
The Phylogeny and Biogeography of Phyla nodiflora (Verbenaceae) Reveals Native and Invasive Lineages throughout the World

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Species
2.2. Phylogenetics
2.2.1. Plant Material Collection
2.2.2. DNA Extraction, Amplification and Sequencing
2.2.3. Sequence Data Preparation
2.2.4. Consensus Tree Analyses
2.2.5. Parsimony Network Analyses
2.3. Biogeography—Delimitation of Native-Range Distributions of P. nodiflora Clades
3. Results
3.1. Phylogenetics
3.1.1. Characteristics of the ITS Sequences and Ribotype Discovery
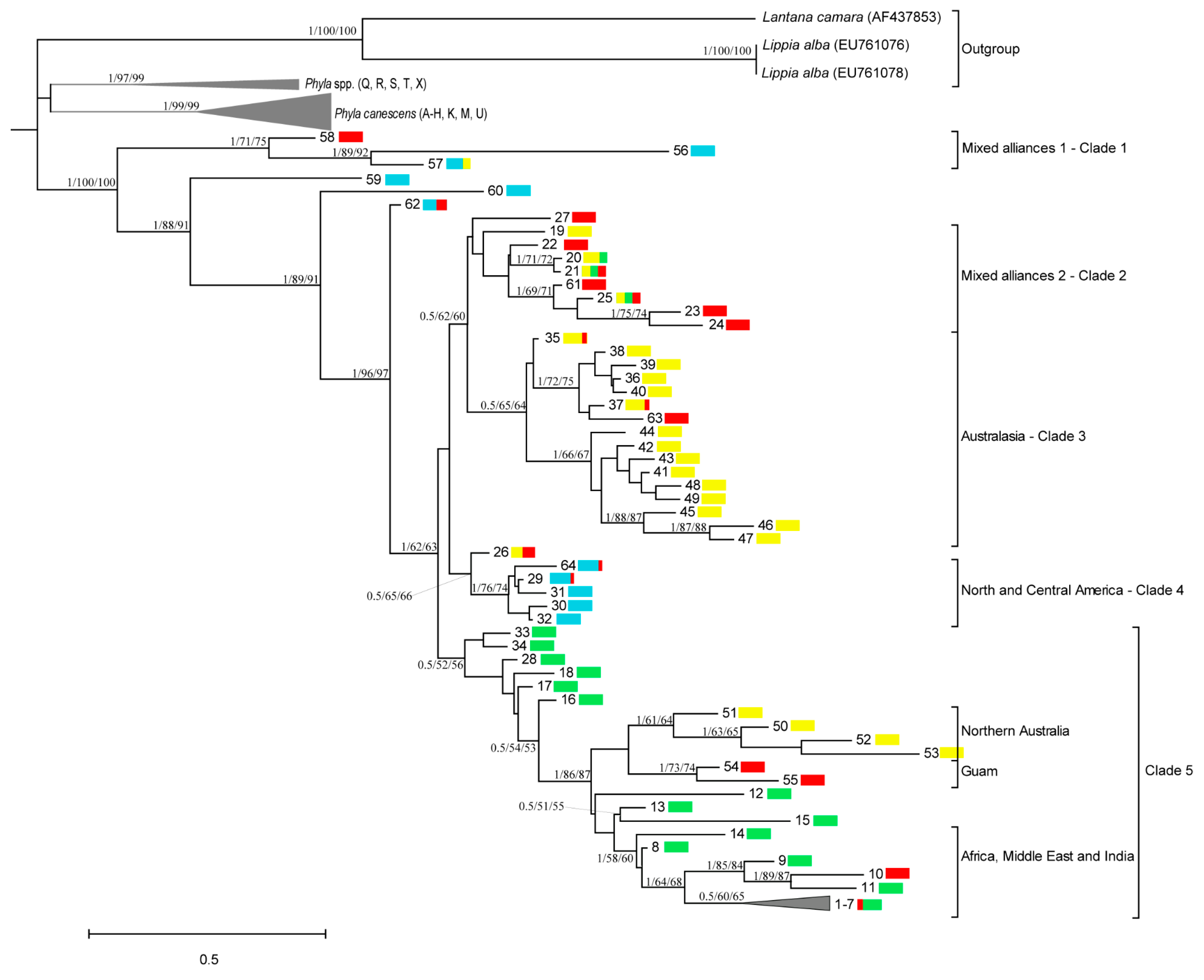
3.1.2. Consensus Trees
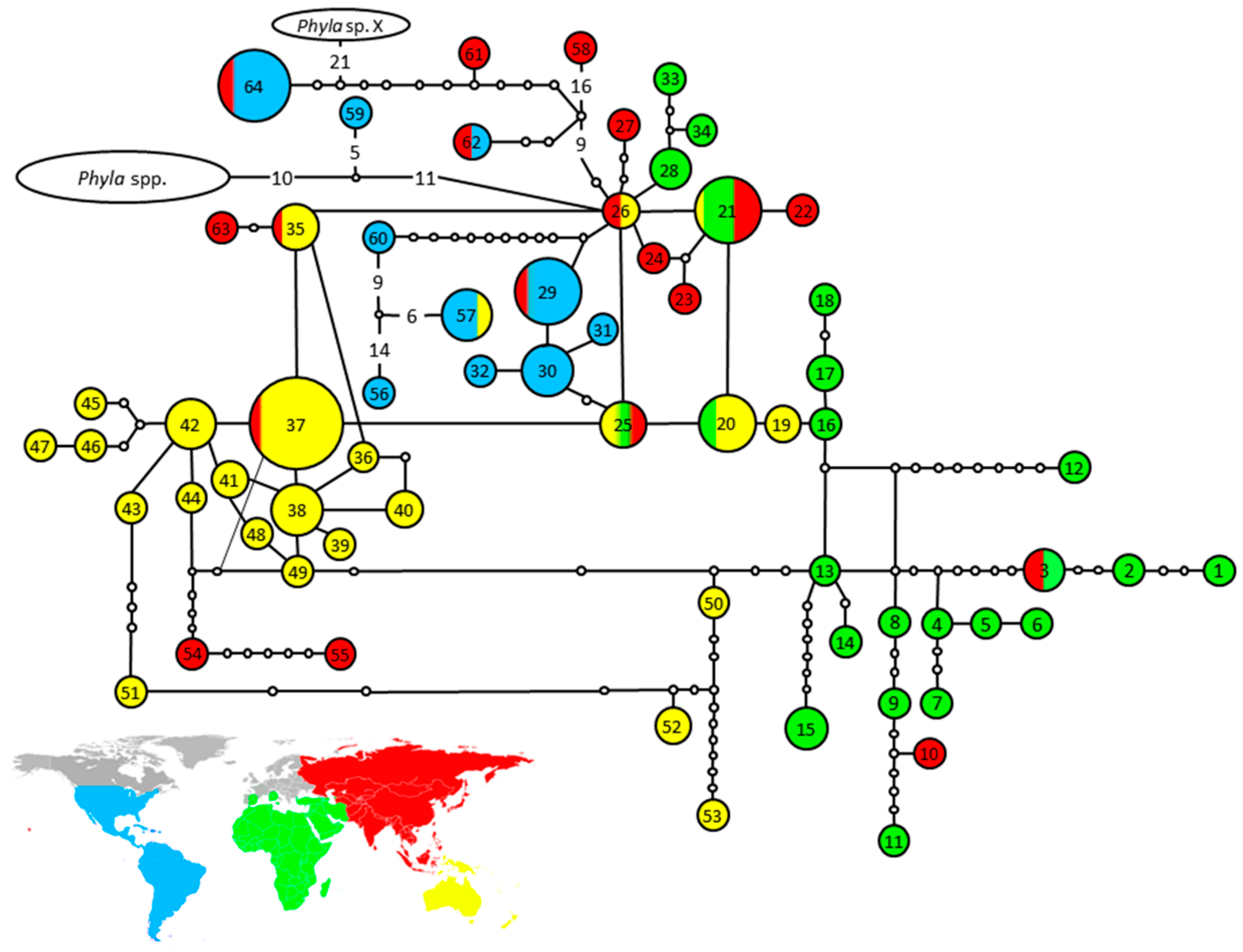
3.1.3. Parsimony Network
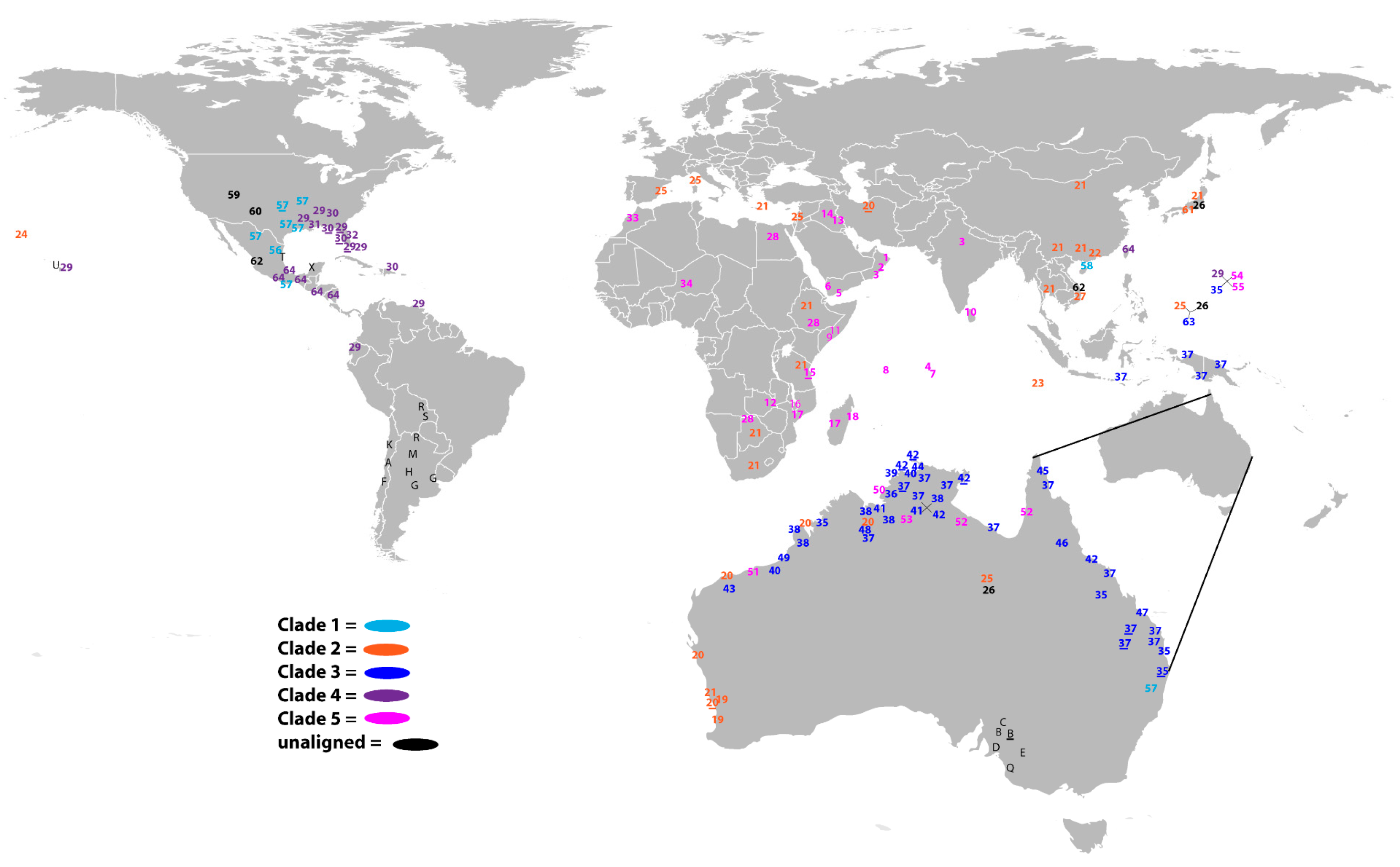
