
Understanding the Distribution, Behavioural Ecology, and Conservation Status of Asian Pelophylax



Abstract
:1. Introduction
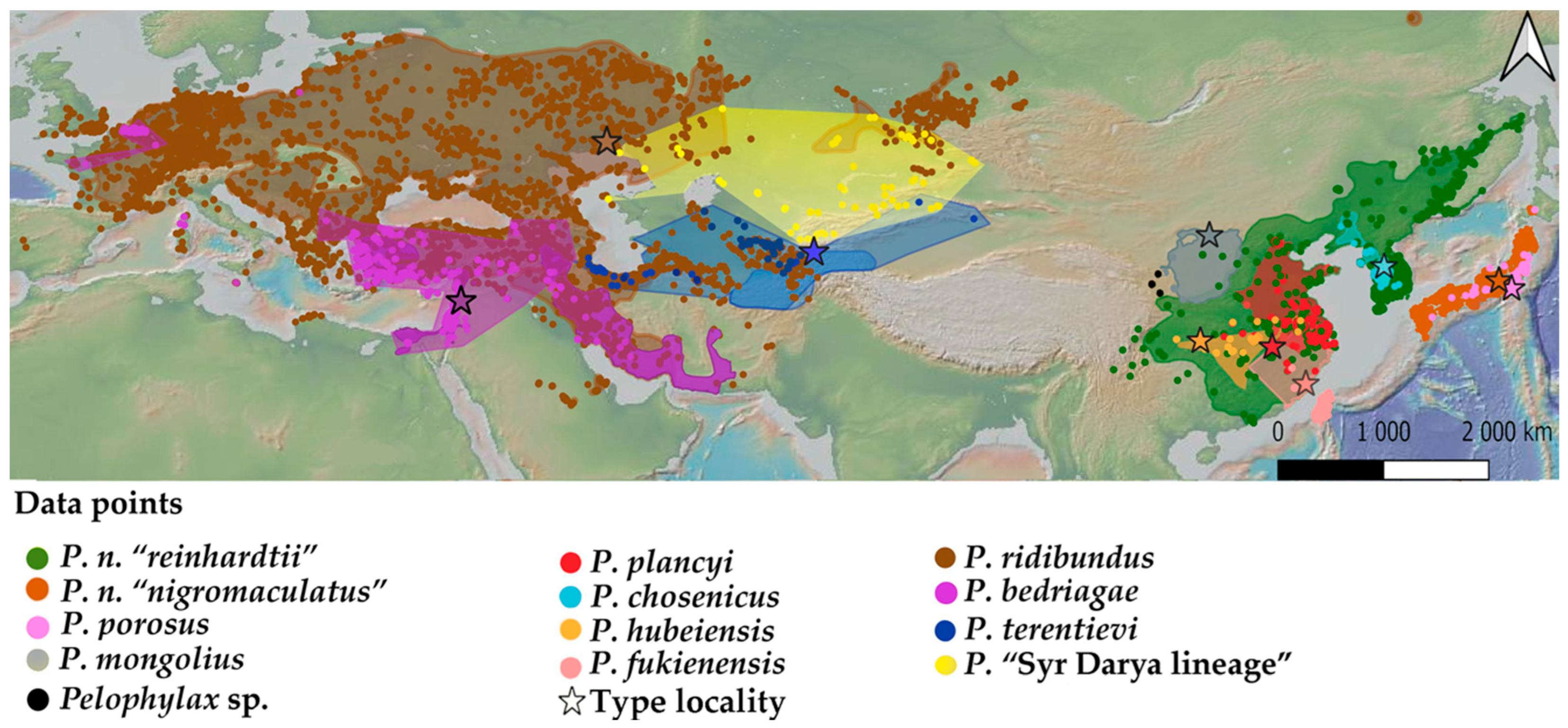
2. Materials and Methods
3. Results
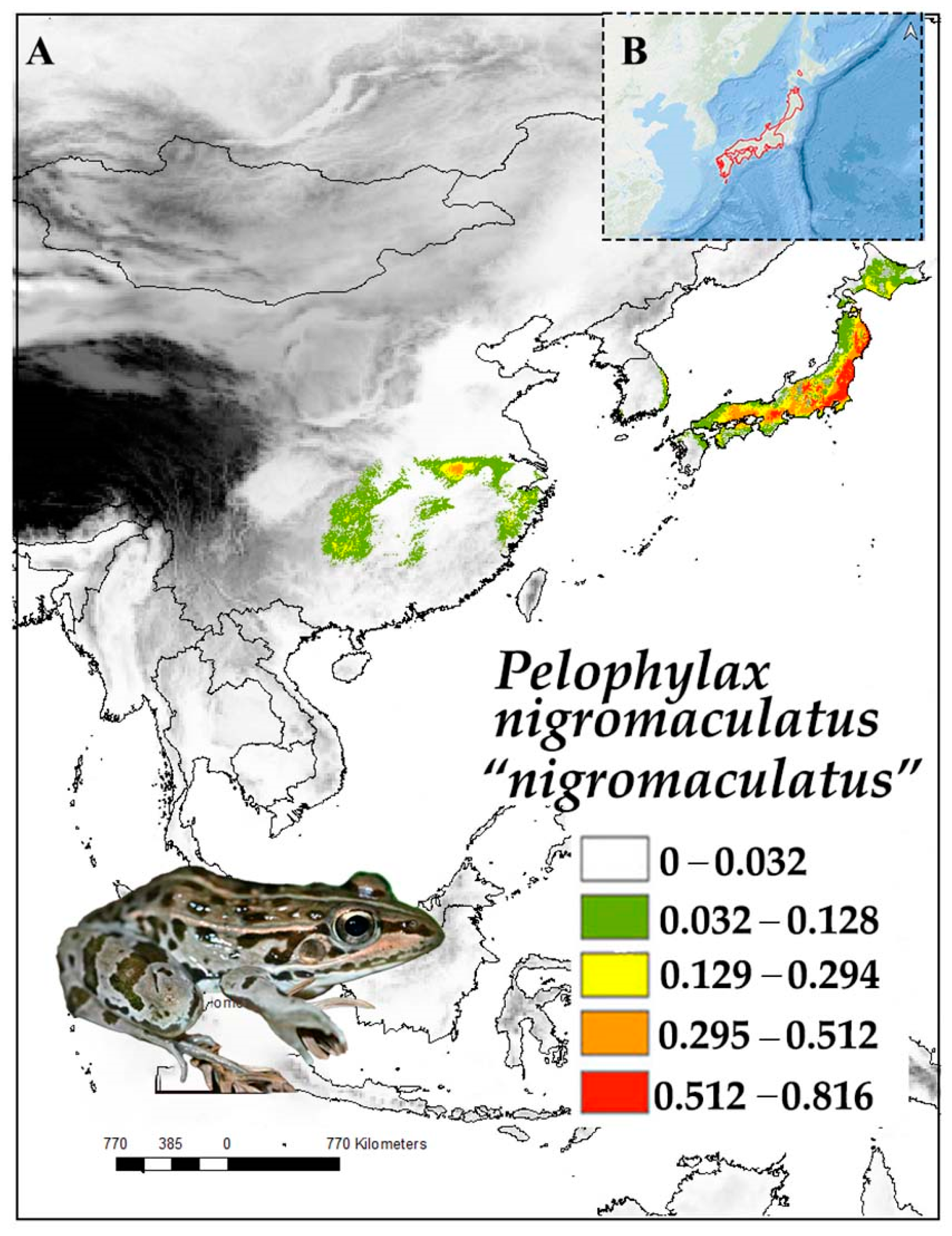
3.1. Pelophylax nigromaculatus “Nigromaculatus”
3.1.1. Distribution
3.1.2. Ecology
3.1.3. Threats and Conservation
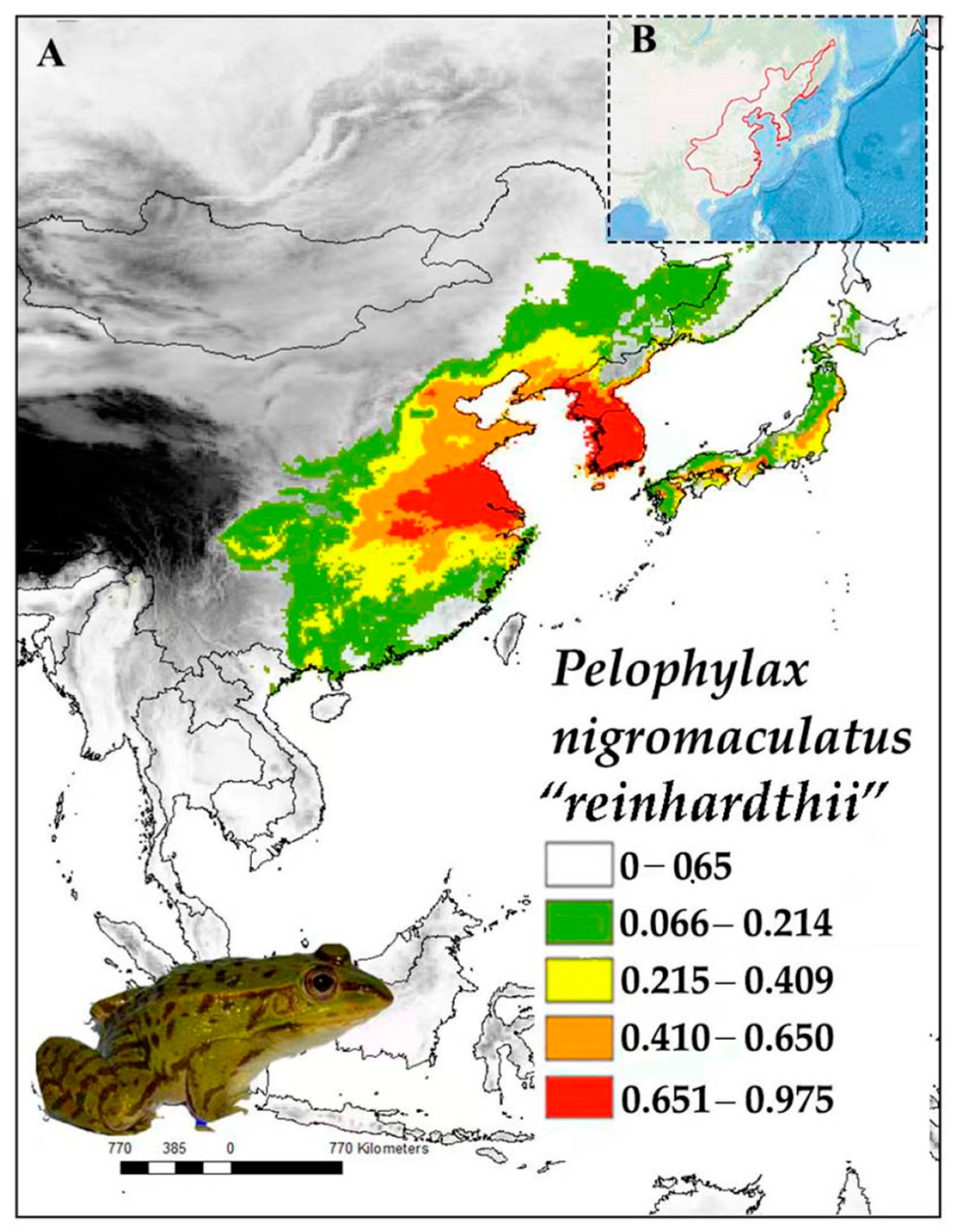
3.2. Pelophylax nigromaculatus “Reinhardtii”
3.2.1. Distribution
3.2.2. Ecology
3.2.3. Threats and Conservation
3.3. Pelophylax porosus
3.3.1. Distribution
3.3.2. Ecology
3.3.3. Threats and Conservation
3.4. Pelophylax mongolius
3.4.1. Distribution
3.4.2. Ecology
3.4.3. Threats and Conservation
3.5. Pelophylax plancyi
3.5.1. Distribution
3.5.2. Ecology
3.5.3. Threats and Conservation
3.6. Pelophylax chosenicus
3.6.1. Distribution
3.6.2. Ecology
3.6.3. Threats and Conservation
3.7. Pelophylax hubeiensis
3.7.1. Distribution
3.7.2. Ecology
3.7.3. Threats and Conservation
3.8. Pelophylax fukienensis
3.8.1. Distribution
3.8.2. Ecology
3.8.3. Threats and Conservation
3.9. Pelophylax ridibundus
3.9.1. Distribution
3.9.2. Ecology
3.9.3. Threats and Conservation
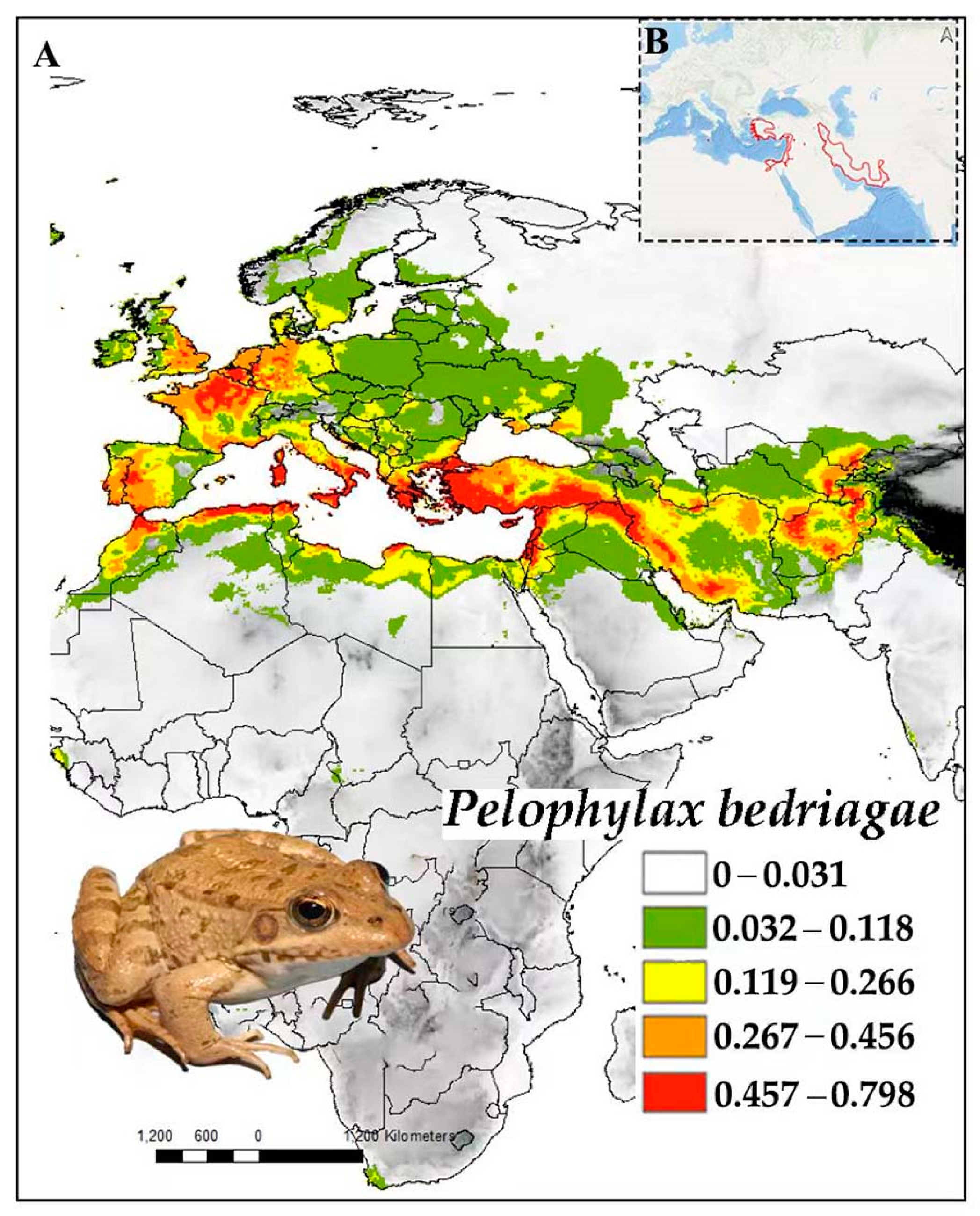
3.10. Pelophylax bedriagae
3.10.1. Distribution
3.10.2. Ecology
3.10.3. Threats and Conservation
3.11. Pelophylax terentievi
3.11.1. Distribution
3.11.2. Ecology
3.11.3. Threats and Conservation
3.12. Pelophylax “Syr Darya” Lineage
3.12.1. Distribution
3.12.2. Ecology
3.12.3. Threats and Conservation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Manne, L.; Pimm, S. Beyond eight forms of rarity: Which species are threatened and which will be next? Anim. Conserv. 2001, 4, 221–229. [Google Scholar] [CrossRef]
- Richardson, D.; Whittaker, R. Conservation biogeography—Foundations, concepts and challenges. Divers. Distrib. 2010, 16, 313–320. [Google Scholar] [CrossRef]
- Guisan, A.; Zimmermann, N.; Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Wiens, J. Speciation and ecology revisited: Phylogenetic niche conservatism and the origin of species. Evolution 2004, 58, 193–197. [Google Scholar]
- Alvarado-Serrano, D.; Knowles, L. Ecological niche models in phylogeographic studies: Applications, advances and precautions. Mol. Ecol. Resour. 2013, 14, 233–248. [Google Scholar] [CrossRef]
- Duellman, W.E. Patterns of Distribution of Amphibians: A Global Perspective; Johns Hopkins University Press: Baltimore, MD, USA, 1999; pp. 1–30. [Google Scholar]
- Fletcher, R.; Hefley, T.; Robertson, E.; Zuckerberg, B.; Mccleery, R.; Dorazio, R. A practical guide for combining data to model species distributions. Ecology 2019, 100, e02710. [Google Scholar] [CrossRef]
- Araújo, M.; Anderson, R.; Barbosa, A.M.; Beale, C.; Dormann, C.; Early, R.; Garcia, R.; Guisan, A.; Maiorano, L.; Naimi, B.; et al. Standards for distribution models in biodiversity assessments. Sci. Adv. 2019, 5, eaat4858. [Google Scholar] [CrossRef]
- Frost, D.; Grant, T.; Faivovich, J.; Bain, R.H.; Haas, A.; Haddad, C.F.B.; Sá, R.; Channing, A.; Wilkinson, M.; Donnellan, S.; et al. The amphibian tree of life. Bull. Am. Mus. Nat. Hist. 2006, 297, 1–291. [Google Scholar] [CrossRef]
- Fu, J.; Wen, L. Impacts of quaternary glaciation, geological history and geography on animal species history in continental East Asia: A phylogeographic review. Mol. Ecol. 2023, 32, 4497–4514. [Google Scholar] [CrossRef]
- Dufresnes, C.; Litvinchuk, S.N. Diversity, distribution and molecular species delimitation in frogs and toads from the Eastern Palaearctic. Zool. J. Linn. Soc. 2022, 195, 695–760. [Google Scholar] [CrossRef]
- Borzée, A. Continental Northeast Asian Amphibians: Origins, Behavioural Ecology, and Conservation; Academic Press: Amsterdam, The Netherlands, 2024. [Google Scholar]
- Fei, L.; Ye, C.; Jiang, J. Colored Atlas of Chinese Amphibians and Their Distributions; Sichuan Science and Technology Press: Chengdu, China, 2012. [Google Scholar]
- Plötner, J.; Baier, F.; Akin, C.; Mazepa, G.; Schreiber, R.; Beerli, P.; Litvinchuk, S.N.; Bilgin, C.C.; Borkin, L.; Uzzell, T. Genetic data reveal that water frogs of Cyprus (Genus Pelophylax) are an endemic species of Messinian origin. Zoosyst. Evol. 2012, 88, 261–283. [Google Scholar] [CrossRef]
- Garcia, V.; Ivy, C.; Fu, J. Syntopic Frogs reveal different patterns of interaction with the landscape: A comparative landscape genetic study of Pelophylax nigromaculatus and Fejervarya limnocharis from central China. Ecol. Evol. 2017, 7, 9294–9306. [Google Scholar] [CrossRef] [PubMed]
- Borzée, A.; Andersen, D.; Jang, Y. Population trend inferred from aural surveys for calling anurans in Korea. PeerJ 2018, 2018, e5568. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.-Y.; Choi, S.; Kim, H.; Suh, J.-H.; Do, M.S. Ecological niche and interspecific competition of two frog species (Pelophylax nigromaculatus and P. chosenicus) in South Korea using the geographic information system. Korean J. Ecol. Environ. 2021, 54, 363–373. [Google Scholar] [CrossRef]
- Nakanishi, K.; Honma, A.; Furukawa, M.; Takakura, K.-I.; Fujii, N.; Morii, K.; Terasawa, Y.; Nishida, T. Habitat partitioning of two closely related pond frogs, Pelophylax nigromaculatus and Pelophylax porosus brevipodus, during their breeding season. Evol. Ecol. 2020, 34, 855–866. [Google Scholar] [CrossRef]
