
Tracking the Recovery of Freshwater Mussel Diversity in Ontario Rivers: Evaluation of a Quadrat-Based Monitoring Protocol

Abstract
:1. Introduction
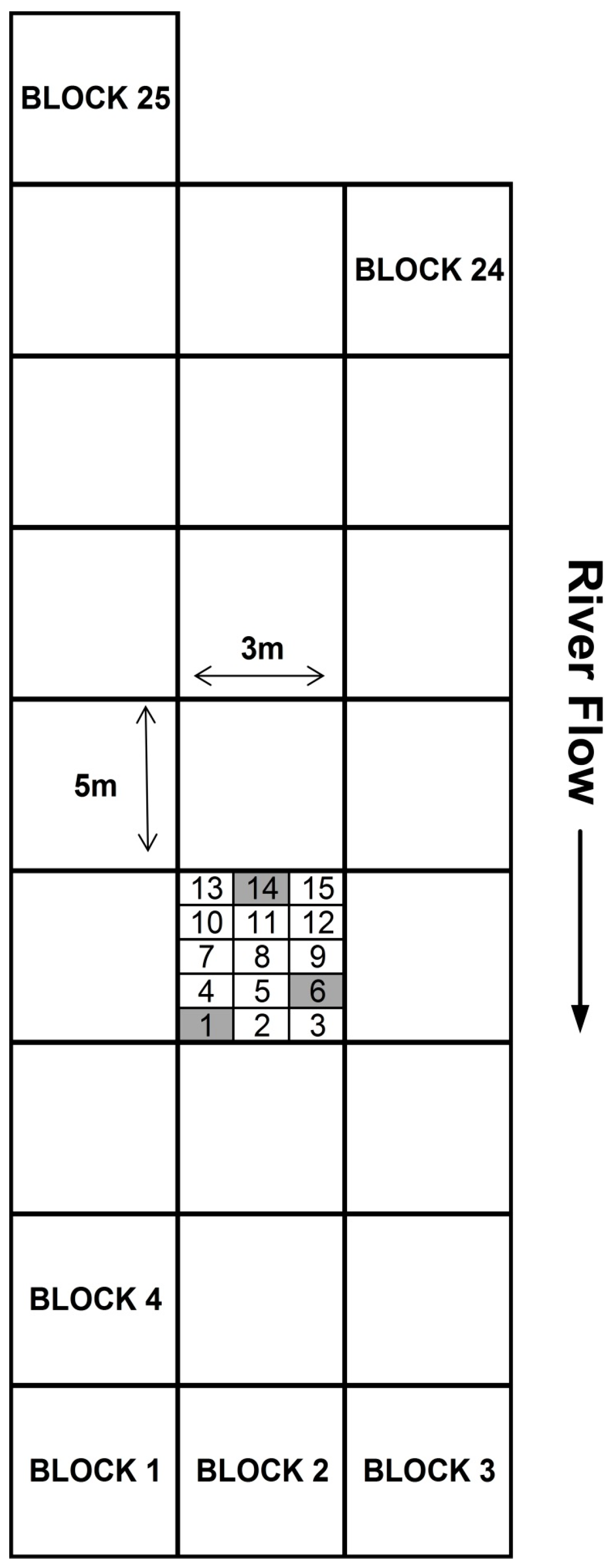
2. Materials and Methods
3. Data Analysis
3.1. Multi-River Dataset
3.2. Census Dataset
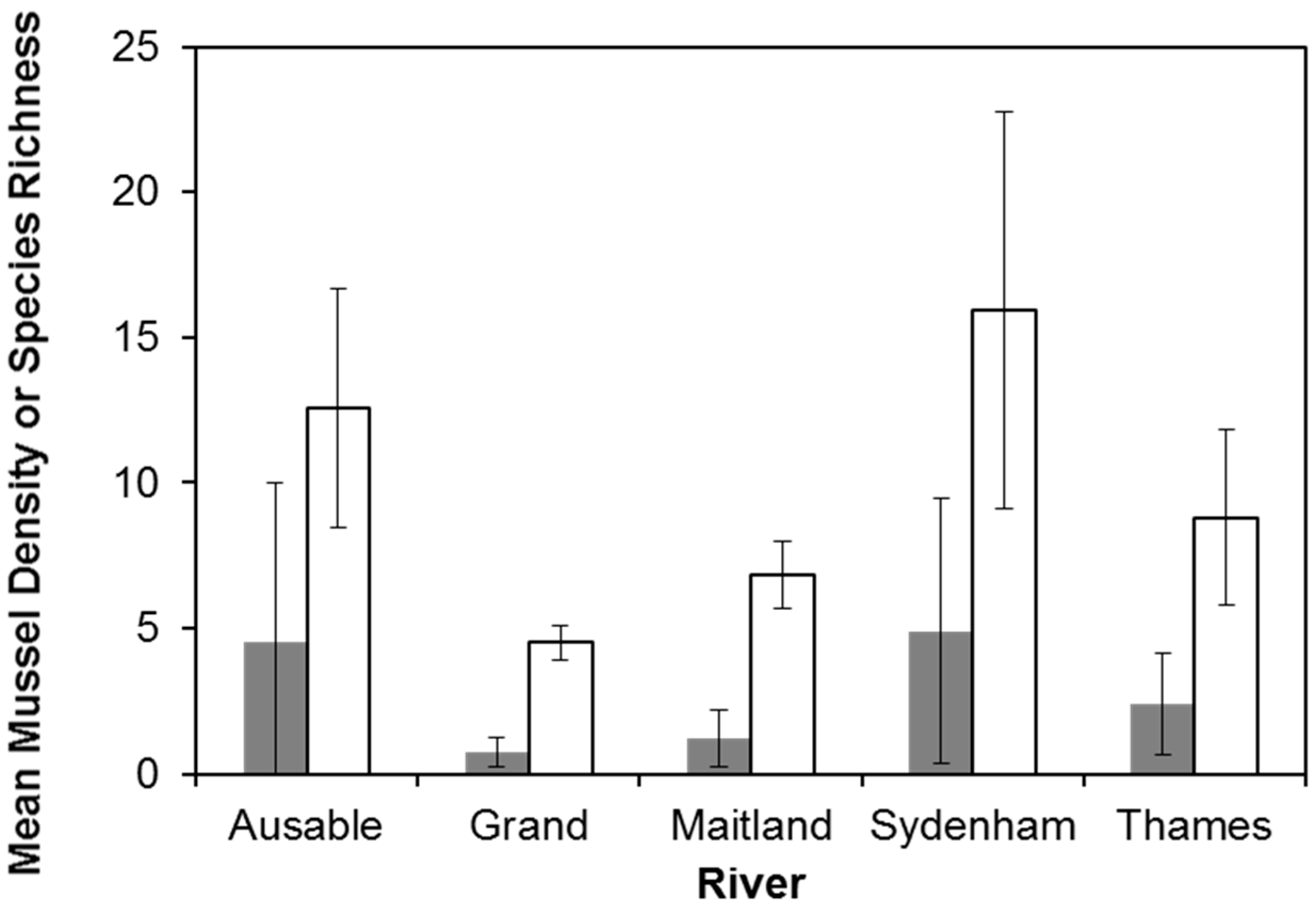
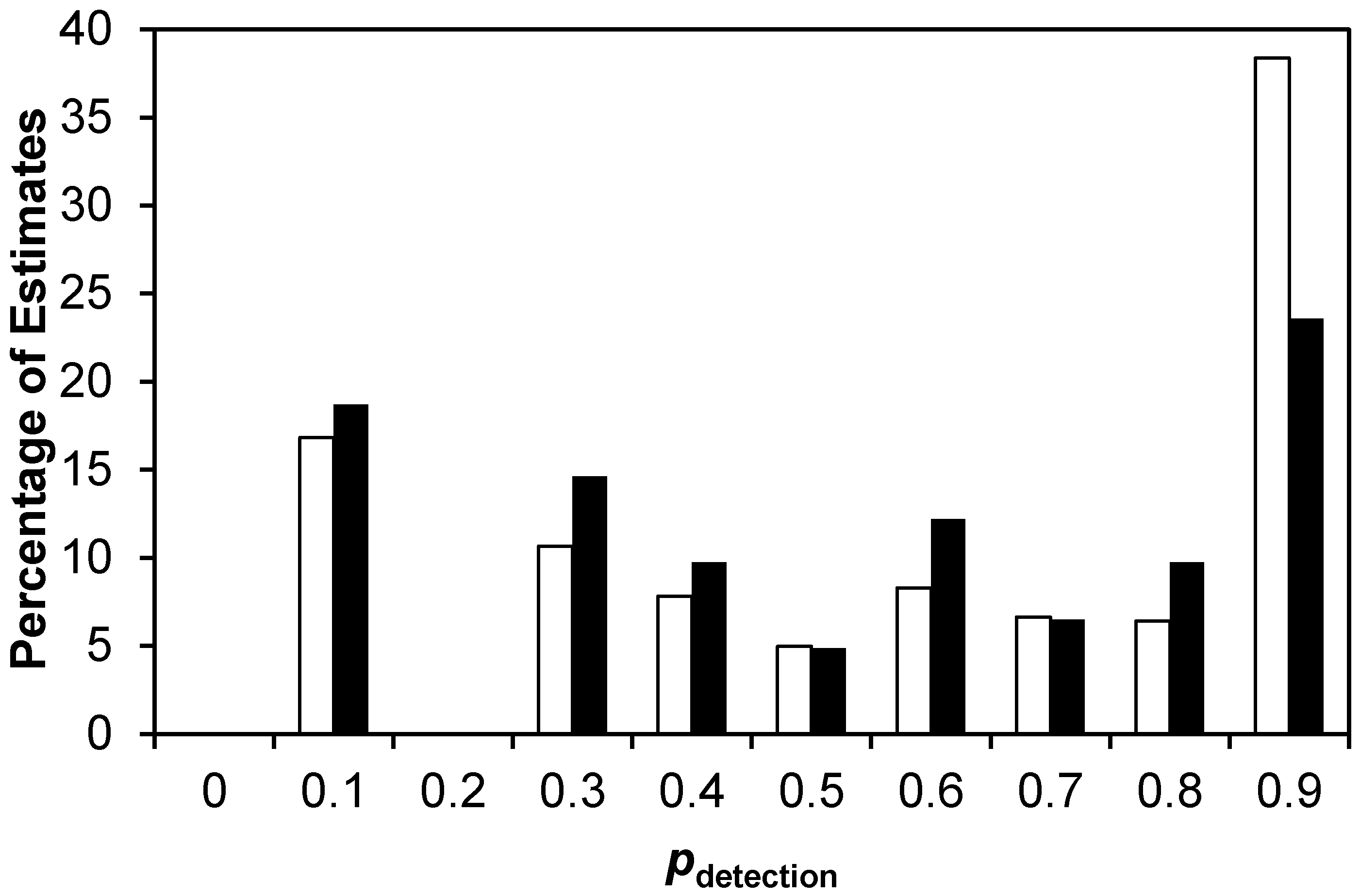
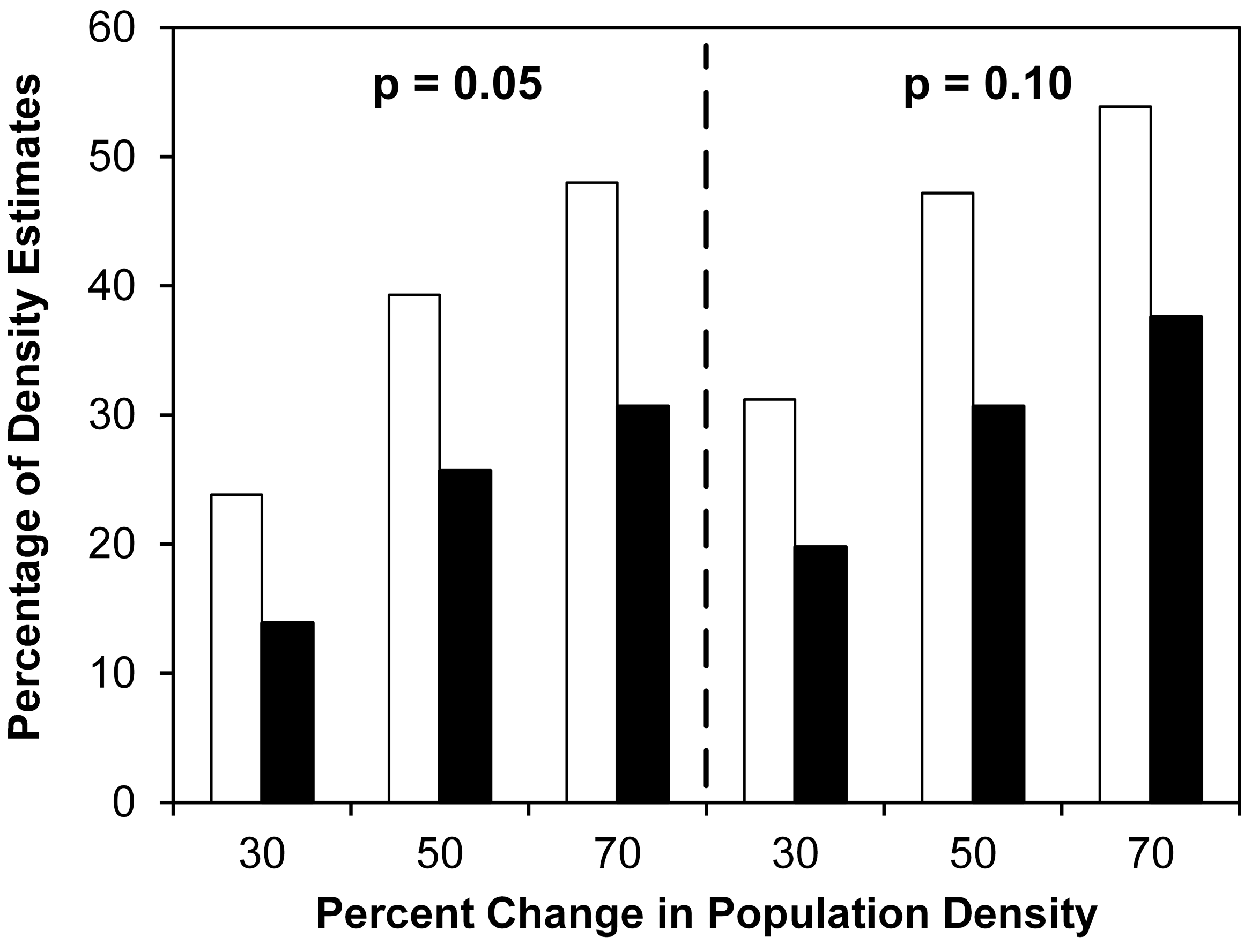
4. Results
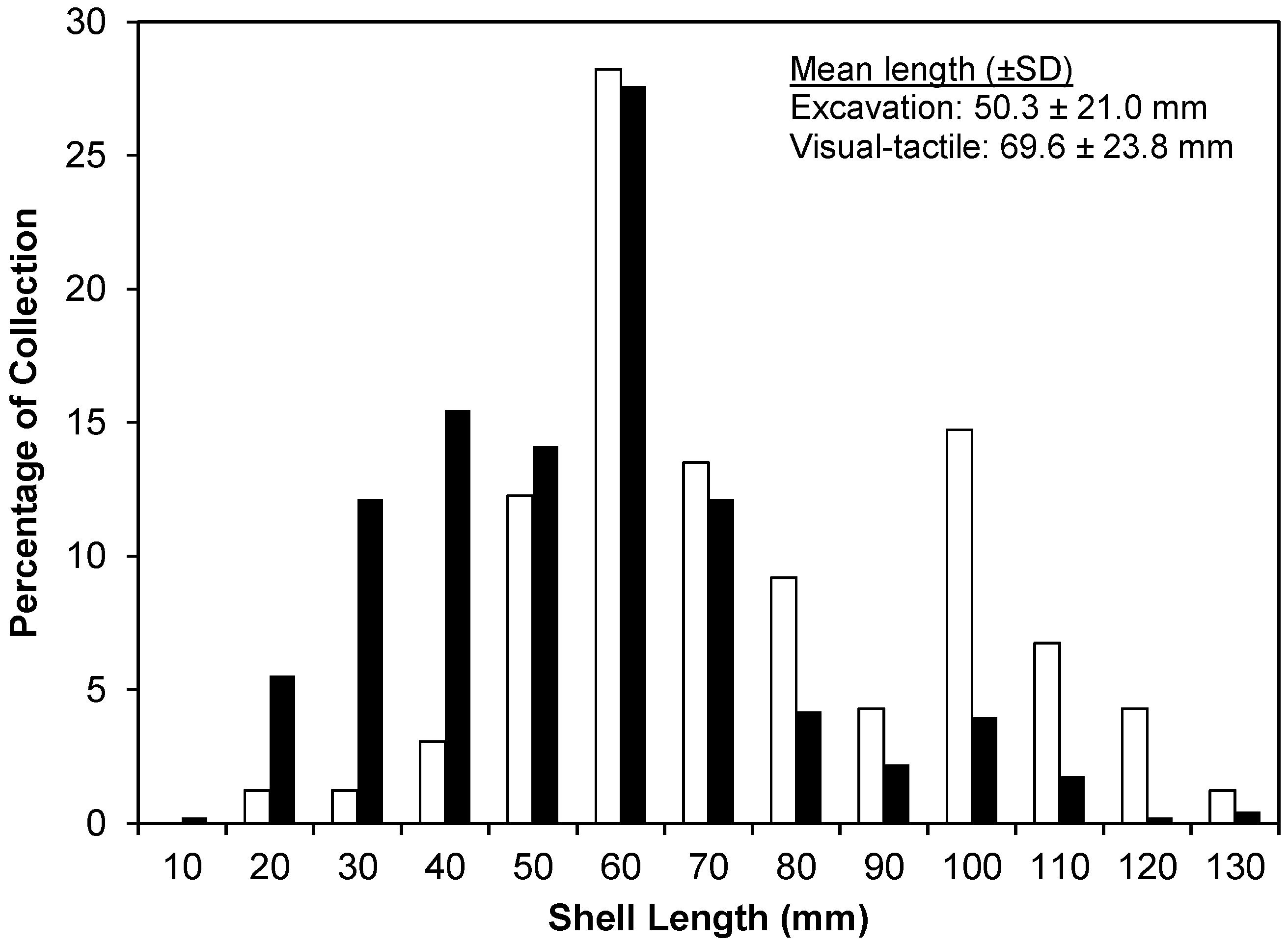
4.1. Multi-River Dataset
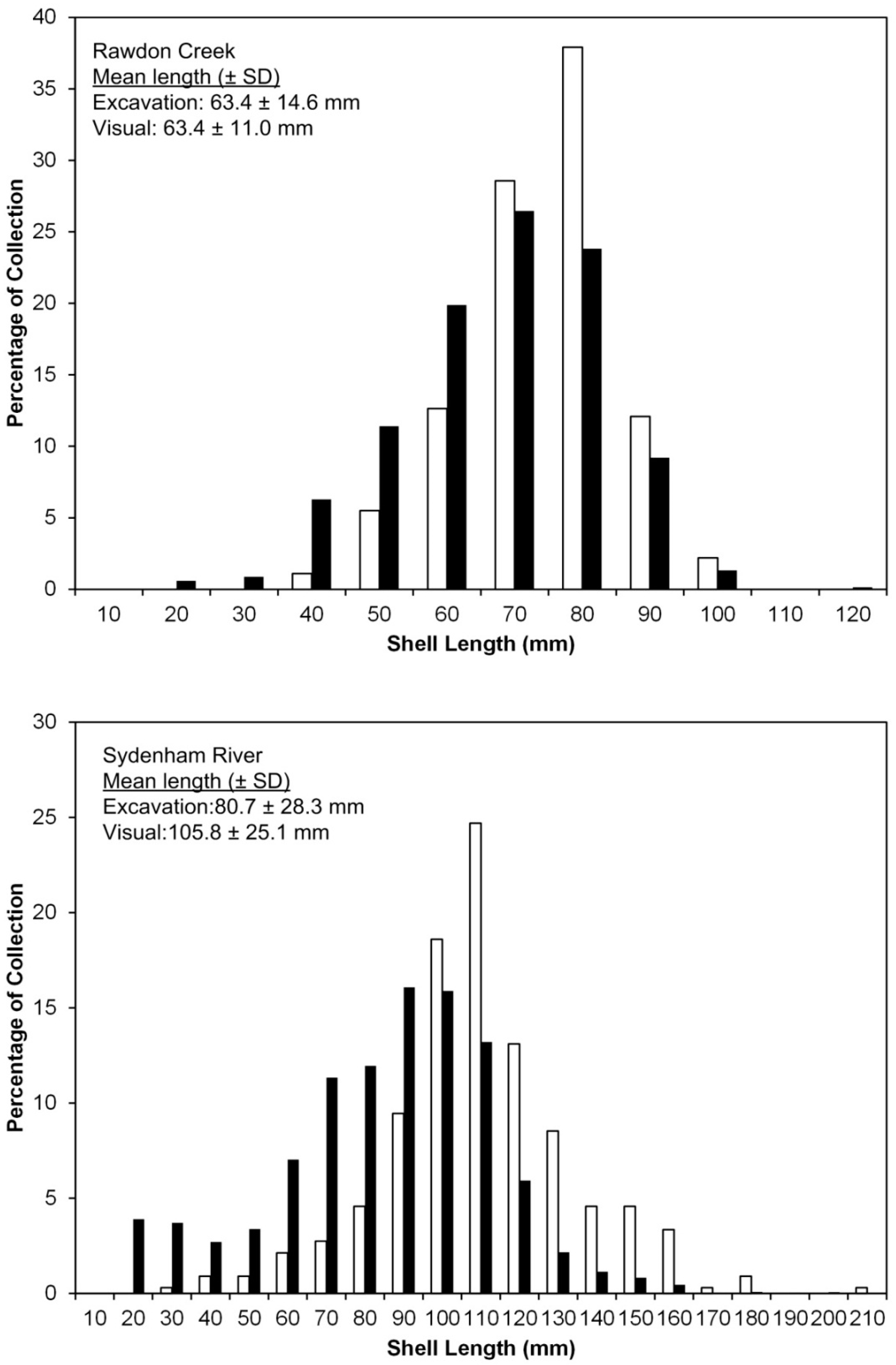
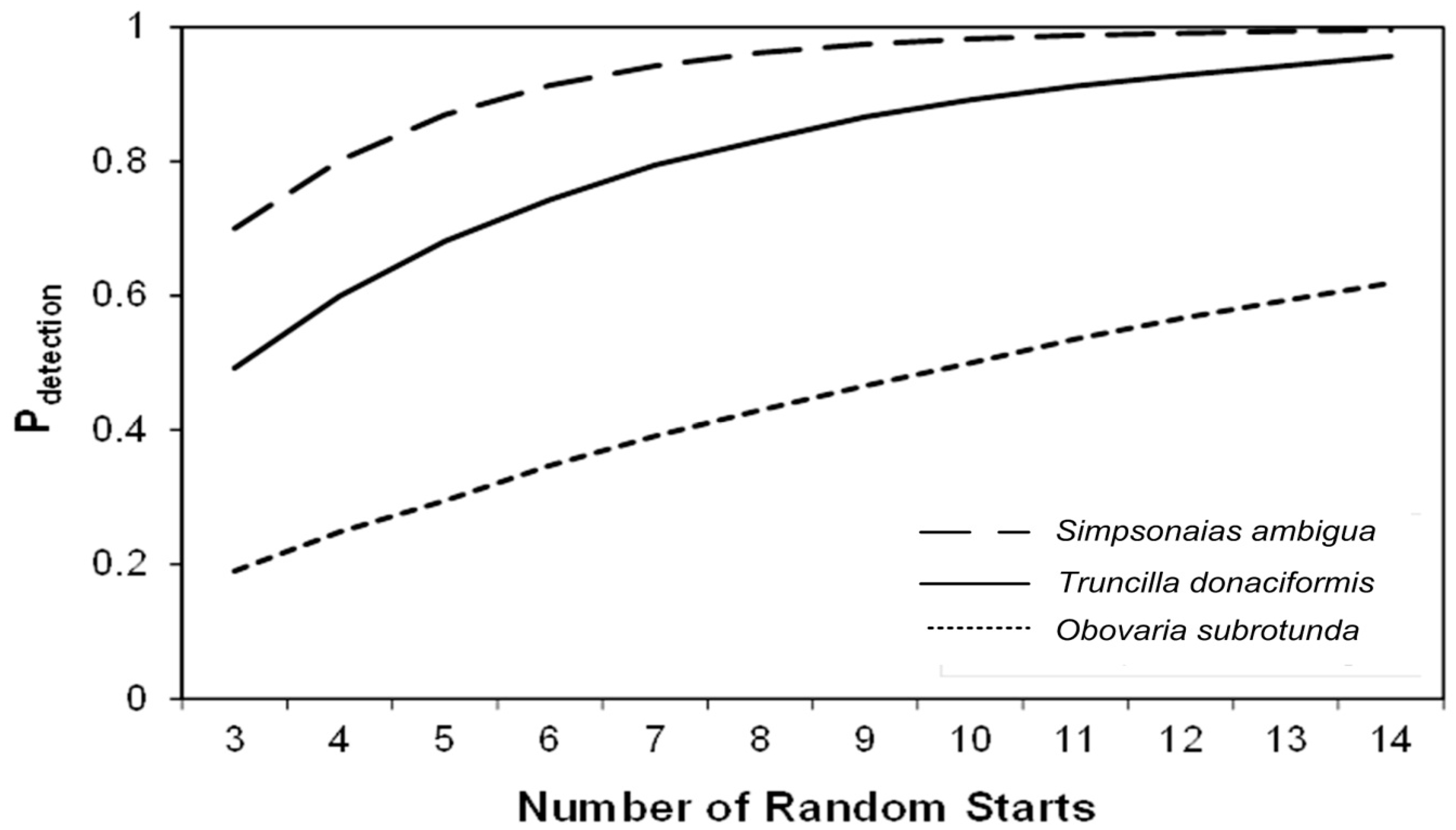
4.2. Census Dataset
4.2.1. Rawdon Creek
4.2.2. Sydenham River
5. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Blocks | Quadrats | N | SOBS | SEST | Q1 | Q2 | N/SOBS | Q1/SOBS 1 |
|---|---|---|---|---|---|---|---|---|---|
| Ausable River | |||||||||
| AR-12 | 25 | 75 | 298 | 16 | 19.9 | 4 | 2 | 18.6 | 25 |
| AR-13 | 25 | 75 | 82 | 9 | 1 | 0 | 9.1 | 11 | |
