
The Phyloperiodic Approach Removes the “Cryptic Species” and Puts forward Multilevel Organismal Diversity



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Nomenclatural Acts
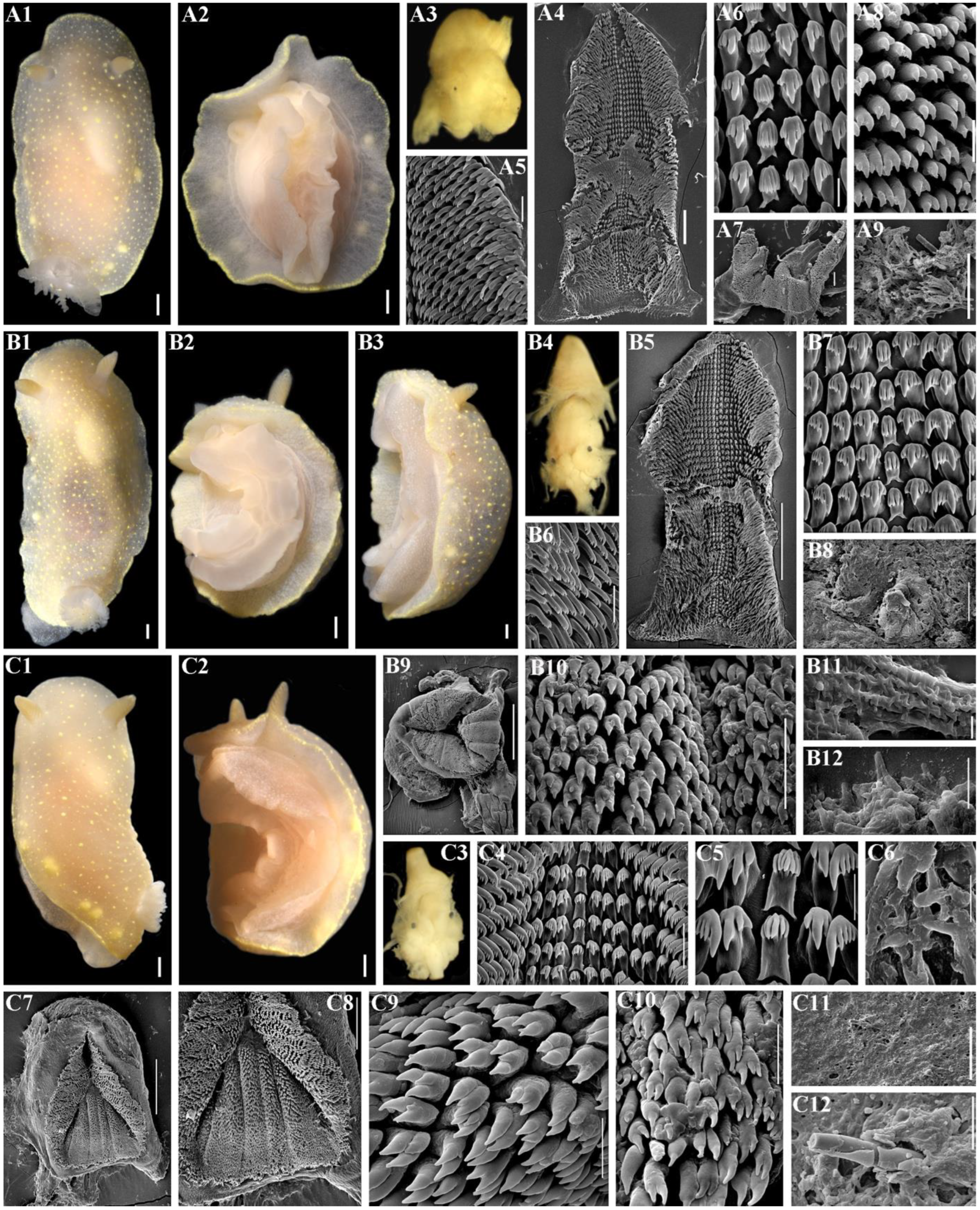
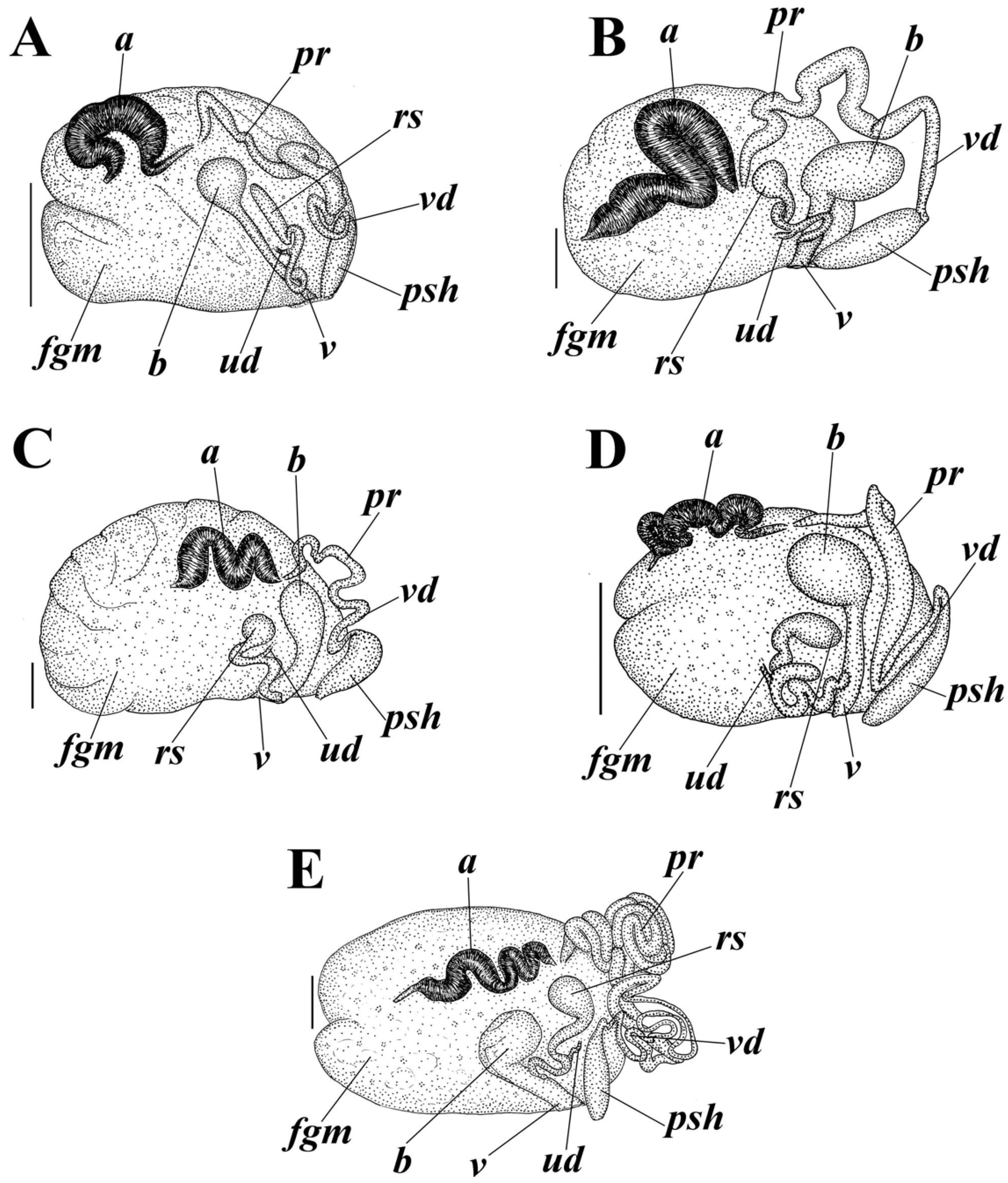
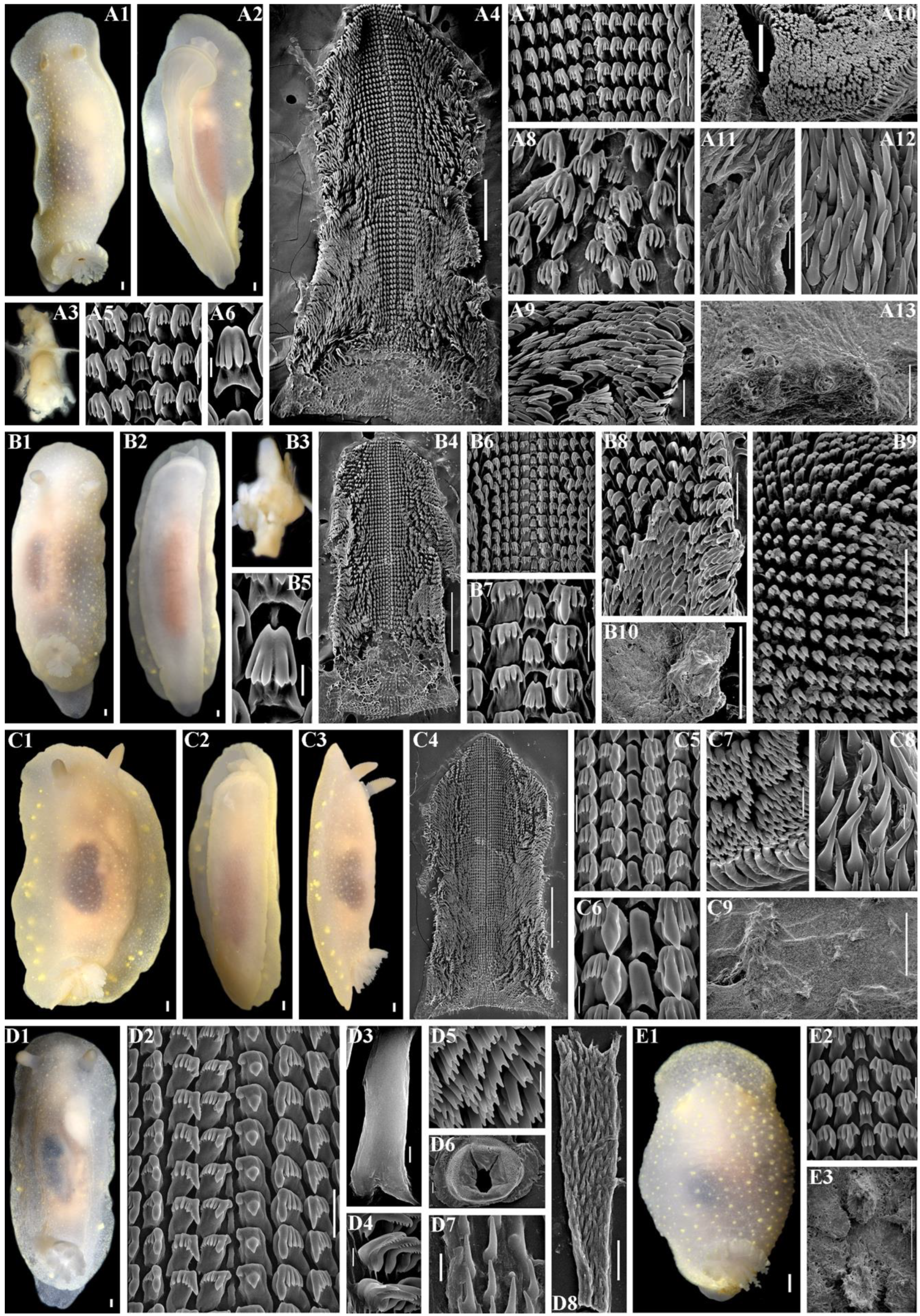
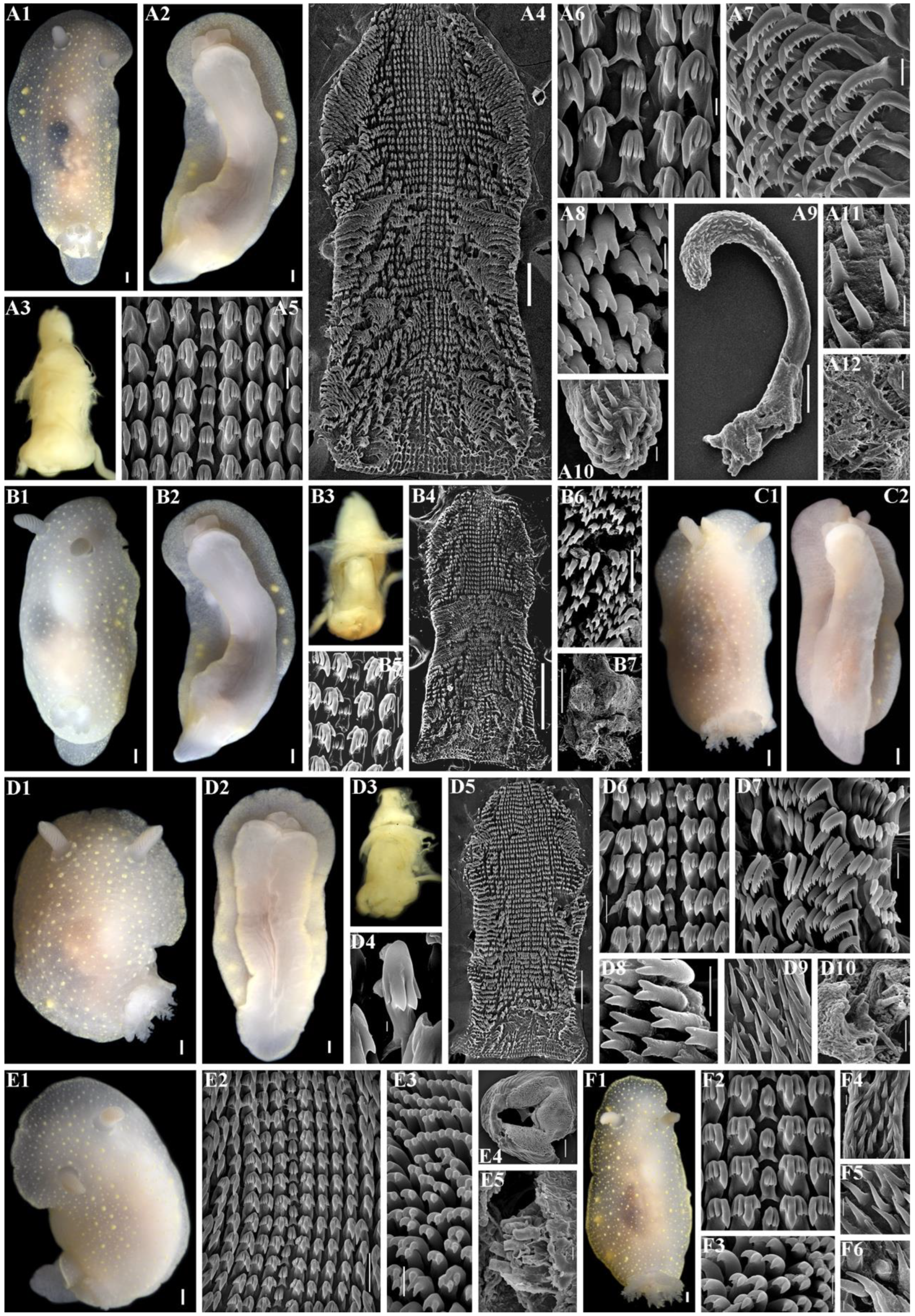
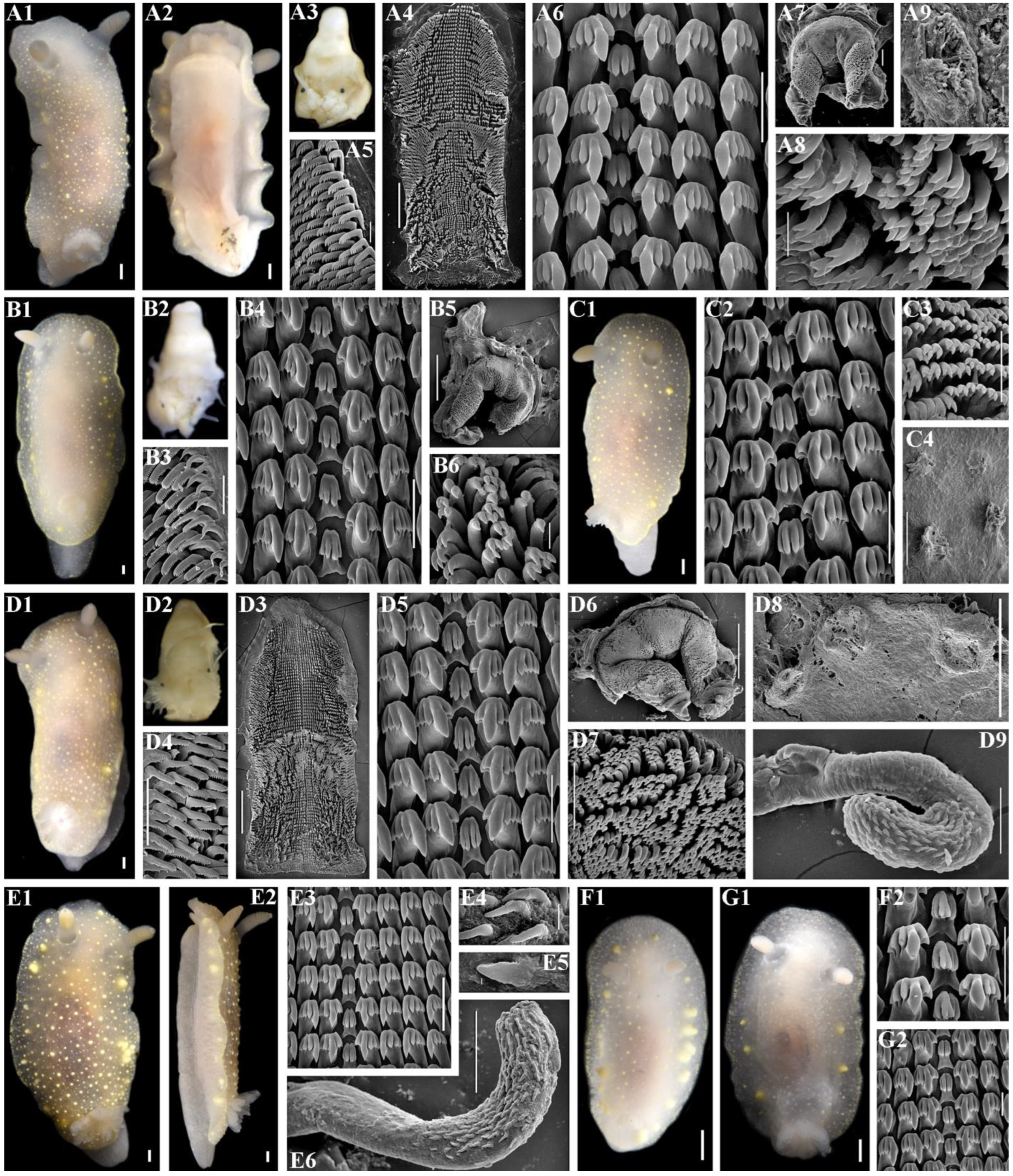
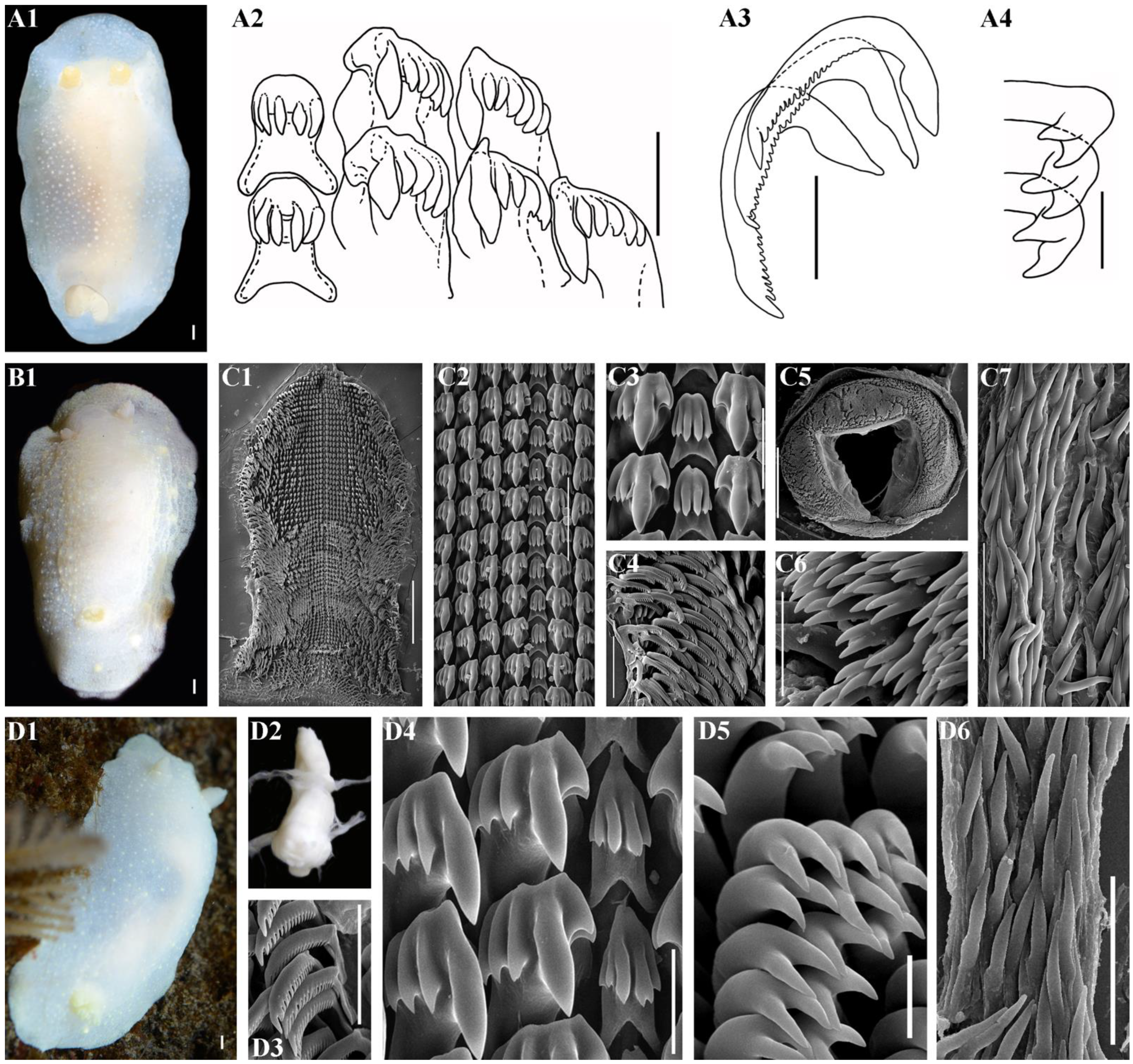
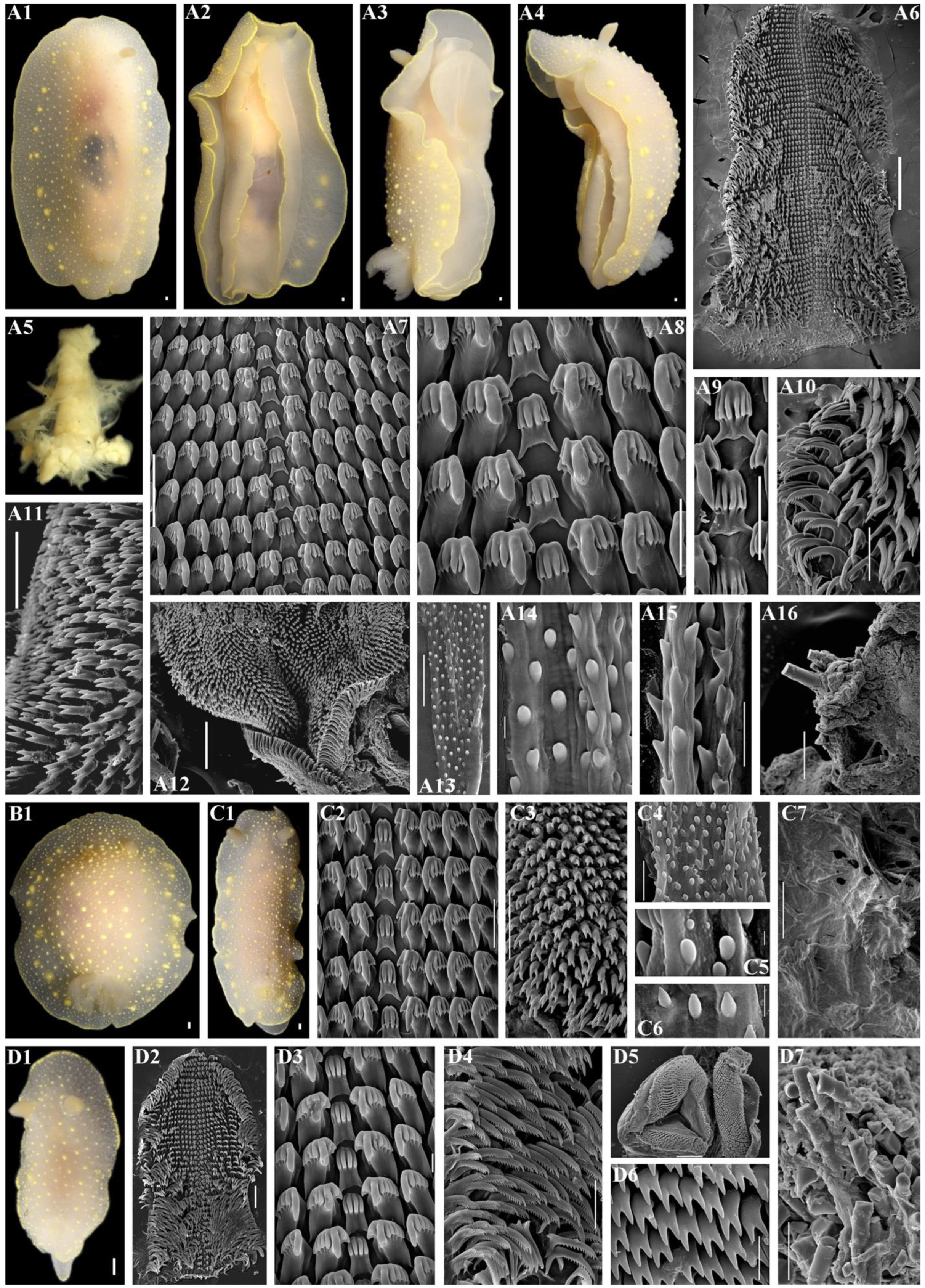
2.3. Morphological Analysis
2.4. Molecular Analysis
3. Results
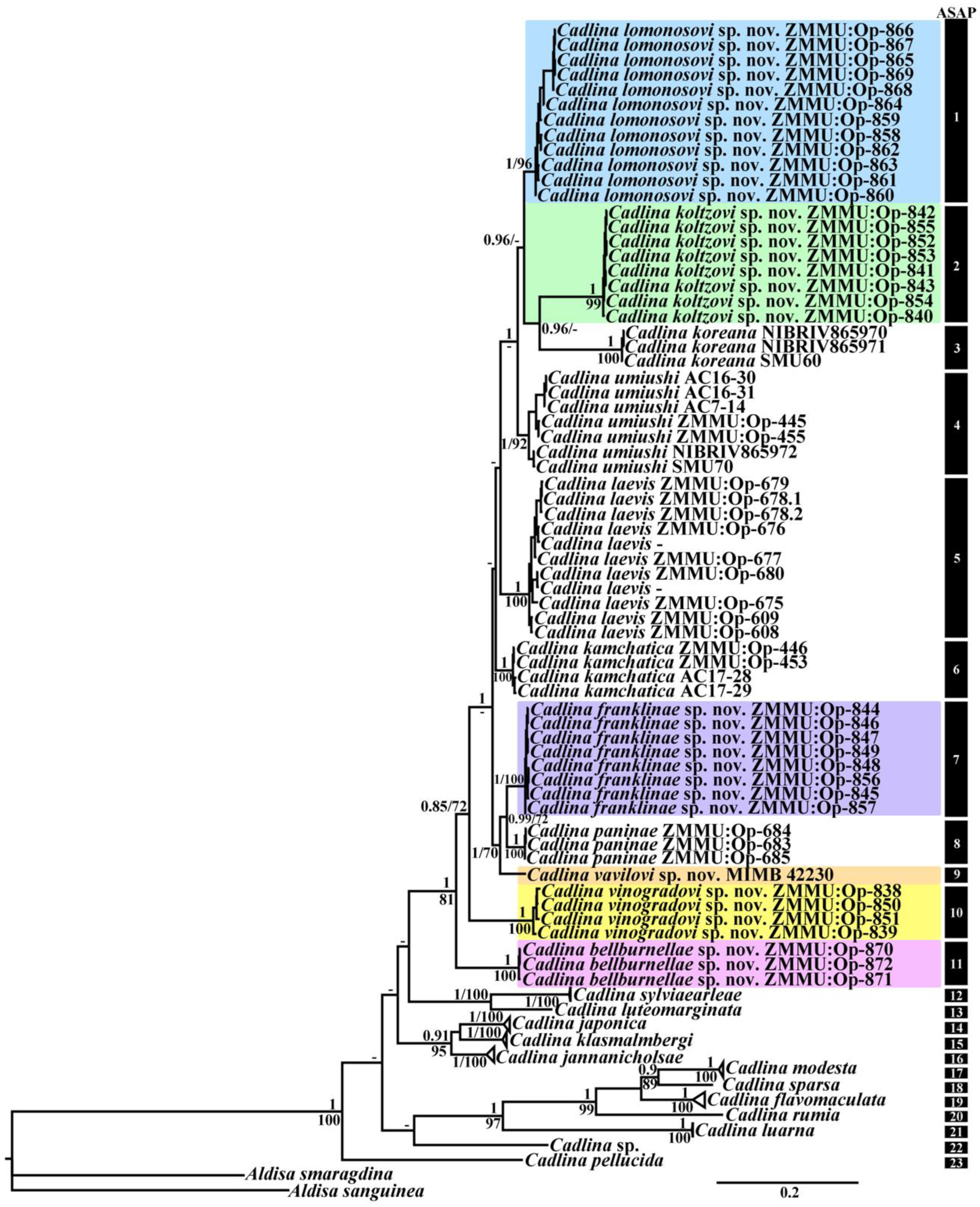
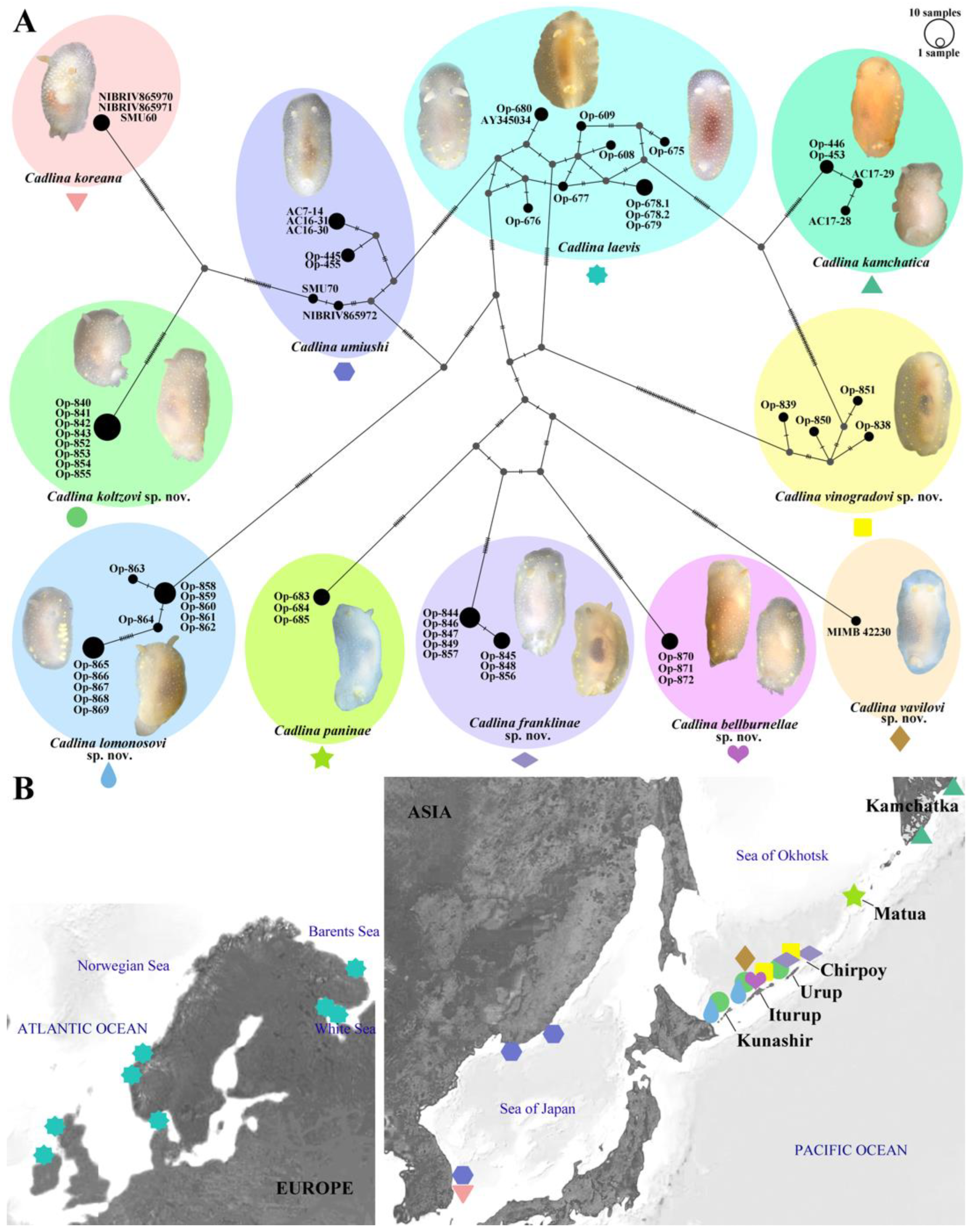
3.1. Molecular Phylogeny
3.2. Practical Guidelines Proposing How to Perform a Taxonomic Study in the Molecular Era
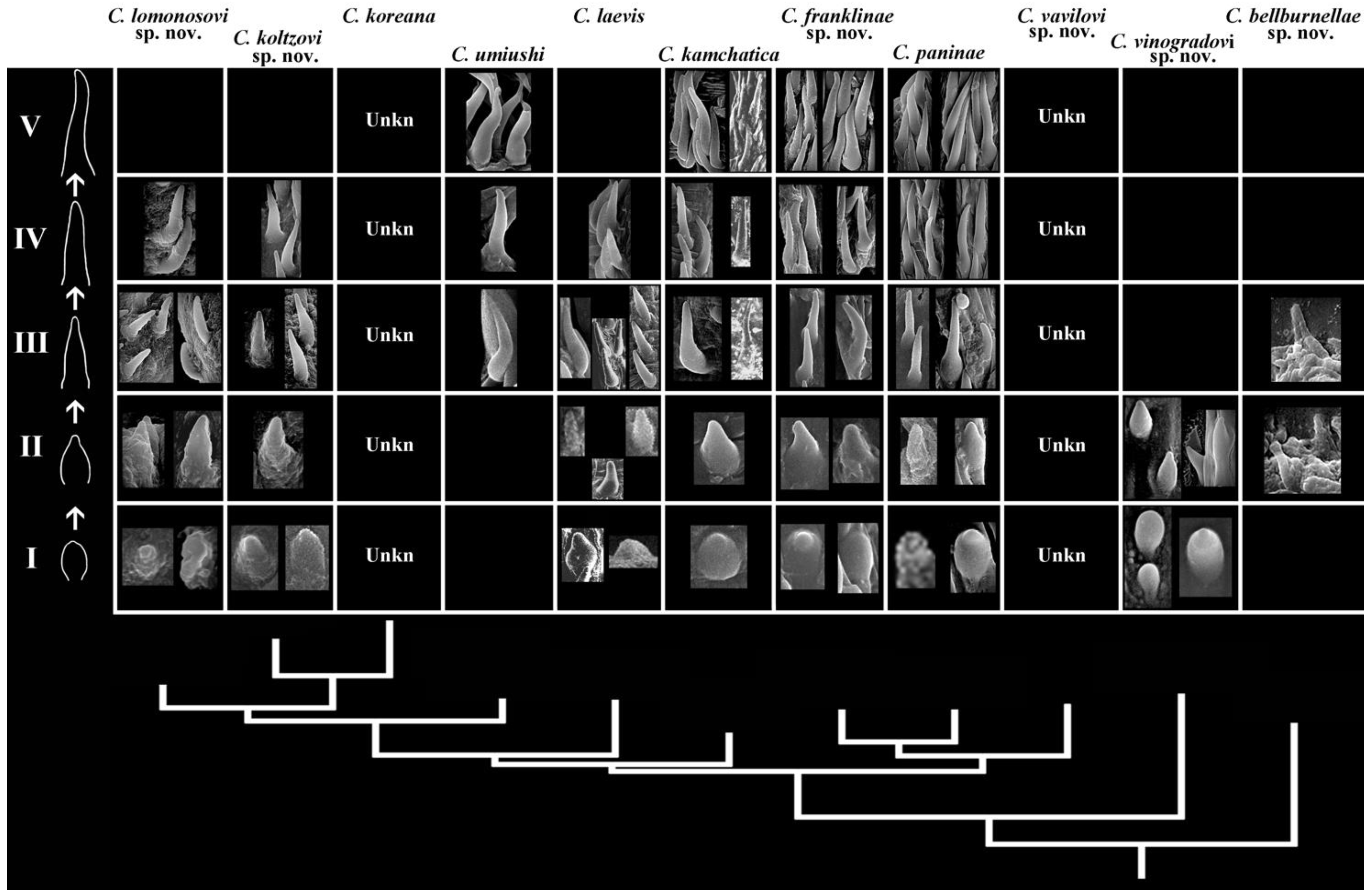
3.3. Phyloperiodic Framework for Recognition of Diagnosable Characters in Species Complexes
- I
- Body white, yellow spots absent, brownish/dark colouration absent, dispersed white spots may be present, yellow notal line absent, light yellow subepidermal glands distinguishable, and rhinophores and gills white;
- II
- Body white, yellow spots absent, brownish/dark colouration absent, dispersed white spots may be present, yellow notal line absent, yellow subepidermal glands barely distinguishable, and rhinophores and gills light yellow;
- III
- Body white, yellow spots absent to hardly distinguishable, dispersed white spots may be present, brownish/dark colouration absent, yellow notal line absent, yellow subepidermal glands distinguishable, and rhinophores and gills white to light yellow;
- IV
- Body white to creamy, yellow spots absent, dispersed white spots may be present, brownish/dark colouration absent, yellow notal line absent, yellow subepidermal glands distinguishable, and rhinophores and gills yellow to light brownish;
- V
- Body white to creamy, faint yellow spots may be present, dispersed white spots may be present, brownish/dark colouration absent, yellow notal line absent, yellow subepidermal glands barely distinguishable, and rhinophores and gills white to light yellow;
- VI
- Body white to creamy, faint yellow spots present, dispersed white spots may be present, brownish/dark colouration absent, yellow notal line partially present, dispersed, yellow subepidermal glands distinguishable, and rhinophores and gills yellow to light brownish;
- VII
- Body white to creamy, distinct yellow spots present, white spots barely distinguishable, brownish/dark colouration absent, yellow notal line present, moderately distinct, yellow subepidermal glands distinguishable, and rhinophores and gills white to light brownish;
- VIII
- Body white to creamy and light brownish, distinct yellow spots present, white spots may form distinct opaque white covering, brownish/dark colouration absent, yellow notal line present, yellow subepidermal glands distinguishable, and rhinophores and gills yellow to light brownish;
- IX
- Body white to brownish, distinct yellow spots present, white spots barely distinguishable, brownish/dark colouration completely absent, yellow notal line distinct, yellow subepidermal glands distinguishable, and rhinophores and gills white to light yellow;
- X
- Body with patchy brownish to uniform yellowish/dark colouration, white spots barely distinguishable, numerous yellow spots present, yellow notal line distinct, yellow subepidermal glands distinguishable, and rhinophores and gills yellow to brownish;
- XI
- Body with almost completely brownish/dark colouration, white and yellow spots, yellow notal line or subepidermal glands barely distinguishable, and rhinophores and gills brownish.
3.4. Systematics







