


A Mitogenome-Based Phylogeny of Pilargidae (Phyllodocida, Polychaeta, Annelida) and Evaluation of the Position of Antonbruunia †



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Extraction, Amplification, Sequencing, and Assembly
2.3. Morphological Analyses
2.4. Phylogenetic Analyses
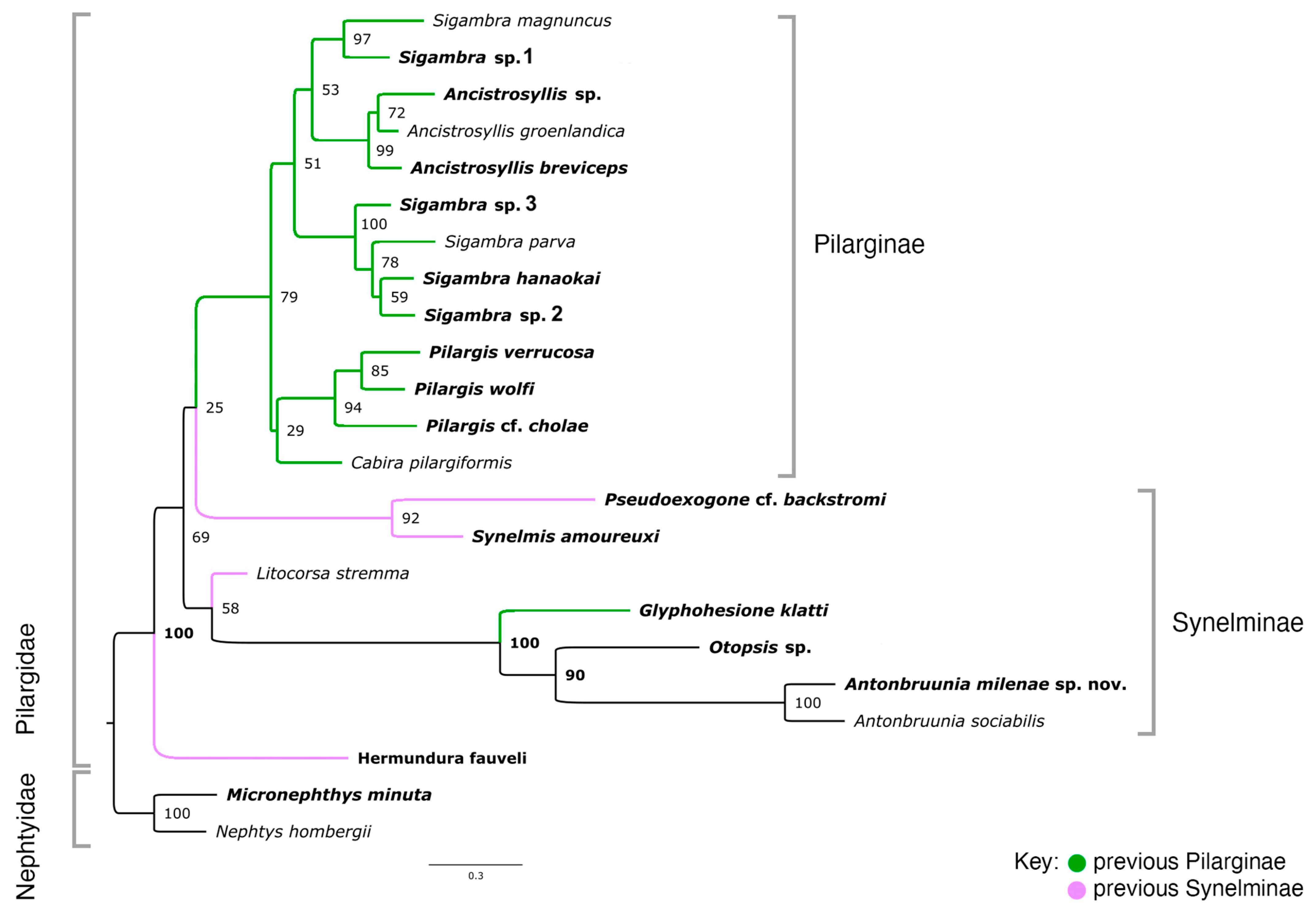
3. Results
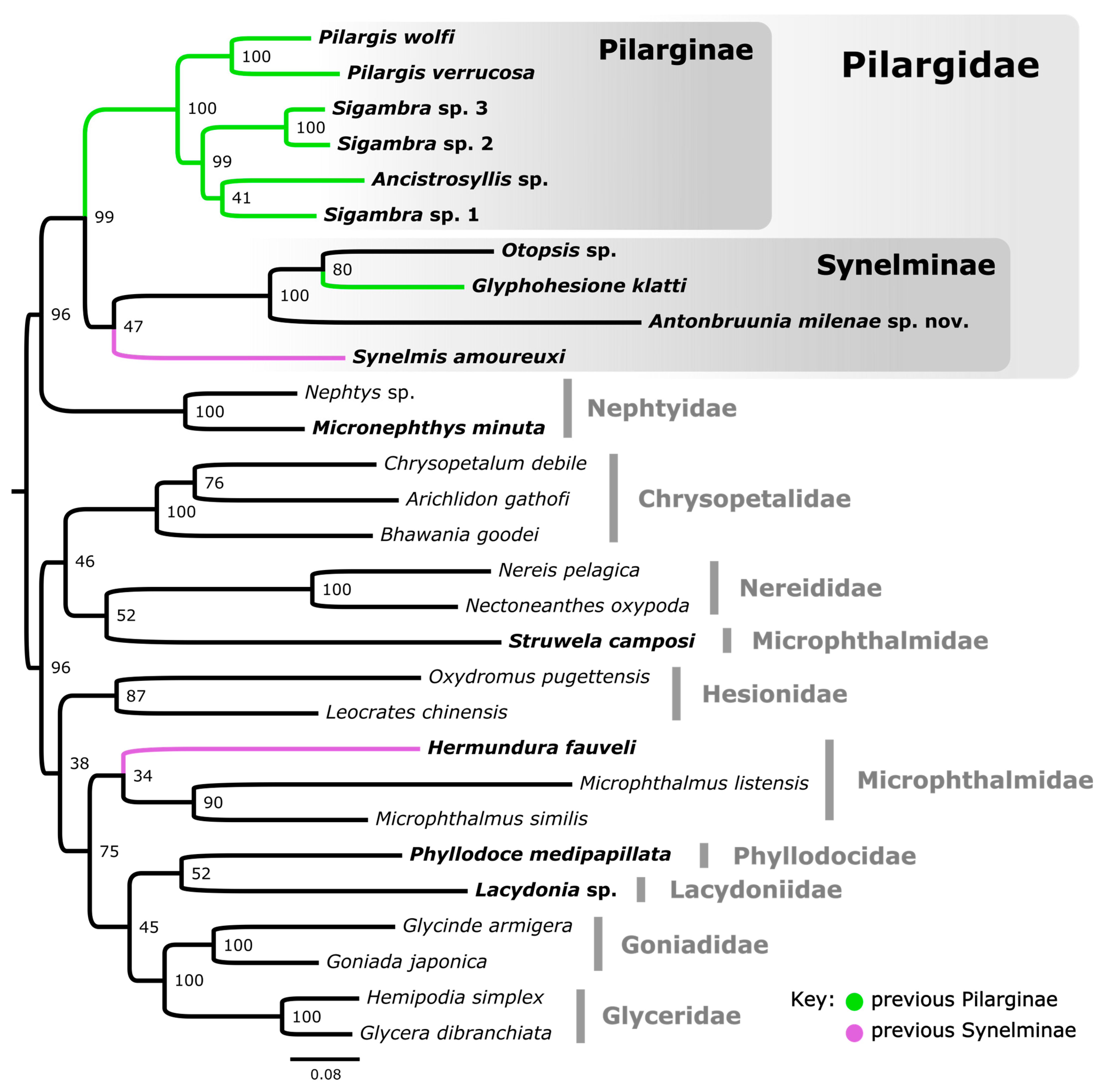
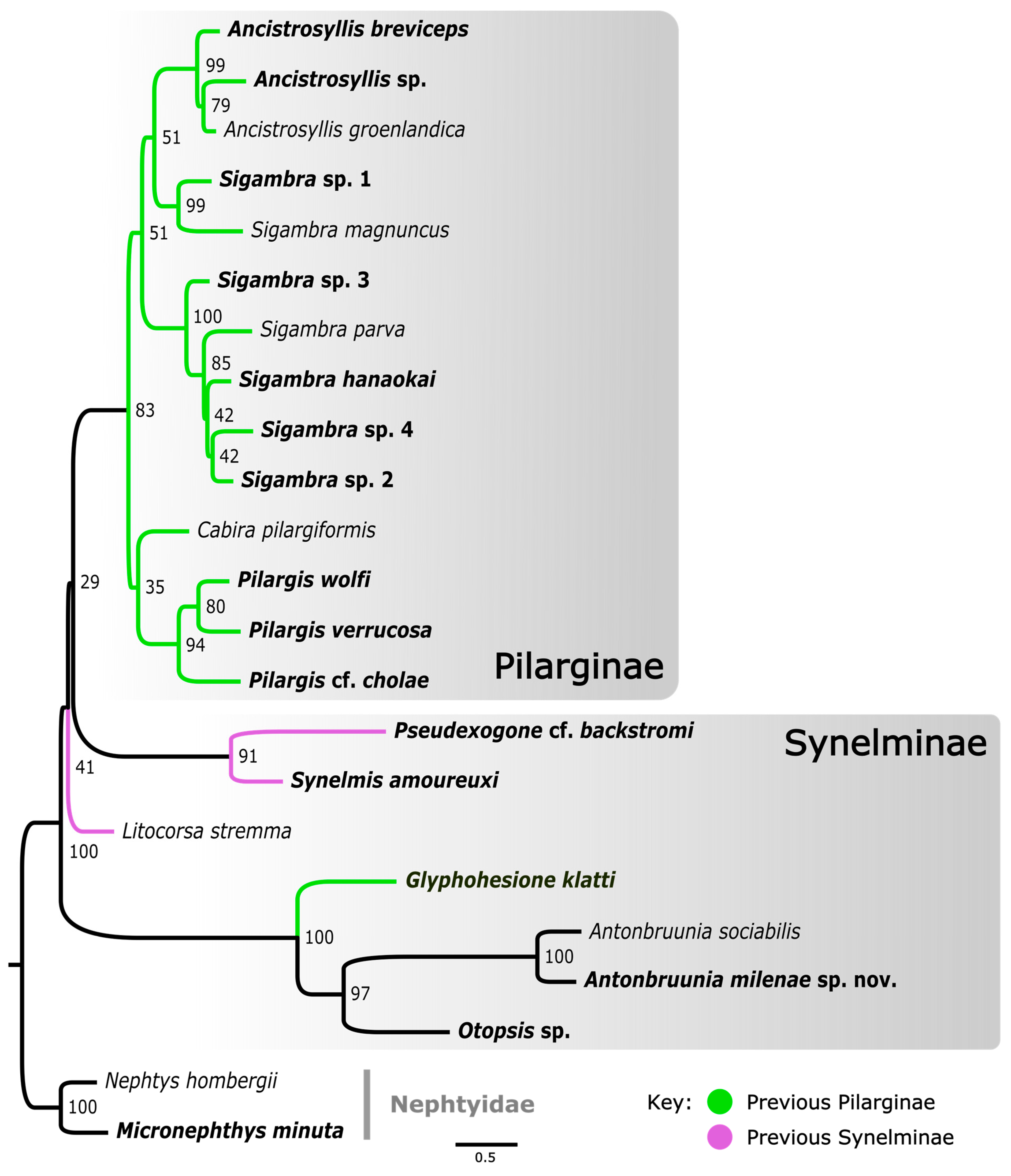
3.1. Phylogenetic Results
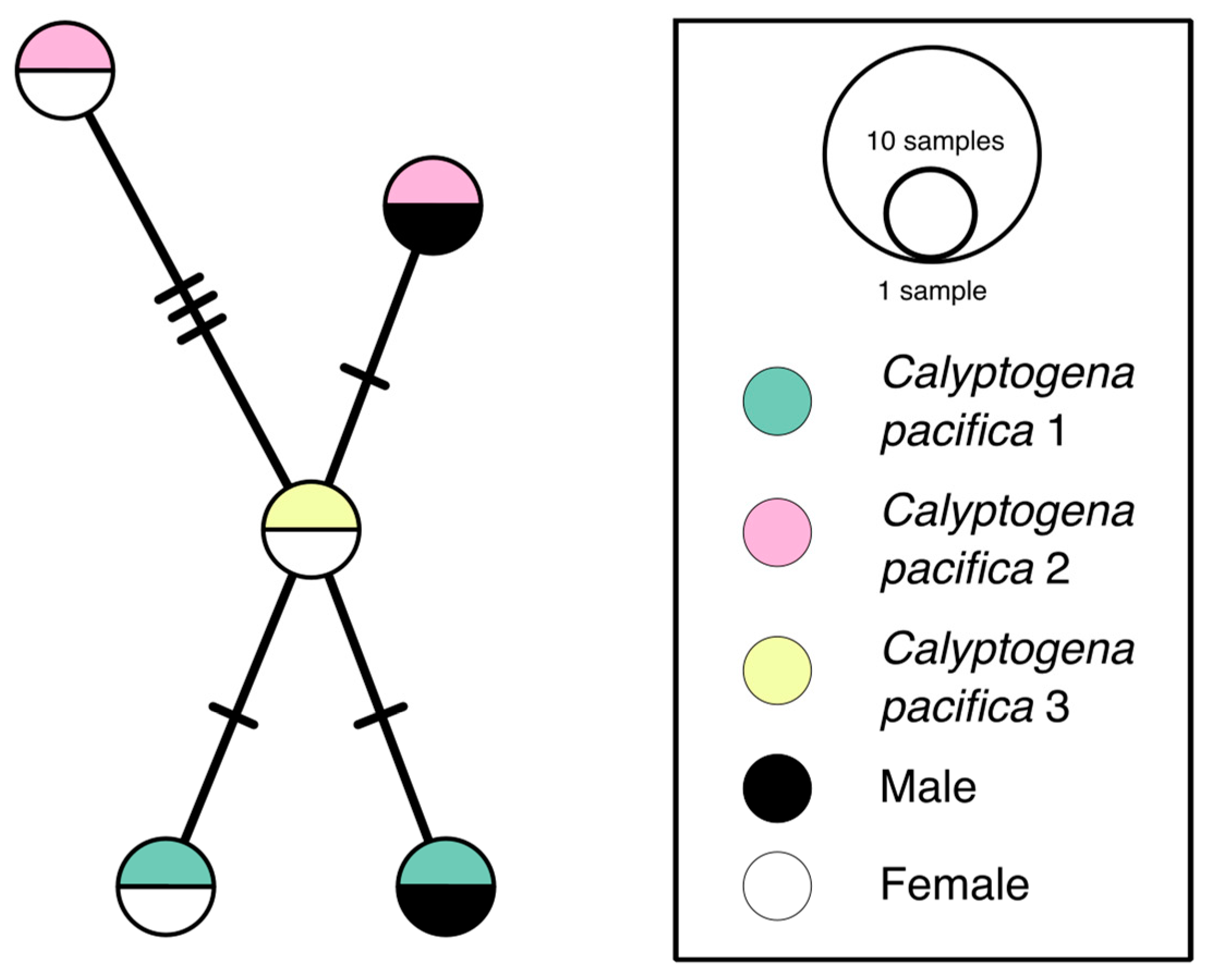
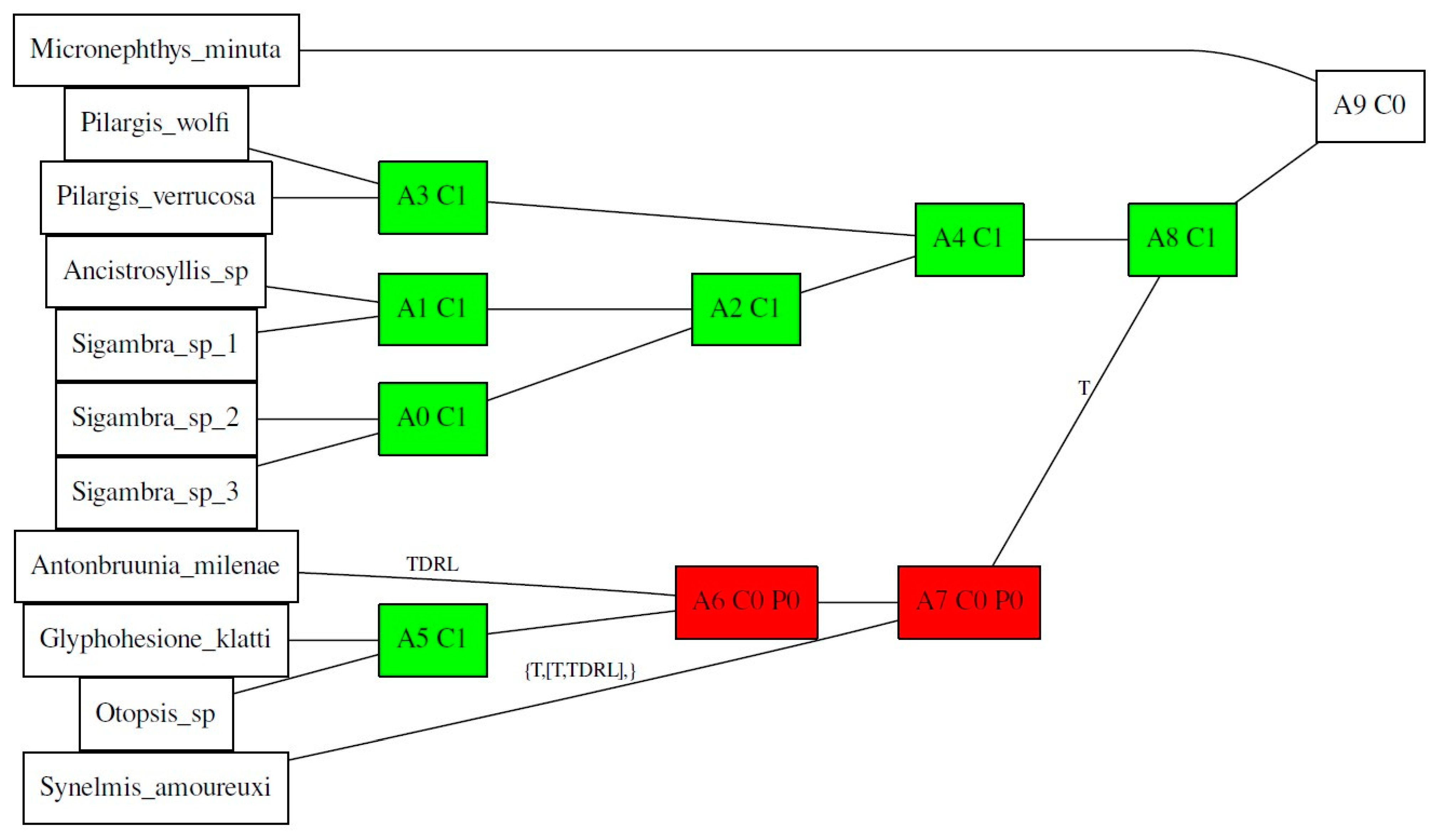
3.2. Haplotype Network
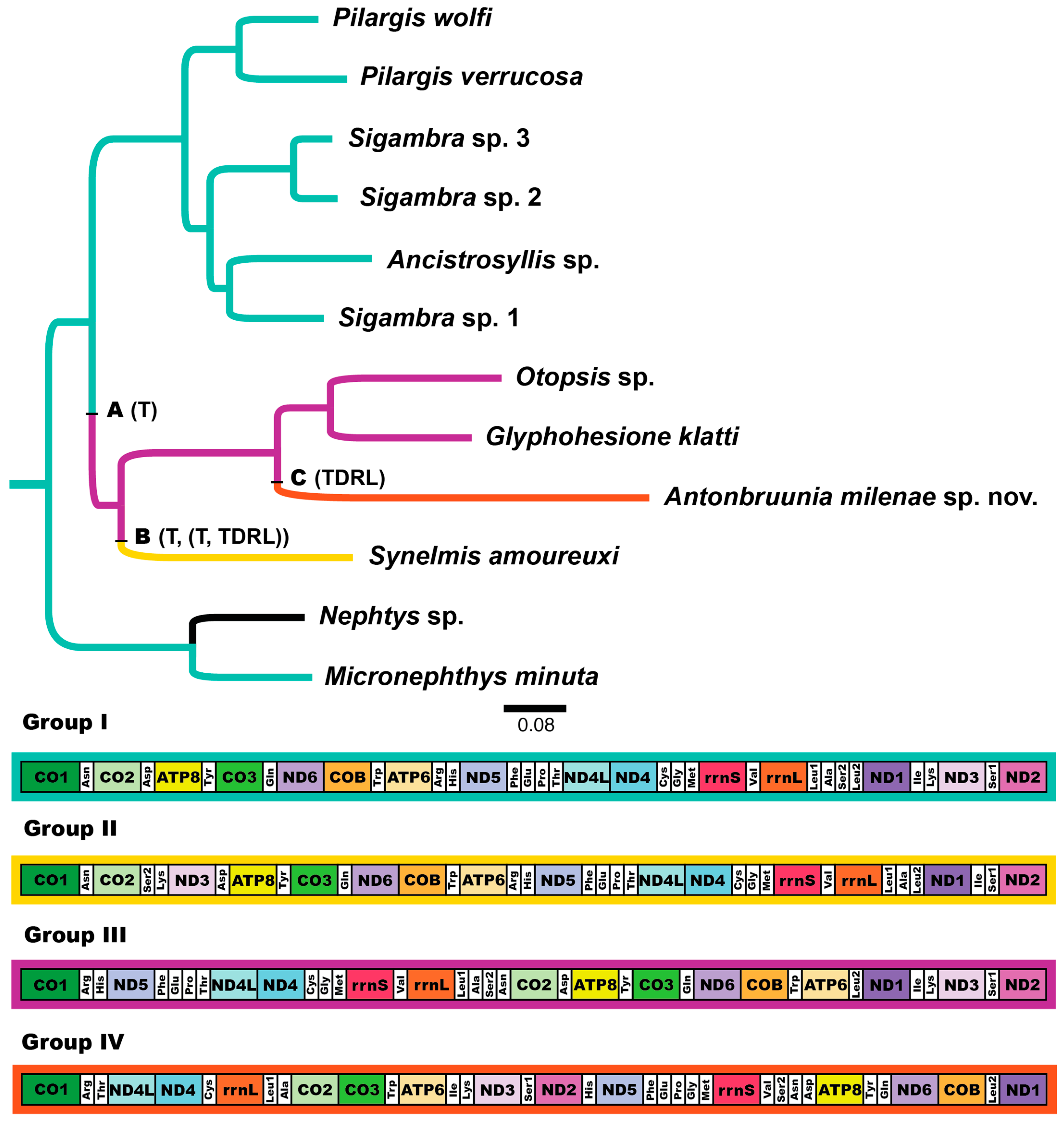
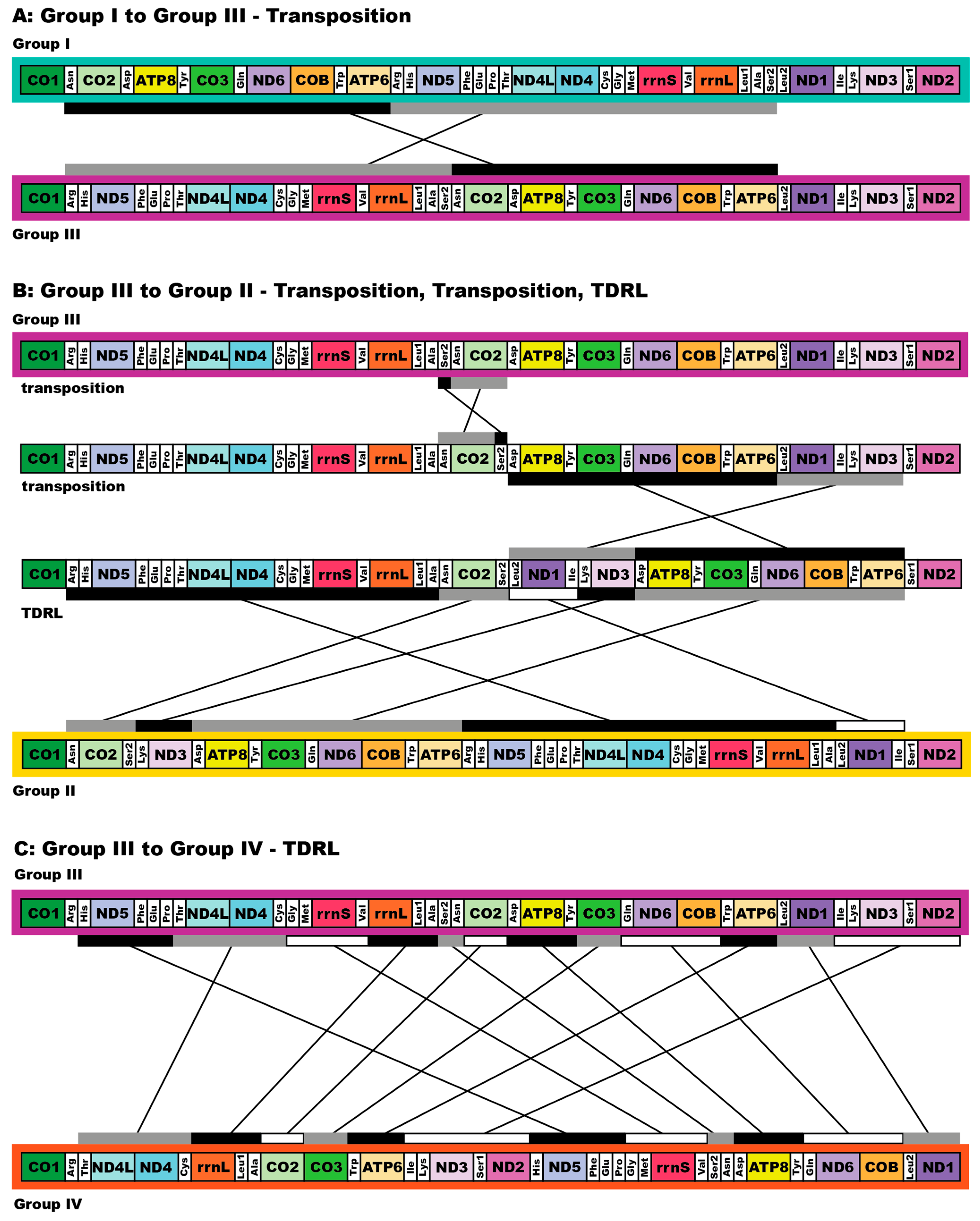
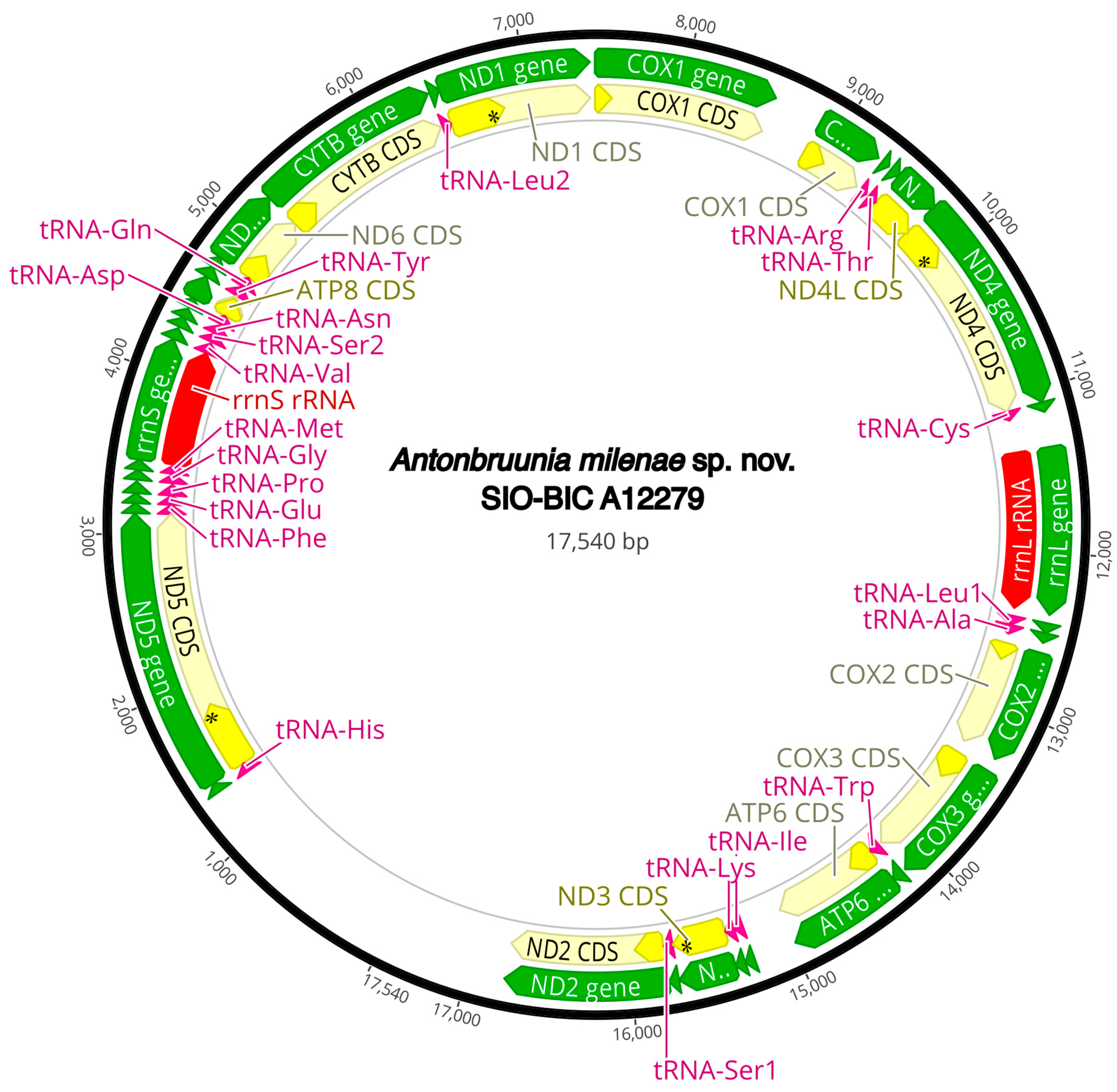
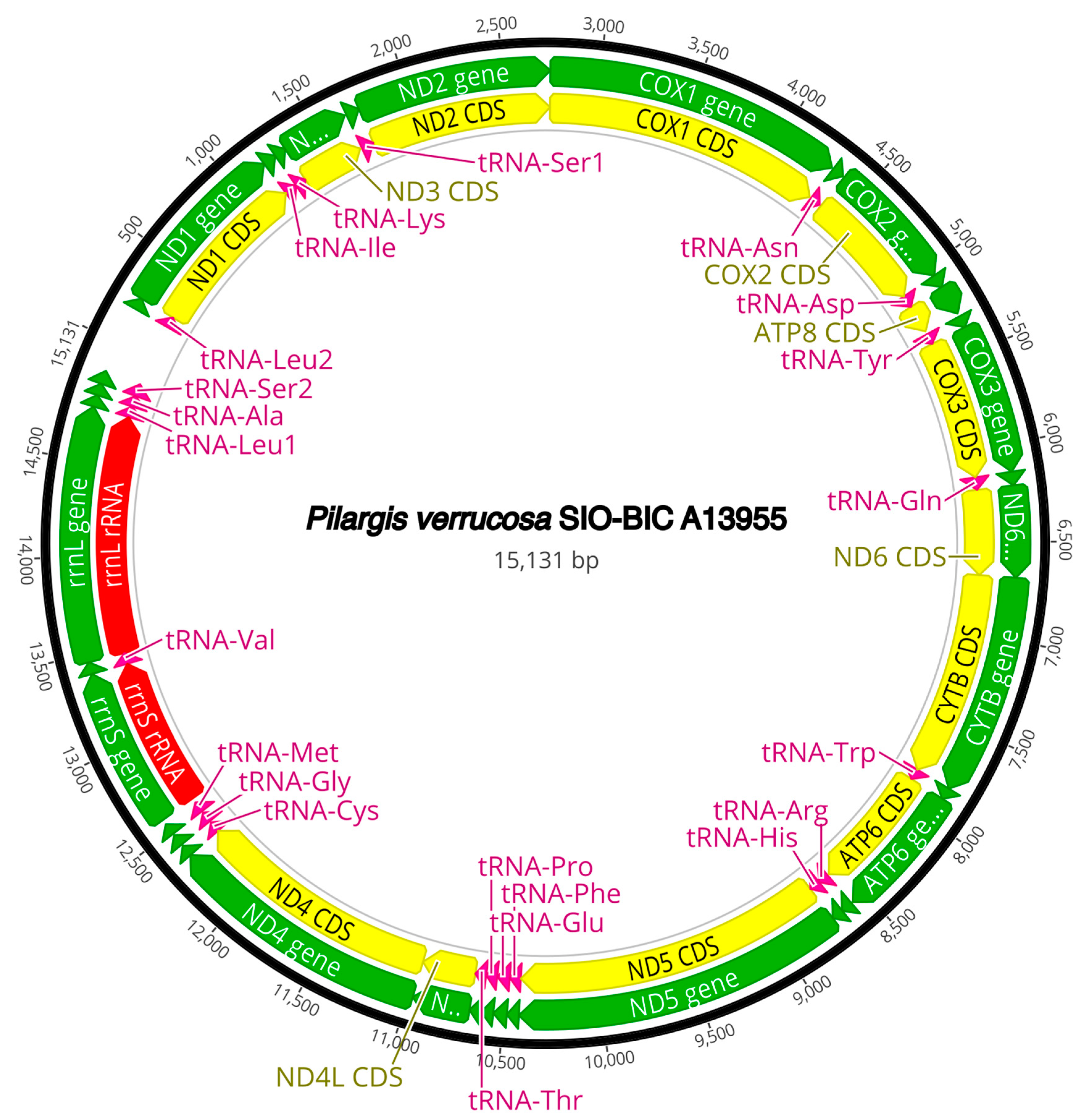
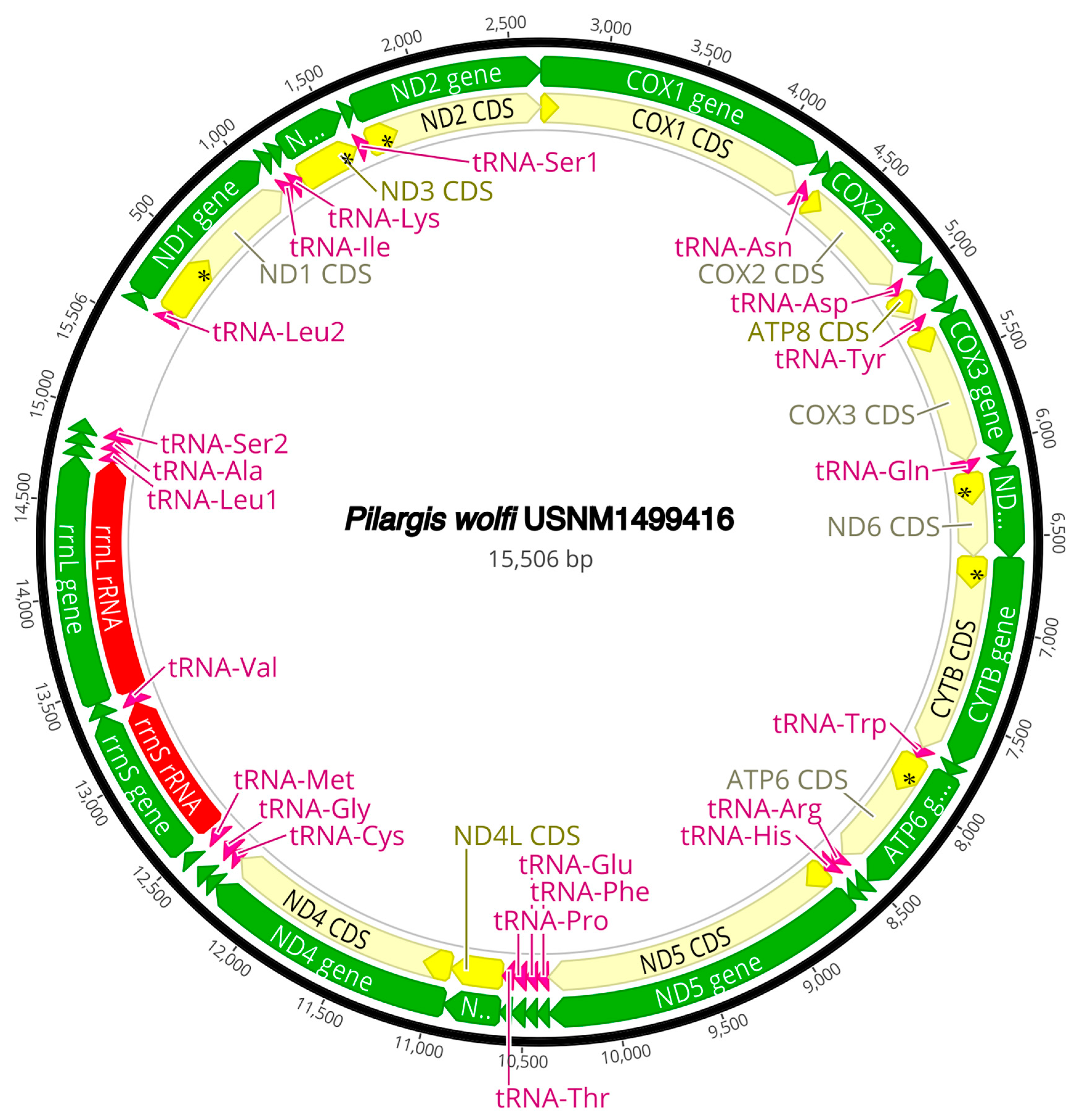
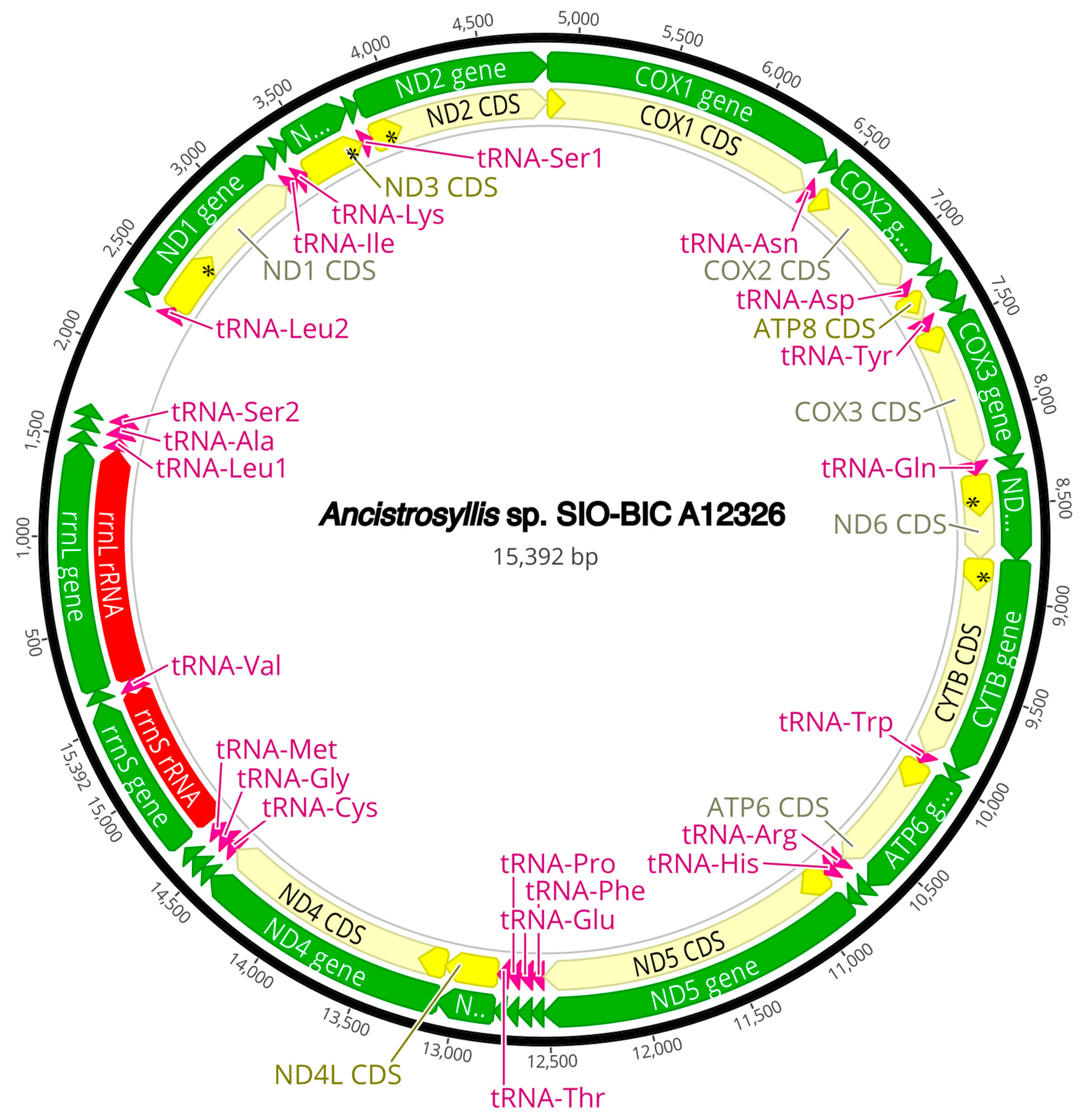
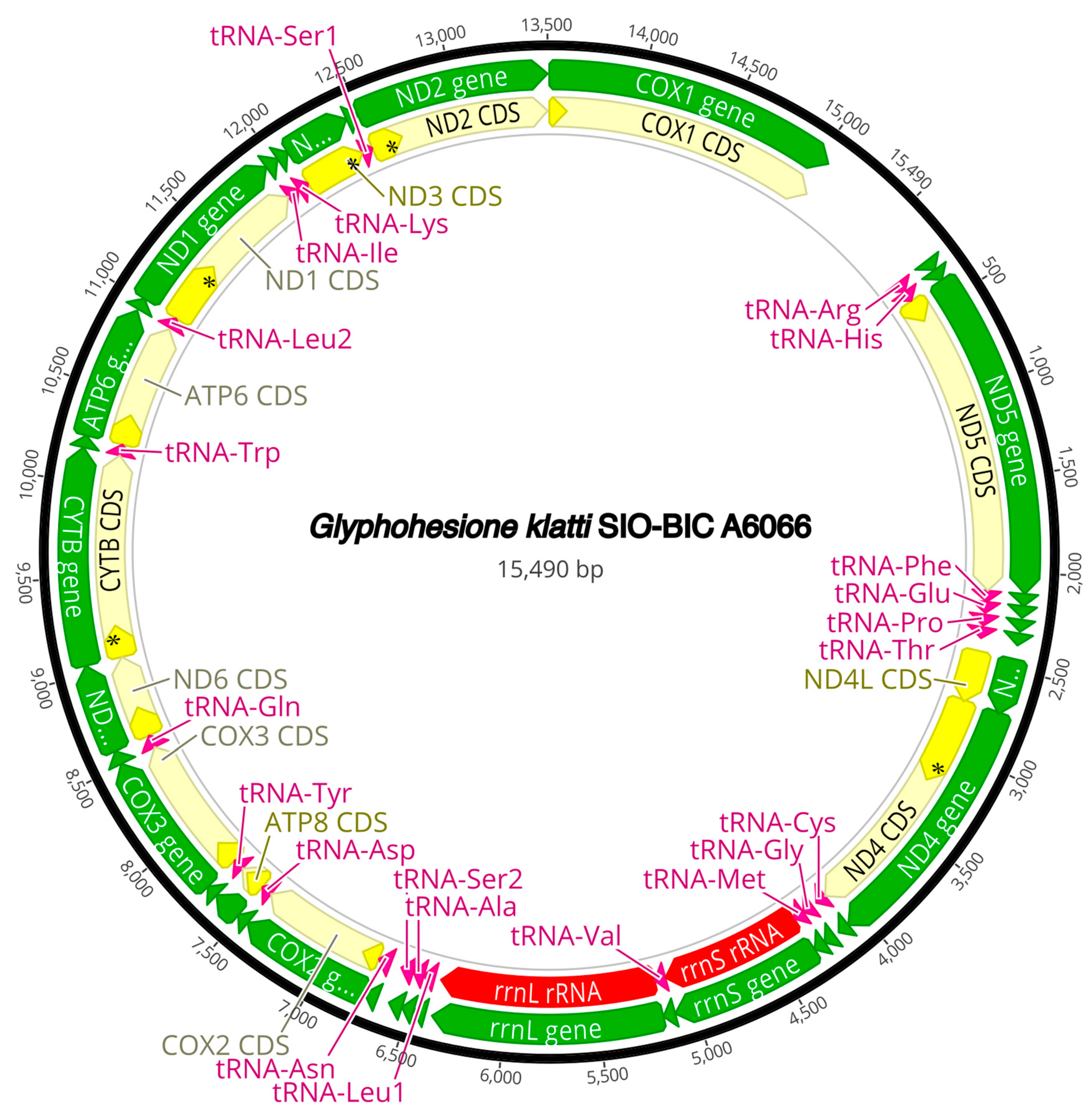
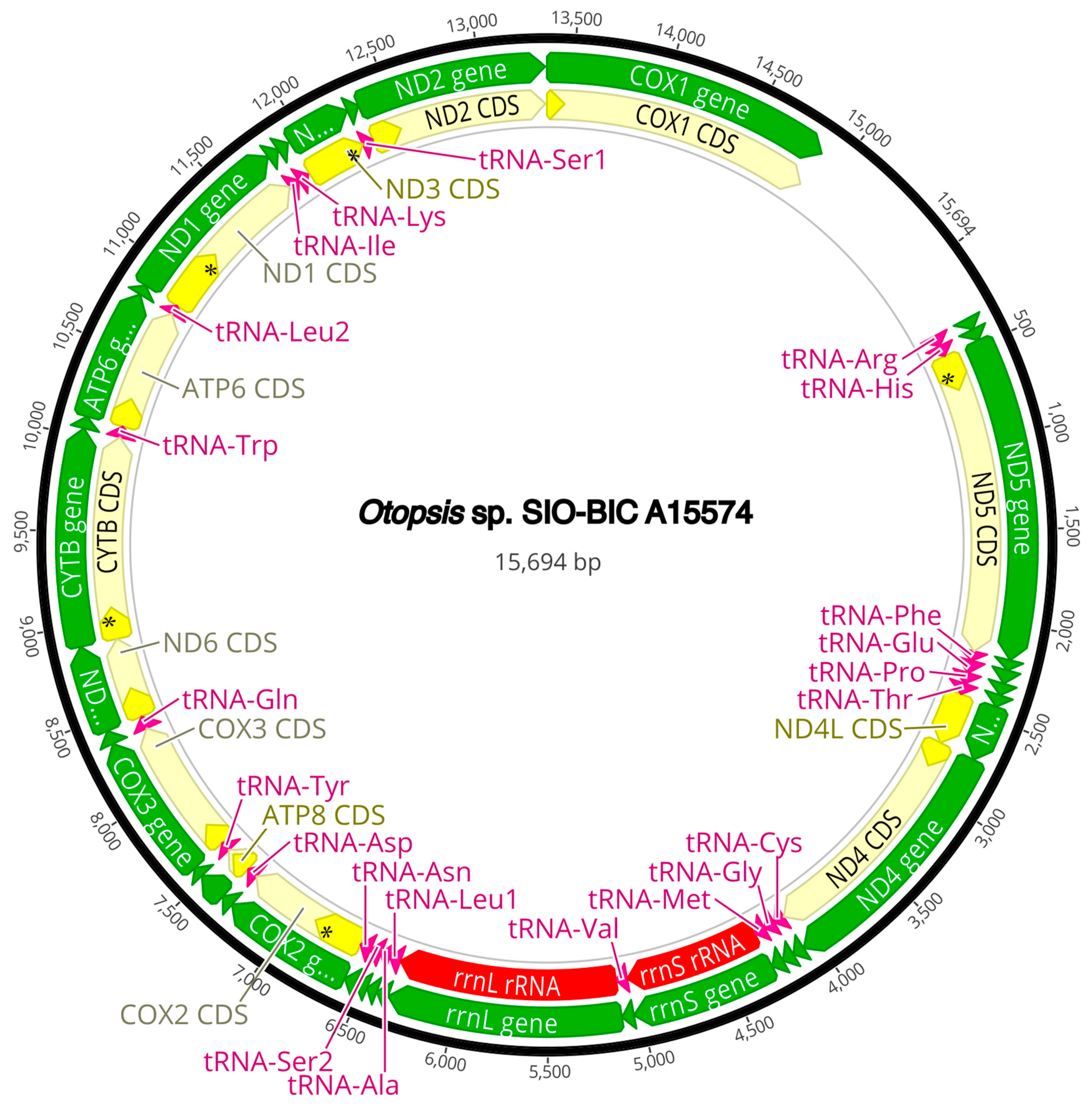
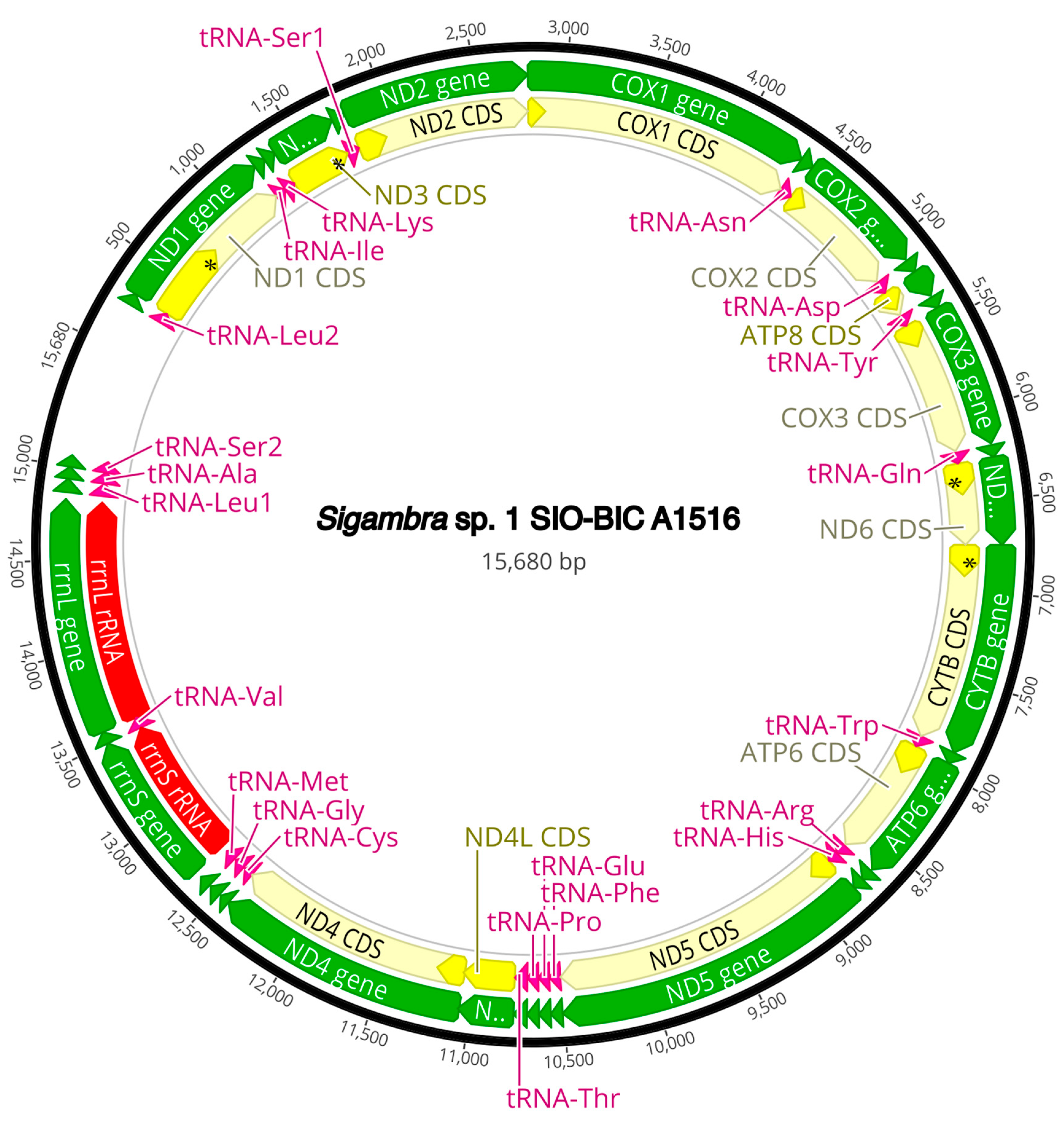
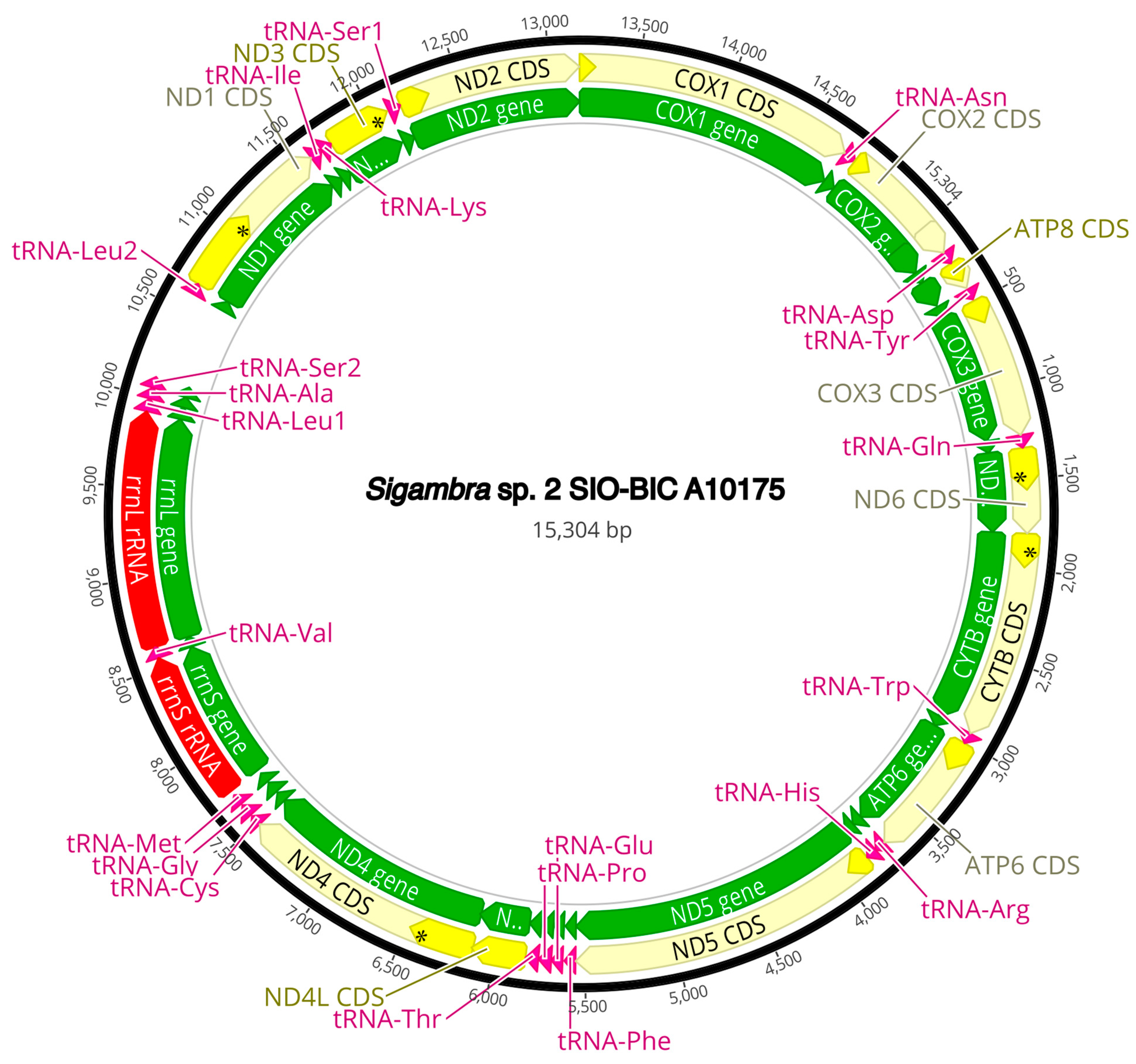
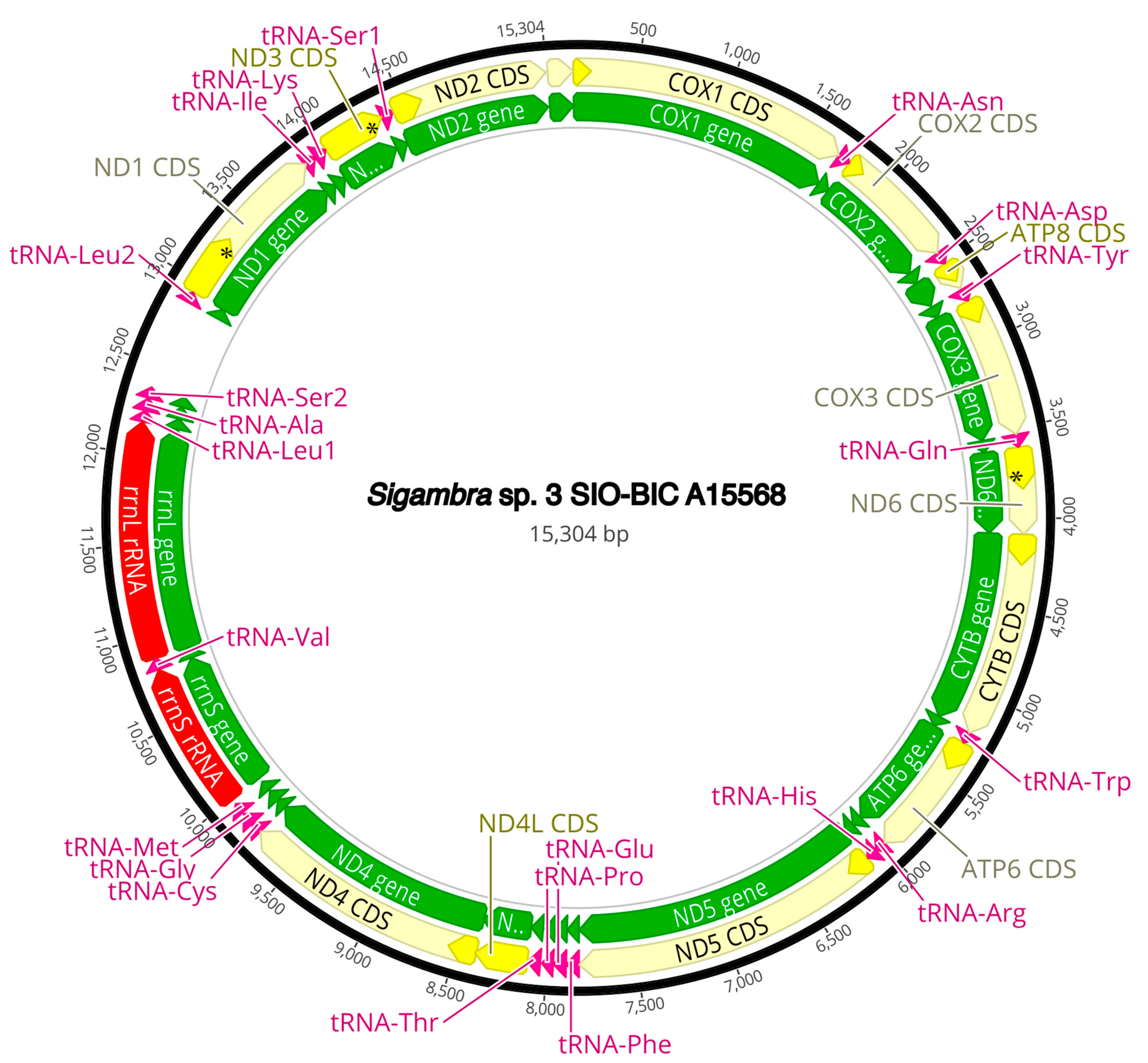
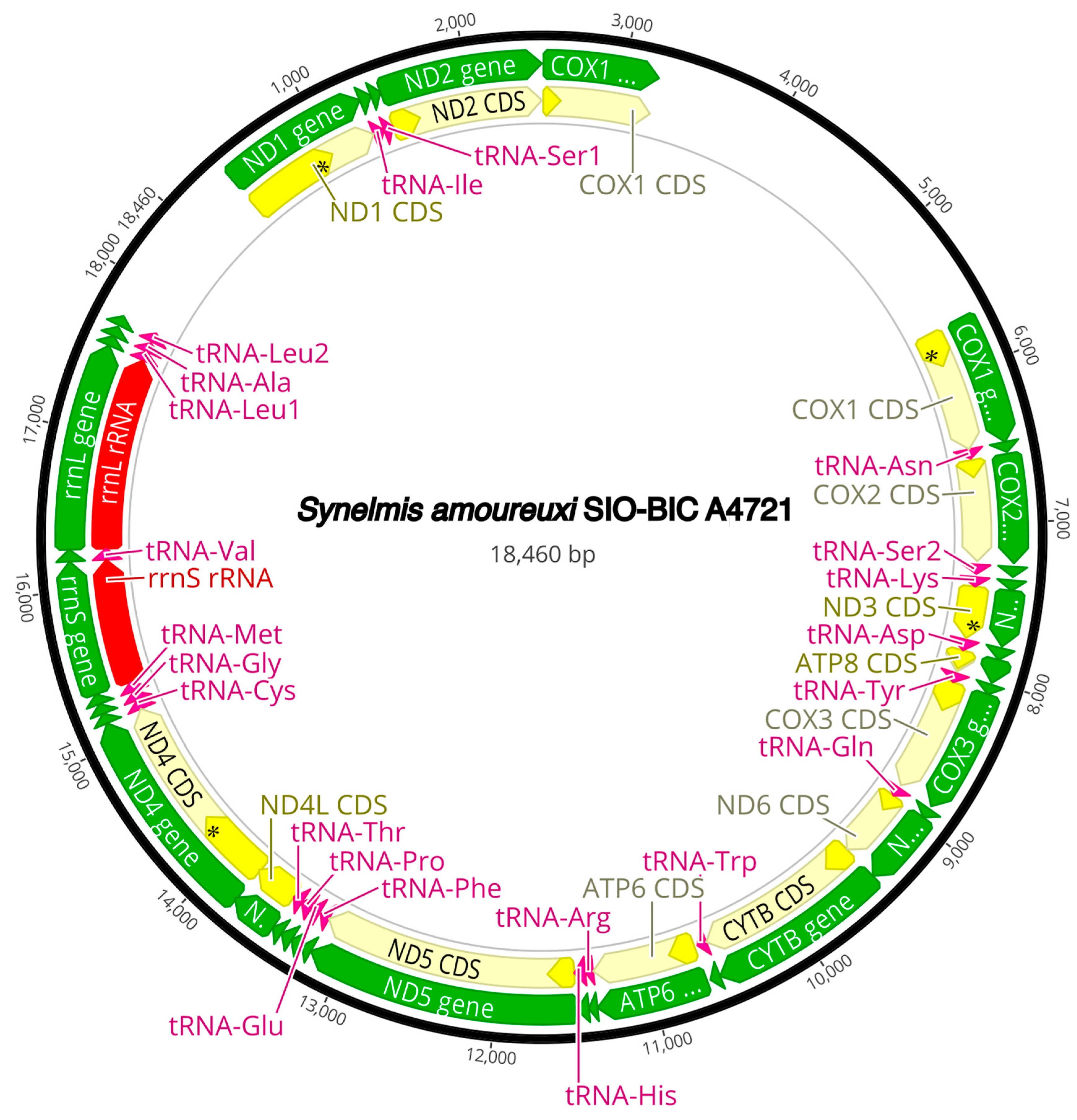
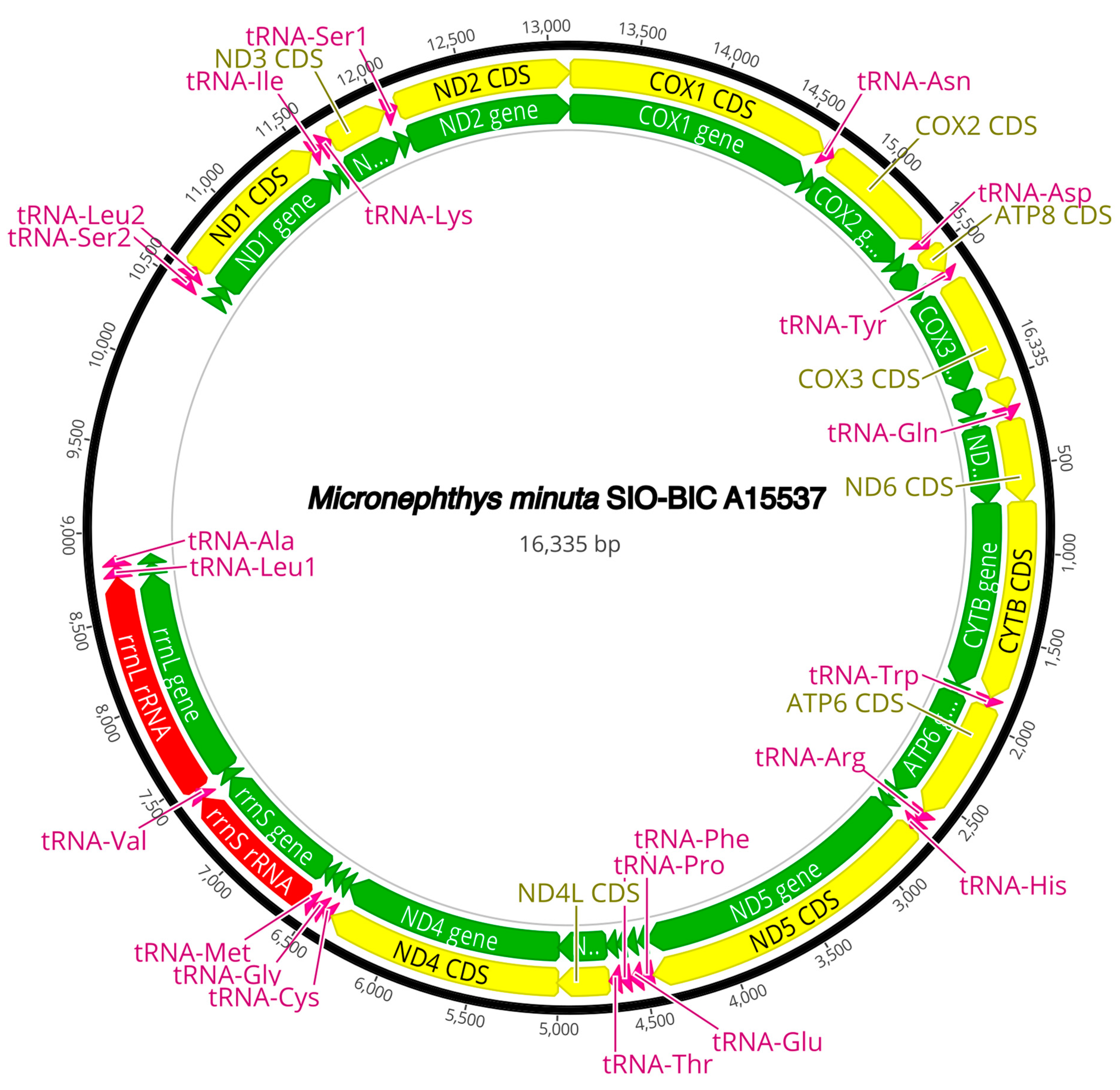
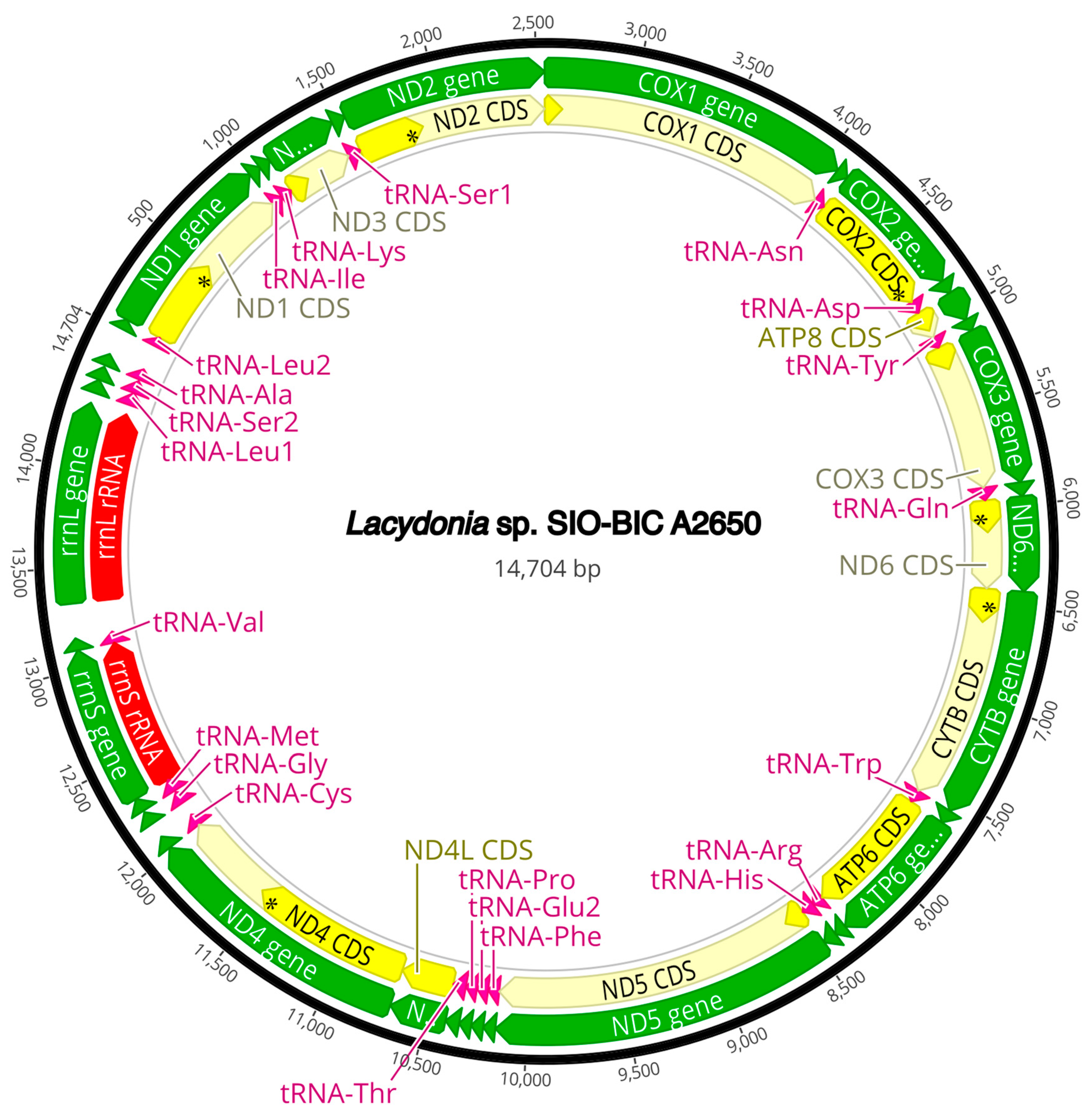
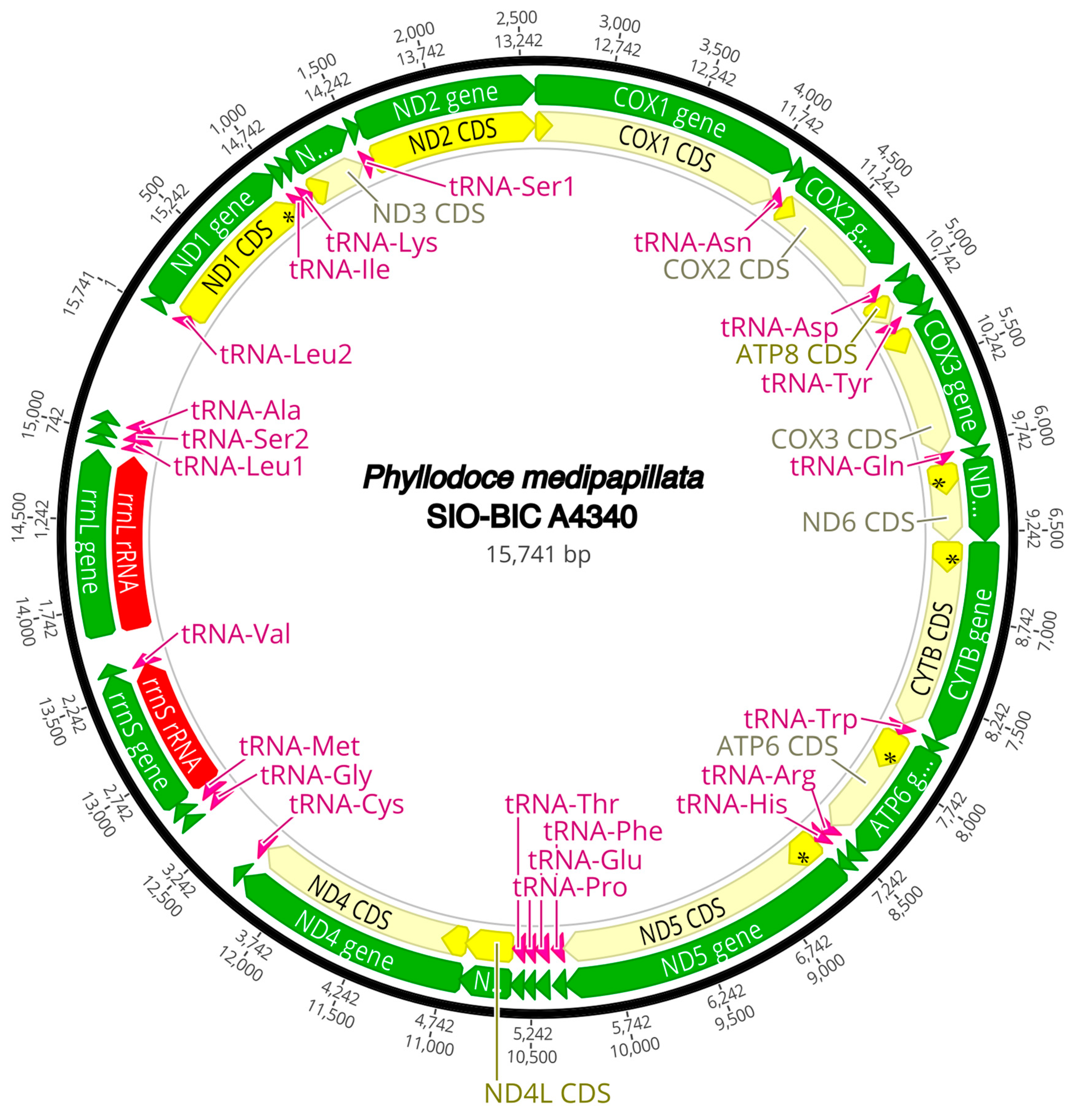
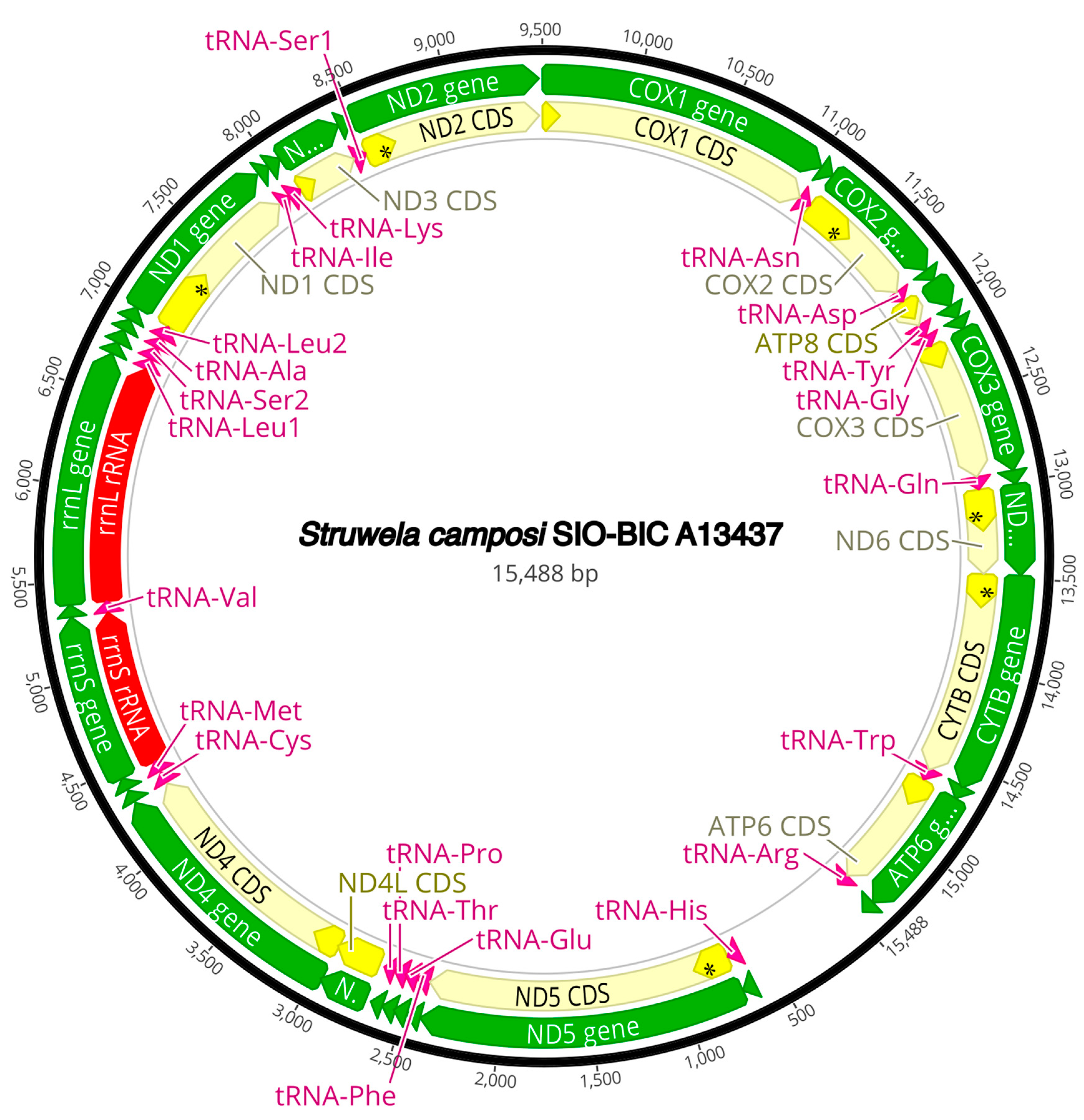
3.3. Mitogenome Organization
3.4. Taxonomy
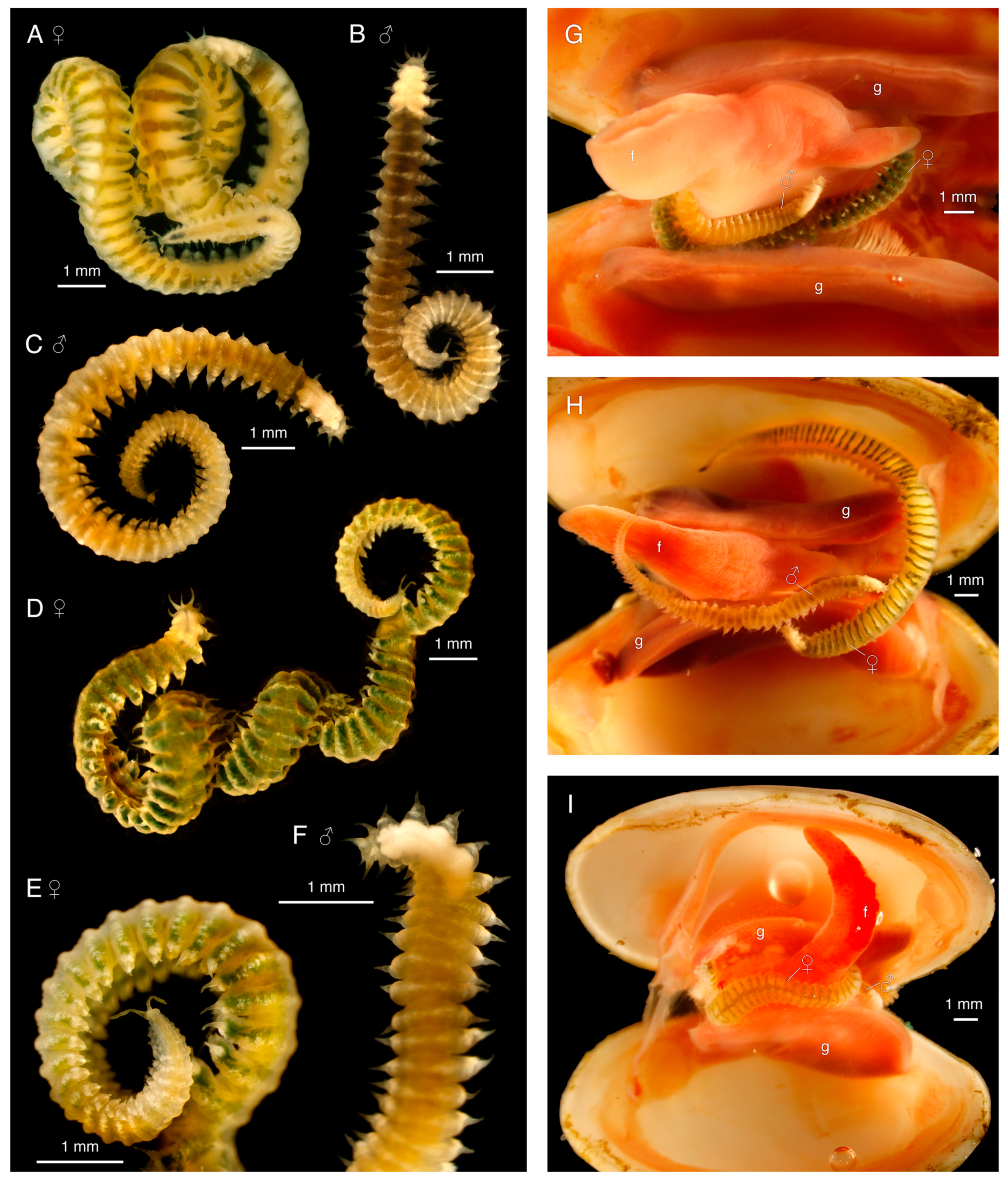
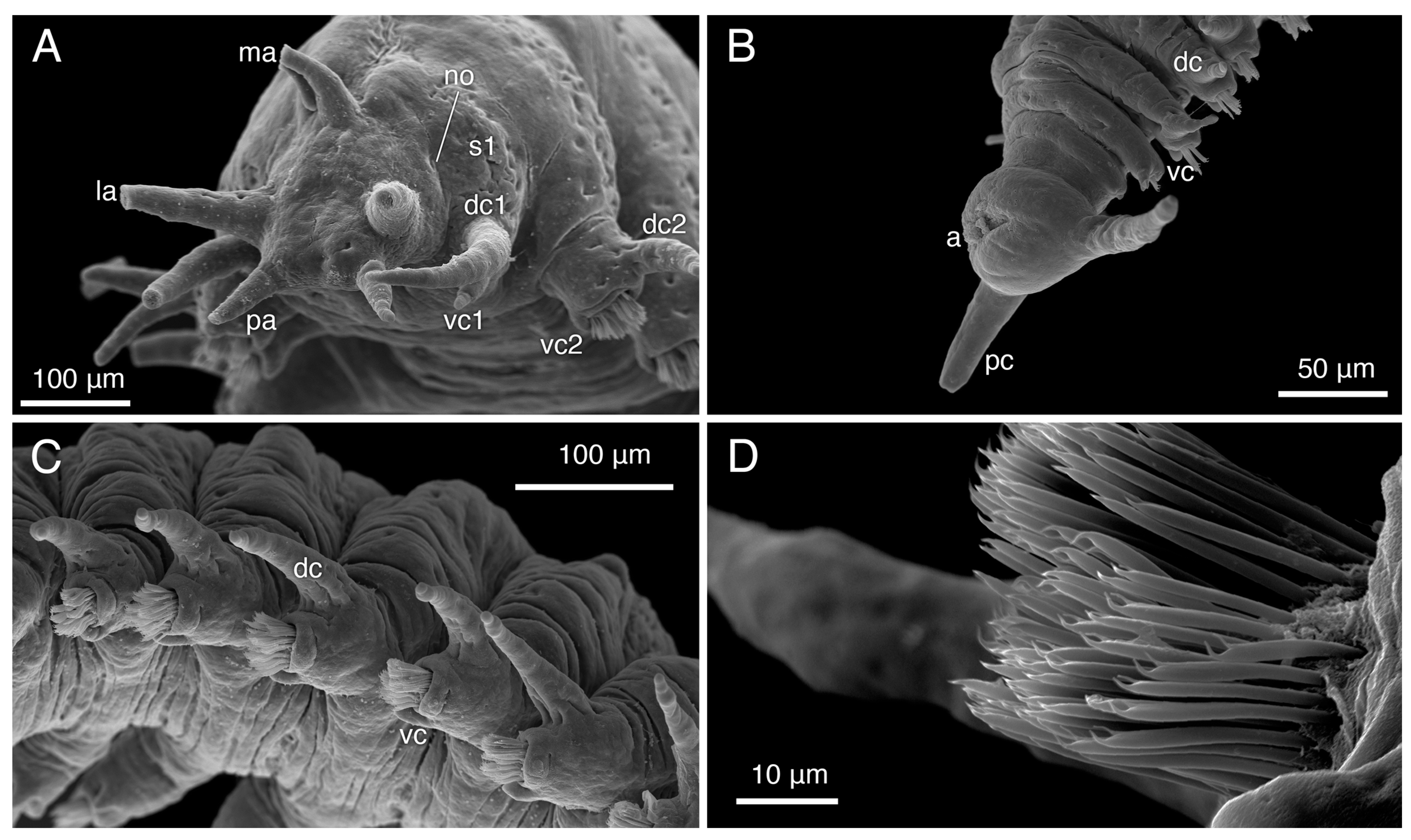
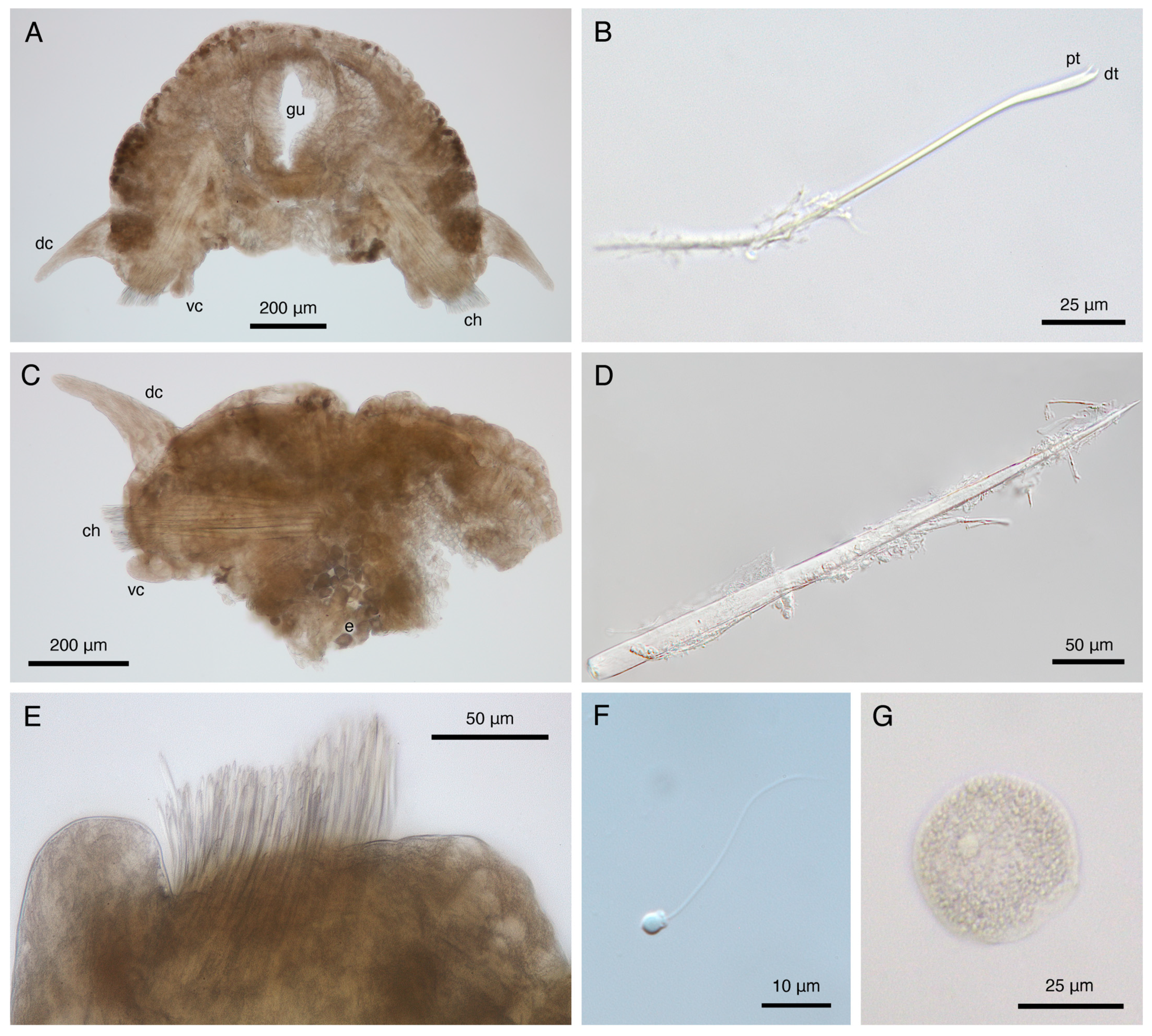
3.4.1. Description
3.4.2. Variation
3.4.3. Habitat
3.4.4. Reproduction
3.4.5. Remarks
3.4.6. Etymology
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Locality | Depth (m) | Host | Worms per Host | Habitat | Reference |
|---|---|---|---|---|---|---|
| A. viridis | Madagascar | 70 | Lucina fosteri | Usually 2 (male + female) | Hypoxic sediment | Hartman and Boss 1965 [8] |
| A. gerdesi | Central Chile | 800 | Calyptogena gallardoi | Usually 1 | Gas hydrate environment | Quiroga and Sellanes, 2009 [9] |
| A. sociabilis | Scotland | 1200 | Thyasira scotiae | 2–9 | Sulfide seep | Mackie, Oliver, and Nygren, 2015 [7] |
| A. milenae sp. nov. | CA, USA | 845 | Calyptogena pacifica | 2 (male + female) | Whale fall | This study |
