
Selective Feeding and Microalgal Consumption Rates by Crown-Of-Thorns Seastar (Acanthaster cf. solaris) Larvae

Abstract
:1. Introduction
2. Material and Methods
2.1. Spawning and Maintenance of CoTS Larvae
2.2. Phytoplankton Cultures and Analysis
2.3. Larval Feeding Experiments
2.4. Modelling
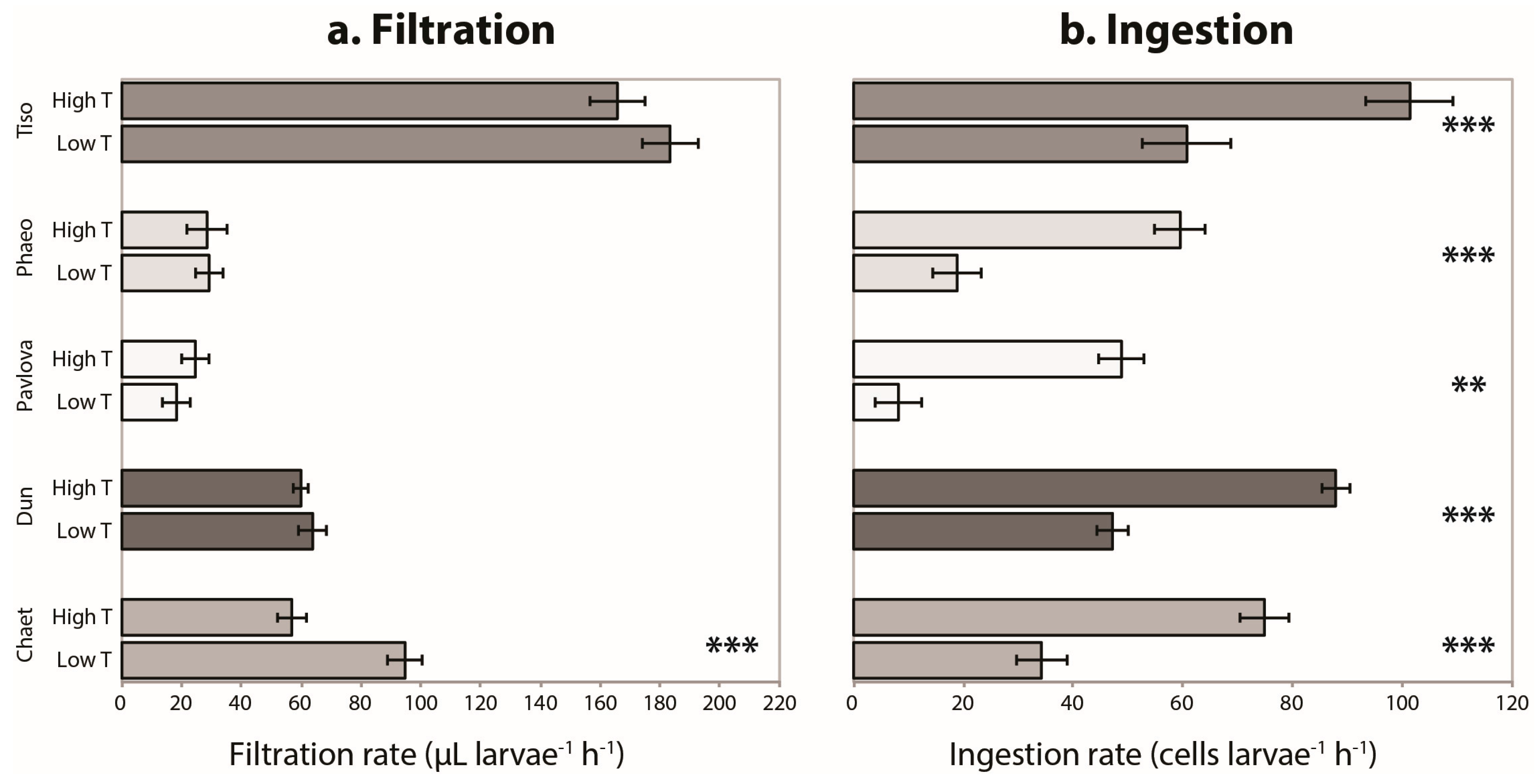
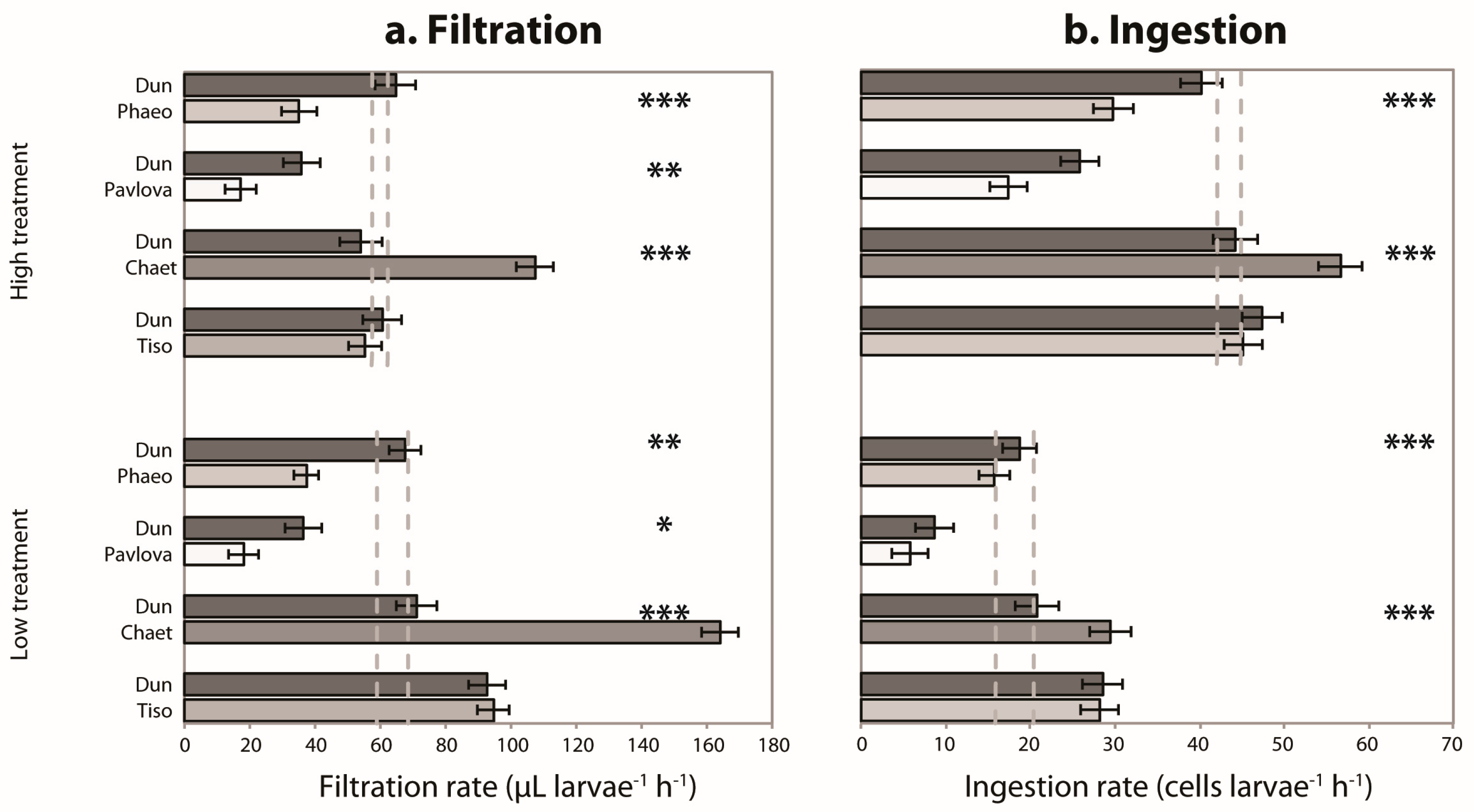
3. Results
3.1. Single Species Experiment
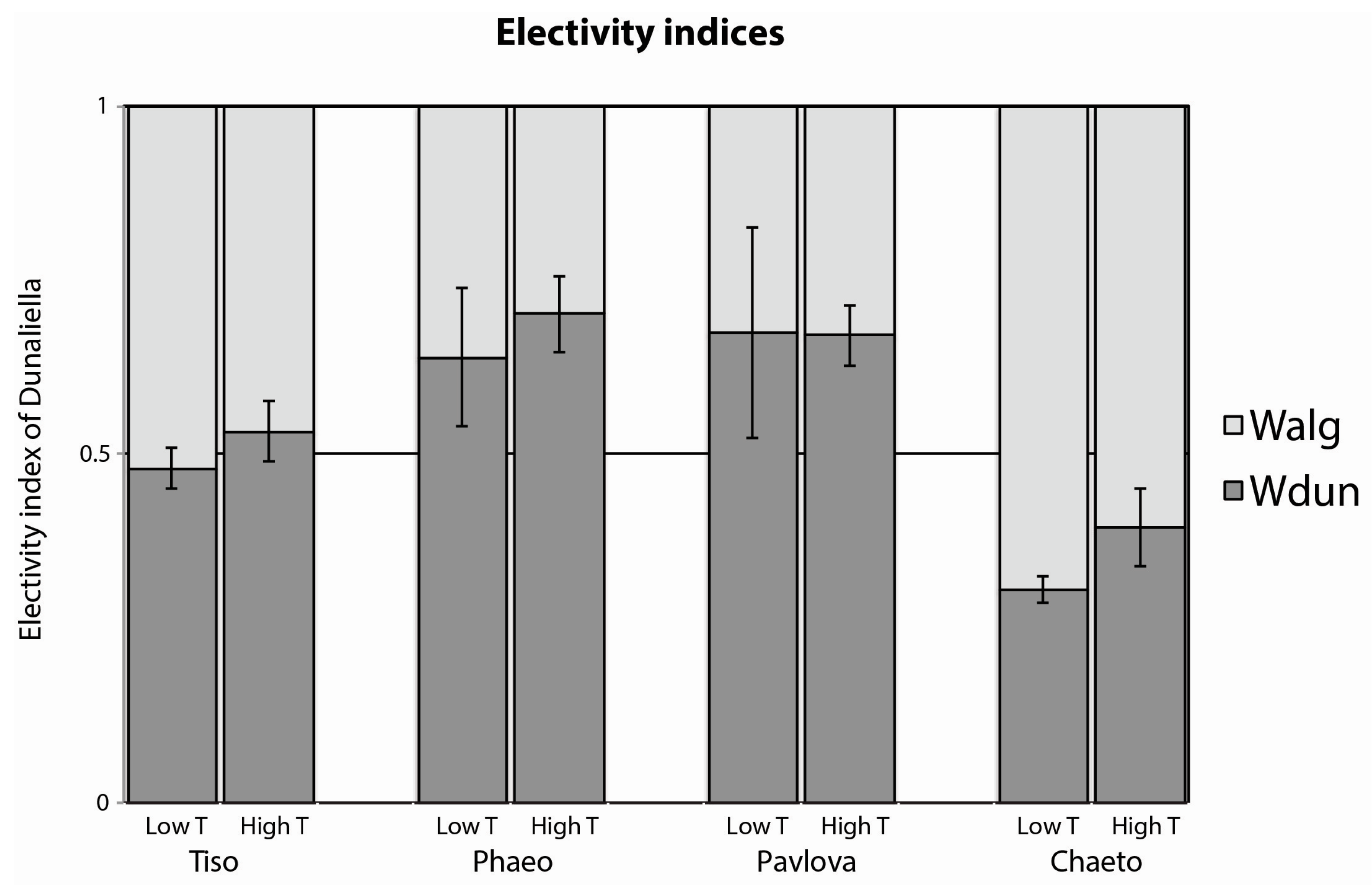
3.2. Mixed Species Experiments
3.3. Algae Quality
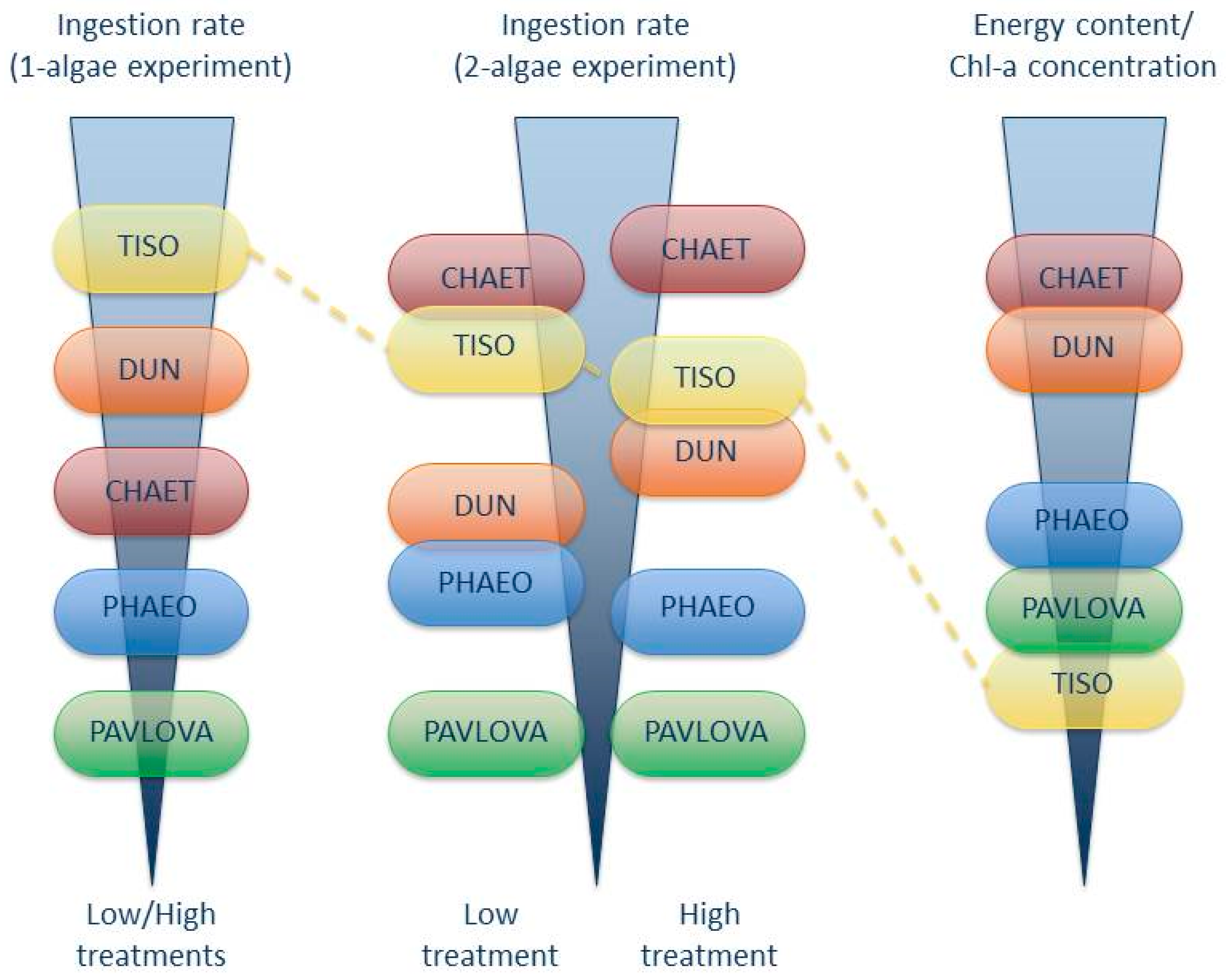
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment/Treatment | Expected Initial Concentration (cells·mL−1) | Actual Measured Concentration (cells·mL−1) | |
|---|---|---|---|
| Mean ± Standard Deviation | Minimum-Maximum | ||
| Single-species | |||
| Low concentration | 1000 | 848 ± 180 | 520–1210 |
| High concentration | 3000 | 2563 ± 500 | 1530–3780 |
| Mixed-species (total concentration) | |||
| Low concentration | 1000 | 947 ± 131 | 740–1310 |
| High concentration | 3000 | 848 ± 180 | 1860–3630 |
| Mixed-species (individual concentrations) | |||
| Low concentration | 500 | 461 ± 110 | 340–760 |
| High concentration | 1500 | 1315 ± 252 | 850–2070 |
References
- De’ath, G.; Fabricius, K.E.; Sweatman, H.; Puotinen, M. The 27-year decline of coral cover on the Great Barrier Reef and its causes. Proc. Natl. Acad. Sci. USA 2012, 109, 17995–17999. [Google Scholar] [CrossRef] [PubMed]
- Birkeland, C. Terrestrial runoff as a cause of outbreaks of Acanthaster planci (Echinodermata: Asteroidea). Mar. Biol. 1982, 69, 175–185. [Google Scholar] [CrossRef]
- Brodie, J. From the effects of terrestrial runoff: A review. Mar. Freshw. Res. 1992, 43, 539. [Google Scholar] [CrossRef]
- Brodie, J.; Fabricius, K.; De'ath, G.; Okaji, K. Are increased nutrient inputs responsible for more outbreaks of crown-of-thorns starfish? An appraisal of the evidence. Mar. Pollut. Bull. 2005, 51, 266–278. [Google Scholar] [CrossRef] [PubMed]