| AR-24 | 25 | 75 | 1356 | 18 | 18.5 | 1 | 1 | 75.3 | 6 |
| AR-27 | 31 | 93 | 122 | 12 | 16.5 | 3 | 1 | 10.2 | 25 |
| AR-28 | 27 | 81 | 379 | 16 | 16.7 | 2 | 3 | 23.7 | 13 |
| AR-29 | 24 | 72 | 9 | 5 | 9.4 | 3 | 1 | 1.8 | 60 |
| AR-33 | 29 | 87 | 24 | 7 | 11.0 | 4 | 2 | 3.4 | 57 |
| AR-34 | 22 | 66 | 133 | 11 | 11.2 | 1 | 2 | 12.1 | 9 |
| AR-5 | 23 | 69 | 114 | 13 | 15.2 | 3 | 2 | 8.8 | 23 |
| AR-7 | 23 | 69 | 870 | 19 | 4 | 0 | 45.8 | 21 | |
| AR-8 | 25 | 75 | 252 | 13 | 13.5 | 1 | 1 | 19.4 | 8 |
| Grand River | |||||||||
| GR-03 | 21 | 63 | 89 | 5 | 5.5 | 1 | 1 | 17.8 | 20 |
| GR-13 | 21 | 63 | 18 | 4 | 6.0 | 2 | 1 | 4.5 | 50 |
| GR-31 | 16 | 48 | 20 | 5 | 7.0 | 2 | 1 | 4.0 | 40 |
| GR-33 | 20 | 60 | 54 | 5 | 1 | 0 | 10.8 | 20 | |
| Maitland River | |||||||||
| MR-01 | 20 | 60 | 15 | 5 | 6.0 | 2 | 2 | 3.0 | 40 |
| MR-02 | 20 | 60 | 165 | 7 | 14.9 | 4 | 1 | 23.6 | 57 |
| MR-09 | 21 | 63 | 126 | 7 | 7.0 | 0 | 1 | 18.0 | 0 |