4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karanovic, T.; Djurakic, M.; Eberhard, S.M. Cryptic species or inadequate taxonomy? Implementation of 2D geometric morphometrics based on integumental organs as landmarks for delimitation and description of copepod taxa. Syst. Biol. 2016, 65, 304–327. [Google Scholar] [CrossRef] [PubMed]
- Zúñiga-Reinoso, A.; Benítez, H.A. The overrated use of the morphological cryptic species concept: An example with Nyctelia darkbeetles (Coleoptera: Tenebrionidae) using geometric morphometrics. Zool. Anz. 2015, 255, 47–53. [Google Scholar] [CrossRef]
- Heethoff, M. Cryptic species—Conceptual or terminological chaos? A response to Struck et al. Trends Ecol. Evol. 2018, 33, 310. [Google Scholar] [CrossRef]
- Korshunova, T.A.; Martynov, A.V.; Bakken, T.; Picton, B.E. External diversity is restrained by internal conservatism: New nudibranch mollusc contributes to the cryptic species problem. Zool. Scr. 2017, 46, 683–692. [Google Scholar] [CrossRef]
- Korshunova, T.A.; Picton, B.; Furfaro, G.; Mariottini, P.; Pontes, M.; Prkić, J.; Fletcher, K.; Malmberg, K.; Lundin, K.; Martynov, A. Multilevel fine scale diversity challenges the ‘cryptic species’ concept. Sci. Rep. 2019, 9, 6732. [Google Scholar] [CrossRef] [PubMed]
- Horsáková, V.; Nekola, J.C.; Horsák, M. When is a “cryptic” species not a cryptic species: A consideration from the Holarctic micro-landsnail genus Euconulus (Gastropoda: Stylommatophora) deciphering “cryptic” pyramidula species. Mol. Phylogenetics Evol. 2019, 132, 307–320. [Google Scholar] [CrossRef]
- Korshunova, T.A.; Driessen, F.; Picton, B.; Martynov, A.V. The multilevel organismal diversity approach deciphers difficult to distinguish nudibranch species complex. Sci. Rep. 2021, 11, 18323. [Google Scholar] [CrossRef]
- Horsáková, V.; Líznarová, E.; Razkin, O.; Nekola, J.C.; Horsák, M. Deciphering “cryptic” nature of European rock-dwelling Pyramidula snails (Gastropoda: Stylommatophora). Contrib. Zool. 2022, 91, 233–260. [Google Scholar] [CrossRef]
- Minelli, A. Two-way exchanges between animal and plant biology, with focus on evo-devo. Front. Ecol. Evol. 2022, 10, 1057355. [Google Scholar] [CrossRef]
- Macleod, N.; Canty, R.J.; Polaszek, A. Morphology-based identification of Bemisia tabaci cryptic species puparia via embedded group-contrast convolution neural network analysis. Syst. Biol. 2022, 71, 1095–1109. [Google Scholar] [CrossRef]
- Kramina, T.E.; Samigullin, T.H.; Meschersky, I.G. Two cryptic species of Lotus (Fabaceae) from the Iberian Peninsula. Wulfenia 2020, 27, 21–45. [Google Scholar]
- Brandão-Dias, P.F.P.; Zhang, Y.M.; Pirro, S.; Vinson, C.C.; Weinersmith, K.L.; Ward, A.K.G.; Forbes, A.A.; Egan, S.P. Describing biodiversity in the genomics era: A new species of Nearctic Cynipidae gall wasp and its genome. Syst. Entomol. 2022, 47, 94–112. [Google Scholar] [CrossRef]
- Christmas, M.J.; Jones, J.C.; Olsson, A.; Wallerman, O.; Bunikis, I.; Kierczak, M.; Peona, V.; Whitley, K.M.; Larva, T.; Suh, A.; et al. Genetic barriers to historical gene flow between cryptic species of alpine bumblebees revealed by comparative population genomics. Mol. Biol. Evol. 2021, 38, 3126–3143. [Google Scholar] [CrossRef] [PubMed]
- Calahorra-Oliart, A.; Ospina-Garcés, S.M.; León-Paniagua, L. Cryptic species in Glossophaga soricina (Chiroptera: Phyllostomidae): Do morphological data support molecular evidence? J. Mammal. 2021, 102, 54–68. [Google Scholar] [CrossRef]
- Chan, K.O.; Hutter, C.R.; Wood, P.L.; Su, Y.C.; Brown, R.M. Gene flow increases phylogenetic structure and inflates cryptic species estimations: A case study on widespread Philippine puddle frogs (Occidozyga laevis). Syst. Biol. 2021, 71, 40–57. [Google Scholar] [CrossRef] [PubMed]