3.2. Biogeography—Delimitation of Native-Range Distributions of P. nodiflora Clades
3.2.1. Clade 1—Mixed Alliances 1: Americas (North, Central, Northern South America)
3.2.2. Clade 2—Mixed Alliances 2: Asia, Africa, Australia
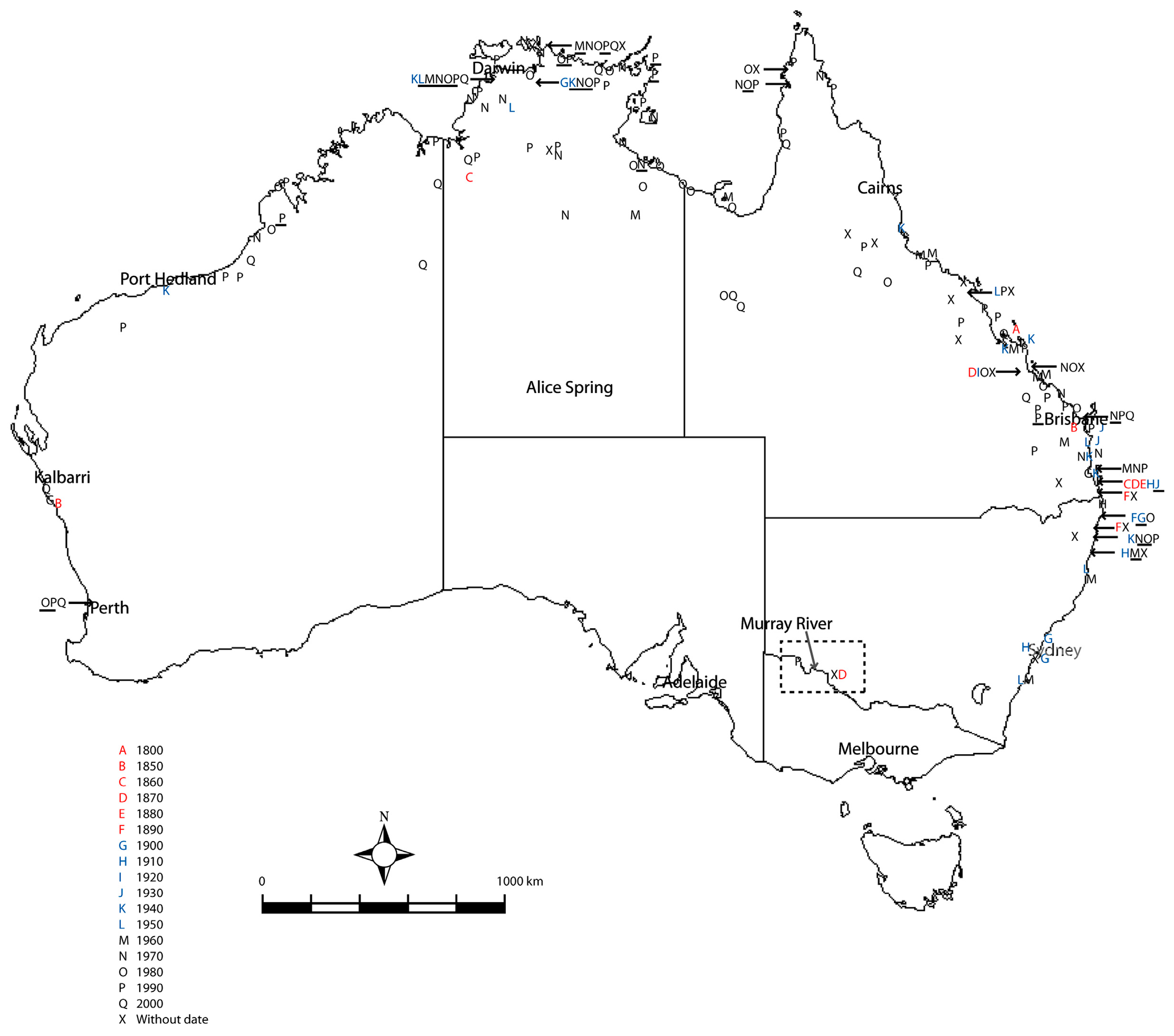
3.2.3. Clade 3: Australasia
3.2.4. Clade 4—North and Central America with Asia
3.2.5. Clade 5—Northern Australia, Guam, Africa, Middle East and India
4. Discussion
4.1. Molecular Markers
4.2. The Global Expansion of Phyla nodiflora Involves Native and Invasive Lineages
4.3. Natural Dispersal
4.4. Human-Assisted Dispersal
4.5. Native Lineages Outside of the Americas Have Had Secondary Emigration Events
4.6. Taxonomic Considerations
4.7. Implications for Conservation