- Devlin, M.J.; Brodie, J. Terrestrial discharge into the Great Barrier Reef Lagoon: Nutrient behavior in coastal waters. Mar. Pollut. Bull. 2005, 51, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Fabricius, K.E.; Okaji, K.; De'ath, G. Three lines of evidence to link outbreaks of the crown-of-thorns seastar Acanthaster planci to the release of larval food limitation. Coral Reefs 2010, 29, 593–605. [Google Scholar] [CrossRef]
- Lucas, J.S. Quantitative studies of feeding and nutrition during larval development of coral reef asteroid Acanthaster planci (L.). J. Exp. Mar. Biol. Ecol. 1982, 65, 173–193. [Google Scholar] [CrossRef]
- Wolfe, K.; Graba-Landry, A.; Dworjanyn, S.A.; Byrne, M. Larval starvation to satiation: Influence of nutrient regime on the success of Acanthaster planci. PLoS ONE 2015, 10, e0122010. [Google Scholar] [CrossRef] [PubMed]
- Uthicke, S.; Logan, M.; Liddy, M.; Francis, D.; Hardy, N.; Lamare, M. Climate change as an unexpected co-factor promoting coral eating seastar (Acanthaster planci) outbreaks. Sci. Rep. 2015, 5, 8402. [Google Scholar] [CrossRef] [PubMed]
- Strathmann, R.R. The feeding behavior of planktotrophic echinoderm larvae: Mechanisms, regulation, and rates of suspensionfeeding. J. Exp. Mar. Bio. Ecol. 1971, 6, 109–160. [Google Scholar] [CrossRef]
- Strathmann, R.R. Larval Feeding in Echinoderms. Am. Zool. 1975, 15, 717–730. [Google Scholar] [CrossRef]
- Ayukai, T. Ingestion of ultraplankton by the planktonic larvae of the crown-of-thorns starfish, Acanthaster planci. Biol. Bull. 1994, 186, 90–100. [Google Scholar] [CrossRef]
- Okaji, K.; Ayukai, T.; Lucas, J.S. Selective feeding by larvae of the crown-of-thorns starfish, Acanthaster planci (L.). Coral Reefs 1997, 16, 47–50. [Google Scholar] [CrossRef]
- Nakajima, R.; Nakatomi, N.; Kurihara, H.; Fox, M.; Smith, J.; Okaji, K. Crown-of-thorns starfish larvae can feed on organic matter released from corals. Diversity 2016, 8, 18. [Google Scholar] [CrossRef]
- Collier, J.L. Flow cytometry and the single cell in phycology. J. Phycol. 2000, 36, 628–644. [Google Scholar] [CrossRef]