- Clavero-Camacho, I.; Palomares-Rius, J.E.; Cantalapiedra-Navarrete, C.; León-Ropero, G.; Martín-Barbarroja, J.; Archidona-Yuste, A.; Castillo, P. Integrative taxonomy reveals hidden cryptic diversity within pin nematodes of the genus Paratylenchus (Nematoda: Tylenchulidae). Plants 2021, 10, 1454. [Google Scholar] [CrossRef] [PubMed]
- Connolly, J.B.; Romeis, J.; Devos, Y.; Glandorf, D.C.; Turner, G.; Coulibaly, M.B. Gene drive in species complexes: Defining target organisms. Trends Biotechnol. 2022, 41, 154–164. [Google Scholar] [CrossRef]
- Costa-Araújo, R.; Silva, J.S.; Boubli, J.P.; Rossi, R.V.; Canale, G.R.; Melo, F.R.; Bertuol, F.; Silva, F.E.; Silva, D.A.; Nash, S.D.; et al. An integrative analysis uncovers a new, pseudo-cryptic species of Amazonian marmoset (Primates: Callitrichidae: Mico) from the arc of deforestation. Sci. Rep. 2021, 11, 15665. [Google Scholar] [CrossRef] [PubMed]
- Irisarri, I.; Darienko, T.; Pröschold, T.; Fürst-Jansen, J.M.R.; Jamy, M.; de Vries, J. Unexpected cryptic species among streptophyte algae most distant to land plants. Proc. R. Soc. B Biol. Sci. 2021, 288, 20212168. [Google Scholar] [CrossRef]
- Maggia, M.E.; Decaëns, T.; Lapied, E.; Dupont, L.; Roy, V.; Schimann, H.; Orivel, J.; Murienne, J.; Baraloto, C.; Cottenie, K.; et al. At each site its diversity: DNA barcoding reveals remarkable earthworm diversity in neotropical rainforests of French Guiana. Appl. Soil Ecol. 2021, 164, 103932. [Google Scholar] [CrossRef]
- Majtyka, T.; Borczyk, B.; Ogielska, M.; Stöck, M. Morphometry of two cryptic tree frog species at their hybrid zone reveals neither intermediate nor transgressive morphotypes. Ecol. Evol. 2022, 12, e8527. [Google Scholar] [CrossRef] [PubMed]
- Parsons, D.J.; Pelletier, T.A.; Wieringa, J.G.; Duckett, D.J.; Carstens, B.C. Analysis of biodiversity data suggest that mammal species are hidden in predictable places. Proc. Natl. Acad. Sci. USA 2022, 119, e2103400119. [Google Scholar] [CrossRef] [PubMed]
- Pérez, M.; Fernández-Míguez, M.; Matallanas, J.; Lloris, D.; Presa, P. Phylogenetic prospecting for cryptic species of the genus Merluccius (Actinopterygii: Merlucciidae). Sci. Rep. 2021, 11, 5929. [Google Scholar] [CrossRef] [PubMed]
- Yuzefovich, A.P.; Artyushin, I.V.; Kruskop, S.V. Not the cryptic species: Diversity of Hipposideros gentilis (Chiroptera: Hipposideridae) in Indochina. Diversity 2021, 13, 218. [Google Scholar] [CrossRef]
- Pfingstl, T.; Schäffer, S.; Bardel-Kahr, I.; Baumann, J. A closer look reveals hidden diversity in the intertidal Caribbean Fortuyniidae (Acari, Oribatida). PLoS ONE 2022, 17, e0268964. [Google Scholar] [CrossRef] [PubMed]
- Pyron, R.A.; Beamer, D.A. Nomenclatural solutions for diagnosing ‘cryptic’ species using molecular and morphological data facilitate a taxonomic revision of the Black-bellied Salamanders (Urodela, Desmognathus ‘quadramaculatus’) from the southern Appalachian Mountains. Bionomina 2022, 27, 1–43. [Google Scholar] [CrossRef]
- Semedo, T.B.F.; Da Silva, M.N.F.; Carmignotto, A.P.; Rossi, R.V. Three new species of spiny mice, genus Neacomys Thomas, 1900 (Rodentia: Cricetidae), from Brazilian Amazonia. Syst. Biodivers. 2021, 19, 1113–1134. [Google Scholar] [CrossRef]
- Yin, Y.; Yao, L.; Hu, Y.; Shao, Z.; Hong, X.; Hebert, P.D.N.; Xue, X. DNA barcoding uncovers cryptic diversity in minute herbivorous mites (Acari, Eriophyoidea). Mol. Ecol. Res. 2022, 22, 1986–1998. [Google Scholar] [CrossRef] [PubMed]
- Martynov, A.V.; Korshunova, T.A. Multilevel Organismal diversity in an Ontogenetic Framework as a Solution for the Species Concept. In Cryptic Species Morphological Stasis, Circumscription, and Hidden Diversity; Monro, A.K., Mayo, S.J., Eds.; Cambridge University Press: Cambridge, UK, 2022; pp. 78–129. [Google Scholar] [CrossRef]
