRef]
- Laksanawimol, P.; Petpiroon, S.; Damrongphol, P. Trade of Seahorses, Hippocampus Spp. (Actinopterygii: Syngnathiformes: Syngnathidae), on the East Coast of the Gulf of Thailand. Acta Icth. Et Piscat. 2013, 43, 229–235. [Google Scholar] [CrossRef]
- Harasti, D. Declining Seahorse Populations Linked to Loss of Essential Marine Habitats. Mar. Ecol. Prog. Ser. 2016, 546, 173–181. [Google Scholar] [CrossRef]
- Masonjones, H.D.; Rose, E.; McRae, L.B.; Dixson, D.L. An Examination of the Population Dynamics of Syngnathid Fishes within Tampa Bay, Florida, USA. Curr. Zool. 2010, 56, 118–133. [Google Scholar] [CrossRef]
- Claassens, L.; Booth, A.J.; Hodgson, A.N. An Endangered Seahorse Selectively Chooses an Artificial Structure. Environ. Biol. Fish 2018, 101, 723–733. [Google Scholar] [CrossRef]
- Salin, K.R.; Yohannan, T.; Nair, C. Captive Rearing of Hippocampus kuda and Issues of Seahorse Conservation in India. In Proceedings of the International Conference & Exposition on Marine Living Resources of India for Food and Medicine, Chennai, India, 27–29 February 2004. [Google Scholar]
- Dody, S.; Manuputty, G.; Limmon, G. Habitat Characteristcs and Density of Spotted Seahorse Hippocampus kuda at Waters of Ternate Island and Surroundings, Maluku Utara, Indonesia. IOP Conf. Ser. Earth Environ. Sci. 2021, 805, 012006. [Google Scholar] [CrossRef]
- Power, M.; Attrill, M.J. Long-Term Trends in the Estuarine Abundance of Nilsson’s Pipefish (Syngnathus rostellatus Nilsson). Estuar. Coast. Shelf S. 2003, 57, 325–333. [Google Scholar] [CrossRef]
- Kuiter, R.H. Seahorses and Their Relatives; Aquatic Photographics: Seaford, UK, 2009; ISBN 9780977537211. [Google Scholar]
- Berglund, A.; Sundin, J.; Rosenqvist, G. Baltic Pipefish Females Need Twice as Many Males as They Get. Behav. Ecol. 2017, 28, 827–832. [Google Scholar] [CrossRef]
- Whitfield, A.K.; Mkare, T.K.; Teske, P.R.; James, N.C.; Cowley, P.D. Life-Histories Explain the Conservation Status of Two Estuary-Associated Pipefishes. Biol. Conserv. 2017, 212, 256–264. [Google Scholar] [CrossRef]
- Castejón-Silvo, I.; Terrados, J.; Nguyen, T.; Jutfelt, F.; Infantes, E. Increased Energy Expenditure Is an Indirect Effect of Habitat Structural Complexity Loss. Funct. Ecol. 2021, 35, 2316–2328. [Google Scholar] [CrossRef]
- Pandolfi, J.M.; Bradbury, R.H.; Sala, E.; Hughes, T.P.; Bjorndal, K.A.; Cooke, R.G.; McArdle, D.; McClenachan, L.; Newman, M.J.H.; Paredes, G.; et al. Global Trajectories of the Long-Term Decline of Coral Reef Ecosystems. Science 2003, 301, 955–958. [Google Scholar] [CrossRef]
- Aylesworth, L.A.; Xavier, J.H.; Oliveira, T.P.R.; Tenorio, G.D.; Diniz, A.F.; Rosa, I.L. Regional-Scale Patterns of Habitat Preference for the Seahorse Hippocampus Reidi in the Tropical Estuarine Environment. Aquat. Ecol. 2015, 49, 499–512. [Google Scholar] [CrossRef]
- Ape, F.; Corriero, G.; Mirto, S.; Pierri, C.; Lazic, T.; Gristina, M. Trophic flexibility and prey selection of the wild long-snouted seahorse Hippocampus guttulatus Cuvier, 1829 in three coastal habitats. Estuar. Coast. Shelf S. 2019, 224, 1–10. [Google Scholar] [CrossRef]
- Lazic, T.; Fosso, B.; Bachir, B.; Corriero, G.; Gristina, M.; Marzano, M.; Pesole, G.; Santamaria, M.; Pierri, C. Hippocampus guttulatus Diet Based on DNA Metabarcoding. Front. Mar. Sci. 2023, 10, 1138279. [Google Scholar] [CrossRef]
- Foster, S.A.; Vincent, A.C. Life history and ecology of seahorses: Implications for conservation and management. J. Fish Biol. 2004, 65, 1–61. [Google Scholar] [CrossRef]
- Joseph, E. A Study of the Systematics and Life History of the Gulf Pipefish, Syngnathus scovelli (Evermann and Kendall). Ph.D. Thesis, The Florida State University, Tallahassee, FL, USA, 1957. [Google Scholar]