- Hofstraat, J.W.; van Zeijl, W.J.M.; de Vreeze, M.E.J.; Peeters, J.C.H.; Peperzak, L.; Colijn, F.; Rademaker, T.W.M. Phytoplankton monitoring by flow cytometry. J. Plankton Res. 1994, 16, 1197–1224. [Google Scholar] [CrossRef]
- Cowles, T.J.; Olson, R.J.; Chisholm, S.W. Food selection by copepods: Discrimination on the basis of food quality. Mar. Biol. 1988, 100, 41–49. [Google Scholar] [CrossRef]
- Baldwin, B.S. Selective particle ingestion by oyster larvae (Crassostrea virginica) feeding on natural seston and cultured algae. Mar. Biol. 1995, 123, 95–107. [Google Scholar] [CrossRef]
- Dionisio Pires, L.M.; Jonker, R.R.; Van Donk, E.; Laanbroek, H.J. Selective grazing by adults and larvae of the zebra mussel (Dreissena polymorpha): Application of flow cytometry to natural seston. Freshw. Biol. 2004, 49, 116–126. [Google Scholar] [CrossRef]
- Fileman, E.S.; Lindeque, P.K.; Harmer, R.A.; Halsband, C.; Atkinson, A. Feeding rates and prey selectivity of planktonic decapod larvae in the Western English Channel. Mar. Biol. 2014, 161, 2479–2494. [Google Scholar] [CrossRef]
- Revelante, N.; Gilmartin, M. Dynamics of phytoplankton in the Great Barrier Reef lagoon. J. Plankton Res. 1982, 4, 47–76. [Google Scholar] [CrossRef]
- Devlin, M.J.; Debose, J.; Ajani, P.; Petus, C.; da Silva, E.T.; Brodie, J.O.N. Phytoplankton in the Great Barrier Reef: Microscopy analysis of community structure in high flow events. Report to the National Environmental Research Program. Reef and Rainforest Research Centre Limited: Cairns, Australia, 2013; p. 68. [Google Scholar]
- Conlan, J.A.; Jones, P.L.; Turchini, G.M.; Hall, M.R.; Francis, D. Changes in the nutritional composition of captive early-mid stage Panulirus ornatus phyllosoma over ecdysis and larval development. Aquaculture 2014, 434, 159–170. [Google Scholar] [CrossRef]
- Angell, A.R.; Mata, L.; de Nys, R.; Paul, N.A. The protein content of seaweeds: A universal nitrogen-to-protein conversion. J. Appl. Phycol. 2016, 28, 511–524. [Google Scholar] [CrossRef]