- Yang, S.; Kim, J.; Min, M.; Suh, J. Genetic diversity and population structure of two Korean pond frog species, Rana nigromaculata and R. plancyi (Anura, Ranidae), with a survey of temporal genetic variation in R. nigromaculata. Korean J. Biol. Sci. 1999, 3, 275–283. [Google Scholar] [CrossRef]
- Wang, S.; Liu, C.; Zhu, W.; Gao, X.; Yiming, L. Tracing the origin of the Black-Spotted Frog, Pelophylax nigromaculatus, in the Xinjiang uyghur autonomous region. Asian Herpetol. Res. 2016, 7, 69–74. [Google Scholar]
- Bae, Y.H.; Park, J.; Othman, S.N.; Yikweon, J.; Borzée, A. Record of invasive Rana huanrenensis Fei, Ye, and Huang, 1990 and Pelophylax nigromaculatus (Hallowell, 1861) on Ulleung Island, Republic of Korea. BioInvasions Rec. 2022, 11, 278–286. [Google Scholar] [CrossRef]
- Lyapkov, S.M.; Ermakov, O.A.; Titov, S.V. Distribution and origin of two forms of the Marsh Frog Pelophylax ridibundus complex (Anura, Ranidae) from Kamchatka based on mitochondrial and nuclear DNA data. Biol. Bull. 2018, 45, 699–705. [Google Scholar] [CrossRef]
- Dufresnes, C.; Mazepa, G. Hybridogenesis in Water Frogs. eLS 2020, 1, 718–726. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Kramer-Schadt, S.; Niedballa, J.; Pilgrim, J.; Schröder, B.; Lindenborn, J.; Reinfelder, V.; Stillfried, M.; Heckmann, I.; Scharf, A.; Augeri, D.; et al. The importance of correcting for sampling bias in MaxEnt species distribution models. Divers. Distrib. 2013, 19, 1366–1379. [Google Scholar] [CrossRef]
- Zhu, G.; Qiao, H. Effect of the maxent model’s complexity on the prediction of species potential distributions. Biodivers. Sci. 2016, 24, 1189–1196. [Google Scholar] [CrossRef]
- Kass, J.M.; Pinilla-Buitrago, G.E.; Paz, A.; Johnson, B.A.; Grisales-Betancur, V.; Meenan, S.I.; Attali, D.; Broennimann, O.; Galante, P.J.; Maitner, B.S.; et al. Wallace 2: A shiny app for modeling species niches and distributions redesigned to facilitate expansion via module contributions. Ecography 2023, 2023, 1–9. [Google Scholar] [CrossRef]
- Liu, C.; Newell, G. Selecting thresholds for the prediction of species occurrence with presence-only data. J. Biogeogr. 2013, 40, 778–789. [Google Scholar] [CrossRef]
- Do, M.S.; Son, S.-J.; Choi, G.; Yoo, N.; Koo, K.-S.; Nam, H.-K. Anuran community patterns in the rice fields of the mid-western region of the Republic of Korea. Glob. Ecol. Conserv. 2021, 26, e01448. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Borzée, A.; Shin, Y.; Bae, Y.; Jeong, D.; Amin, H.; Min, M.-S.; Othman, S.N. From Korean to northeast Asian endemicity: On the occurrence of Pelophylax chosenicus in China. Front. Biogeogr. 2024, 16, e62301. [Google Scholar]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Dolgener, N.; Freudenberger, L.; Schluck, M.; Schneeweiss, N.; Ibisch, P.L.; Tiedemann, R. Environmental niche factor analysis (ENFA) relates environmental parameters to abundance and genetic diversity in ane amphibian, the fire-bellied-toad (Bombina bombina). Conserv. Genet. 2014, 15, 11–21. [Google Scholar] [CrossRef]
- Dubois, A.; Ohler, A. Frogs of the Subgenus Pelophylax (Amphibia, Anura, Genus Rana): A catalogue of available and valid scientific names, with comments on name-bearing types, complete synonymies, proposed common names, and maps showing all type localities. Zool. Pol. 1994, 39, 139–204. [Google Scholar]
- Matsui, M.; Maeda, N. Frogs and Toads of Japan; Bun-Ichi Sogo Shuppan: Tokyo, Japan, 1990. [Google Scholar]
- Hirai, T.; Matsui, M. Feeding relationships between Hyla japonica and Rana nigromaculata in rice fields of Japan. J. Herpetol. 2002, 36, 662–667. [Google Scholar] [CrossRef]
- Kanai, R.; Funao, T.; Nakanishi, K.; Sawad, H. Distribution survey of frogs by its call in paddy fields in Shiga, central Japan. Jpn. J. Environ. Entomol. 2014, 25, 121–126. [Google Scholar]
- Hirai, T.; Matsui, M. Feeding habits of the pond frog, Rana nigromaculata, inhabiting rice fields in kyoto, Japan. Copeia 1999, 1999, 940–947. [Google Scholar] [CrossRef]
- Togane, D.; Fukuyama, K.; Takai, K.; Kuramoto, N. Body size and age structure in two populations of Tokyo Daruma Pond Frog, Pelophylax porosus porosus. Curr. Herpetol. 2018, 37, 58–68. [Google Scholar] [CrossRef]
- Shimoyama, R. Preliminary report on male territoriality in the pond frog, Rana nigromaculata, in the breeding season. Jpn. J. Herpetol. 1982, 9, 99–102. [Google Scholar] [CrossRef] [PubMed]
- IUCN SSC Red List of Treatened Species. Pelophylax nigromaculatus (amended version of 2020 assessment). In The IUCN Red List of Threatened Species 2023; e.T58679A63862076; ICUN: Gland, Switzerland, 2023. [Google Scholar]
- Tsuji, M.; Ushimaru, A.; Osawa, T.; Mitsuhashi, H. Paddy-associated frog declines via urbanization: A test of the dispersal-dependent-decline hypothesis. Landsc. Urban Plan. 2011, 103, 318–325. [Google Scholar] [CrossRef]