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A. Methods for the Chloroplast Regions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Forward | Reverse | Tm | Length | Source | ||
|---|---|---|---|---|---|---|---|
| 3′rps16/5′trnK | (UUU) F | AAAGTGGGTTTTTATGATCC | (UUU) R | TTAAAAGCCGAGTACTCTACC | 56 | 631 | [89] |
| trnL-trnF | UniE | GGTTCAAGTCCCTCTATCCC | UniF | ATTTGAACTGGTGACACGAG | 57 | 273–392 | [90] |
| petB intron | petB intron F | AGAGATGGTTCTACTTCGTC | petB intron R | ACTTTCATCTCGTACAGCTC | 57 | 552 | [88] |
References
- Pyšek, P.; Richardson, D.M.; Rejmánek, M.; Webster, G.L.; Williamson, M.; Kirschner, J. Alien plants in checklists and floras: Towards better communication between taxonomists and ecologists. Taxon 2004, 53, 131–143. [Google Scholar] [CrossRef]
- Gormally, C.; Donovan, J.H.A. Genetic structure of a widely dispersed beach annual, Cakile edentula (Brassicaceae). Am. J. Bot. 2011, 98, 1657–1662. [Google Scholar] [CrossRef] [PubMed]
- Der, J.P.; Thomson, J.A.; Stratford, J.K.; Wolf, P.G. Global chloroplast phylogeny and biogeography of bracken (Pteridium; Dennstaedtiaceae). Am. J. Bot. 2009, 96, 1041–1049. [Google Scholar] [CrossRef] [PubMed]
- Horrocks, M.; Rechtman, R.B. Sweet potato (Ipomoea batatas) and banana (Musa sp.) microfossils in deposits from the Kona Field System, Island of Hawaii. JAS 2009, 36, 1115–1126. [Google Scholar] [CrossRef]
- Carlton, J.T. Biological invasions and cryptogenic species. Ecology 1996, 77, 1653–1655. [Google Scholar] [CrossRef]
- Kennedy, K.L. A Systematic Study of the Genus Phyla Lour (Verbenaceae: Verbenoideae, Lantanae). Ph.D. Thesis, The University of Texas at Austin, Austin, TX, USA, 1992. [Google Scholar]
- O’Leary, N.; Múlgura, M.E. A Taxonomic Revision of the Genus Phyla (Verbenaceae). Ann. Mo. Bot. Gard. 2012, 98, 578–596. [Google Scholar] [CrossRef]
- Munir, A.A. A taxonomic revision of the genus Phyla Lour.(Verbenaceae) in Australia. J. Adel. Bot. Gard. 1993, 15, 109–128. [Google Scholar]
- Greene, E. Neglected generic types I. Pittonia 1899, 4, 45–51. [Google Scholar]
- Gupta, A.; Sadasivaiah, B.; Bhat, G. Phyla nodiflora. Version 2016-3. Available online: www.iucnredlist.org (accessed on 24 February 2017).