- Bureau, D.P.; Kaushik, S.J.; Cho, C.Y. Bioenergetics. In Fish Nutrition; Halver, J.E., Hardy, R.W., Eds.; Academic Press: San Diego, CA, USA, 2002. [Google Scholar]
- Schaffelke, B.; Carleton, J.; Skuza, M.; Zagorskis, I.; Furnas, M.J. Water quality in the inshore Great Barrier Reef lagoon: Implications for long-term monitoring and management. Mar. Pollut. Bull. 2012, 65, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Okaji, K. Feeding ecology in the early life stages of the crown-of-thorns starfish, Acanthaster planci (L.). PhD dissertation, James Cook University, Townsville, Australia, 1996. [Google Scholar]
- Coughlan, J. The estimation of filtering rate from the clearance of suspensions. Mar. Biol. 1969, 2, 356–358. [Google Scholar] [CrossRef]
- Vanderploeg, H.a.; Scavia, D. Calculation and use of selectivity coefficients of feeding: Zooplankton grazing. Ecol. Modell. 1979, 7, 135–149. [Google Scholar] [CrossRef]
- Bolker, B.M.; Brooks, M.E.; Clark, C.J.; Geange, S.W.; Poulsen, J.R.; Stevens, M.H.H.; White, J.S.S. Generalized linear mixed models: A practical guide for ecology and evolution. Trends Ecol. Evol. 2009, 24, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Marin, V.; Huntley, M.E.; Frost, B. Measuring feeding rates of pelagic herbivores: Analysis of experimental design and methods. Mar. Biol. 1986, 58, 49–58. [Google Scholar] [CrossRef]
- Nakagawa, S.; Schielzeth, H. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Meth. Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model selection and multimodel inference: A practical information theoretic approach, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Burnham, K.P.; Anderson, D.R.; Huyvaert, K.P. AIC model selection and multimodel inference in behavioral ecology: Some background, observations, and comparisons. Behav. Ecol. Sociobiol. 2011, 65, 23–35. [Google Scholar] [CrossRef]
- R Development Core Team. R: A language and environment for statistical computing; R Foundation for Statistical Computing: Vienna, Austria; ISBN 3–900051–07–0. 2014; Available online: http://www.R-project.org/. (accessed on 21 January 2015).