- Peters, W.C.H. Über Fledertheire (Pteropus gouldii, Rhinolophus deckenii, Vespertilio lobipes, Vesperugo temminckii) und Amphibien (Hypsilurus godeffroyi, Lygosoma scutatum, Stenostoma narisostre, Onychocephalus unguirostris, Ahaetulla polylepis, Pseudechis scutellatus); Monatsberichte der Königlichen Preussische Akademie des Wissenschaften zu Berlin: Berlin, Germany, 1867; pp. 703–712. [Google Scholar]
- Peter, C.; Norliza, A.; Ngeian, J.; Tuen, A.A.; Minton, G. Naturalists, Explorers and Field Scientists in South-East Asia and Australasia; Springer: Cham, Switzerland, 2016. [Google Scholar]
- Komaki, S.; Kurabayashi, A.; Islam, M.M.; Tojo, K.; Sumida, M. Distributional change and epidemic introgression in overlapping areas of Japanese Pond Frog species over 30 Years. Zoolog. Sci. 2012, 29, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Liu, H.T.; Ji, Y.J.; Shi, C.M.; Yang, Y.H.; Zhang, D.X. Black-Spotted Pond Frog (Pelophylax nigromaculatus) on the chinese loess plateau represents a cryptic species: Evidence from polecular phylogeny and ecological niche modeling. J. Syst. Evol. 2015, 53, 339–350. [Google Scholar] [CrossRef]
- Zhang, H.; Yan, J.; Zhang, G.; Zhou, K. Phylogeography and demographic history of Chinese Black-Spotted Frog populations (Pelophylax nigromaculata): Evidence for independent refugia expansion and secondary contact. BMC Evol. Biol. 2008, 8, 21. [Google Scholar] [CrossRef]
- Liu, K.; Wang, F.; Chen, W.; Tu, L.; Min, M.S.; Bi, K.; Fu, J. Rampant historical mitochondrial genome introgression between two species of green pond frogs, Pelophylax nigromaculatus and P. plancyi. BMC Evol. Biol. 2010, 10, 201. [Google Scholar] [CrossRef] [PubMed]
- Groffen, J.; Andersen, D.; Borzée, A. Breeding phenology and landscape use in all amphibian species from the Republic of Korea based on open-source data. Front. Environ. Sci. 2022, 10, 846481. [Google Scholar] [CrossRef]
- Yoo, E.; Yikweon, J. Abiotic effects on calling phenology of three frog species in Korea. Anim. Cells Syst. 2011, 16, 1–8. [Google Scholar] [CrossRef]
- Yoon, I.B.; Kim, J.I.; Yang, S.Y. Study on the food habits of Rana nigromaculata Hallowell and Rana plancyi chosenica Okada (Salientia; Ranidae) in Korea. Korean J. Environ. Biol. 1998, 16, 69–76. [Google Scholar]
- Borzée, A.; Litvinchuk, S.N.; Ri, K.; Andersen, D.; Nam, T.Y.; Jon, G.H.; Man, H.S.; Choe, J.S.; Kwon, S.; Othman, S.N.; et al. Update on distribution and conservation status of amphibians in the Democratic People’s Republic of Korea: Conclusions based on field surveys, environmental modelling, molecular analyses and call properties. Animals 2021, 11, 2057. [Google Scholar] [CrossRef] [PubMed]
- Min, M.; Yan, H.; Zhiping, M.; Yanhong, L.; Zhou, C. Skeletochronological study of age, longevity and growth in a population of Rana nigromaculata (Amphibia: Anura) in Sichuan, China. Asian Herpetol. Res. 2012, 3, 258–264. [Google Scholar]
- Liao, W.B.; Zhou, C.; Yang, Z.; Hu, J.; Lu, X. Age, size and growth in two populations of the Dark-Spotted Frog Rana nigromaculata at different altitudes in southwestern China. Herpetol. J. 2010, 20, 77–82. [Google Scholar]
- Park, J.-K.; Chung, K.; Kim, J.; Do, Y. Population structure and morphological pattern of the Black-Spotted Pond Frog (Pelophylax nigromaculatus) inhabiting watershed areas of the Geum River in South Korea. Sustainability 2022, 14, 16530. [Google Scholar] [CrossRef]
- Huang, M.-Y.; Zhao, Q.; Duan, R.-Y.; Liu, Y.; Wan, Y.-Y. The effect of atrazine on intestinal histology, microbial community and short chain fatty acids in Pelophylax nigromaculatus tadpoles. Environ. Pollut. 2021, 288, 117702. [Google Scholar] [CrossRef]
- Liu, P.-Y.; Meng, T.; Li, Y.; Cai, M.; Li, X.-H.; Chen, J.; Qin, Z.-F. Tetrabromoethylcyclohexane affects gonadal differentiation and development in the frog Pelophylax nigromaculatus. Aquat. Toxicol. 2017, 192, 40–47. [Google Scholar] [CrossRef]
- Fan, X.; Lin, Z.; Li, X.; Wei, L.; Ding, G.-H. Effects of predation by invasive western mosquitofish (Gambusia affinis) on survival of eggs, embryos and tadpoles of Pelophylax nigromaculatus and Duttaphrynus melanostictus in south China. Asian Herpetol. Res. 2016, 7, 46–52. [Google Scholar]
- Groffen, J.; Kong, S.; Yikweon, J.; Borzée, A. The invasive american Bullfrog (Lithobates catesbeianus) in the Republic of Korea: History and recommendations for population control. Manag. Biol. Invasions 2019, 10, 517–535. [Google Scholar] [CrossRef]
- Shin, Y.; Kim, K.; Groffen, J.; Woo, D.; Song, E.; Borzée, A. Citizen science and roadkill trends in the Korean herpetofauna: The importance of spatially biased and unstandardized Data. Front. Ecol. Evol. 2022, 10, 944318. [Google Scholar] [CrossRef]
- Takahashi, K.; Takeuchi, H. Selection of calling sites by Pelophylax porosus porosus (Anura: Ranidae). Acta Biol. 2021, 28, 41–51. [Google Scholar] [CrossRef]
- Matsushima, N.; Hasegawa, M.; Nishihiro, J. Effects of landscape heterogeneity at multiple spatial scales on paddy field-breeding frogs in a large alluvial Plain in Japan. Wetlands 2022, 42, 106. [Google Scholar] [CrossRef]
- IUCN SSC Amphibian Specialist Group. Pelophylax porosus (amended version of 2020 assessment). In The IUCN Red List of Threatened Species 2023; e.T58699A179261024; ICUN: Gland, Switzerland, 2023. [Google Scholar]