- Korshunova, T.A.; Sanamyan, N.P.; Sanamyan, K.E.; Bakken, T.; Lundin, K.; Fletcher, K.; Martynov, A.V. Biodiversity hotspot in cold waters: A review of the genus Cuthonella with descriptions of seven new species (Mollusca, Nudibranchia). Contrib. Zool. 2021, 90, 216–283. [Google Scholar] [CrossRef]
- Korshunova, T.A.; Martynov, A.V. Increased information on biodiversity from the neglected part of the North Pacific contributes to the understanding of phylogeny and taxonomy of nudibranch molluscs. Can. J. Zool. 2022, 100, 436–451. [Google Scholar] [CrossRef]
- Korshunova, T.A.; Fletcher, K.; Picton, B.; Lundin, K.; Kashio, S.; Sanamyan, N.; Sanamyan, K.; Padula, V.; Schroedl, M.; Martynov, A. The Emperor Cadlina, hidden diversity and gill cavity evolution: New insights for taxonomy and phylogeny of dorid nudibranchs (Mollusca: Gastropoda). Zool. J. Linn. Soc. 2020, 189, 762–827. [Google Scholar] [CrossRef]
- Gómez Daglio, L.; Dawson, M.N. Integrative taxonomy: Ghosts of past, present and future. J. Mar. Biol. Assoc. UK 2019, 99, 1237–1246. [Google Scholar] [CrossRef]
- Korshunova, T.; Grøtan, V.V.; Johnson, K.B.; Bakken, T.; Picton, B.E.; Martynov, A. Similar ones are not related and vice versa—New Dendronotus taxa (Nudibranchia: Dendronotidae) from the North Atlantic ocean provide a platform for discussion of global marine biodiversity patterns. Diversity 2023, 15, 504. [Google Scholar] [CrossRef]
- Taylor, W.R. A ‘periodic table’ for protein structures. Nature 2002, 416, 657–660. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.J.; Hassan, M.; Jernigan, R.L.; Jia, K.; Kihara, D.; Kloczkowski, A.; Kotelnikov, S.; Kozakov, D.; Liang, J.; Liwo, A.; et al. Protein folds vs. protein folding: Differing questions, different challenges. Proc. Natl. Acad. Sci. USA 2023, 120, e2214423119. [Google Scholar] [CrossRef]
- Wang, S.S.; Ellington, A.D. Pattern generation with nucleic acid chemical reaction networks. Chem. Rev. 2019, 119, 6370–6383. [Google Scholar] [CrossRef] [PubMed]
- Xia, B.; Yanai, I. A periodic table of cell types. Development 2019, 146, dev169854. [Google Scholar] [CrossRef]
- Gante, H.F. How fish get their stripes again and again. Science 2018, 362, 396–397. [Google Scholar] [CrossRef]
- Inaba, M.; Chuong, C.M. Avian pigment pattern formation: Developmental control of macro- (across the body) and micro- (within a feather) level of pigment patterns. Front. Cell Dev. Biol. 2020, 8, 620. [Google Scholar] [CrossRef]
- Freudstein, J.V.; Broe, M.B.; Folk, R.A.; Sinn, B.T. Biodiversity and the species concept—Lineages are not enough. Syst. Biol. 2017, 66, 644–656. [Google Scholar] [CrossRef]
- Sáez, A.G.; Probert, I.; Geisen, M.; Quinn, P.; Young, J.R.; Medlin, L.K. Pseudo-cryptic speciation in coccolithophores. Proc. Natl. Acad. Sci. USA 2003, 100, 7163–7168. [Google Scholar] [CrossRef] [PubMed]
- Bickford, D.; Lohman, D.J.; Sodhi, N.S.; Ng, P.K.; Meier, R.; Winker, K.; Ingram, K.K.; Das, I. Cryptic species as a window on diversity and conservation. Trends Ecol. Evol. 2007, 22, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Fišer, C.; Robinson, C.T.; Malard, F. Cryptic species as a window into the paradigm shift of the species concept. Mol. Ecol. 2018, 27, 613–635. [Google Scholar] [CrossRef] [PubMed]
- Jörger, K.M.; Schrödl, M. How to describe a cryptic species? Practical challenges of molecular taxonomy. Front. Zool. 2013, 10, 59. [Google Scholar] [CrossRef] [PubMed]