- Pratchett, M.S.; Caballes, C.F.; Rivera-Posada, J.A.; Sweatman, H.P. Limits to understanding and managing outbreaks of crown-of-thorns starfish (Acanthaster spp.). Oceanogr. Mar. Biol. 2014, 52, 133–199. [Google Scholar]
- Mellin, C.; Lurgi, M.; Matthews, S.; MacNeil, M.A.; Caley, M.J.; Bax, N.; Przeslawski, R.; Fordham, D.A. Forecasting marine invasions under climate change: Biotic interactions and demographic processes matter. Biol. Conserv. 2016. [Google Scholar] [CrossRef]
- Pratchett, M.S.; Dworjanyn, S.A.; Mos, B.; Caballes, C.F.; Thompson, C.A.; Blowes, S. Larval survivorship and settlement of crown-of-thorns starfish (Acanthaster cf. solaris) at varying algal cell densities. Diversity 2017, 9, 2. [Google Scholar] [CrossRef]
- Hay, M.E. Marine chemical ecology: Chemical signals and cues structure marine populations, communities, and ecosystems. Ann. Rev. Mar. Sci. 2009, 1, 193–212. [Google Scholar] [CrossRef] [PubMed]
- Labarbera, M. Feeding currents and particle capture mechanisms in suspension feeding animals. Integr. Comp. Biol. 1984, 24, 71–84. [Google Scholar] [CrossRef]
- Rubenstein, D.I.; Koehl, M.A.R. The mechanisms of filter feeding: Some theoretical considerations. Am. Nat. 1977, 111, 981. [Google Scholar] [CrossRef]
- Rassoulzadegan, F.; Fenaux, L.; Strathmann, R.R. Effect of flavor and size on selection of food by suspension-feeding plutei. Limnol. Oceanog. 1984, 29, 357–360. [Google Scholar] [CrossRef]
- Strathmann, R.R. Time and extent of ciliary response to particles in a non-filtering feeding mechanism. Biol. Bull. 2007, 212, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Frost, B.W. Effects of size and concentration of food particles on the feeding behavior of the marine planktonic copepod Calanus pacificus. Limnol. Oceanogr. 1972, 17, 805–815. [Google Scholar] [CrossRef]
- Nell, J.A.; Connor, W.A.O. The evaluation of fresh algae and stored algal concentrates as a food source for Sydney rock oyster, Succostrea commercialis (Iredale & Roughley), larvae. Aquaculture 1991, 99, 277–284. [Google Scholar]