| MR-14 | 20 | 60 | 64 | 8 | 8.2 | 1 | 2 | 8.0 | 13 |
| MR-16 | 21 | 63 | 55 | 8 | 10.2 | 3 | 2 | 6.9 | 38 |
| MR-21 | 20 | 60 | 26 | 6 | 6.2 | 1 | 3 | 4.3 | 17 |
| Sydenham River | |||||||||
| SR1 | 28 | 84 | 85 | 16 | 8 | 0 | 5.3 | 50 | |
| SR10 | 25 | 75 | 246 | 17 | 21.4 | 3 | 1 | 14.5 | 18 |
| SR12 | 26 | 78 | 233 | 20 | 20.0 | 0 | 3 | 11.7 | 0 |
| SR13 | 25 | 75 | 151 | 9 | 4 | 0 | 16.8 | 44 | |
| SR15 | 26 | 78 | 165 | 6 | 6.0 | 0 | 1 | 27.5 | 0 |
| SR17 | 27 | 81 | 318 | 19 | 19.0 | 0 | 3 | 16.7 | 0 |
| SR19 | 25 | 75 | 816 | 23 | 23.2 | 1 | 2 | 35.5 | 4 |
| SR2 | 26 | 78 | 276 | 15 | 27.3 | 5 | 1 | 18.4 | 33 |
| SR20 | 27 | 81 | 42 | 9 | 4 | 0 | 4.7 | 44 | |
| SR21 | 28 | 84 | 10 | 2 | 1 | 0 | 5.0 | 50 | |
| SR3 | 23 | 69 | 230 | 21 | 7 | 0 | 11.0 | 33 | |
| SR5 | 23 | 69 | 769 | 21 | 21.2 | 1 | 2 | 36.6 | 5 |
| SR6 | 26 | 78 | 761 | 23 | 24.5 | 3 | 3 | 33.1 | 13 |
| SR7 | 27 | 81 | 1138 | 23 | 29.2 | 5 | 2 | 49.5 | 22 |
| Thames River | |||||||||