| Gene | Primer Name | Primer Sequence | Reference |
|---|---|---|---|
| COI | polyLCO (F) | 5′-GAYTATWTTCAACAAATCATAAAGATATTGG-3′ | [116] |
| COI | polyHCO (R) | 5′-TAMACTTCWGGGTGACCAAARAA TCA-3′ | [116] |
| 16S | 16S-AnnF (F) | 5′-GCGGTATCCTGACCGTRCWAAGGTA-3′ | [117] |
| 16S | 16Sb (R) | 5′-CTCCGGTTTGAACTCAGATCA -3′ | [118] |
| 16S | 16SarL (F) | 5′-CGCCTGTTTATCAAAAACAT-3′ | [119] |
| 16S | 16SbrH (R) | 5′-CCGGTCTGAACTCAGATCACGT-3′ | [119] |
| 18S | 18S-1F (F) | 5′-TACCTGGTTGATCCTGCCAGTAG-3′ | [120] |
| 18S | 18S-5R (R) | 5′-CTTGGCAAATGCTTTCGC-3′ | [120] |
| 18S | 18S-3F (F) | 5′-GTTCGATTCCGGAGAGGGA-3′ | [120] |
| 18S | 18S-bi (R) | 5′-GAGTCTCGTTCGTTATCGGA-3′ | [121] |
| 18S | 18S-a2.0 (F) | 5′-ATGGTTGCAAAGCTGAAAC-3′ | [121] |
| 18S | 18S-9R (R) | 5′-GATCCTTCCGCAGGTTCACCTAC-3′ | [120] |
| COI intron | AntonbruuniaCOIintron-205F | 5′-GGGGGTTTAACAGGGATTGTC-3′ | This Study |
| COI intron | AntonbruuniaCOIintron-683R | 5′-TTGTTTATGGAGTGTTAACCCTCT-3′ | This Study |
| COI intron | Synelmis-Intron3,011F | 5′-GGTTAGAACGAGTGCCGCTA-3′ | This Study |
| COI intron | Synelmis-Intron3,714R | 5′-CTCCTGCTCCGTATCACACC-3′ | This Study |
| COI intron | Synelmis-Intron4,990F | 5′-AGTAACCCTCGCTCCAAAGC-3′ | This Study |
| COI intron | Synelmis-Intron5,755R | 5′-CCAAGTGTGCCGAATGGTTC-3′ | This Study |
| Voucher | COI | 16S rRNA | 18S rRNA |
|---|---|---|---|
| A12280 | PolyLCO/PolyHCO | - | - |
| A12283 | PolyLCO/PolyHCO | - | - |
| A13955 | PolyLCO/PolyHCO | AnnF/16Sb | 18S-1F/18S-5R, 18S-3F/18S-bi, 18S-a2.0/18S-9R |
| A15568 | PolyLCO/PolyHCO | - | - |
| A16333 | PolyLCO/PolyHCO | AnnF/16Sb | - |
| A16381 | PolyLCO/PolyHCO | AnnF/16Sb | 18S-1F/18S-5R, 18S-3F/18S-bi, 18S-a2.0/18S-9R |
| A16382 | PolyLCO/PolyHCO | - | 18S-3F/18S-bi, 18S-a2.0/18S-9R |
| USNM1499416 | PolyLCO/PolyHCO | AnnF/16Sb | 18S-1F/18S-5R, 18S-3F/18S-bi, 18S-a2.0/18S-9R |
| USNM1643733 | - | AnnF/16Sb | 18S-1F/18S-5R, 18S-3F/18S-bi, 18S-a2.0/18S-9R |
| A6261 | - | 16SarL/16SbrH | - |
| A5771 | - | 16SarL/16SbrH | 18S-1F/18S-5R |
| A5926 | - | 16SarL/16SbrH | 18S-1F/18S-5R, 18S-3F/18S-bi |
| A4846 | - | - | 18S-1F/18S-5R |