- Woodall, L.C.; Otero-Ferrer, F.; Correia, M.; Curtis, J.M.; Garrick-Maidment, N.; Shaw, P.W.; Koldewey, H.J. A synthesis of European seahorse taxonomy, population structure, and habitat use as a basis for assessment, monitoring and conservation. Mar. Biol. 2018, 165, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Frando, A.; Malavasi, S.; Zucchetta, M.; Franzoi, M.; Torricelli, P. Environmental influences on fish assemblage in the Venice Lagoon, Italy. Chem Ecol. 2006, 22, S105–S118. [Google Scholar] [CrossRef]
- Pineiro-Corbeira, C.; Iglesias, L.; Nogueira, R.; Campos, S.; Jimenez, A.; Regueira, M.; Berreiro, R.; Planas, M. Structure and trophic niches in mobile epifauna assemblages associated with seaweeds and habitats of syngnathid fishes in Cíes archipelago (Atlantic Islands Marine National Park, Northwest Iberia). Front. Mar. Sci. 2021, 8, 773367. [Google Scholar] [CrossRef]
- Planas, M. Ecological traits and trophic plasticity in the greater pipefish Syngnathus acus in the NW Iberian Peninsula. Biology 2022, 11, 712. [Google Scholar] [CrossRef] [PubMed]
- Louisy, P. Hippocampus Guttulatus, l’espèce Commune de l’étang de Thau, Hippo-Thau Bilan Scientifique 2005–2009; CPIE: Bassin de Thau, France, 2011. [Google Scholar]
- Faleiro, F.; Baptista, M.; Santos, C.; Aurélio, M.L.; Pimentel, M.; Pegado, M.R.; Paula, J.R.; Calado, R.; Rosa, R. Seahorses under a changing ocean: The impact of warming and acidification on the behaviour and physiology of a poor-swimming bony-armoured fish. Conserv. Physiol. 2015, 3, cov009. [Google Scholar] [CrossRef] [PubMed]
- Lazic, T.; Pierri, C.; Corriero, G.; Balech, B.; Cardone, F.; Deflorio, M.; Fosso, B.; Gissi, C.; Marzano, M.; Nonnis Marzano, F.; et al. Evaluating the efficiency of DNA metabarcoding to analyze the diet of Hippocampus guttulatus (Teleostea: Syngnathidae). Life 2021, 11, 998. [Google Scholar] [CrossRef]
- Diaz-Ruiz, S.; Aguirre-Leon, A.; Perez-Soli, O. Distribution and abundance of Syngnathus louisianae and Syngnathus scovelli (Syngnathidae) in Tamiahua Lagoon, Gulf of Mexico. Cienc. Mar. 2000, 26, 125–143. [Google Scholar] [CrossRef]
- Lourie, S.A.; Pollom, R.A.; Foster, S.J. A global revision of the seahorses Hippocampus Rafinesque 1810 (Actinopterygii: Syngnathiformes): Taxonomy and biogeography with recommendations for further research. Zootaxa 2016, 4146, 1–66. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station | Habitat | Depth | Syngnathid Species |
|---|---|---|---|
| St 1 | Muddy–sandy seabed with patchy meadows of C. nodosa and Chaetomorpha spp. and abundant fine vegetal detritus. Dense population of Cerithium vulgatum and Hexaplex trunculus. The site is weakly affected by tidal streams, with hydrological features mainly referring to the lagoon ecosystem. | 0.5–1.5 m | Hippocampus hippocampus Nerophis ophidion Syngnathus abaster |
| St 2 | Muddy seabed with stable occurrence of Chaetomorpha spp. Patchy meadows of C. nodosa were also present. Structured macrobenthic community with dense populations of H. trunculus, Haminoea japonica, and Gammarus insensibilis. The site is not affected by tidal streams, with hydrological features mainly referring to the lagoon ecosystem. | 1–3 m | N. ophidion S. abaster |
| St 3 | Muddy seabed with stable occurrence of Chaetomorpha spp. Patchy meadows of C. nodosa were also present. Structured macrobenthic community with dense populations of H. trunculus, H. japonica, and G. insensibilis. The site is not affected by tidal streams, with hydrological features mainly referring to the lagoon ecosystem. | 1–3.5 m | N. ophidion S. abaster |