- Marx, H.E.; O’Leary, N.; Yuan, Y.-W.; Lu-Irving, P.; Tank, D.C.; Múlgura, M.E.; Olmstead, R.G. A molecular phylogeny and classification of Verbenaceae. Am. J. Bot. 2010, 97, 1647–1663. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, B. Shoalwater Bay: Settlers in a Queensland Wilderness; Central Queensland University Press: Rockhampton, Australia, 1996. [Google Scholar]
- Hawkins, J.; Boutaoui, N.; Cheung, K.; van Klinken, R.; Hughes, C. Intercontinental dispersal prior to human translocation revealed in a cryptogenic invasive tree. New Phytol. 2007, 175, 575–587. [Google Scholar] [CrossRef] [PubMed]
- Webb, D. What are the criteria for presuming native status? Watsonia 1985, 15, 231–236. [Google Scholar]
- Bean, A.R. A new system for determining which plant species are indigenous in Australia. Aust. Syst. Bot. 2007, 20, 1–43. [Google Scholar] [CrossRef]
- Steven, W.; Ulloa, C.; Pool, A.; Montiel, O. Flora de Nicaragua. In Monographs in Systematic Botany from the Missouri Botanical Garden; Missouri Botanical Garden: St. Louis, MO, USA, 1978. [Google Scholar]
- Macdonald, M.J. Ecology of Phyla canescens (Verbenaceae) in Australia. Ph.D. Thesis, University of New England, Armidale, Australia, 2008. [Google Scholar]
- Gross, C.; Gorrell, L.; Macdonald, M.; Fatemi, M. Honeybees facilitate the invasion of Phyla canescens (Verbenaceae) in Australia–no bees, no seed! Weed Res. 2010, 50, 364–372. [Google Scholar] [CrossRef]
- Roalson, E.; Friar, E. Phylogenetic relationships and biogeographic patterns in North American members of Carex section Acrocystis (Cyperaceae) using nrDNA ITS and ETS sequence data. Plant Syst. Evol. 2004, 243, 175–187. [Google Scholar] [CrossRef]
- Buckler, E.S.; Ippolito, A.; Holtsford, T.P. The evolution of ribosomal DNA divergent paralogues and phylogenetic implications. Genetics 1997, 145, 821–832. [Google Scholar] [PubMed]
- Consaul, L.L.; Gillespie, L.J.; Waterway, M.J. Evolution and polyploid origins in North American Arctic Puccinellia (Poaceae) based on nuclear ribosomal spacer and chloroplast DNA sequences. Am. J. Bot. 2010, 97, 324–336. [Google Scholar] [CrossRef] [PubMed]
- Mäder, G.; Zamberlan, P.M.; Fagundes, N.J.; Magnus, T.; Salzano, F.M.; Bonatto, S.L.; Freitas, L.B. The use and limits of ITS data in the analysis of intraspecific variation in Passiflora L. (Passifloraceae). Genet. Mol. Biol. 2010, 33, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Feliner, G.N.; Rosselló, J.A. Better the devil you know? Guidelines for insightful utilization of nrDNA ITS in species-level evolutionary studies in plants. Mol. Phylogen. Evol. 2007, 44, 911–919. [Google Scholar] [CrossRef] [PubMed]
- Chenna, R.; Sugawara, H.; Koike, T.; Lopez, R.; Gibson, T.J.; Higgins, D.G.; Thompson, J.D. Multiple sequence alignment with the Clustal series of programs. Nucleic Acids Res. 2003, 31, 3497–3500. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.; Clamp, M.; Barton, G.J. Jalview Version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed]
- Swofford, D.L. PAUP*. Phylogenetic Analysis Using Parsimony (* and Other Methods), Version 4; Sinauer Associates: Sunderland, MA, USA, 2003.
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed]
- Ogden, T.H.; Rosenberg, M.S. How should gaps be treated in parsimony? A comparison of approaches using simulation. Mol. Phylogen. Evol. 2007, 42, 817–826. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Posada, D.; Crandall, K.A. Modeltest: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. Population Genetics, Molecular Evolution, and the Neutral Theory: Selected Papers; University of Chicago Press: Chicago, IL, USA, 1994; p. 704. [Google Scholar]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [PubMed]
- Posada, D.; Crandall, K.A. Intraspecific gene genealogies: Trees grafting into networks. Trends Ecol. Evol. 2001, 16, 37–45. [Google Scholar] [CrossRef]
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef] [PubMed]
- Templeton, A.R.; Crandall, K.A.; Sing, C.F. A cladistic analysis of phenotypic associations with haplotypes inferred from restriction endonuclease mapping and DNA sequence data. III. Cladogram estimation. Genetics 1992, 132, 619–633. [Google Scholar] [PubMed]
- Joly, S.; Stevens, M.I.; van Vuuren, B.J. Haplotype networks can be misleading in the presence of missing data. Syst. Biol. 2007, 56, 857–862. [Google Scholar] [CrossRef] [PubMed]
- Tobler, M.; Honorio, E.; Janovec, J.; Reynel, C. Implications of collection patterns of botanical specimens on their usefulness for conservation planning: An example of two neotropical plant families (Moraceae and Myristicaceae) in Peru. Biodivers. Conserv. 2007, 16, 659–677. [Google Scholar] [CrossRef]
- Genc, H.; Nation, J.L.; Emmel, T.C. Life history and biology of Phyciodes phaon (Lepidoptera: Nymphalidae). Fla. Entomol. 2003, 86, 445–449. [Google Scholar] [CrossRef]
- Bentham, G. Flora Australiensis: A Description of the Plants of the Australian Territory; Reeve & Co.: London, UK, 1870; Volume 5, pp. 31–70. [Google Scholar]
- Song, Z.C.; Wang, W.M.; Huang, F. Fossil pollen records of extant angiosperms in China. Bot. Rev. 2004, 70, 425–458. [Google Scholar]
- Robinson, G.S.; Ackery, P.R.; Kitching, I.J.; Beccaloni, G.W.; Hernández, L.M. HOSTS―A Database of the World’s Lepidopteran Hostplants. Available online: http://www.nhm.ac.uk/hosts (accessed on 23 February 2017).