| TM04-15 | 23 | 69 | 75 | 9 | 9.0 | 0 | 0 | 8.3 | 0 |
| TM-05-01 | 23 | 69 | 96 | 14 | 16.0 | 2 | 1 | 6.9 | 14 |
| TR 97-11 | 22 | 66 | 16 | 7 | 9.2 | 3 | 2 | 2.3 | 43 |
| TR 97-3 | 22 | 66 | 146 | 7 | 7.2 | 1 | 2 | 20.9 | 14 |
| TR 97-18 | 21 | 63 | 87 | 7 | 3 | 0 | 12.4 | 43 | |
References
- Williams, J.D.; Warren, M.L.; Cummings, K.S.; Harris, J.L.; Neves, R.J. Conservation status of the freshwater mussels of the United States and Canada. Fisheries 1992, 18, 6–22. [Google Scholar] [CrossRef]
- Metcalfe-Smith, J.L.; MacKenzie, A.; Carmichael, I.; McGoldrick, D. Photo Field Guide to the Freshwater Mussels of Ontario; St. Thomas Field Naturalist Club Inc.: St. Thomas, ON, Canada, 2005. [Google Scholar]
- Committee on the Status of Endangered Wildlife in Canada (COSEWIC). Wildlife Species Search 2013. Available online: http://www.cosewic.gc.ca/eng/sct1/searchform_e.cfm (accessed on 17 September 2013).
- Ontario Ministry of Natural Resources and Forestry (MNRF). Species at Risk in Ontario (SARO) List. 2014. Available online: http://www.ontario.ca/environment-and-energy/species-risk-ontario-list (accessed on 16 September 2014). [Google Scholar]
- Schloesser, D.W.; Nalepa, T.F. Dramatic decline of unionid bivalves in the offshore waters of western Lake Erie after infestation by the zebra mussel, Dreissena polymorpha. Can. J. Fish. Aquat. Sci. 1994, 51, 2234–2242. [Google Scholar] [CrossRef]
- Clarke, A.H. Ontario’s Sydenham River, an important refugium for native freshwater mussels against competition from the zebra mussel, Dreissena polymorpha. Malacol. Data Net 1992, 3, 43–55. [Google Scholar]
- McNichols-O’Rourke, K.A.; Robinson, A.; Morris, T.J. Summary of Freshwater Mussel Timed Search Surveys in Southwestern Ontario in 2010 and 2011; Canadian Manuscript Report Fisheries and Aquatic Sciences 3009; Fisheries and Oceans Canada: Ottawa, ON, Canada, 2012.