- Cerca, J.; Meyer, C.; Purschke, G.; Struck, T.H. Delimitation of cryptic species drastically reduces the geographical ranges of marine interstitial ghost-worms (Stygocapitella; Annelida, Sedentaria). Mol. Phyl. Evol. 2020, 143, 106663. [Google Scholar] [CrossRef] [PubMed]
- Minelli, A. Taxonomy needs pluralism, but a controlled and manageable one. Megataxa 2020, 1, 9–18. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis Version 11. Mol. Biol. Evol. 2012, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML web servers. Syst. Biol. 2008, 75, 758–771. [Google Scholar] [CrossRef]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble species by automatic partitioning. Mol. Ecol. Resour. 2021, 21, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Korshunova, T.A.; Martynov, A.; Bakken, T.; Evertsen, J.; Fletcher, K.; Mudianta, I.W.; Saito, H.; Lundin, K.; Schrödl, M.; Picton, B. Polyphyly of the traditional family Flabellinidae affects a major group of Nudibranchia: Aeolidacean taxonomic reassessment with descriptions of several new families, genera, and species (Mollusca, Gastropoda). ZooKeys 2017, 717, 1–139. [Google Scholar] [CrossRef] [PubMed]
- Martynov, A.V.; Lundin, K.; Korshunova, T.A. Ontogeny, phylotypic periods, paedomorphosis, and ontogenetic systematics. Front. Ecol. Evol. 2022, 10, 806414. [Google Scholar] [CrossRef]
- Martynov, A.V.; Korshunova, T.A. A new deep-sea genus of the family Polyceridae (Nudibranchia) possesses a gill cavity, with implications for cryptobranch condition and a ‘Periodic Table’ approach to taxonomy. J. Molluscan Stud. 2015, 81, 365–379. [Google Scholar] [CrossRef]
- Korshunova, T.A.; Malmberg, K.; Prkić, J.; Petani, A.; Fletcher, K.; Lundin, K.; Martynov, A. Fine-scale species delimitation: Speciation in process and periodic patterns in nudibranch diversity. ZooKeys 2020, 917, 15–50. [Google Scholar] [CrossRef] [PubMed]
- Scerri, E. Recent attempts to change the periodic table. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2020, 378, 20190300. [Google Scholar] [CrossRef] [PubMed]
- Martynov, A.V. From ‘tree-thinking’ to ‘cycle-thinking’: Ontogenetic systematics of nudibranch molluscs. Thalassas 2011, 27, 193–224. [Google Scholar]
- Ekimova, I.; Valdés, Á.; Stanovova, M.; Mikhlina, A.; Antokhina, T.; Neretina, T.; Chichvarkhina, O.; Schepetov, D. Connected across the ocean: Taxonomy and biogeography of deep water Nudibranchia from the Northwest Pacific reveal trans Pacific links and two undescribed species. Org. Divers. Evol. 2021, 21, 753–782. [Google Scholar] [CrossRef]
- Gill, B.A.; Kondratieff, B.C.; Casner, K.L.; Encalada, A.C.; Flecker, A.S.; Gannon, D.G.; Ghalambor, C.K.; Guayasamin, J.M.; Poff, N.L.; Simmons, M.P.; et al. Cryptic species diversity reveals biogeographic support for the “mountain passes are higher in the tropics” hypothesis. Proc. R. Soc. B Biol. Sci. 2016, 283, 20160553. [Google Scholar] [CrossRef] [PubMed]
- Walters, A.D.; Cannizzaro, A.G.; Trujillo, D.A.; Berg, D.J. Addressing the Linnean shortfall in a cryptic species complex. Zool. J. Linn. Soc. 2021, 192, 277–305. [Google Scholar] [CrossRef]
- Golo, R.; Vergés, A.; Díaz-Tapia, P.; Cebrian, E. Implications of taxonomic misidentification for future invasion predictions: Evidence from one of the most harmful invasive marine algae. Mar. Pollut. Bull. 2023, 191, 114970. [Google Scholar] [CrossRef] [PubMed]
- Karanovic, T.; Bláha, M. Taming extreme morphological variability through coupling of molecular phylogeny and quantitative phenotype analysis as a new avenue for taxonomy. Sci. Rep. 2019, 9, 2429. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, C.G.; Rauch, C.; Pola, M.; Malaquias, M. Integrative taxonomy reveals a cryptic species of the nudibranch genus Polycera (Polyceridae) in European waters. J. Biol. Assoc. UK. 2020, 100, 733–752. [Google Scholar] [CrossRef]