- Delaunay, F. The effect of monospecific algal diets on growth and fatty acid composition of Pecten maximus (L.) larvae. J. Exp. Mar. Bio. Ecol. 1993, 173, 163–179. [Google Scholar] [CrossRef]
- Ponis, E.; Robert, R.; Parisi, G.; Tredici, M. Assessment of the performance of Pacific oyster (Crassostrea gigas) larvae fed with fresh and preserved Pavlova lutheri concentrates. Aquaculture Inter. 2003, 11, 69–79. [Google Scholar] [CrossRef]
- Byrne, M.; Sewell, M.A.; Prowse, T.A.A. Nutritional ecology of sea urchin larvae: Influence of endogenous and exogenous nutrition on echinopluteal growth and phenotypic plasticity in Tripneustes gratilla. Funct. Ecol. 2008, 22, 643–648. [Google Scholar] [CrossRef]
- Duy, N.D.Q.; Francis, D.S.; Pirozzi, I.; Southgate, P.C. Use of micro-algae concentrates for hatchery culture of sandfish, Holothuria scabra. Aquaculture 2016, 464, 145–152. [Google Scholar] [CrossRef]
- Suzuki, G.; Yasuda, N.; Ikehara, K.; Fukuoka, K.; Kameda, T.; Kai, S.; Nagai, S.; Watanabe, A.; Nakamura, T.; Kitazawa, S.; et al. Detection of a high-density brachiolaria-stage larval population of Crown-of-Thorns sea star (Acanthaster planci) in Sekisei lagoon (Okinawa, Japan). Diversity 2016, 8, 9. [Google Scholar] [CrossRef]




| Model | k | LL | AICc | wAICc | Rm | Rc |
|---|---|---|---|---|---|---|
| Filtration rate | ||||||
| Species * IC + Species * LA + (1|date) | 17 | −324.8 | 744.7 | 0.920 | 65.70 | 81.80 |
| Species * IC + Species * LA + LA + (1|date) | 18 | −322.4 | 749.5 | 0.080 | 64.50 | 82.10 |
| 1 + (1|date) | 3 | −426.1 | 858.4 | 0.000 | 0.00 | 39.80 |
| Ingestion rate | ||||||
| Species * LA + IC + (1|date) | 13 | −339 | 740.3 | 0.996 | 73.70 | 90.90 |
| Species + IC + LA + (1|date) | 9 | −357.4 | 751.2 | 0.004 | 73.70 | 91.40 |
| 1 + (1|date) | 3 | −447.3 | 900.9 | 0.000 | 0.00 | 41.20 |
| Model | k | LL | AICc | wAICc | Rm | Rc |
|---|---|---|---|---|---|---|
| Filtration rate | ||||||
| Species * IC * IC dun + LA + (1|date) | 22 | −317.1 | 779.9 | 0.980 | 27.0 | 63.5 |