| Primer Pair | Initial Denaturation | Cycles of Denaturation (Denaturation, Annealing, Elongation) | Final Extension | Max Amplified Base Pairs |
|---|---|---|---|---|
| polyLCO/polyHCO | 95 °C for 3 min | 40 (95 °C for 40 s, 42 °C for 45 s, 72 °C for 50 s) | 72 °C for 5 min | 684 |
| 16S-AnnF/16Sb | 94 °C for 2 min | 35 (94 °C for 40 s, 60 °C for 40 s, 70 °C for 45 s) | 72 °C for 7 min | 442 |
| 16SarL/16SbrH | 95 °C for 3 min | 35 (95 °C for 40 s, 50 °C for 40 s, 72 °C for 50 s) | 72 °C for 5 min | 530 |
| 18S-1F/18S-5R and 18S-a2.0/18S-9R | 95 °C for 3 min | 40 (95 °C for 30 s, 50 °C for 30 s, 72 °C for 90 s) | 72 °C for 8 min | 1792 |
| 18S-3F/18S-bi | 95 °C for 3 min | 40 (95 °C for 30 s, 52 °C for 30 s, 72 °C for 90 s) | 72 °C for 8 min | |
| AntonbruuniaCOIintron-205F/ AntonbruuniaCOIintron-683R | 95 °C for 3 min | 40 (95 °C for 40 s, 48 °C for 40 s, 72 °C for 50 s) | 72 °C for 5 min | 451 |
| Synelmis-Intron3,011F/ Synelmis-Intron3,714R | 95 °C for 3 min | 40 (95 °C for 40 s, 51 °C for 40 s, 72 °C for 50 s) | 72 °C for 5 min | 673 |
| Synelmis-Intron4,990F/ Synelmis-Intron5,755R | 95 °C for 3 min | 40 (95 °C for 40 s, 51 °C for 40 s, 72 °C for 50 s) | 72 °C for 5 min | 730 |
| Species | Total Number of Raw Reads (Paired-End) | Total Length of Raw Reads (Paired-End) | Individual Length of Raw Reads | Number of Reads Post-Trimmomatic (Paired-End) |
|---|---|---|---|---|
| Antonbruunia milenae sp. nov. | 3,702,263 | 555,339,450 | 150 | 3,662,081 (98.91%) |
| Ancistrosyllis sp. | 4,226,103 | 633,915,450 | 150 | 4,160,625 (98.45%) |
| Otopsis sp. | 10,035,386 | 1,505,307,900 | 150 | 9,912,137 (98.77%) |
| Sigambra sp. 1 | 8,748,561 | 1,312,284,150 | 150 | 8,609,906 (98.42%) |
| Sigambra sp. 2 | 9,106,118 | 1,505,307,900 | 150 | 8,996,276 (98.79%) |
| Sigambra sp. 3 | 6,241,943 | 936,291,450 | 150 | 6,134,740 (98.28%) |
| Glyphohesione klatti | 9,850,116 | 1,477,517,400 | 150 | 9,682,419 (98.30%) |
| Synelmis amoureuxi | 10,165,381 | 1,524,807,150 | 150 | 10,023,655 (98.61%) |
| Pilargis verrucosa | 8,696,621 | 1,304,493,150 | 150 | 8,479,066 (97.50%) |
| Pilargis wolfi | 7,213,725 | 1,082,058,750 | 150 | 7,062,447 (97.90%) |