- Korshunova, T.A.; Bakken, T.; Grøtan, V.; Johnson, K.; Lundin, K.; Martynov, A.V. A synoptic review of the family Dendronotidae (Mollusca: Nudibranchia): A multilevel organismal diversity approach. Contrib. Zool. 2020, 90, 93–153. [Google Scholar] [CrossRef]
- Maroni, P.J.; Wilson, N.G. Multiple Doris “kerguelenensis” (Nudibranchia) species span the Antarctic Polar Front. Ecol. Evol. 2022, 12, e9333. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J.; Sato, M.; Ruedy, R.; Lo, K.; Lea, D.W.; Medina-Elizade, M. Global temperature change. Proc. Natl. Acad. Sci. USA 2006, 103, 14288–14293. [Google Scholar] [CrossRef] [PubMed]
- Eyring, V.; Cox, P.M.; Flato, G.M.; Gleckler, P.J.; Abramowitz, G.; Caldwell, P.; Collins, W.D.; Gier, B.K.; Hall, A.D.; Hoffman, F.M.; et al. Taking climate model evaluation to the next level. Nat. Clim. Change 2019, 9, 102–110. [Google Scholar] [CrossRef]
- Supran, G.; Rahmstorf, S.; Oreskes, N. Assessing ExxonMobil’s global warming projections. Science 2023, 379, eabk0063. [Google Scholar] [CrossRef] [PubMed]
- Japan Oceanographic Data Center Portal. 2023. Available online: https://jdoss1.jodc.go.jp/vpage/bts.html (accessed on 9 February 2023).
- Harada, N.; Katsuki, K.; Nakagawa, M.; Matsumoto, A.; Seki, O.; Addison, J.A.; Finney, B.P.; Sato, M. Holocene sea surface temperature and sea ice extent in the Okhotsk and Bering Seas. Prog. Oceanogr. 2014, 126, 242–253. [Google Scholar] [CrossRef]
- Do, T.D.; Jung, D.-W.; Kil, H.-J.; Kim, C.-B. A report of a new species and new record of Cadlina (Nudibranchia, Cadlinidae) from South Korea. ZooKeys 2020, 996, 1–18. [Google Scholar] [CrossRef]
- Fraga, M.F.; Ballestar, E.; Paz, M.F.; Ropero, S.; Setien, F.; Ballestar, M.L.; Heine-Sucer, D.; Cigudosa, J.C.; Urioste, M.; Benitez, J.; et al. Epigenetic differences arise during the lifetime of monozygotic twins. Proc. Natl. Acad. Sci. USA 2005, 26, 10604–10609. [Google Scholar] [CrossRef] [PubMed]
- Gibert, A.; Louty, F.; Buscail, R.; Baguette, M.; Schatz, B.; Bertrand, J.A.M. Extracting quantitative information from images taken in the wild: A case study of two vicariants of the Ophrys aveyronensis species complex. Diversity 2022, 14, 400. [Google Scholar] [CrossRef]
- Silva, T.S.R.; Hamer, M.T.; Guénard, B. A checklist of Nylanderia (Hymenoptera: Formicidae: Formicinae) from Hong Kong and Macao SARs, with an illustrated identification key for species in Southeast China and Taiwan. Zootaxa 2023, 5301, 501–539. [Google Scholar] [CrossRef] [PubMed]
- González-Wevar, C.A.; Segovia, N.I.; Rosenfeld, S.; Maturana, C.S.; Jeldres, V.; Pinochet, R.; Saucède, T.; Morley, S.A.; Brickle, P.; Wilson, N.G.; et al. Seven snail species hidden in one: Biogeographic diversity in an apparently widespread periwinkle in the Southern Ocean. J. Biogeogr. 2022, 49, 1521–1534. [Google Scholar] [CrossRef]
- Lindeque, P.K.; Parry, H.E.; Harmer, R.A.; Somerfield, P.J.; Atkinson, A. Next Generation sequencing reveals the hidden diversity of zooplankton assemblages. PLoS ONE 2013, 8, e81327. [Google Scholar] [CrossRef] [PubMed]
- Korshunova, T.A.; Martynov, A.V. Consolidated data on the phylogeny and evolution of the family Tritoniidae (Gastropoda: Nudibranchia) contribute to genera reassessment and clarify the taxonomic status of the neuroscience models Tritonia and Tochuina. PLoS ONE 2020, 15, e0242103. [Google Scholar] [CrossRef]
- Edwards, D.; Morris, J.L.; Richardson, J.B.; Kenrick, P. Cryptospores and cryptophytes reveal hidden diversity in early land floras. New Phytol. 2014, 202, 50–78. [Google Scholar] [CrossRef] [PubMed]