| Species * IC * IC dun + LA + LA + (1|date) | 23 | −315 | 788.7 | 0.012 | 29.0 | 68.6 |
| Species * IC + Species * IC dun + Species * LA + (1|date) | 21 | −328.3 | 790.2 | 0.006 | 27.5 | 67.5 |
| Species * IC + Species * IC dun + LA + (1|date) | 17 | −349.7 | 792.3 | 0.002 | 26.8 | 64.3 |
| 1 + (1|date) | 3 | −422.4 | 851.1 | 0.000 | 0.0 | 38.0 |
| Ingestion rate | ||||||
| Species * IC * IC dun + LA + (1|date) | 22 | −243.8 | 633.3 | 0.980 | 71.4 | 87.2 |
| Species * IC + Species * IC dun + LA + (1|date) | 17 | −274.6 | 642.3 | 0.011 | 69.2 | 86.7 |
| Species * IC * IC dun + LA + LA + (1|date) | 23 | −242.4 | 643.6 | 0.006 | 71.3 | 88.0 |
| 1 + (1|date) | 3 | −399.4 | 805 | 0.000 | 0.0 | 27.8 |
| Model | k | LL | AICc | wAICc | Rm | Rc |
|---|---|---|---|---|---|---|
| Filtration rate | ||||||
| Species * IC * IC dun + LA + (1|date) | 22 | −317.3 | 780.4 | 0.985 | 75.0 | 79.4 |
| Species * IC * IC dun + LA + LA + (1|date) | 23 | −316 | 790.7 | 0.006 | 74.3 | 79.6 |
| Species * IC + Species * IC dun + (1|date) | 16 | −353.3 | 790.9 | 0.005 | 75.5 | 80.1 |
| Species * IC + Species * IC dun + LA + (1|date) | 17 | −349.7 | 792.4 | 0.002 | 75.1 | 80.0 |
| Species * IC + Species * IC dun + Species * LA + (1|date) | 21 | −329.6 | 793.0 | 0.002 | 74.7 | 79.6 |
| 1 + (1|date) | 3 | −461.6 | 929.4 | 0.000 | 0.0 | 39.9 |
| Ingestion rate | ||||||
| Species * IC * IC dun + LA + (1|date) | 22 | −243.2 | 632.2 | 0.980 | 83.4 | 91.9 |
| Species * IC + Species * IC dun + LA + (1|date) | 17 | −274 | 640.9 | 0.012 | 81.2 | 91.2 |
| Species * IC * IC dun + LA + LA + (1|date) | 23 | −242.2 | 643.1 | 0.004 | 83.1 | 92.1 |
| Species * IC + Species * IC dun + (1|date) | 16 | −279.6 | 643.5 | 0.003 | 78.9 | 91.4 |
| 1 + (1|date) | 3 | −410.1 | 826.5 | 0.000 | 0.0 | 45.0 |
| Variable (unit) | Dunaliella sp. | T. lutea | Chaetoceros sp. | P. lutheri | P. tricornutum |
|---|---|---|---|---|---|
| Length (μm) | 7.92 | 5.01 | 9.18 | 5.91 | 18.92 |
| (7.37–8.47) | (4.77–5.25) | (8.65–9.72) | (5.51–6.3) | (17.94–19.9) | |