- Grey, G. Journals of Two Expeditions of Discovery in North-West and Western Australia, during the Years 1837, 38, and 39; T and W Boone: London, UK, 1841; Volume II. [Google Scholar]
- Knibbs, G.H. Commonwealth Bureau of Census and Statistics Melbourne; Australian Government: Melbourne, Australia, 1909.
- Day, K.J.; Forster, B.A. Report on the land units of Croker Island, N.T.; Department of Northern Australia, Land Conservation Section: Canberra, Australia, 1975.
- Edwards, E.D.; Newland, J.; Regan, L. Lepidoptera: Hesperioidea, Papilionoidea; CSIRO Publishing: Melbourne, Australia, 2001; Volume 31.6, pp. 545, 615. [Google Scholar]
- Leigh, C.; Walton, C.S. Lippia (Phyla canescens) in Queensland; Land Protection, Department of Natural Resources, Mines, and Energy: Brisbane, Australia, 2004.
- Price, J.; Macdonald, M.; Gross, C.; Whalley, R.D.; Simpson, I. Vegetative reproduction facilitates early expansion of Phyla canescens in a semi-arid floodplain. Biol. Invasions 2011, 13, 285–289. [Google Scholar] [CrossRef]
- Xu, C.Y.; Julien, M.H.; Fatemi, M.; Girod, C.; Van Klinken, R.D.; Gross, C.L.; Novak, S.J. Phenotypic divergence during the invasion of Phyla canescens in Australia and France: Evidence for selection-driven evolution. Ecol. Lett. 2010, 13, 32–44. [Google Scholar] [CrossRef] [PubMed]
- Cilento, R.; Lack, C. Triumph in the Tropics: An Historical Sketch of Queensland/compiled and edited by Sir Raphael Cilento; with the assistance of Clem Lack; for the Historical Committee of the Centenary Celebrations Council of Queensland; Smith & Paterson: Brisbane, Australia, 1959; p. 446. [Google Scholar]
- Mueller, F. North Australian botany, observations on, by Dr Frederick (sic) Mueller, botanist to the NW Australian Government Expedition, under the command of Mr Surveyor Gregory; in a letter to Sir WJ Hooker (published with the sanction of the Colonial Office). Hooker’s J. Bot. Kew Gard. Misc. 1856, 8, 321–331. [Google Scholar]
- Mueller, F. Botanical report on the North Australian exploring expedition, under the command of A.C. Gregory Esq. J. Proc. Linn. Soc. Lond. Bot. 1858, 2, 137–144. [Google Scholar] [CrossRef]
- Knibbs, G.H. Official Year Book of the Commonwealth of Australia Containing Authoritative Statistics for the Period 1901–1908 and Corrected Statistics for the Period 1788 to 1900. No. 2; Melbourne, Australia, 1909. [Google Scholar]
- Dupont, L.; Behling, H.; Kim, J.-H. Thirty thousand years of vegetation development and climate change in Angola (Ocean Drilling Program Site 1078). CliPa 2008, 4, 107–124. [Google Scholar]
- Dangol, D.; Gurung, S. Ethnobotany of the Tharu tribe of Chitwan district, Nepal. Int. J. Pharm. 1991, 29, 203–209. [Google Scholar] [CrossRef]
- Shukla, S.; Patel, R.; Kukkar, R. Study of phytochemical and diuretic potential of methanol and aqueous extracts of aerial parts of Phyla nodiflora Linn. Int. J. Pharm. Pharm. Sci. 2009, 1, 85–91. [Google Scholar]