- Naito, R.; Yamasaki, M.; Lmanishi, A.; Natuhara, Y.; Morimoto, Y. Effects of water management, connectivity, and surrounding land use on habitat use by frogs in rice paddies in Japan. Zoolog. Sci. 2012, 29, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Okochi, I. Captive breeding and the re-inforcement to an endangered population of Rana porosa brevipoda Ito (Ranidae: Amphibia). Jpn. J. Conserv. Ecol. 1997, 2, 135–146. [Google Scholar]
- Naito, J.; Sakamura, A.; Nakayama, T.; Matsubara, C. The conservation on the Daruma Pond Frog (Rana porosa brevipoda) in biotope area of Haizuka Dam. Hibakagaku 2014, 250, 1–27. [Google Scholar]
- Liu, C. Amphibians of Western China; Chicago Natural History Museum: Chicago, IL, USA, 1950; Volume 2, ISBN 0430-3776. [Google Scholar]
- Zhou, S.-B.; Zhang, Z.-H.; Hui, W.; Qing, M.; Xu, S.-J.; Guan, P.; Shi, J.-S. New record species of anura from China—Pelophylax chosenicus. Chin. J. Zool. 2022, 57, 440–446. [Google Scholar]
- Zhou, S.; He, L.; Ma, S.; Xu, S.; Zhai, Q.; Guan, P.; Wang, H.; Shi, J. Taxonomic status of Rana nigromaculata mongolia and the validity of Pelophylax tenggerensis (Anura, Ranidae). Zootaxa 2022, 5165, 486–500. [Google Scholar] [CrossRef]
- Duan, R.Y.; Kong, X.Q.; Huang, M.Y.; Varela, S.; Ji, X. The potential effects of climate change on amphibian distribution, range fragmentation and turnover in China. PeerJ 2016, 2016, e2185. [Google Scholar] [CrossRef] [PubMed]
- Mou, Y.; Zhao, E.M.; Jiang, Y.M. A Study of vocalization on thirteen species of four genera, Anura. Herpetology 1992, 15, 26. [Google Scholar]
- Zhou, S.-B.; Zhang, Q.-Y.; Hu, Z.-Q.; Xia, Z.-Y.; Miao, Q.; Guan, P.; Shi, J.-S. The validity of Pelophylax chosenicus (Okada, 1931) and P. hubeiensis (Fei & Ye, 1982) (Amphibia, Ranidae). Herpetozoa 2023, 36, 143–152. [Google Scholar] [CrossRef]
- Borzée, A.; Ren, J.-L.; Li, J.-T.; Groffen, J.; Yang, Y.; Jang, Y. Microhabitat segregation between Black-Spotted Pond Frogs, Pelophylax nigromaculatus, and Gold-Striped Pond Frogs, P. plancyi (Anura: Ranidae). Reptil. Amphib. 2019, 26, 119–120. [Google Scholar] [CrossRef]
- IUCN SSC Amphibian Specialist Group. Pelophylax plancyi (amended version of 2020 assessment). In The IUCN Red List of Threatened Species 2023; e.T58699A179261024; ICUN: Gland, Switzerland, 2023. [Google Scholar]
- Borzée, A.; Kyong, C.N.; Kil, H.K.; Jang, Y. Impact of water quality on the occurrence of two endangered Korean anurans: Dryophytes suweonensis and Pelophylax chosenicus. Herpetologica 2018, 74, 1–7. [Google Scholar] [CrossRef]
- Wei, X.; Huang, M.; Yue, Q.; Ma, S.; Li, B.; Mu, Z.; Peng, C.; Gao, W.; Liu, W.; Zheng, J.; et al. Long-term urbanization impacts the Eastern Golden Frog (Pelophylax plancyi) in Shanghai City: Demographic history, genetic structure, and implications for amphibian conservation in intensively urbanizing environments. Evol. Appl. 2021, 14, 117–135. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Li, B.; Shu, X.; Pei, E.; Yuan, X.; Sun, Y.; Wang, T.; Wang, Z. Responses of anuran communities to rapid urban growth in Shanghai, China. Urban For. Urban Green. 2016, 20, 365–374. [Google Scholar] [CrossRef]
- Shu, X.; Zhang, W.; Li, B.; Pei, E.; Yuan, X.; Wang, T.; Wang, Z. Major factors affecting the distribution of anuran communities in the urban, suburban and rural areas of Shanghai, China. Asian Herpetol. Res. 2016, 7, 287–294. [Google Scholar]
- Okada, Y. Frogs in Korea. J. Chosen Biol. Soc. 1928, 6, 15–46. [Google Scholar]
- Andersen, D.; Chuang, M.F.; Choe, M.; Kim, A.; Kwon, S.; Jang, Y.; Borzée, A. Elevational distribution of amphibians: Resolving distributions, patterns, and species communities in the Republic of Korea. Zool. Stud. 2022, 61, e25. [Google Scholar]
- Ra, N.Y.; Sung, H.C.; Cheong, S.K.; Lee, J.H.; Eom, J.; Park, D. Habitat use and home range of the endangered Gold-Spotted Pond Frog (Rana chosenica). Zoolog. Sci. 2008, 25, 894–903. [Google Scholar] [CrossRef] [PubMed]
- Eom, J.; Lee, J.-B.; Ra, N.-Y.; Park, D.-S. Preferred feeding sites and prey of the adult Gold-Spotted Pond Frog, Rana plancyi chosenica. J. Ecol. Environ. 2007, 30, 357–361. [Google Scholar] [CrossRef]
- Koo, K.S.; Kwon, S.; Kwak, M.; Oh, J. First record of heterospecific amplexus behaviour between Pelophylax chosenicus (Anura: Ranidae) and Dryophytes japonicus (Anura: Hylidae) in Paju, Republic of Korea. Herpetol. Notes 2021, 14, 1225–1226. [Google Scholar]
- IUCN SSC Amphibian Specialist Group. Pelophylax chosenicus (amended version of 2020 assessment). In The IUCN Red List of Threatened Species 2023; e.T58699A179261024; ICUN: Gland, Switzerland, 2023. [Google Scholar]
- Park, C.-D.; Kwon, K.; Yoo, N.; Lee, J.-H.; Kang, D.; Park, J.-S.; Yoo, J.; Kim, K.-S.; Yoon, J.-D. Post-release monitoring after reintroduction of captive-reared Korean endangered Frog, Pelophylax chosenicus. Proc. Natl. Inst. Ecol. Repub. Korea 2021, 2, 114–119. [Google Scholar]
- Yoo, N.; Do, M.S.; Nam, H.-K.; Choi, G.; Son, S.-J.; Yoo, J. Habitat characteristics of anuran species inhabiting rice fields of western Mid-South Korea. Korean J. Ecol. Environ. 2019, 52, 366–377. [Google Scholar] [CrossRef]