| Width (μm) | 5.43 | 5.7 | 6.5 | 5.17 | 3.48 |
| (5.15–5.71) | (5.46–5.94) | (6.1–6.9) | (4.94–5.4) | (3.31–3.64) | |
| Carbon (pg/cell) | 33.29 | 15.39 | 49.87 | 22.97 | 24.2 |
| (33.01–33.58) | (15.09–15.68) | (48.56–51.18) | (22.14–23.79) | (23.71–24.69) | |
| Nitrogen (pg/cell) | 6.58 | 2.53 | 7.9 | 3.57 | 3.85 |
| (6.54–6.61) | (2.44–2.62) | (7.44–8.36) | (3.52–3.62) | (3.73–3.97) | |
| C/N | 5.06 | 6.07 | 6.31 | 6.42 | 6.27 |
| (4.99–5.13) | (5.97–6.17) | (6.11–6.51) | (6.28–6.57) | (6.2–6.34) | |
| Chlorophyll (pg/cell) | 1.05 | 0.35 | 0.75 | 0.58 | 0.31 |
| (0.99–1.12) | (0.32–0.38) | (0.7–0.8) | (0.55–0.6) | (0.3–0.33) | |
| Lipid (pg/cell) | 15.58 | 6.79 | 17.33 | 10.54 | 9.93 |
| (15.49–15.66) | (5.48–8.09) | (16.53–18.13) | (8.96–12.12) | (9.5–10.36) | |
| Protein (pg/cell) | 32.9 | 12.65 | 39.5 | 17.85 | 19.25 |
| (32.7–33.05) | (12.2–13.1) | (37.2–41.8) | (17.6–18.1) | (18.65–19.85) | |
| Energy (10−9 kJ/cell) | 1.39 | 0.57 | 1.62 | 0.84 | 0.85 |
| (1.38–1.40) | (0.50–0.63) | (1.53–1.70) | (0.77–0.91) | (0.81–0.88) |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mellin, C.; Lugrin, C.; Okaji, K.; Francis, D.S.; Uthicke, S. Selective Feeding and Microalgal Consumption Rates by Crown-Of-Thorns Seastar (Acanthaster cf. solaris) Larvae. Diversity 2017, 9, 8. https://doi.org/10.3390/d9010008
Mellin C, Lugrin C, Okaji K, Francis DS, Uthicke S. Selective Feeding and Microalgal Consumption Rates by Crown-Of-Thorns Seastar (Acanthaster cf. solaris) Larvae. Diversity. 2017; 9(1):8. https://doi.org/10.3390/d9010008
Chicago/Turabian StyleMellin, Camille, Claire Lugrin, Ken Okaji, David S. Francis, and Sven Uthicke. 2017. "Selective Feeding and Microalgal Consumption Rates by Crown-Of-Thorns Seastar (Acanthaster cf. solaris) Larvae" Diversity 9, no. 1: 8. https://doi.org/10.3390/d9010008
APA StyleMellin, C., Lugrin, C., Okaji, K., Francis, D. S., & Uthicke, S. (2017). Selective Feeding and Microalgal Consumption Rates by Crown-Of-Thorns Seastar (Acanthaster cf. solaris) Larvae. Diversity, 9(1), 8. https://doi.org/10.3390/d9010008