| St 4 | Muddy–sandy seabed with C. nodosa meadows extending to the whole channel mouth area with abundant fine vegetal detritus. Dense population of Cerithium vulgatum and H. trunculus. Strong tidal flow with hydrological features changing according to tidal phases. | 0.5–1.5 m | H. guttulatus H. hippocampus N. ophidion S. abaster |
| St 5 | Muddy–sandy seabed with patchy meadows of C. nodosa and Chaetomorpha sp. Dense populations of C. vulgatum. and H. trunculus be found, along with a more complex malacocenosis. The site is weakly affected by tidal streams, with hydrological features mainly referring to the lagoon ecosystem. | 0.5–2 m | H. hippocampus N. ophidion S. abaster |
| St 6 | Muddy–sandy seabed with patchy meadows of C. nodosa and abundant fine vegetal detritus. Dense populations of C. vulgatum. and H. trunculus were found. Moderate tidal flow with hydrological features changing according to tidal phases. | 1–2.5 m | H. guttulatus H. hippocampus N. ophidion S. abaster |
| St 7 | Muddy seabed with seasonal occurrence of algal coverage (Ulva spp., Chaetomorpha spp.). Poor seabed macrobenthic community. Dense population of Ciona intestinalis, Styela plicata, Aiptasia mutabilis, and Ficopomatus enigmaticus colonise artificial rocky substrates. The site is not affected by tidal streams, with hydrological features mainly referring to the lagoon ecosystem. | 1–2 m | |
| St 8 | Sandy bottom with a thick coverage of organic matter mainly composed of P. oceanica leaves and rhizome fibres. Tidal flow is weaker than at Station 9. | 0.5–1 m | H. guttulatus H. hippocampus |
| St 9 | Sandy bottom with scattered accumulation of vegetal detritus, gravel, and compacted organic matter. Closer to the inner mouth, the channel is affected by a sparsely packed grassland of C. nodosa. Strong tidal flow with hydrological features changing according to tidal phases. | 0.5–2 m | H. guttulatus H. hippocampus |
| St 10 | Coarse shell-bearing sands with terrestrial and marine vegetal detritus, including tree branches, P. oceanica rhizomes, and detached thalli of Codium spp. High seabed slope, reaching 3.5 m in depth in the central part. Strong tidal flow velocity leaves large portions of rocks uncovered. Hydrological features change according to tidal phases. | 0.5–3.5 m | |
| St 11 | Coarse sandy bottom with scattered vegetal detritus, without areas of accumulation. Hard substrates colonised by dense populations of anthozoans (Anemonia viridis, Paranemonia cinerea). Strong tidal flow velocity. Hydrological features change according to tidal phases. | 0.5–1.5 m |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lazic, T.; Pierri, C.; Corriero, G.; Gravina, M.F.; Gristina, M.; Ravisato, M.; Macali, A. Abundance, Distribution, and Habitat Preference of Syngnathid Species in Sabaudia Lake (Tyrrhenian Sea). Diversity 2023, 15, 972. https://doi.org/10.3390/d15090972
Lazic T, Pierri C, Corriero G, Gravina MF, Gristina M, Ravisato M, Macali A. Abundance, Distribution, and Habitat Preference of Syngnathid Species in Sabaudia Lake (Tyrrhenian Sea). Diversity. 2023; 15(9):972. https://doi.org/10.3390/d15090972
Chicago/Turabian StyleLazic, Tamara, Cataldo Pierri, Giuseppe Corriero, Maria Flavia Gravina, Michele Gristina, Miriam Ravisato, and Armando Macali. 2023. "Abundance, Distribution, and Habitat Preference of Syngnathid Species in Sabaudia Lake (Tyrrhenian Sea)" Diversity 15, no. 9: 972. https://doi.org/10.3390/d15090972
APA StyleLazic, T., Pierri, C., Corriero, G., Gravina, M. F., Gristina, M., Ravisato, M., & Macali, A. (2023). Abundance, Distribution, and Habitat Preference of Syngnathid Species in Sabaudia Lake (Tyrrhenian Sea). Diversity, 15(9), 972. https://doi.org/10.3390/d15090972