- Fong, J.J.; Cheng, T.L.; Bataille, A.; Pessier, A.P.; Waldman, B.; Vredenburg, V.T. Early 1900s detection of Batrachochytrium dendrobatidis in Korean amphibians. PLoS ONE 2015, 10, e0115656. [Google Scholar] [CrossRef] [PubMed]
- Fei, L.; Ye, C. The distributional characteristics of amphibian in Hubei province, including description of a new frog. Acta Zool 1982, 28, 293–301. [Google Scholar]
- IUCN SSC Amphibian Specialist Group. Pelophylax hubeiensis (amended version of 2020 assessment). In The IUCN Red List of Threatened Species 2023; e.T58699A179261024; ICUN: Gland, Switzerland, 2023. [Google Scholar]
- Pope, C. Four new frogs from Fukien Province, China. Am. Museum Novit. 1929, 1–5. [Google Scholar]
- Eo, S.H.E.; Lee, B.J.; Park, C.D.; Jung, J.H.; Hong, N.; Lee, W.S. Taxonomic identity of the Glandirana emeljanovi (Anura, Ranidae) in Korea revealed by the complete mitochondrial genome sequence analysis. Mitochondrial DNA Part B Resour. 2019, 4, 961–962. [Google Scholar] [CrossRef]
- Jang-Liaw, N.-H.; Chou, W.-H. Anuran fauna of Taiwan and adjacent islands based on valid specimen records. Coll. Res. 2015, 28, 5–53. [Google Scholar]
- Sumida, M.; Ogata, M.; Nishioka, M. Molecular phylogenetic relationships of pond frogs distributed in the Palearctic region inferred from DNA sequences of mitochondrial 12S ribosomal RNA and cytochrome b genes. Mol. Phylogenet. Evol. 2000, 16, 278–285. [Google Scholar] [CrossRef]
- IUCN SSC Amphibian Specialist Group. Pelophylax fukienensis (amended version of 2020 assessment). In The IUCN Red List of Threatened Species 2023; e.T58699A179261024; ICUN: Gland, Switzerland, 2023. [Google Scholar]
- Arif, S.M. Endoparasitic nematohelminthes collected from inside frog Rana ridibunda (Pallas, 1771) in two rivers near Baghdad/Iraq. J. Pharm. Sci. Res. 2019, 11, 2080–2083. [Google Scholar]
- Jablonski, D.; Van Bemmel, S.; Masroor, R. Notes on the herpetofauna of Kunduz Province, northeastern Afghanistan, with the first record of Varanus griseus caspius (Eichwald, 1831). Herpetol. Notes 2021, 14, 63–66. [Google Scholar]
- Kaptyonkina, A.G.; Dujsebayeva, T.N.; Akhmedenov, K.M.; Khromov, V.A.; Krainyuk, V.N.; Sarzhanov, F.; Starikov, S.V.; Tarasovskaya, N.E.; Timoshenko, A.Y.; Titov, S.V. The range of Marsh Frogs (Complex Pelophylax ridibundus, Amphibia, Ranidae) in Kazakhstan: Progressive dispersal or cyclic fluctuations? Proc. Zool. Inst. Russ. Acad. Sci. 2022, 326, 211–238. [Google Scholar] [CrossRef]
- Khalaf. Reptiles of Iraq: With Some Notes on the Amphibians; Ar-Rabitta Press: Charlottesville, VA, USA, 1959. [Google Scholar]
- Mohammadi, Z.; Khajeh, A.; Ghorbani, F.; Kami, H.G. A biosystematic sudy of new records of the Marsh Frog Pelophylax ridibundus (Pallas, 1771) (Amphibia: Ranidae) from southeast of Iran. J. Asia-Pac. Biodivers. 2015, 31, 178–182. [Google Scholar] [CrossRef]
- Papežík, P.; Sciberras, A.; Benovics, M.; Sciberras, J.; Deidun, A.; Mikulicek, P. Far from home: Tracing the origin of non-native water frogs (Genus Pelophylax) in Malta by molecular markers. Biol. Invasions 2024, 26, 1045–1059. [Google Scholar] [CrossRef]
- Plötner, J. Die Westpaläarktischen Wasserfrösche: Von Märtyrern der Wissenschaft zur Biologischen Sensation; Laurenti: Bielefeld, Germany, 2005. [Google Scholar]
- Sánchez-Montes, G.; Recuero, E.; Gutiérrez-Rodríguez, J.; Gomez-Mestre, I.; Martinez-Solano, I. Species assignment in the Pelophylax ridibundus × P.perezi hybridogenetic complex based on 16 newly characterized microsatellite markers. Herpetol. J. 2016, 26, 99–108. [Google Scholar]
- Ualiyeva, D.; Ermakov, O.A.; Litvinchuk, S.N.; Guo, X.; Ivanov, A.Y.; Xu, R.; Li, J.; Xu, F.; Arifulova, I.I.; Kaptyonkina, A.G.; et al. Diversity, phylogenetic relationships and distribution of Marsh Frogs (the Pelophylax ridibundus complex) from Kazakhstan and northwest China. Diversity 2022, 14, 869. [Google Scholar] [CrossRef]
- Bayrakci, Y.; Çiçek, K. Breeding ecology and larval development of Marsh Frogs (Pelophylax ridibundus s.l.) from East Mediterranean, Türkiye. Commagene J. Biol. 2023, 7, 65–72. [Google Scholar] [CrossRef]
- Baran, İ.; Atatür, M.K. Turkish Herpetofauna: Amphibians and Reptiles; Turkey Ministry of Environment: Ankara, Turkey, 1997. [Google Scholar]
- Borkin, L. Cryptic species (a case study of amphibians). Entomol. Rev 2004, 84, 75–98. [Google Scholar]
- Dufresnes, C.; Leuenberger, J.; Amrhein, V.; Bühler, C.; Thiébaud, J.; Bohnenstengel, T.; Dubey, S. Invasion genetics of Marsh Frogs (Pelophylax ridibundus sensu lato) in Switzerland. Biol. J. Linn. Soc. 2018, 123, 402–410. [Google Scholar] [CrossRef]
- Dujsebayeva, T.N.; Ivanov, A.Y.; Kaptyonkina, A.G.; Ualiyeva, D.A.; Krainyuk, V.N.; Cherednichenko, A.V.; Khromov, V.A. The Marsh Frogs (Pelophylax ridibundus complex) in central Kazakhstan: Expansion and retreat. Russ. J. Ecosyst. Ecol. 2021, 6, 1–18. [Google Scholar] [CrossRef]
- Holsbeek, G.; Jooris, R. Potential impact of genome exclusion by alien species in the hybridogenetic Water Frogs (Pelophylax esculentus complex). Biol. Invasions 2010, 12, 1–13. [Google Scholar] [CrossRef]
- Hotz, H.; Beerli, P.; Uzzell, T.; Guex, G.-D.; Pruvost, N.; Schreiber, R.; Plötner, J. Balancing a cline by influx of migrants: A genetic transition in Water Frogs of eastern Greece. J. Hered. 2012, 104, 57–71. [Google Scholar] [CrossRef]