- Engel, P.; Stepanauskas, R.; Moran, N.A. Hidden diversity in honey bee gut symbionts detected by single-cell genomics. PLoS Genet. 2014, 10, e1004596. [Google Scholar] [CrossRef]
- Muggia, L.; Leavitt, S.; Barreno, E. The hidden diversity of lichenised Trebouxiophyceae (Chlorophyta). Phycologia 2018, 57, 503–524. [Google Scholar] [CrossRef]
- Schulz, F.; Alteio, L.; Goudeau, D.; Ryan, E.M.; Yu, F.B.; Malmstrom, R.R.; Blanchard, J.; Woyke, T. Hidden diversity of soil giant viruses. Nature Comm. 2018, 9, 4881. [Google Scholar] [CrossRef]
- Santamaría, R.I.; Bustos, P.; Van Cauwenberghe, J.; González, V. Hidden diversity of double-stranded DNA phages in symbiotic Rhizobium species. Philos. Trans. R. Soc. B Biol. Sci. 2021, 377, 20200468. [Google Scholar] [CrossRef] [PubMed]
- Ampai, N.; Rujirawan, A.; Yodthong, S.; Termprayoon, K.; Stuart, B.L.; Wood, P.L., Jr.; Aowphol, A. Hidden diversity of rock geckos within the Cnemaspis siamensis species group (Gekkonidae, Squamata): Genetic and morphological data from southern Thailand reveal two new insular species and verify the phylogenetic affinities of C. chanardi and C. kamolnorranathi. ZooKeys 2022, 1125, 115–158. [Google Scholar] [CrossRef] [PubMed]
- Claridge, A.W.; Mifsud, G.; Dawson, J.; Saxon, M.J. Use of infrared digital cameras to investigate the behaviour of cryptic species. Wildl. Res. 2004, 31, 645–650. [Google Scholar] [CrossRef]
- Willan, R.C. The taxonomy of two host-specific, cryptic dendronotoid nudibranch species (Mollusca: Gastropoda) from Australia including a new species description. Zool. J. Linn. Soc. 1988, 94, 39–63. [Google Scholar] [CrossRef]
- Karp, D. Detecting small and cryptic animals by combining thermography and a wildlife detection dog. Sci. Rep. 2020, 10, 5220. [Google Scholar] [CrossRef] [PubMed]
- Jaklitsch, W.M.; Voglmayr, H. Hidden diversity in Thyridaria and a new circumscription of the Thyridariaceae. Stud. Mycol. 2016, 85, 35–64. [Google Scholar] [CrossRef] [PubMed]
- Padró, J.; Esteven, A.S.; Soto, I.M. DNA barcodes reveal the hidden arthropod diversity in a threatened cactus forest of the central Andes. Biodivers. Conserv. 2022, 32, 567–587. [Google Scholar] [CrossRef]
- Villalobos-Barrantes, H.M.; Meriño, B.M.; Walter, H.E.; Guerrero, P.C. Independent evolutionary lineages in a globular cactus species complex reveals hidden diversity in a central Chile biodiversity hotspot. Genes 2022, 13, 240. [Google Scholar] [CrossRef]
- Ferreira, V.S.; Ivie, M.A. Lessons from a museum’s cabinet: DNA barcoding and collections-based life stage associations reveals a hidden diversity in the Puerto Rican Bank paedomorphic Lycidae (Coleoptera: Elateroidea: Leptolycini). Insect Syst. Divers. 2022, 6, 1–36. [Google Scholar] [CrossRef]
- Jiang, L.; Zhou, M.; Sarker, K.K.; Huang, J.; Chen, W.; Li, C. Mitochondrial genome uncovered hidden genetic diversity in Microdous chalmersi (Teleostei: Odontobutidae). Fishes 2023, 8, 228. [Google Scholar] [CrossRef]
- Winemiller, K.O.; Fitzgerald, D.B.; Bower, L.M.; Pianka, E.R. Functional traits, convergent evolution, and periodic tables of niches. Ecol. Lett. 2015, 18, 737–751. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves-Souza, T.; Chaves, L.S.; Boldorini, G.X.; Reginaldo, N.F.; Gusmão, A.F.; Perônico, P.B.; Sanders, N.J.; Teresa, F.B. Bringing light onto the Raunkiæran shortfall: A comprehensive review of traits used in functional animal ecology. Ecol. Evol. 2023, 13, e10016. [Google Scholar] [CrossRef] [PubMed]
- Cummings, R.D. A periodic table of monosaccharides. Glycobiology 2024, 34, cwad088. [Google Scholar] [CrossRef]