| Hermundura fauveli | 7,387,747 | 1,108,162,050 | 150 | 7,253,809 (98.19%) |
| Micronephthys minuta | 6,284,087 | 942,613,050 | 150 | 6,159,575 (98.02%) |
| Struwela camposi | 8,715,828 | 1,307,374,200 | 150 | 8,631,491 (99.03%) |
| Lacydonia sp. | 8,319,473 | 1,247,920,950 | 150 | 8,213,908 (98.73%) |
| Phyllodoce medipapillata | 9,488,143 | 1,423,221,450 | 150 | 9,378,425 (98.84%) |
| Mitochondrial Genome Phylogeny Dataset | |||||
|---|---|---|---|---|---|
| Gene | Data Type | Substitution Model | Stationary Frequencies | Proportion of Invariant Sites | Rate Heterogeneity |
| ATP6 | AA | MTZOA | +F (empirical) | - | +GAMMA (mean) |
| ATP8 | AA | MTART | - | - | +GAMMA (mean) |
| COI | AA | MTZOA | - | +I (ML estimate) | +GAMMA (mean) |
| COII | AA | MTZOA | - | - | +GAMMA (mean) |
| COIII | AA | MTZOA | - | +I (ML estimate) | +GAMMA (mean) |
| CYTB | AA | MTZOA | - | +I (ML estimate) | +GAMMA (mean) |
| ND1 | AA | MTART | - | +I (ML estimate) | +GAMMA (mean) |
| ND2 | AA | MTMAM | +F (empirical) | - | +GAMMA (mean) |
| ND3 | AA | MTART | - | +I (ML estimate) | +GAMMA (mean) |
| ND4 | AA | MTZOA | +F (empirical) | - | +GAMMA (mean) |
| ND4L | AA | MTMAM | - | - | +GAMMA (mean) |
| ND5 | AA | MTART | +F (empirical) | +I (ML estimate) | +GAMMA (mean) |
| ND6 | AA | MTART | - | +I (ML estimate) | +GAMMA (mean) |
| 12S | NT | TIM2 | - | - | +GAMMA (mean) |
| 16S | NT | TN93 | - | +I (ML estimate) | +GAMMA (mean) |
| 18S | NT | TN93 | - | +I (ML estimate) | +GAMMA (mean) |
| Concatenated COI, 16S rRNA, and 18S rRNA Phylogeny Dataset | |||||
| Gene | Data Type | Substitution Model | Stationary Frequencies | Proportion of Invariant Sites | Rate Heterogeneity |
| COI | NT | GTR | - | +I (ML estimate) | +GAMMA (mean) |
| 16S | NT | TIM3 | - | +I (ML estimate) | +GAMMA (mean) |
| 18S | NT | TIM3 | - | +I (ML estimate) | +GAMMA (mean) |

Appendix B
















References
- Capa, M.; Hutchings, P. Annelid Diversity: Historical Overview and Future Perspectives. Diversity 2021, 13, 129. [Google Scholar] [CrossRef]
- Rouse, G.; Pleijel, F.; Tilic, E. Annelida; Oxford University Press: Oxford, UK, 2022; ISBN 0192591622. [Google Scholar]