- Jablonski, D.; Khalili, F.; Masroor, R. The herpetofaunal diversity of Takhar Province, Afghanistan. Herpetozoa 2023, 90, 73–90. [Google Scholar]
- Domeneghetti, D.; Bruni, G.; Fasola, M.; Bellati, A. Discovery of alien Water Frogs (gen. Pelophylax) in Umbria, with first report of P. shqipericus for Italy. Acta Herpetol. 2013, 8, 171–176. [Google Scholar]
- Laghi, P.; Miserocchi, D.; Valli, M. Determinazione genetica della presenza delle rane verdi alloctone Pelophylax ridibundus e Pelophylax kurtmuelleri (Amphibia, Anura, Ranidae) in due località della Romagna. Quad. Mus. Stor. Nat. Ferrara 2013, 1, 75–78. [Google Scholar]
- Pagano, A.; Joly, P.; Plénet, S.; Lehman, A.; Grolet, O. Breeding habitat partitioning in the Rana esculenta complex: The intermediate niche hypothesis supported. Ecoscience 2001, 8, 294–300. [Google Scholar] [CrossRef]
- Dubey, S.; Leuenberger, J.; Perrin, N. Multiple origins of invasive and ‘native’ Water Frogs (Pelophylax spp.) in Switzerland. Biol. J. Linn. Soc. 2014, 112, 442–449. [Google Scholar] [CrossRef]
- Holsbeek, G.; Mergeay, J.; Hotz, H.; Plötner, J.; Volckaert, F.; De Meester, L. A cryptic invasion within an invasion and widespread introgression in the European Water Frog complex: Consequences of uncontrolled commercial trade and weak international legislation. Mol. Ecol. 2008, 17, 5023–5035. [Google Scholar] [CrossRef]
- Holsbeek, G.; Mergeay, J.; Volckaert, F.; De Meester, L. Genetic detection of multiple exotic Water frog species in Belgium illustrates the need for monitoring and immediate action. Biol. Invasions 2010, 12, 1459–1463. [Google Scholar] [CrossRef]
- Ermakov, O.; Ivanov, A.; Titov, S.; Svinin, A.; Litvinchuk, S.N. New Multiplex PCR method for identification of East European Green Frog species and their hybrids. Russ. J. Herpetol. 2019, 26, 367–370. [Google Scholar] [CrossRef]
- Sah, A.K.; Gautam, B.; Bhattarai, S. Biological aspects of the Marsh Frog Pelophylax ridibundus population from the ahadegan wetland, Khuzestan Province, Iran. J. Anim. Divers. 2020, 2, 18–26. [Google Scholar]
- Lukanov, S.P.; Tzankov, N.D.; Simeonovska-Nikolova, D.M. Effects of environmental factors on mating call characteristics of the Marsh Frog Pelophylax ridibundus (Pallas, 1771) (Amphibia: Ranidae) in Bulgaria. Acta Zool. Bulg. 2014, 66, 209–216. [Google Scholar]
- Balint, N.; Citrea, L.; Memetea, A.; Jurj, N.; Condure, N. Feeding biology of the Pelophylax ridibundus Pallas 1771, (Anura, Ranidae) in Dobromir, Romania. Biharean Biol. 2008, 2, 27–37. [Google Scholar]
- Najibzadeh, M.; Darvish, J. Comparison of habitat, mating behavior and laying of three species of anuran amphibians (Rana (Pelophylax) ridibunda, Hyla savignyi and Bufo (Pseudepidalea) variabilis) in the Lorestan Province. J. Anim. Res. 2014, 27, 291–299. [Google Scholar]
- Erismis, U. Abundance, demography and population structure of Pelophylax ridibundus (Anura: Ranidae) in 26-August National Park (Turkey). North. West. J. Zool. 2011, 7, 5–16. [Google Scholar]
- Altun, C.; Altunışık, A. Comparison of the Marsh Frog (Pelophylax ridibundus) populations living in different altitudes in terms of age, size, and some growth parameters. Biol. Bull. 2023, 2023, 1–8. [Google Scholar]
- IUCN SSC Amphibian Specialist Group. Pelophylax ridibundus (amended version of 2020 assessment). In The IUCN Red List of Threatened Species 2023; e.T58699A179261024; ICUN: Gland, Switzerland, 2023. [Google Scholar]
- Quilodran, C.; Montoya-Burgos, J.; Currat, M. Modelling interspecific hybridization with genome exclusion to identify conservation actions: The case of native and invasive Pelophylax Waterfrogs. Evol. Appl. 2015, 8, 199–210. [Google Scholar] [CrossRef]
- Quilodran, C.; Austerlitz, F.; Currat, M.; Montoya-Burgos, J. Cryptic biological invasions: A general model of hybridization. Sci. Rep. 2018, 8, 2414. [Google Scholar] [CrossRef]
- Zhelev, Z.; Tsonev, S.; Georgieva, K. Health status of Pelophylax ridibundus (Amphibia: Ranidae) in a rice paddy ecosystem in southern Bulgaria and its importance in assessing environmental state: Haematological parameters. Environ. Sci. Pollut. Res. 2019, 25, 7884–7895. [Google Scholar] [CrossRef]
- Zhelev, Z.; Tsonev, S.; Boyadz, P. Using of fluctuating asymmetry in adult Pelophylax ridibundus (Amphibia: Anura:Ranidae) meristic traits as a method for a developmental stability of populationand environmental quality of their habitat: Industrial area in southern Bulgaria. Turkish J. Zool. 2022, 46, 220–227. [Google Scholar] [CrossRef]
- Lukanov, S.; Popgeorgiev, G.; Tzankov, N. First bioacoustic and morphological data for the presence of Pelophylax bedriagae in Bulgaria. Acta Sci. Nat. 2018, 5, 54–63. [Google Scholar] [CrossRef]
- Schneider, H. Calls and reproductive behaviour of the Water Frogs of Damascus, Syria (Amphibia: Anura: Rana Bedriagae Camerano, 1882). Zool. Middle East 1997, 15, 51–66. [Google Scholar] [CrossRef]
- Pesarakloo, A.; Najibzadeh, M.; Rastegar-Pouyani, N.; Rastegar-Pouyani, E. Taxonomic survey of Water Frog populations of Pelophylax bedriagae (Anura: Ranidae) in Western Iran: A morphometric and bioacoustic approach. Biologia 2018, 73, 673–681. [Google Scholar] [CrossRef]