- Morris, T.; Burridge, J.M. Recovery Strategy for Northern Riffleshell, Snuffbox, Round Pigtoe, Mudpuppy Mussel and Rayed Bean in Canada; Species at Risk Act Recovery Strategy Series; Fisheries and Oceans Canada: Ottawa, ON, Canada, 2006.
- Department of Fisheries and Oceans Canada (DFO). Report on the Progress of Recovery Strategy Implementation for the Wavyrayed Lampmussel, Northern Riffleshell, Snuffbox, Round Pigtoe, Mudpuppy Mussel and Rayed Bean in Canada for the Period 2006–2011; Species at Risk Act Recovery Strategy Report Series; Fisheries and Oceans Canada: Ottawa, ON, Canada, 2013.
- Cudmore, B.; Mandrak, N.E.; Morris, T.J.; Edwards, A. Allowable Harm Analysis Workshops for Freshwater Species at Risk in Central and Arctic Region; Canadian Science Advisory Secretariat Proceedings Series 2006/026; Fisheries and Oceans Canada: Ottawa, ON, Canada, 2006.
- Department of Fisheries and Oceans Canada (DFO). Assessment of Methods for the Identification of Critical Habitat for Freshwater Mussels; Canadian Science Advisory Secretariat Science Advisory Report 2011/047; Fisheries and Oceans Canada: Ottawa, ON, Canada, 2011.
- Thompson, S.K. Sampling, 2nd ed.; John Wiley and Sons: New York, NY, USA, 2002. [Google Scholar]
- Christman, M.C. A review of quadrat-based sampling of rare, geographically clustered populations. J. Agric. Biol. Ecol. Stat. 2000, 5, 168–201. [Google Scholar] [CrossRef]
- Smith, D.R.; Villella, R.F.; Lemarie, D.P. Survey protocol for assessment of endangered freshwater mussels in the Allegheny River, Pennsylvania. J. N. Am. Benthol. Soc. 2001, 20, 118–132. [Google Scholar] [CrossRef]
- Pooler, P.S.; Smith, D.R. Optimal sampling design for estimating spatial distribution and abundance of a freshwater mussel population. J. N. Am. Benthol. Soc. 2005, 24, 525–537. [Google Scholar] [CrossRef]
- Metcalfe-Smith, J.L.; Staton, S.K.; Mackie, G.L.; Lane, N.M. Changes in the biodiversity of freshwater mussels in the Canadian waters of the Lower Great Lakes drainage basin over the past 140 years. J. Gt. Lakes Res. 1998, 24, 845–858. [Google Scholar] [CrossRef]
- Metcalfe-Smith, J.L.; McGoldrick, D.J.; Zanatta, D.T.; Grapentine, L.C. Development of a Monitoring Program for Tracking the Recovery of Endangered Mussels in the Sydenham River, Ontario; Water Science and Technology Directorate Contribution 07-510; Environment Canada: Burlington, ON, Canada, 2007.
- Reid, S.M. Search effort and imperfect detection: Influence on timed-search mussel (Bivalvia: Unionidae) surveys in Canadian rivers. Knowl. Manag. Aquat. Ecosyst. 2016, 417, 17. [Google Scholar] [CrossRef]
- Green, R.H.; Young, R.C. Sampling to detect rare species. Ecol. Appl. 1993, 3, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Zar, J.H. Biostatistical Analysis, 2nd ed.; Prentice-Hall: Engelwood Cliffs, CA, USA, 1984. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Chao, A.; Colwell, R.K.; Lin, C.W.; Gotelli, N.J. Sufficient sampling for asymptotic minimum species richness estimators. Ecology 2009, 90, 1125–1133. [Google Scholar] [CrossRef]
- Lopez, L.C.S.; Fracasso, M.P.D.; Mesquita, D.O.; Palma, A.R.T.; Riul, P. The relationship between percentage of singletons and sampling effort: A new approach to reduce the bias of richness estimates. Ecol. Indic. 2012, 14, 164–169. [Google Scholar] [CrossRef]
- Coddington, J.A.; Agnarsson, I.; Miller, J.A.; Kunter, M.; Hormiga, G. Undersampling bias: The null hypothesis for singleton species in tropical arthropod surveys. J. Anim. Ecol. 2009, 78, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Strayer, D.L.; Smith, D.R. A Guide to Sampling Freshwater Mussel Populations; Monograph 8; American Fisheries Society: Bethesda, MD, USA, 2003. [Google Scholar]
- Jones, N.E.; Petreman, I.C. The capacity to detect change stream fish communities characteristics at the site-level in the Lake Ontario basin. Environ. Manag. 2012, 50, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Committee on the Status of Endangered Wildlife in Canada (COSEWIC). COSEWIC Assessment Process, Categories and Guidelines. Version: November 2015. Available online: http://www.cosewic.gc.ca/eng/sct1/searchform_e.cfm (accessed on 17 October 2016).
- Peterman, R. Statistical power analysis can improve fisheries research and management. Can. J. Fish. Aquat. Sci. 1990, 47, 2–15. [Google Scholar] [CrossRef]
- Jackson, D.A.; Harvey, H.H. Qualitative and quantitative sampling of lake fish communities. Can. J. Fish. Aquat. Sci. 1997, 54, 2807–2813. [Google Scholar] [CrossRef]
- Manly, B.F.J. Randomization, Bootstrap and Monte Carlo Methods in Biology, 3rd ed.; Chapman and Hall/CRC: Boca Raton, FL, USA, 2007. [Google Scholar]
- Legendre, P.; Legendre, L. Numerical Ecology, 2nd ed.; Elsevier Science: Amsterdam, The Netherlands, 1998. [Google Scholar]
- Hood, G.M. PopTools Version 3.2.5. 2010. Available online: http://www.poptools.org (accessed on 19 March 2014).