- Watson, C.; Tilic, E.; Rouse, G.W. Revision of Hyalopale (Chrysopetalidae; Phyllodocida; Annelida): An Amphi-Atlantic Hyalopale bispinosa Species Complex and Five New Species from Reefs of the Caribbean Sea and Indo-Pacific Oceans. Zootaxa 2019, 4671, 339–368. [Google Scholar] [CrossRef]
- Pearson, K.A.M.; Rouse, G.W. Vampire Worms; A Revision of Galapagomystides (Phyllodocidae, Annelida), with the Description of Three New Species. Zootaxa 2022, 5128, 451–485. [Google Scholar] [CrossRef] [PubMed]
- Summers, M.M.; Rouse, G.W. Phylogeny of Myzostomida (Annelida) and Their Relationships with Echinoderm Hosts. BMC Evol. Biol. 2014, 14, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Tilic, E.; Stiller, J.; Campos, E.; Pleijel, F.; Rouse, G.W. Phylogenomics Resolves Ambiguous Relationships within Aciculata (Errantia, Annelida). Mol. Phylogenetics Evol. 2022, 166, 107339. [Google Scholar] [CrossRef] [PubMed]
- Mackie, A.S.Y.; Oliver, P.G.; Nygren, A. Antonbruunia sociabilis sp. nov. (Annelida: Antonbruunidae) Associated with the Chemosynthetic Deep-Sea Bivalve Thyasira scotiae Oliver & Drewery, 2014, and a Re-Examination of the Systematic Affinities of Antonbruunidae. Zootaxa 2015, 3995, 20–36. [Google Scholar] [CrossRef] [PubMed]
- Hartman, O.; Boss, K.J. Antonbruunia viridis, a New Inquiline Annelid with Dwarf Males, Inhabiting a New Species of Pelecypod, Lucina fosteri, in the Mozambique Channel. Ann. Mag. Nat. Hist. 1965, 8, 177–186. [Google Scholar] [CrossRef]
- Quiroga, E.; Sellanes, J. Two New Polychaete Species Living in the Mantle Cavity of Calyptogena gallardoi (Bivalvia: Vesicomyidae) at a Methane Seep Site off Central Chile (∼36° S). Sci. Mar. 2009, 73, 399–407. [Google Scholar] [CrossRef]
- Fauchald, K. The Polychaete Worms, Definitions and Keys to the Orders, Families and Genera; Science Series; Natural History Museum of Los Angeles County: Los Angeles, CA, USA, 1977; Volume 28, pp. 1–188. [Google Scholar]
- Glasby, C.J. Family Revision and Cladistic Analysis of the Nereidoidea (Polychaeta: Phyllodocida). Invertebr. Syst. 1993, 7, 1551–1573. [Google Scholar] [CrossRef]