- Schneider, H. Calls of the Levantine Frog, Rana bedriagae, at Birket Ata, Israel (Amphibia: Anura). Zool. Middle East 1999, 19, 101–116. [Google Scholar] [CrossRef]
- Plötner, J.; Akın Pekşen, Ç.; Baier, F.; Uzzell, T.; Bilgin, C. Genetic evidence for human-mediated introduction of Anatolian Water Frogs (Pelophylax cf. bedriagae) to Cyprus (Amphibia: Ranidae). Zool. Middle East 2015, 61, 125–132. [Google Scholar] [CrossRef]
- Goldberg, S. Notes on reproduction of Levant Green Frogs, Pelophylax bedriagae (Anura: Ranidae), from Israel. Bull. Chic. Herpetol. Soc. 2023, 58, 26–27. [Google Scholar]
- Dufresnes, C.; Denoël, M.; Di Santo, L.; Dubey, S. Multiple uprising invasions of Pelophylax Water Frogs, potentially inducing a new hybridogenetic complex. Sci. Rep. 2017, 7, 6506. [Google Scholar] [CrossRef]
- Ibrahim, A. First Record of Pelophylax bedriagae (Amphibia: Ranidae) in the Suez Canal area, Egypt. Herpetol. Notes 2011, 4, 331–332. [Google Scholar]
- Pesarakloo, A.; Najibzadeh, M. Life history of the Levant Water Frog, Pelophylax bedriagae (Amphibia: Anura: Ranidae) in Western Iran. J. Anim. Divers. 2019, 1, 11–19. [Google Scholar] [CrossRef]
- Pesarakloo, A.; Rastegar-Pouyani, E.; Rastegar-Pouyani, N.; Kami, H.; Najibzadeh, M.; Khosravani, A.; Oraie, H. The first taxonomic revaluation of the Iranian Water Frogs of the genus Pelophylax (Anura: Ranidae) using sequences of the mitochondrial genome. Mitochondrial DNA Part A DNA Mapp. Seq. Anal. 2017, 28, 392–398. [Google Scholar] [CrossRef] [PubMed]
- Khaloei, M.; Fathinia, B.; Shafaeipour, A. Skeletochronology of the Levant Green Frog, Pelophylax bedriagae (Camerano, 1882), in southern Iran. Curr. Herpetol. 2023, 42, 55–63. [Google Scholar] [CrossRef]
- Disi, A.; Amr, Z. Morphometrics, distribution and ecology of the amphibians in Jordan. Vertebr. Zool. 2010, 60, 147–162. [Google Scholar] [CrossRef]
- Özcan, L.; Yorulmaz, B.; Ayaz, D. Data on Food Composition of the Levant Marsh Frog (Pelophylax bedriagae) in Southwestern Anatolia. North. West. J. Zool. 2021, 17, 227–231. [Google Scholar]
- IUCN SSC Amphibian Specialist Group. Pelophylax bedriagae (amended version of 2020 assessment). In The IUCN Red List of Threatened Species 2023; e.T58699A179261024; ICUN: Gland, Switzerland, 2023. [Google Scholar]
- Başkale, E.; Kaya, U. Decline of the Levantine Frog, Pelophylax bedriagae Camerano, 1882, in the western Aegean region of Turkey changes in population size and implications for conservation: (Amphibia: Ranidae). Zool. Middle East. 2012, 57, 69–76. [Google Scholar] [CrossRef]
- Innal, D.; Sozbilen, D.; Ozmen, O. Prevalence and pathology of Lernaea cyprinacea (Crustacea: Lernaeidae) parasitizing tadpoles of Pelophylax bedriagae (Anura: Ranidae) in Antalya, Turkey. Phyllomedusa 2019, 18, 177–183. [Google Scholar] [CrossRef]
- Mezhzherin, S.V. New species of Green Frogs Rana terentievi sp. nova (Amphibia, Ranidae) from South Tajikistan. Proc. Acad. Sci. Ukr. 1992, 5, 150–153. [Google Scholar]
- Pesarakloo, A. Central Asia or Minor Asia: Origin of the Water Frogs in the Southern Coast of the Caspian Sea. Soc. Sci. Res. Netw. 2022, preprint, 4178613. [Google Scholar]
- Mazepa, G. Evolution of Water Frogs Pelophylax in Central Asia: How Hybridization and Mitochondrial Introgression among Ecologically Divergent Species Promote Occupation of Novel Environment. Master’s Thesis, Uppsala University, Uppsala, Sweden, 2013. [Google Scholar]
- Kidov, A.A.; Ivolga, R.A.; Kondratova, T.E.; Ivanov, A.A. Age, Growth, and fertility in Terentiev’s Frog (Pelophylax terentievi, Amphibia, Ranidae). Biol. Bull. 2023, 50, 1854–1863. [Google Scholar] [CrossRef]
- Wagner, P.; Bauer, A.; Leviton, A.; Wilms, T.; Böhme, W. A Checklist of the amphibians and reptiles of Afghanistan—Exploring herpetodiversity using biodiversity archives. Proc. Calif. Acad. Sci. 2016, 63, 457–565. [Google Scholar]
- IUCN SSC Amphibian Specialist Group. Pelophylax terentievi (amended version of 2020 assessment). In The IUCN Red List of Threatened Species 2023; e.T58699A179261024; ICUN: Gland, Switzerland, 2023. [Google Scholar]
- Akin, C.P. Molecular Evolution and Phylogeography of the Eastern Mediterranean Water Frog (Pelophylax) Complex. Ph.D. Thesis, School of Natural and Applied Sciences of Middle East Technical University, Ankara, Turkey, 2015. [Google Scholar]
- Akin, C.; Bilgin, C.C.; Beerli, P.; Westaway, R.; Ohst, T.; Litvinchuk, S.N.; Uzzell, T.; Bilgin, M.; Hotz, H.; Guex, G.D.; et al. Phylogeographic patterns of genetic diversity in eastern mediterranean water frogs were determined by geological processes and climate change in the late cenozoic. J. Biogeogr. 2010, 37, 2111–2124. [Google Scholar] [CrossRef] [PubMed]
- Ualiyeva, D.A.; Ivanov, A.Y.; Ermakov, O.A. A development of a PCR-RFLP test system for the identification of mitochondrial lines of the Pelophylax ridibundus Lake Frog in Kazakhstan. Univ. Proc. Volga Reg. Nat. Sci. 2022, 37, 76–84. [Google Scholar] [CrossRef]