- Morrison, L.W.; Smith, D.R.; Young, C.C.; Nichols, D.W. Evaluating sampling designs by computer simulation: A case study with the Missouri bladderpod. Popul. Ecol. 2008, 50, 417–425. [Google Scholar] [CrossRef]
- Krebs, C.J. Ecological Methodology; Harper and Row Publishers: New York, NY, USA, 1989. [Google Scholar]
- Rosenberg, M.S.; Anderson, C.D. PASSaGE: Pattern analysis, spatial statistics and geographic exegesis. Version 2. Methods Ecol. Evol. 2011, 2, 229–232. [Google Scholar] [CrossRef]
- Smith, D.R.; Villella, R.F.; Lemarie, D.P. Application of adaptive cluster sampling to low-density populations of freshwater mussels. Environ. Ecol. Stat. 2003, 10, 7–15. [Google Scholar] [CrossRef]
- Villella, R.F.; Smith, D.R. Two-phase sampling to estimate river-wide populations of freshwater mussels. J. N. Am. Benthol. Soc. 2005, 24, 357–368. [Google Scholar] [CrossRef]
- Thompson, W.L. Future directions in estimating abundance of rare or elusive species. In Sampling Rare or Elusive Species. Concepts, Designs, and Techniques for Estimating Population Parameters; Thompson, W.L., Ed.; Island Press: Washington, DC, USA, 2004; pp. 389–399. [Google Scholar]
- Witmer, G.W. Wildlife population monitoring: Some practical considerations. Wildl. Res. 2005, 32, 259–263. [Google Scholar] [CrossRef]
- Smith, D.R.; Villella, R.F.; Lemarie, D.P.; von Oettingen, S. How much excavation is needed to monitor freshwater mussels? In Proceedings of the First Mollusk Conservation Society Symposium, Chattanooga, TN, USA, 16–20 March 1999; Ohio Biological Society: Columbus, OH, USA, 2000; pp. 203–218. [Google Scholar]
- Wisniewski, J.M.; Rankin, N.M.; Weiler, D.A.; Strickland, B.A.; Chandler, H.C. Occupancy and detection of benthic invertebrates: A case study of unionids in the lower Flint River, Georgia, USA. Freshw. Sci. 2013, 32, 1122–1135. [Google Scholar] [CrossRef]
- Metcalfe-Smith, J.L.; Maio, J.; Staton, S.; Mackie, G. Effect of sampling effort on the efficiency of the timed search method for sampling freshwater mussel communities. J. N. Am. Benthol. Soc. 2000, 19, 725–732. [Google Scholar] [CrossRef]
- Epp, J.M.; Morris, T.J.; McNichols-O’Rourke, K.A. A Preliminary Search for Epioblasma torulosa rangiana (Northern Riffleshell) in the Maitland River Watershed; Canadian Manuscript Report Fisheries and Aquatic Sciences 3025; Fisheries and Oceans Canada: Ottawa, ON, Canada, 2013.
- Randklev, C.R.; Johnson, M.S.; Tsakiris, E.T.; Groce, J.; Wilkins, N. Status of the freshwater mussel (Unionidae) communities of the Leon River, Texas. Aquat. Conserv. Mar. Freshw. Ecosyst. 2013, 23, 390–404. [Google Scholar] [CrossRef]
- Smith, T.A.; Crabtree, D. Freshwater mussel (Unionidae: Bivalvia) distributions and densities in French Creek, Pennsylvania. Northeast. Nat. 2010, 17, 387–414. [Google Scholar] [CrossRef]
- Guillera-Arroita, G.; Lahoz-Monfort, J.J. Designing studies to detect differences in species occupancy: Power analysis under imperfect detection. Methods Ecol. Evol. 2012, 3, 860–869. [Google Scholar] [CrossRef]
- Salehi, M.M.; Smith, D.R. Two-stage sequential sampling: A neighbourhood-free adaptive sampling procedure. J. Agric. Biol. Ecol. Stat. 2005, 10, 84–103. [Google Scholar] [CrossRef]
- Committee on the Status of Endangered Wildlife in Canada (COSEWIC). COSEWIC Assessment and Status Report on the Wavy-Rayed Lampmussel Lampsilis fasciola in Canada; Committee on the Status of Endangered Wildlife in Canada: Ottawa, ON, Canada, 2010. [Google Scholar]









| Tribe | Species | Rivers 1 | Sites | Density | CV | p Detection |
|---|---|---|---|---|---|---|
| Alasmidontini | Alasmidonta marginata | A,G,M,S,T | 30 | 0.09 (0.21) | 0.27 (0.17) | 0.62 (0.30) |
| Alasmidonta viridis | G,M,S | 5 | 0.06 (0.06) | 0.48 (0.09) | 0.45 (0.27) | |
| Anodontoides ferussacianus | M,S | 1 | 0.07 | 0.18 | ||
| Lasmigona complanata | A,S,T | 15 | 0.24 (0.38) | 0.26 (0.17) | 0.64 (0.33) | |
| Lasmigona compressa | A,G,M | 9 | 0.03 (0.01) | 0.49 (0.08) | 0.21 (0.10) | |
| Lasmigona costata | A,G,M,S,T | 35 | 0.58 (0.78) | 0.16 (0.15) | 0.82 (0.27) | |
| Pyganodan grandis | A,G,M,S | 14 | 0.08 (0.14) | 0.36 (0.17) | 0.47 (0.29) | |
| Simpsonaias ambigua * | S | 7 | 0.03 (0.01) | 0.43 (0.16) | 0.31 (0.15) | |
| Strophitus undulatus | A,G,M,S,T | 18 | 0.09 (0.08) | 0.27 (0.15) | 0.52 (0.28) | |
| Utterbackia imbecillis | S | 1 | 0.01 | 0.32 | ||
| Amblemini | Amblema plicata | A,S,T | 24 | 1.11 (2.28) | 0.14 (0.14) | 0.84 (0.27) |
| Lampsilini | Actinonaias ligamentina | M,S,T | 16 | 0.53 (0.61) | 0.19 (0.16) | 0.79 (0.31) |
| Epioblasma torulosa rangiana * | A,S | 10 | 0.07 (0.08) | 0.31 (0.16) | 0.50 (0.27) | |