- Salazar-Vallejo, S.I. Pilargidae (Annelida: Polychaeta) de Mexico: Lista de Especies, Nueva Especie y Biografia. Cah. Biol. Mar. 1986, 27, 192–210. [Google Scholar]
- Glasby, C.J.; Salazar-Vallejo, S.I. Pilargidae Saint-Joseph, 1899. In Handbook of Zoology. Annelida. Pleistoannelida, Errantia II, Phyllodocida; Purschke, G., Westheide, W., Böggemann, M., Eds.; De Gruyter: Berlin, Germany, 2022; pp. 308–320. [Google Scholar]
- Miura, T.; Laubier, L. Nautiliniellid Polychaetes Collected from the Hatsushima Cold-Seep Site in Sagami Bay, with Descriptions of New Genera and Species. Zool. Sci. 1990, 7, 319–325. [Google Scholar]
- Fitzhugh, K.; Wolf, P.S. Gross Morphology of the Brain of Pilargid Polychaetes: Taxonomic and Systematic Implications. Am. Mus. Novit. 1990, 2992, 16–23. [Google Scholar]
- Beesley, P.L.; Ross, G.J.B.; Glasby, C.J. (Eds.) Polychaetes & Allies: The Southern Synthesis; CSIRO Publishing: Clayton, VIC, Australia, 2000; Volume 4. [Google Scholar]
- Pleijel, F.; Dahlgren, T. Position and Delineation of Chrysopetalidae and Hesionidae (Annelida, Polychaeta, Phyllodocida). Cladistics 1998, 14, 129–150. [Google Scholar] [CrossRef] [PubMed]
- Rouse, G.W.; Pleijel, F. Polychaetes; Oxford University Press: Oxford, UK, 2001; ISBN 0198506082. [Google Scholar]
- Kükenthal, W. Annelida: Volume 1. Annelida Basal Groups and Pleistoannelida, Sedentaria I; Purschke, G., Westheide, W., Böggemann, M., Eds.; Walter de Gruyter GmbH & Co.: Berlin, Germany, 2019; ISBN 3110291460. [Google Scholar]
- Ravara, A.; Wiklund, H.; Cunha, M.R.; Pleijel, F. Phylogenetic Relationships within Nephtyidae (Polychaeta, Annelida). Zool. Scr. 2010, 39, 394–405. [Google Scholar] [CrossRef]
- Bernardino, A.F.; Li, Y.; Smith, C.R.; Halanych, K.M. Multiple Introns in a Deep-Sea Annelid (Decemunciger: Ampharetidae) Mitochondrial Genome. Sci. Rep. 2017, 7, 4295. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Liu, X. HZPLY558-13.COI-5P. Available online: http://www.boldsystems.org/index.php/Public_RecordView?processid=HZPLY558-13 (accessed on 20 December 2023).