- Lymberakis, P.; Poulakakis, N.; Manthalou, G.; Tsigenopoulos, C.S.; Magoulas, A.; Mylonas, M. Mitochondrial phylogeography of Rana (Pelophylax) populations in the eastern Mediterranean region. Mol. Phylogenet. Evol. 2007, 44, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Andersen, D.; Maslova, I.; Purevdorj, Z.; Li, J.T.; Messenger, K.R.; Ren, J.L.; Jang, Y.; Borzée, A. East palearctic treefrog past and present habitat suitability using ecological niche models. PeerJ 2022, 10, e12999. [Google Scholar] [CrossRef] [PubMed]
- Litvinchuk, S.; Skorinov, D.; Ivanov, A.; Ermakov, O. Detection of glacial refugia and post-glacial colonization routes of morphologically cryptic Marsh Frog species (Anura: Ranidae: Pelophylax) using environmental niche modeling. Diversity 2024, 16, 94. [Google Scholar] [CrossRef]
- Jablonski, D.; Gkontas, I.; Poursanidis, D.; Lymberakis, P.; Poulakakis, N. Stability in the Balkans: Phylogeography of the endemic Greek Stream Frog, Rana graeca. Biol. J. Linn. Soc. 2021, 132, 829–846. [Google Scholar] [CrossRef]
- Olalla-Tárraga, M.; McInnes, L.; Bini, L.; Diniz-Filho, J.A.; Fritz, S.; Hawkins, B.; Hortal, J.; Orme, D.; Rahbek, C.; Rodriguez, M.; et al. Climatic niche conservatism and the evolutionary dynamics in species range boundaries: Global congruence across mammals and amphibians. J. Biogeogr. 2011, 38, 2237–2247. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change. Future Global Climate: Scenario-Based Projections and Near-Term Information; Cambridge University Press: Cambridge, UK, 2023. [Google Scholar]
- Luedtke, J.A.; Chanson, J.; Neam, K.; Hobin, L.; Maciel, A.O.; Catenazzi, A.; Borzée, A.; Hamidy, A.; Aowphol, A.; Jean, A. Ongoing declines for the world’s amphibians in the face of emerging threats. Nature 2023, 622, 308–314. [Google Scholar] [CrossRef]
- Borzée, A.; Heo, K.; Jang, Y. Relationship between agro-environmental variables and breeding Hylids in rice paddies. Sci. Rep. 2018, 8, 8049. [Google Scholar] [CrossRef]
- Naito, R.; Sakai, M.; Morimoto, Y. Negative effects of deep roadside ditches on Pelophylax porosa brevipoda dispersal and migration in comparison with Hyla japonica in a rice paddy area in Japan. Zoolog. Sci. 2012, 29, 599–603. [Google Scholar] [CrossRef] [PubMed]
- Naito, R.; Sakai, M.; Natuhara, Y.; Morimoto, Y.; Shibata, S. Microhabitat use by Hyla japonica and Pelophylax porosa brevipoda at levees in rice paddy areas of Japan. Zoolog. Sci. 2013, 30, 386–391. [Google Scholar] [CrossRef] [PubMed]
- Borzée, A.; Andersen, D.; Groffen, J.; Kim, H.T.; Bae, Y.; Jang, Y. Climate change-based models predict range shifts in the distribution of the only Asian Plethodontid salamander: Karsenia koreana. Sci. Rep. 2019, 9, 11838. [Google Scholar] [CrossRef] [PubMed]
- Dufresnes, C.; Monod-Broca, B.; Bellati, A.; Canestrelli, D.; Ambu, J.; Wielstra, B.; Dubey, S.; Crochet, P.A.; Denoël, M.; Jablonski, D. Piecing the barcoding puzzle of Palearctic Water Frogs (Pelophylax) sheds light on amphibian biogeography and global invasions. Glob. Chang. Biol. 2024, 30, e17180. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | P. ridibundus | P. bedriagae | P. terentievi | Pelophylax “Syr Darya lineage” | P. n. “reinhardtii” | P. n. “nigromaculatus” | P. porosus | P. mongolius | P. plancyi | P. chosenicus | P. hubeiensis | P. fukienensis |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Elevation | 0.6 | 0.8 | 0.9 | 2.2 | 18 | 2.74 | 1 | 59 | 11 | 2.9 | 4.8 | 60 |
| Bio 1 | 5.2 | 3.1 | 17 | 0.1 | 21 | 10.2 | 12 | 2.9 | 2.87 | 3 | 28 | 1 |
| Bio 2 | 4.4 | 1.6 | 8.2 | 11 | 1.5 | 0.6 | 1 | 0.3 | 8.8 | 8.7 | 12 | 0 |
| Bio 3 | 1 | 10.4 | 1.4 | 5.5 | 11 | 0.1 | 3 | 6 | 34.3 | 40 | 25 | 7 |
| Bio 5 | 1 | 5.8 | 0 | 1.7 | 16 | 2.4 | 0 | 2.3 | 1 | 0 | 0 | 2 |
| Bio 8 | 17 | 17.2 | 9 | 0.5 | 1 | 1.1 | 1 | 1 | 14 | 19 | 13 | 1 |
| Bio 9 | 6.1 | 27.8 | 1.3 | 4.9 | 0.6 | 0 | 2 | 0.2 | 1.3 | 1.1 | 1.3 | 0 |
| Bio 11 | 12.8 | 0.1 | 3.6 | 4.3 | 0 | 0 | 1 | 14 | 0.4 | 0.9 | 5.1 | 20 |
| Bio 12 | 4.8 | 12.6 | 10 | 14 | 6 | 0.2 | 3 | 2.9 | 3 | 0.2 | 3.9 | 2 |
| Bio 14 | 39 | 0.3 | 1.1 | 0.9 | 0.7 | 0.1 | 2 | 6 | 18.4 | 23 | 22 | 5 |
| Bio 15 | 1.2 | 1.2 | 10 | 1 | 5 | 16.1 | 12 | 2.1 | 2.63 | 0.2 | 1.2 | 1 |
| Bio 18 | 0.6 | 1.3 | 37 | 9.2 | 0 | 0 | 0 | 0.2 | 0 | 1.3 | 1.5 | 0 |
| Bio 19 | 1 | 1.7 | 0.2 | 34 | 20 | 65.2 | 60 | 0.6 | 1.9 | 0.3 | 1.5 | 1 |
| AUC | 0.992 | 0.985 | 0.994 | 0.976 | 0.895 | 0.953 | 0.997 | 0.98 | 0.99 | 0.997 | 0.981 | 0.997 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amin, H.; Borzée, A. Understanding the Distribution, Behavioural Ecology, and Conservation Status of Asian Pelophylax. Diversity 2024, 16, 259. https://doi.org/10.3390/d16050259
Amin H, Borzée A. Understanding the Distribution, Behavioural Ecology, and Conservation Status of Asian Pelophylax. Diversity. 2024; 16(5):259. https://doi.org/10.3390/d16050259
Chicago/Turabian StyleAmin, Hina, and Amaël Borzée. 2024. "Understanding the Distribution, Behavioural Ecology, and Conservation Status of Asian Pelophylax" Diversity 16, no. 5: 259. https://doi.org/10.3390/d16050259
APA StyleAmin, H., & Borzée, A. (2024). Understanding the Distribution, Behavioural Ecology, and Conservation Status of Asian Pelophylax. Diversity, 16(5), 259. https://doi.org/10.3390/d16050259