- Poveda-Cuellar, J.L.; Conde-Saldaña, C.C.; Villa-Navarro, F.A.; Lujan, N.K.; Dergam dos Santos, J.A. Phylogenetic revision of whisker-cheeked suckermouth catfishes (Loricariidae: Lasiancistrus) from east of the Andes: Five species where once there were two. Zool. J. Linn. Soc. 2023, 199, 1–25. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Distances within Species | C. bellburnellae sp. nov. | C. franklinae sp. nov. | C. koltzovi sp. nov. | C. lomonosovi sp. nov. | C. vavilovi sp. nov. | C. vinogradovi sp. nov. | |
|---|---|---|---|---|---|---|---|
| C. bellburnellae sp. nov. | 0 | - | 7.90–8.05 | 8.05–8.21 | 7.45–7.75 | 8.99 | 8.81–9.57 |
| C. franklinae sp. nov. | 0–0.30 | 7.90–8.05 | - | 7.14–7.45 | 5.93–6.23 | 4.82–4.98 | 8.66–9.57 |
| C. koltzovi sp. nov. | 0–0.15 | 8.05–8.21 | 7.14–7.45 | - | 6.38–7.14 | 8.83 | 10.18–11.09 |
| C. lomonosovi sp. nov. | 0–1.52 | 7.45–7.75 | 5.93–6.23 | 6.38–7.14 | - | 6.10–6.42 | 8.81–9.73 |
| C. vavilovi sp. nov. | - | 8.99 | 4.82–4.98 | 8.83 | 6.10–6.42 | - | 8.83–9.31 |
| C. vinogradovi sp. nov. | 0.61–1.06 | 8.81–9.57 | 8.66–9.57 | 10.18–11.09 | 8.81–9.73 | 8.83–9.31 | - |
| C. flavomaculata | 0.85 | 13.61–13.98 | 16.33–16.72 | 13.95–14.13 | 15.48–16.41 | 17.34–17.52 | 15.65–16.57 |
| C. jannanicholsae | 0.17–0.91 | 10.49–10.83 | 11.40–12.18 | 10.64–11.25 | 10.33–11.40 | 12.52–12.86 | 11.85–12.61 |
| C. japonica | 0–0.78 | 11.41–12.01 | 11.41–12.46 | 11.88–12.77 | 11.09–12.61 | 13.32–13.48 | 11.88–12.77 |
| C. kamchatica | 0–0.30 | 7.90–8.21 | 4.56–5.02 | 7.45–7.90 | 5.62–6.08 | 5.30–5.62 | 8.05–8.97 |
| C. klasmalmbergi | 0–0.30 | 11.85–12.19 | 12.46–12.96 | 12.31–12.77 | 11.85–12.61 | 14.45–14.61 | 12.77–13.73 |
| C. koreana | 0 | 8.13 | 8.28–8.44 | 6.72–6.88 | 6.41–7.03 | 9.47 | 10.00–10.63 |
| C. laevis | 0–1.37 | 7.14–7.90 | 4.41–5.02 | 6.69–7.45 | 4.56–5.32 | 5.14–5.78 | 7.75–8.68 |
| C. luarna | 0 | 13.68 | 16.11–16.26 | 13.68–13.83 | 15.81–16.41 | 17.34 | 15.35–16.26 |
| C. luteomarginata | - | 11.57 | 13.24–13.39 | 11.26–11.42 | 10.50–11.57 | 13.32 | 12.48–13.24 |
| C. modesta | 0–0.61 | 14.29–14.89 | 15.96–16.57 | 14.59–15.35 | 15.35–16.72 | 17.34–17.98 | 16.57–17.48 |
| C. paninae | 0 | 6.53 | 2.74–2.89 | 7.14–7.29 | 5.02–5.17 | 4.82 | 8.36–9.12 |
| C. pellucida | - | 11.51 | 1.83–11.99 | 11.04–11.20 | 10.73–11.51 | 13.68 | 11.83–12.78 |
| C. rumia | - | 13.83 | 15.35–15.50 | 13.83–13.98 | 14.13–14.44 | 16.05 | 15.05–15.65 |
| C. sp. (Africa) | - | 12.94 | 14.92–14.76 | 14.46–14.61 | 13.70–14.46 | 17.01 | 12.94–13.24 |
| C. sparsa | - | 14.74 | 17.63–17.78 | 15.05–15.20 | 15.96–16.72 | 17.98 | 14.35–15.81 |
| C. sylviaearleae | 0 | 14.29 | 14.29–14.44 | 13.37–13.53 | 12.46–12.77 | 16.05 | 14.74–15.35 |
| C. umiushi | 0–1.56 | 6.88–7.60 | 5.47–6.38 | 5.63–6.99 | 3.28–4.10 | 5.46–6.58 | 7.97–9.42 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Korshunova, T.; Martynov, A. The Phyloperiodic Approach Removes the “Cryptic Species” and Puts forward Multilevel Organismal Diversity. Diversity 2024, 16, 220. https://doi.org/10.3390/d16040220
Korshunova T, Martynov A. The Phyloperiodic Approach Removes the “Cryptic Species” and Puts forward Multilevel Organismal Diversity. Diversity. 2024; 16(4):220. https://doi.org/10.3390/d16040220
Chicago/Turabian StyleKorshunova, Tatiana, and Alexander Martynov. 2024. "The Phyloperiodic Approach Removes the “Cryptic Species” and Puts forward Multilevel Organismal Diversity" Diversity 16, no. 4: 220. https://doi.org/10.3390/d16040220
APA StyleKorshunova, T., & Martynov, A. (2024). The Phyloperiodic Approach Removes the “Cryptic Species” and Puts forward Multilevel Organismal Diversity. Diversity, 16(4), 220. https://doi.org/10.3390/d16040220