- Böggemann, M. Polychaetes (Annelida) of the Abyssal SE Atlantic. Org. Divers. Evol. 2009, 9, 251–428. [Google Scholar] [CrossRef]
- Vijapure, T.; Sukumaran, S.; Manohar, C.S. Molecular Characterization and Phylogenetics of Indian Polychaete Fauna: Scope for Implementation in Ecological Monitoring. Aquat. Ecol. 2019, 53, 665–677. [Google Scholar] [CrossRef]
- Aylagas, E.; Borja, Á.; Irigoien, X.; Rodríguez-Ezpeleta, N. Benchmarking DNA Metabarcoding for Biodiversity-Based Monitoring and Assessment. Front. Mar. Sci. 2016, 3, 96. [Google Scholar] [CrossRef]
- Struck, T.H.; Schult, N.; Kusen, T.; Hickman, E.; Bleidorn, C.; McHugh, D.; Halanych, K.M. Annelid Phylogeny and the Status of Sipuncula and Echiura. BMC Evol. Biol. 2007, 7, 57. [Google Scholar] [CrossRef]
- Paulay, G.; O’Mahoney, M.J.V. BBPS179-19.COI-5P. Available online: http://www.boldsystems.org/index.php/Public_RecordView?processid=BBPS179-19 (accessed on 20 December 2023).
- Alves, P.R.; Halanych, K.M.; Silva, E.P.; Santos, C.S.G. Nereididae (Annelida) Phylogeny Based on Molecular Data. Org. Divers. Evol. 2023, 23, 529–541. [Google Scholar] [CrossRef]
- Cejp, B.; Ravara, A.; Aguado, M.T. First Mitochondrial Genomes of Chrysopetalidae (Annelida) from Shallow-Water and Deep-Sea Chemosynthetic Environments. Gene 2022, 815, 146159. [Google Scholar] [CrossRef] [PubMed]
- Alves, P.R.; Halanych, K.M.; Santos, C.S.G. The Phylogeny of Nereididae (Annelida) Based on Mitochondrial Genomes. Zool. Scr. 2020, 49, 366–378. [Google Scholar] [CrossRef]
- Villalobos-Guerrero, T.F.; Huč, S.; Tilic, E.; Hiley, A.S.; Rouse, G.W. A Remarkable New Deep-Sea Nereidid (Annelida: Nereididae) with Gills. PLoS ONE 2024, in press. [Google Scholar]
- Li, X.; Yang, D.; Qiu, J.-W.; Liu, P.; Meng, D.; Zhu, H.; Guo, L.; Luo, S.; Wang, Z.; Ke, C. Mitochondrial Genome of Leocrates chinensis Kinberg, 1866 (Annelida: Hesionidae). Mitochondrial DNA Part B 2023, 8, 172–176. [Google Scholar] [CrossRef] [PubMed]
- Ruta, C.; Nygren, A.; Rousset, V.; Sundberg, P.; Tillier, A.; Wiklund, H.; Pleijel, F. Phylogeny of Hesionidae (Aciculata, Polychaeta), Assessed from Morphology, 18S RDNA, 28S RDNA, 16S RDNA and COI. Zool. Scr. 2007, 36, 99–107. [Google Scholar] [CrossRef]
- Richter, S.; Schwarz, F.; Hering, L.; Böggemann, M.; Bleidorn, C. The Utility of Genome Skimming for Phylogenomic Analyses as Demonstrated for Glycerid Relationships (Annelida, Glyceridae). Genome Biol. Evol. 2015, 7, 3443–3462. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, M.; Liu, H.; Li, B.; Guo, L.; Meng, Z.; Lin, H. The Complete Mitochondrial Genome of the Polychaete, Goniada japonica (Phyllodocida, Goniadidae). Mitochondrial DNA Part A 2016, 27, 2850–2851. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An Integrated and Extendable Desktop Software Platform for the Organization and Analysis of Sequence Data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Allio, R.; Schomaker-Bastos, A.; Romiguier, J.; Prosdocimi, F.; Nabholz, B.; Delsuc, F. MitoFinder: Efficient Automated Large-Scale Extraction of Mitogenomic Data in Target Enrichment Phylogenomics. Mol. Ecol. Resour. 2020, 20, 892–905. [Google Scholar] [CrossRef]
- Li, D.; Luo, R.; Liu, C.M.; Leung, C.M.; Ting, H.F.; Sadakane, K.; Yamashita, H.; Lam, T.W. MEGAHIT v1.0: A Fast and Scalable Metagenome Assembler Driven by Advanced Methodologies and Community Practices. Methods 2016, 102, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Laslett, D.; Canbäck, B. ARWEN: A Program to Detect TRNA Genes in Metazoan Mitochondrial Nucleotide Sequences. Bioinformatics 2008, 24, 172–175. [Google Scholar] [CrossRef]
- Dierckxsens, N.; Mardulyn, P.; Smits, G. NOVOPlasty: De Novo Assembly of Organelle Genomes from Whole Genome Data. Nucleic Acids Res. 2017, 45, e18. [Google Scholar] [CrossRef] [PubMed]
- Meng, G.; Li, Y.; Yang, C.; Liu, S. MitoZ: A Toolkit for Animal Mitochondrial Genome Assembly, Annotation and Visualization. Nucleic Acids Res. 2019, 47, e63. [Google Scholar] [CrossRef]
- Bernt, M.; Donath, A.; Jühling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Pütz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de Novo Metazoan Mitochondrial Genome Annotation. Mol. Phylogenetics Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef]
- Hahn, C.; Bachmann, L.; Chevreux, B. Reconstructing Mitochondrial Genomes Directly from Genomic Next-Generation Sequencing Reads—A Baiting and Iterative Mapping Approach. Nucleic Acids Res. 2013, 41, e129. [Google Scholar] [CrossRef]
- Li, H. Minimap2: Pairwise Alignment for Nucleotide Sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef]
- Li, H. New Strategies to Improve Minimap2 Alignment Accuracy. Bioinformatics 2021, 37, 4572–4574. [Google Scholar] [CrossRef]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve Years of SAMtools and BCFtools. Gigascience 2021, 10, giab008. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New Capabilities and Interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Swofford, D. PAUP*; Version 4.0b10; Phylogenetic Analysis Using Parsimony (*and Other Methods); Oxford University Press: Oxford, UK, 2002. [Google Scholar]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. Version 3.61. Available online: http://www.mesquiteproject.org (accessed on 26 December 2023).
- Katoh, K.; Misawa, K.; Kuma, K.I.; Miyata, T. MAFFT: A Novel Method for Rapid Multiple Sequence Alignment Based on Fast Fourier Transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Edgar, R.C. Quality Measures for Protein Alignment Benchmarks. Nucleic Acids Res. 2010, 38, 2145–2153. [Google Scholar] [CrossRef]
- Edler, D.; Klein, J.; Antonelli, A.; Silvestro, D. RaxmlGUI 2.0: A Graphical Interface and Toolkit for Phylogenetic Analyses Using RAxML. Methods Ecol. Evol. 2020, 12, 373–377. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 11 March 2021).
- Leigh, J.W.; Bryant, D. POPART: Full-Feature Software for Haplotype Network Construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Bernt, M.; Merkle, D.; Middendorf, M. An Algorithm for Inferring Mitogenome Rearrangements in a Phylogenetic Tree. In RECOMB International Workshop on Comparative Genomics, Proceedings of the Comparative Genomics, Paris, France, 13–15 October 2008; Nelson, C.E., Vialette, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 143–157. [Google Scholar]
- Bernt, M.; Merkle, D.; Ramsch, K.; Fritzsch, G.; Perseke, M.; Bernhard, D.; Schlegel, M.; Stadler, P.F.; Middendorf, M. CREx: Inferring Genomic Rearrangements Based on Common Intervals. Bioinformatics 2007, 23, 2957–2958. [Google Scholar] [CrossRef]
- von Haeseler, A.; Schmidt, H.A.; Bui, M.Q.; Nguyen, L.T. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2014, 32, 268–274. [Google Scholar]
- Chernomor, O.; von Haeseler, A.; Minh, B.Q. Terrace Aware Data Structure for Phylogenomic Inference from Supermatrices. Syst. Biol. 2016, 65, 997–1008. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Shimodaira, H. An Approximately Unbiased Test of Phylogenetic Tree Selection. Syst. Biol. 2002, 51, 492–508. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, R.P.; Barbosa, A.D.C.; Freitas, R.; Zanol, J.; Glasby, C.J.; Ruta, C. Pilargidae (Annelida: Phyllodocida) from Coastal and Deep Waters of the Southwestern Atlantic, with Descriptions of Two New Species. Zootaxa 2020, 4878, 056–076. [Google Scholar] [CrossRef]
- Hartman, O. Polychaetous Annelids, Part VIII: Pilargiidae. In Allan Hancock Pacific Expeditions; University of Southern California Press: Los Angeles, CA, USA, 1947; Volume 10, pp. 483–523. [Google Scholar]
- Glasby, C.J.; Hocknull, S.A. New Records and a New Species of Hermundura Müller, 1858, the Senior Synonym of Loandalia Monro, 1936 (Annelida: Phyllodocida: Pilargidae) from Northern Australia and New Guinea. Beagle Rec. Mus. Art Gall. N. Territ. 2010, 26, 57–67. [Google Scholar]
- Salazar-Vallejo, S.; Bailey Brock, J.H.; Dreyer, J.C. Revision of Pseudexogone Augener, 1922 (Annelida, Polychaeta, Syllidae), and Its Transfer to Pilargidae. Zoosystema 2007, 29, 535–553. [Google Scholar]
- Vallès, Y.; Halanych, K.M.; Boore, J.L. Group II Introns Break New Boundaries: Presence in a Bilaterian’s Genome. PLoS ONE 2008, 3, e1488. [Google Scholar] [CrossRef] [PubMed]
- Saint-Joseph, A. d’Anthoine de Note Sur Une Nouvelle Famille d’Annélides Polychètes. Bull. Muséum D’histoire Nat. Paris 1899, 5, 41–42. [Google Scholar]
- Webster, H.E. The Annelida Chaetopoda of the Virginian Coast. Trans. Albany Inst. 1879, 9, 202–269. [Google Scholar]
- McIntosh, W.C. On the Annelida Obtained during the Cruise of H.M.S. ‘Valorous’ to Davis Strait in 1875. Trans. Linn. Soc. London. Second. Ser. Zool. 1878, 1, 499–511. [Google Scholar] [CrossRef]
- Misra, A. Polychaete Fauna of West Bengal. In Zoological Survey of India; Gosh, A.K., Ed.; Director: Calcutta, India, 1999; Volume 10, pp. 125–225. [Google Scholar]
- Ditlevsen, H.; Annelids, I. The Danish Ingolf Expedition; Nature Publishing Group: Copenhagen, Denmark, 1917; Volume 4, pp. 1–71. [Google Scholar]
- Friedrich, H. Zwei Neue Bestandteile in Der Fauna Der Nordsee. Zool. Anz. 1950, 145, 171–177. [Google Scholar]
- Müller, F. Einiges Über Die Annelidenfauna Der Insel Santa Catharina an Der Brasilianischen Küste. Arch. Naturgeschichte 1858, 24, 211–220. [Google Scholar]
- Pearson, T.H. Litocorsa stremma a New Genus and Species of Pilargid (Polychaeta: Annelida) from the West Coast of Scotland, with Notes on Two Other Pilargid Species. J. Nat. Hist. 1970, 4, 69–77. [Google Scholar] [CrossRef]
- Augener, H. Litorale Polychaeten von Juan Fernandez. In The Natural History of Juan Fernandez and Easter Island; Creative Media Partners, LLC: Sheridan, WY, USA, 1922; Volume 3.2, pp. 161–218. [Google Scholar]
- Chamberlin, R.V. The Annelida Polychaeta; The Museum of Comparative Zoology at Harvard College: Cambridge, MA, USA, 1919; Volume 48. [Google Scholar]
- Fauvel, P. Annelida Polychaeta of the Indian Museum, Calcutta. In Memoirs of the Indian Museum; Manager of Publications: Calcutta, India, 1932; Volume 12, pp. 1–261. [Google Scholar]
- Salazar-Vallejo, S.I.; Nishi, E.; Anguspanich, S. Rediscovery of Talehsapia annandalei (Polychaeta: Pilargidae) in Songkhla Lagoon, Thailand. Pac. Sci. 2001, 55, 267–273. [Google Scholar] [CrossRef]
- Pettibone, M.H. Revision of the Pilargidae (Annelida: Polychaeta), Including Descriptions of New Species, and Redescriptions of the Pelagic Podarmus ploa Chamberlain (Polynoidae). In Proceedings of the United States National Museum; Smithsonian Institution Press: Washington, DC, USA, 1966; Volume 118, pp. 155–207. [Google Scholar]
- Mandal, S.; Harkantra, S.N.; Salazar-Vallejo, S.I. Cabira rangarajani n. sp. (Polychaeta: Pilargidae) from the Goa Coast, Central West Coast of India. Zootaxa 2007, 1446, 21–29. [Google Scholar] [CrossRef]
- Plathong, J.; Dean, H.K.; Plathong, S. Four New Species of Pilargidae (Annelida: Pilarginae) from the Gulf of Thailand. Zootaxa 2021, 5071, 537–562. [Google Scholar] [CrossRef]
- Licher, F. Resurrection of Glyphohesione Friedrich, 1950, with Redescription of G. klatti Friedrich, 1950 and Description of G. longocirrata (Polychaeta: Hesionidae). Proc. Biol. Soc. Wash. 1994, 107, 600–608. [Google Scholar]
- Salazar-Vallejo, S.I. Revision of Synelmis Chamberlin, 1919 (Annelida, Polychaeta, Pilargidae). Zoosystema 2003, 25, 17–42. [Google Scholar]
- Ehlers, E. Polychaeten von Java Und Amboina. Ein Beitrag Zur Kenntnis Der Malaiischen Strandfauna. In Abhandlungen der koeniglichen Gesellschaft der Wissenschaften zu Goettingen, Ser. neue folge; Forgotten Books: London, UK, 1920; Volume 10, pp. 1–73. [Google Scholar]
- Aguado, M.T. Antonbruunidae Fauchald, 1977. In Handbook of Zoology. Annelida Vol. 4: Pleistoannelida, Errantia II, Phyllodocida; Purschke, G., Boggemann, M., Westheide, W., Eds.; De Gruyter: Berlin, Germany, 2022; pp. 394–399. [Google Scholar]
- Katzmann, W.; Laubier, L.; Ramos, J. Pilargidae (Annélides Polychètes Errantes) de Méditerranée. Bull. L’institute Océanographique 1974, 71, 1–40. [Google Scholar]
- Uschakov, P. V A New Deep-Water Species of Otopsis (Polychaeta, Pilargidae) from the Kurile-Kamtchatka Trench. Zool. Zhurnal 1971, 50, 279–281. [Google Scholar]
- Salazar-Vallejo, S.I.; De León-González, J.A.; Carrera-Parra, L.F. Phylogeny of Microphthalminae Hartmann-Schröder, 1971, and Revision of Hesionella Hartman, 1939, and Struwela Hartmann-Schröder, 1959 (Annelida, Errantia). PeerJ 2019, 7, e7723. [Google Scholar] [CrossRef]
- Darbyshire, T.; Mackie, A.S.Y. Species of Litocorsa (Polychaeta: Pilargidae) from the Indian Ocean and South China Sea. Hydrobiologia 2003, 496, 63–73. [Google Scholar] [CrossRef]
- Monro, C.C.A. Polychaete Worms II; Discovery Reports; Cambridge University Press: Cambridge, UK, 1936; Volume 12, pp. 59–197. [Google Scholar]
- Marks, S.; Hocknull, S. New Species of Loandalia (Polychaeta: Pilargidae) from Queensland, Australia. Zootaxa 2006, 1119, 59–68. [Google Scholar] [CrossRef]
- Hartmann-Schröder, G. Zur Ökologie Der Polychaeten Des Mangrove-Estero-Gebietes von El Salvador. Beitr. Neotropischen Fauna 1959, 1, 69–183. [Google Scholar] [CrossRef]
- Emerson, R.R.; Fauchald, K. A Revision of the Genus Loandalia Monro with Description of a New Genus and Species of Pilargiid Polychaete. Bull. South. Calif. Acad. Sci. 1971, 70, 18–22. [Google Scholar]
- Solis-Weiss, V. Parandalia bennei (Pilargidae) and Spiophanes lowai (Spionidae), New Species of Polychaetous Annelids from Mazatlan Bay, Pacific Coast of Mexico. Proc. Biol. Soc. Wash. 1983, 96, 370–378. [Google Scholar]
- Berkeley, E.; Berkeley, C. On a Collection of Polychaeta from Southern California. Bull. South. Calif. Acad. Sci. 1941, 40, 16–60. [Google Scholar]
- Thomas, P.J. Polychaetous Worms from the Arabian Sea, 1: A New Species of the Genus Loandalia Monro. Bull. Dep. Mar. Biol. Oceanogr. 1963, 1, 29–34. [Google Scholar]
- Intes, A.; Le Loeuff, P. Les Annélides Polychètes de Côte d’Ivoire. I. Polychètes Errantes—Compte Rendu Systématique. Cah. ORSTOM Série Océanographie 1975, 13, 267–321. [Google Scholar]
- León-González, J.A. de Poliquetos de Fondos Blandos de La Costa Occidental de Baja California Sur, Mexico. I. Pilargidae. Cah. Biol. Mar. 1991, 32, 311–321. [Google Scholar]
- Salazar-Vallejo, S.I.; Reyes-Barragán, M.P. Parandalia vivianneae n. sp. and P. tricuspis (Müller), Two Estuarine Polychaetes (Polychaeta: Pilargidae) from Eastern Mexico. Rev. Biol. Trop. 1990, 38, 87–90. [Google Scholar]
- Struck, T.H.; Golombek, A.; Hoesel, C.; Dimitrov, D.; Elgetany, A.H. Mitochondrial Genome Evolution in Annelida—A Systematic Study on Conservative and Variable Gene Orders and the Factors Influencing Its Evolution. Syst. Biol. 2023, 72, 925–945. [Google Scholar] [CrossRef]
- Britayev, T.A.; Zamishliak, E.A. Association of the Commensal Scaleworm Gastrolepidia clavigera (Polychaeta: Polynoidae) with Holothurians near the Coast of South Vietnam. Ophelia 1996, 45, 175–190. [Google Scholar] [CrossRef]
- Ruff, R.E. A New Species of Bathynoe (Polychaeta: Polynoidae) from the Northeast Pacific Ocean Commensal with Two Species of Deep-Water Asteroids. Ophelia Suppl. 1991, 5, 219–230. [Google Scholar]
- Martin, D.; Rosell, D.; Uriz, M.J. Harmothoe hyalonemae sp. nov. (Polychaeta, Polynoidae), an Exclusive Inhabitant of Different Atlanto-Mediterranean Species of Hyalonema (Porifera, Hexactinellida). Ophelia 1992, 35, 169–185. [Google Scholar] [CrossRef]
- Tilic, E.; Rouse, G.W. Hardly Venus’s Servant—Morphological Adaptations of Veneriserva to an Endoparasitic Lifestyle and Its Phylogenetic Position within Dorvilleidae (Annelida). Org. Divers. Evol. 2024. [Google Scholar] [CrossRef]
- Dimock, R.V.J. Intraspecific Agression and the Distribution of a Symbiotic Polychaete on Its Hosts. In Proceedings of the Symbiosis in the Sea; Vernberg, W.B., Ed.; University or South Carolina Press: Columbia, SC, USA, 1974; pp. 29–44. [Google Scholar]
- Pernet, B.; Jaeckle, W.B. Size and Organic Content of Eggs of Marine Annelids, and the Underestimation of Egg Energy Content by Dichromate Oxidation. Biol. Bull. 2004, 207, 67–71. [Google Scholar] [CrossRef] [PubMed]
- McHugh, D.; Fong, P.P. Do Life History Traits Account for Diversity of Polychaete Annelids? Invertebr. Biol. 2002, 121, 325–338. [Google Scholar] [CrossRef]
- Rouse, G.W.; Scripps Institution of Oceanography, UCSD, La Jolla, CA, USA. Personal Observation, 2020.