| Epioblasma triquetra * | A,S | 13 | 0.07 (0.14) | 0.38 (0.15) | 0.43 (0.27) | |
| Lampsilis cardium | A,M,S,T | 24 | 0.16 (0.16) | 0.25 (0.18) | 0.69 (0.28) | |
| Lampsilis fasciola * | A,G,T | 26 | 0.15 (0.18) | 0.24 (0.16) | 0.62 (0.26) | |
| Lampsilis siliquoidea | A,G,S | 17 | 0.10 (0.09) | 0.24 (0.19) | 0.67 (0.31) | |
| Leptodea fragilis | A,S,T | 17 | 0.17 (0.14) | 0.22 (0.16) | 0.69 (0.32) | |
| Ligumia recta | A,S,T | 21 | 0.12 (0.11) | 0.22 (0.15) | 0.63 (0.31) | |
| Obliquaria reflexa | S,T | 2 | 0.03 | 0.41 | 0.32 | |
| Potamilus alatus | A,S,T | 18 | 0.05 (0.05) | 0.35 (0.16) | 0.42 (0.27) | |
| Ptychobranchus fascioloris * | A,M,S | 14 | 0.19 (0.32) | 0.26 (0.16) | 0.69 (0.28) | |
| Truncilla donaciformis * | S | 2 | 0.06 | 0.22 | 0.57 | |
| Truncilla truncata | A,S,T | 12 | 0.08 (0.11) | 0.34 (0.14) | 0.49 (0.25) | |
| Villosa fabalis * | S,T | 11 | 0.74 (1.09) | 0.22 (0.18) | 0.69 (0.35) | |
| Villosa iris * | A,M,S,T | 12 | 0.4 (0.72) | 0.19 (0.14) | 0.66 (0.30) | |
| Pleurobemini | Cyclonaias tuberculata | A,S,T | 19 | 0.91 (1.46) | 0.17 (0.15) | 0.82 (0.26) |
| Elliptio dilatata | A,G,S,T | 21 | 0.47 (0.60) | 0.17 (0.12) | 0.82 (0.26) | |
| Fusconaia flava | A,S,T | 22 | 0.33 (0.55) | 0.22 (0.15) | 0.78 (0.29) | |
| Pleurobema sintoxia * | S,T | 8 | 0.05 (0.06) | 0.39 (0.14) | 0.44 (0.29) | |
| Quadrulini | Quadrula pustulosa | A,S,T | 9 | 0.16 (0.32) | 0.27 (0.13) | 0.58 (0.25) |
| Quadrula quadrula * | A,S,T | 13 | 0.22 (0.19) | 0.22 (0.15) | 0.79 (0.28) | |
| All Mussels | 3.51 (4.28) | 0.06 (0.04) |
| Species | Rawdon Creek (Total = 866 Mussels) | Sydenham River (Total = 6180 Mussels) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| x | s2/x | FO (%) | Visual (%) | Excavation (%) | x | s2/x | FO (%) | Visual (%) | Excavation (%) | |
| Actinonaias ligamentina | 0.54 | 1.8 | 32.0 | 9.3 | 90.7 | |||||
| Alasmidonta marginata | 0.17 | 1.1 | 0.5 | 3.1 | 96.9 | |||||
| Amblema plicata | 1.78 | 2.1 | 70.7 | 8.8 | 91.2 | |||||
| Cyclonaias tuberculata | 6.98 | 5.0 | 96.0 | 5.3 | 94.7 | |||||
| Elliptio complanata | 1.97 | 2.5 | 68.3 | 6.8 | 93.2 | |||||
| Elliptio dilatata | 1.40 | 1.5 | 68.3 | 0 | 100 | |||||
| Epioblasma torulosa rangiana * | 0.16 | 1.1 | 14.1 | 1.6 | 98.4 | |||||
| Epioblasma triquetra * | 0.19 | 1.1 | 16.8 | 0 | 100 | |||||
| Fusconaia flava | 0.43 | 1.2 | 31.2 | 2.5 | 97.5 | |||||
| Lampsilis cardium | 0.06 | 1.0 | 5.6 | 23.8 | 76.2 | 0.02 | 1.0 | 1.6 | 16.7 | 83.3 |
| Lampsilis siliquoidea | 0.02 | 1.0 | 1.9 | 37.5 | 62.5 | |||||
| Lasmigona complanata | 0.12 | 1.1 | 10.7 | 15.9 | 84.1 | |||||
| Lasmigona costata | 0.01 | 1.0 | 1.1 | 40 | 60 | 0.99 | 2.8 | 68.5 | 5.9 | 94.1 |
| Leptodea fragilis | 0.18 | 1.1 | 16.3 | 2.9 | 97.1 | |||||
| Ligumia recta | 0.003 | 1.0 | 0.3 | 0 | 100 | 0.24 | 1.2 | 19.5 | 15.7 | 84.3 |
| Obovaria subrotunda * | 0.003 | 1.0 | 0.3 | 0 | 100 | |||||
| Pleurobema sintoxia * | 0.08 | 1.1 | 6.9 | 6.9 | 93.1 | |||||
| Potamilus alatus | 0.08 | 1.1 | 6.9 | 14.2 | 85.8 | |||||
| Ptychobranchus fuscioloris * | 0.46 | 1.1 | 36.0 | 1.2 | 98.8 | |||||
| Pyganodon grandis | 0.01 | 1.0 | 1.1 | 0 | 100 | |||||
| Quadrula pustulosa | 0.17 | 1.2 | 14.4 | 3.2 | 96.8 | |||||
| Quadrula quadrula * | 0.74 | 1.7 | 42.1 | 9.9 | 90.1 | |||||
| Simpsonaias ambigua * | 0.02 | 1.0 | 1.3 | 0 | 100 | |||||
| Strophitus undulatus | 0.01 | 1.0 | 0.8 | 66.7 | 33.3 | 0.04 | 1.0 | 4.0 | 0 | 100 |
| Truncilla donaciformis * | 0.03 | 4.3 | 1.1 | 0 | 100 | |||||
| Truncilla truncata | 0.01 | 1 | 0.5 | 0 | 100 | |||||
| Villosa fabalis * | 0.69 | 1.6 | 41.9 | 0 | 100 | |||||
| Villosa iris * | 0.24 | 1.3 | 19.5 | 7.8 | 92.2 | |||||
| All Mussels | 2.31 | 2.7 | 7.2 | 92.8 | 16.48 | 11.5 | 5.3 | 94.7 | ||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reid, S.M.; Morris, T.J. Tracking the Recovery of Freshwater Mussel Diversity in Ontario Rivers: Evaluation of a Quadrat-Based Monitoring Protocol. Diversity 2017, 9, 5. https://doi.org/10.3390/d9010005
Reid SM, Morris TJ. Tracking the Recovery of Freshwater Mussel Diversity in Ontario Rivers: Evaluation of a Quadrat-Based Monitoring Protocol. Diversity. 2017; 9(1):5. https://doi.org/10.3390/d9010005
Chicago/Turabian StyleReid, Scott M., and Todd J. Morris. 2017. "Tracking the Recovery of Freshwater Mussel Diversity in Ontario Rivers: Evaluation of a Quadrat-Based Monitoring Protocol" Diversity 9, no. 1: 5. https://doi.org/10.3390/d9010005
APA StyleReid, S. M., & Morris, T. J. (2017). Tracking the Recovery of Freshwater Mussel Diversity in Ontario Rivers: Evaluation of a Quadrat-Based Monitoring Protocol. Diversity, 9(1), 5. https://doi.org/10.3390/d9010005