- Dall, W.H. On Some New or Interesting West American Shells Obtained from the Dredgings of the U.s. Fish Commission Steamer Albatross in 1888, and from Other Sources. In Proceedings of the United States National Museum; Smithsonian Institution Press; Washington, DC, USA, 1891; Volume 14, pp. 173–191. [Google Scholar]
- Suzuki, K. Three New Species of Non-Marine Shells from the Tertiary Formations of Hokkaidō and Karahuto. Jpn. J. Geol. Geogr. 1941, 18, 53–58. [Google Scholar]
- Taboada, S.; Silva, A.S.; Neal, L.; Cristobo, J.; Ríos, P.; Álvarez-Campos, P.; Hestetun, J.T.; Koutsouveli, V.; Sherlock, E.; Riesgo, A. Insights into the Symbiotic Relationship between Scale Worms and Carnivorous Sponges (Cladorhizidae, Chondrocladia). Deep. Sea Res. Part I Oceanogr. Res. Pap. 2020, 156, 103191. [Google Scholar] [CrossRef]
- Eeckhaut, I.; Fievez, L.; Müller, M.C.M. Larval Development of Myzostoma cirriferum (Myzostomida). J. Morphol. 2003, 258, 269–283. [Google Scholar] [CrossRef]
- Rouse, G.W.; Grygier, M.J. Myzostoma seymourcollegiorum n. sp. (Myzostomida) from Southern Australia, with a Description of Its Larval Development. Zootaxa 2005, 1010, 53–64. [Google Scholar] [CrossRef]
- Salazar-Vallejo, S.I.; Harris, L.H. Revision of Pilargis de Saint-Joseph, 1899 (Annelida, Polychaeta, Pilargidae). J. Nat. Hist. 2006, 40, 119–159. [Google Scholar] [CrossRef]
- Carr, C.M.; Hardy, S.M.; Brown, T.M.; Macdonald, T.A.; Hebert, P.D.N. A Tri-Oceanic Perspective: DNA Barcoding Reveals Geographic Structure and Cryptic Diversity in Canadian Polychaetes. PLoS ONE 2011, 6, e22232. [Google Scholar] [CrossRef] [PubMed]
- Sjölin, E.; Erséus, C.; Källersjö, M. Phylogeny of Tubificidae (Annelida, Clitellata) Based on Mitochondrial and Nuclear Sequence Data. Mol. Phylogenetics Evol. 2005, 35, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Palumbi, S.R. Nucleic Acids II: The Polymerase Chain Reaction. In Molecular Systematics, 2nd ed.; Sinauer: Sunderland, MA, USA, 1996; pp. 205–247. [Google Scholar]
- Palumbi, S.R.; Martin, A.; Romano, S.; McMillan, W.O.; Stice, L.; Grabowski, G. The Simple Fool’s Guide to PCR; University of Hawaii: Honolulu, HI, USA, 1991. [Google Scholar]
- Giribet, G.; Carranza, S.; Baguna, J.; Riutort, M.; Ribera, C. First Molecular Evidence for the Existence of a Tardigrada + Arthropoda Clade. Mol. Biol. Evol. 1996, 13, 76–84. [Google Scholar] [CrossRef]
- Whiting, M.F.; Carpenter, J.C.; Wheeler, Q.D.; Ard, V. The Strepsiptera Problem: Phylogeny of the Holometabolous Insect Orders Inferred from 18S and 28S Ribosomal DNA Sequences and Morphology. Syst. Biol. 1997, 46, 1–68. [Google Scholar] [CrossRef]











| Species | Voucher | Locality | Depth (m) | COI | 16S rRNA | 18S rRNA |
|---|---|---|---|---|---|---|
| Micronephthys minuta | SIO-BIC A15537 | Disko Island, Greenland (69.2901, −53.2143) | 190 | OR126034 | OR159884 | OR159893 |
| Nephtys hombergii | MB36000118 [20] | Ria Aveiro, Portugal (40.642, −8.754) | - | GU179410 | - | GU179377 |
| MB36000136 [20] | Bohuslän, Sweden (58.217, 10.595) | - | - | GU179354 | - | |
| Nephtys sp. | AB-2017 [21] | NE Pacific (48.5, −127.0) | - | KY972420 | - | - |
| Antonbruunia milenae sp. nov. | SIO-BIC: A12278, | San Diego, CA, USA (32.7769, −117.4881) | 845 | OR126047 | - | - |
| A12279, | OR126043 | OR159883 | OR148950 | |||
| A12280, | OR126045 | - | - | |||
| A12281 *, | OR126046 * | - | - | |||
| A12283 | OR126044 | - | - | |||
| Antonbruunia sociabilis | NMW<GBR>:Z.2012.074.13 [7] | Scotland (57.9500, −15.5500) | 1200 | - | KP992845 | KP992843 |
| Pilargis verrucosa | SIO-BIC A13955 | Koster, Sweden (58.895, 10.989) | - | OR126029 | OR159877 | OR159892 |
| Pilargis wolfi | USNM1499416 | Virginia, USA (37.5848, −75.6486) | - | USNM1499416 | OR159878 | OR159891 |
| Pilargis cf. cholae | SIO-BIC A4846 | Carrie Bow Cay, Belize (16.8134, −88.0817) | 1 | OR126031 | OR159876 | OR159890 |
| Cabira pilargiformis | BIOUG06647-G01 [22] | Dongying, China (37.948, 119.361) | 16 | HZPLY558-13 | - | - |
| Sigambra magnuncus | [23] | W Africa (−5.34, 3.68) | GQ426642 | GQ426621 | GQ426593 | |
| Sigambra hanaokai | SIO-BIC: A5771, A15561 | Tanabe Bay, Japan (33.6997, 135.3268) | 32–36 | OR126015, OR126016 | OR159880 - | OR159886 - |
| Sigambra parva | Vd4(c)-Pil [24] | NW India (22.3000, 69.3900) | - | KY775644 | - | - |
| Sigambra sp. 1 | SIO-BIC: A1516, | W Costa Rica (8.9308, −84.3072) | 967–1005 | OR126027, | OR123440 | PP034425 |
| A9597, | W Costa Rica (9.1174, −84.8396) | 1795 | OR126028, | - | - | |
| A9843 | W Costa Rica (8.9309, −84.3133) | 999 | OR126026 | - | - | |
| Sigambra sp. 2 | SIO-BIC A10175 | W Costa Rica (7.6802, −85.9118) | 528.2 | OR126014 | OR123444 | PP034426 |
| Sigambra sp. 3 | SIO-BIC: A15568, | Rovinj, Croatia (45.0915, 13.6244) | 25 | OR126019, | OR159881 | OR159887 |
| A15569, | OR126020, | - | - | |||
| A15570, | OR126021, | - | - | |||
| A15571, | OR126018, | - | - | |||
| A15572, | OR126023, | - | - | |||
| A15573 | OR126022, | - | - | |||
| A5926 | Banyuls-sur-Mer, France (42.4958, 3.1400) | 24 | OR126017 | OR159882 | OR159888 | |
| Sigambra sp. 4 | A16333 | Hong Kong, China (22.4692, 114.2117) | 1 | PP050561 | PP034423 | - |
| Litocorsa stremma | Itsastk13-P23 [25] | N Spain (44.75, −4.96) | - | KT307655 | - | - |
| Ancistrosyllis sp. | SIO-BIC A12326 | Santa Monica, CA, USA (33.8385, −118.6904) | 711 | OR126024 | OR123445 | PP034427 |
| Ancistrosyllis breviceps | SIO-BIC A6261 | Santa Barbara, CA, USA (34.3700, −120.0300) | 400 | OR126025 | OR159879 | OR159889 |
| Ancistrosyllis groenlandica | Itsastk13-P43 [25], | N Spain, (44.75, −4.96) | - | KT307624 | - | - |
| [26] | - | - | - | - | DQ790075 | |
| Glyphohesione klatti | SIO-BIC A6066 | N Trondheim, Norway (63.5086, 10.4419) | 35 | OR126035 | OR123443 | PP034428 |
| Otopsis sp. | SIO-BIC: A15574, | Monterey, CA, USA (36.7721, −122.0831) | 2892–2937 | OR126036, | OR123441 | PP034429 |
| A15575, | OR126037, | - | - | |||
| A15576, | OR126039, | - | - | |||
| A15577, | OR126042, | - | - | |||
| A15578, | OR126040, | - | - | |||
| A15579, | OR126041, | - | - | |||
| A15580 | OR126038 | - | - | |||
| Synelmis amoureuxi | SIO-BIC A4721 | Carrie Bow Cay, Belize (16.8032, −88.0778) | 10 | OR126033 | OR123442 | PP034430 |
| Pseudexogone cf. backstromi | SIO-BIC A2813 | S Pacific (−17.4904, −129.8267) | 18–20 | OR126032 | OR159875 | OR159895 |
| Hermundura americana | A16381 A16382 | Pagan River, Chesapeake Bay, VA, USA | - - | PP050562 PP050563 | PP034424 - | PP033026 PP033027 |
| Hermundura fauveli | USNM1643733 [27] | Bremerton, WA, USA (47.5471, −122.6620) | - | BBPS179-19 | OR159885 | OR159894 |
| Species | Voucher | Locality | Depth (m) | 18S rRNA | Mitochondrial Genome |
|---|---|---|---|---|---|
| Micronephthys minuta | SIO-BIC A15537 | Disko Island, Greenland (69.2901, −53.2143) | 190 | OR159893 | OR123448 |
| Nephtys sp. | AB-2017 [21] | NE Pacific (48.5, −127.0) | - | - | KY972376, KY972406, KY972433, KY972447, KY972462, KY972476, KY972490, KY972504, KY972532 |
| Antonbruunia milenae sp. nov. | SIO-BIC A12279 | San Diego, CA, USA (32.7769, −117.4881) | 845 | OR148950 | OR123446 |
| Pilargis verrucosa | SIO-BIC A13955 | Koster, Sweden (58.895, 10.989) | - | OR159892 | OR123439 |
| Pilargis wolfi | USNM1499416 | Virginia, USA (37.5848, −75.6486) | - | OR159891 | OR123447 |
| Sigambra sp. 1 | SIO-BIC A1516 | W Costa Rica (8.9308, −84.3072) | 967–1005 | PP034425 | OR123440 |
| Sigambra sp. 2 | SIO-BIC A10175 | W Costa Rica (7.6802, −85.9118) | 528.2 | PP034426 | OR123444 |
| Sigambra sp. 3 | SIO-BIC A15568 | Rovinj, Croatia (45.0915, 13.6244) | 25 | OR159887 | PP035855 |
| Ancistrosyllis sp. | SIO-BIC A12326 | Santa Monica, CA, USA (33.8385, −118.6904) | 711 | PP034427 | OR123445 |
| Glyphohesione klatti | SIO-BIC A6066 | N Trondheim, Norway (63.5086, 10.4419) | 35 | PP034428 | OR123443 |
| Otopsis sp. | SIO-BIC A15574 | Monterey, CA, USA (36.7721, −122.0831) | 2892–2937 | PP034429 | OR123441 |
| Synelmis amoureuxi | SIO-BIC A4721 | Carrie Bow Cay, Belize (16.8032, −88.0778) | 10 | PP034430 | OR123442 |
| Lacydonia sp. | SIO-BIC A2650 | Hydrate Ridge, OR, USA (44.6701, −125.0987) | 587–610 | PP034431 | PP035856 |
| Phyllodoce medipapillata | SIO-BIC A4340 | La Jolla, CA, USA (32.8145, −117.2740) | 1 | PP034432 | PP035857 |
| Struwela camposi | SIO-BIC A13437 | San Felipe, Mexico (31.0700, −114.8400) | 1 | PP033480 | PP035858 |
| Hermundura fauveli | USNM1643733 | Bremerton, WA, USA (47.5471, −122.6620) | - | OR159894 | OR124844 |
| Microphthalmus listensis | 85792 [28] | List, Germany (55.0156, 8.4379) | - | - | MN855111, MN855121, MN855130, MN855139, MN855149, MN855158, MN855175, MN855183, MN855191, |
| 76292 [28] | List, Germany (55.0232, 8.4401) | - | - | MN855200, MN855210 MN855206 | |
| Microphthalmus similis | 76290 | List, Germany (55.0232, 8.4401) | - | - | MN855112, MN855122, MN855131, MN855140, MN855150, MN855160, MN855168, MN855184, MN855192, MN855201 |
| Chrysopetalum debile | [29] | Mallorca, Spain | 1 | MZ450125 | MZ506670 |
| Arichlidon gathofi | AG01 [28] | - | - | - | OQ732695 |
| 13915 [30] | NC, USA (33.9993, −76.7026) | - | - | MN855107, MN855116, MN855126, MN855135, MN855144, MN855154, MN855164, MN855171, MN855178, | |
| 18126 [30] | Bocas del Toro, Panama (9.3339, −82.2205) | - | - | MN855188, MN855196 MN855127, MN855197, MN855205 | |
| Bhawania goodei | 27046 [30] | Bocas del Toro, Panama (9.3667, −82.3000) | - | - | MN855118, MN855128, MN855146, MN855156, MN855166, MN855173, MN855180, MN855189, MN855198, MN855208 |
| Nereis pelagica | SIO-BIC A6054 [31] | Trondheim, Norway (63.4410, 10.3482) | 0 | OR437939 | OL782598 |
| Nectoneanthes oxypoda | SIO-BIC A13109 [31] | Osaka Bay, Japan (34.3300, 135.1400) | - | OR437940 | OL782599 |
| Oxydromus pugettensis | 23185 [30] | Snug Harbor, WA, USA (48.5755, −123.1699) | - | - | MN855113, MN855123, MN855132, MN855141, MN855151, MN855161, MN855176, MN855185, MN855193, MN855202, MN855211 |
| [26] | - | - | DQ790086 | - | |
| Leocrates chinensis | XMU-Pol-2021-354 [32] | Hong Kong, China (22.2360, 114.1852) | 9.1 | - | NC066969 |
| SMNH83510 [33] | Lifou, New Caledonia | - | DQ442589 | - | |
| Glycinde armigera | FS17 [34] | Bellingham Bay, WA, USA | - | KT989339 | KT989325 |
| Goniada japonica | [35] | Hailing Island, China | - | - | KP867019 |
| Hemipodia simplex | FS13 [34] | Bamfield, Canada | - | KT989342 | KT989322 |
| Glycera dibranchiata | FS05 [34] | Loagy Bay, MA, USA | - | KT989350 | KT989318 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huč, S.; Hiley, A.S.; McCowin, M.F.; Rouse, G.W. A Mitogenome-Based Phylogeny of Pilargidae (Phyllodocida, Polychaeta, Annelida) and Evaluation of the Position of Antonbruunia. Diversity 2024, 16, 134. https://doi.org/10.3390/d16030134
Huč S, Hiley AS, McCowin MF, Rouse GW. A Mitogenome-Based Phylogeny of Pilargidae (Phyllodocida, Polychaeta, Annelida) and Evaluation of the Position of Antonbruunia. Diversity. 2024; 16(3):134. https://doi.org/10.3390/d16030134
Chicago/Turabian StyleHuč, Sonja, Avery S. Hiley, Marina F. McCowin, and Greg W. Rouse. 2024. "A Mitogenome-Based Phylogeny of Pilargidae (Phyllodocida, Polychaeta, Annelida) and Evaluation of the Position of Antonbruunia" Diversity 16, no. 3: 134. https://doi.org/10.3390/d16030134
APA StyleHuč, S., Hiley, A. S., McCowin, M. F., & Rouse, G. W. (2024). A Mitogenome-Based Phylogeny of Pilargidae (Phyllodocida, Polychaeta, Annelida) and Evaluation of the Position of Antonbruunia. Diversity, 16(3), 134. https://doi.org/10.3390/d16030134