- Wang, Y.-C.; Huang, T.-L. Screening of anti-Helicobacter pylori herbs deriving from Taiwanese folk medicinal plants. FEMS Immunol. Med. Microbiol. 2005, 43, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.B.A.; Gouthaman, T.; Rao, A.S.; Rao, M.V. Micropropagation of Phyla nodiflora (L.) Greene: An important medicinal plant. Iran. J. Biotechnol. 2005, 3, 186–190. [Google Scholar]
- Abuodha, J.; Musila, W.; van der Hagen, H.; van der Meulen, F. Floristic composition and vegetation ecology of the Malindi Bay coastal dune field, Kenya. J. Coast. Conserv. 2003, 9, 97–112. [Google Scholar] [CrossRef]
- Estes, J.R.; Brown, L.S. Entomophilous, intrafloral pollination in Phyla incisa. Am. J. Bot. 1973, 60, 228–230. [Google Scholar] [CrossRef]
- Pascual, M.; Slowing, K.; Carretero, E.; Mata, D.S.; Villar, A. Lippia: Traditional uses, chemistry and pharmacology: A review. J. Ethnopharmacol. 2001, 76, 201–214. [Google Scholar] [CrossRef]
- Smith, D.R. Mutation rates in plastid genomes: They are lower than you might think. Gen. Biol. Evol. 2015, 7, 1227–1234. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, K.H.; Li, W.-H.; Sharp, P.M. Rates of nucleotide substitution vary greatly among plant mitochondrial, chloroplast, and nuclear DNAs. Proc. Natl. Acad. Sci. USA 1987, 84, 9054–9058. [Google Scholar] [CrossRef] [PubMed]
- Call, A.; Sun, Y.X.; Yu, Y.; Pearman, P.B.; Thomas, D.T.; Trigiano, R.N.; Carbone, I.; Xiang, Q.Y.J. Genetic structure and post-glacial expansion of Cornus florida L. (Cornaceae): integrative evidence from phylogeography, population demographic history, and species distribution modeling. J. Syst. Evol. 2015, 54, 136–151. [Google Scholar] [CrossRef]
- Rossetto, M.; McPherson, H.; Siow, J.; Kooyman, R.; Merwe, M.; Wilson, P.D. Where did all the trees come from? A novel multispecies approach reveals the impacts of biogeographical history and functional diversity on rain forest assembly. J. Biogeogr. 2015, 42, 2172–2186. [Google Scholar] [CrossRef]
- Diazgranados, M.; Barber, J.C. Geography shapes the phylogeny of frailejones (Espeletiinae Cuatrec., Asteraceae): A remarkable example of recent rapid radiation in sky islands. PeerJ 2017, 5, e2968. [Google Scholar] [CrossRef] [PubMed]
- Verdcourt, B. Flora of Tropical East Africa. In Verbenaceae; CRC Press: London, UK, 1992. [Google Scholar]
- Renner, S. Plant dispersal across the tropical Atlantic by wind and sea currents. Int. J. Plant Sci. 2004, 165, S23–S33. [Google Scholar] [CrossRef]
- Mummenhoff, K.; Linder, P.; Friesen, N.; Bowman, J.L.; Lee, J.-Y.; Franzke, A. Molecular evidence for bicontinental hybridogenous genomic constitution in Lepidium sensu stricto (Brassicaceae) species from Australia and New Zealand. Am. J. Bot. 2004, 91, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Pettingill, O.S. Additional information on the food of the American woodcock. Wilson Bull. 1939, 51, 78–82. [Google Scholar]
- Rundle, W.D.; Sayre, M.W. Feeding ecology of migrant Soras in southeastern Missouri. J. Wildl. Manag. 1983, 47, 1153–1159. [Google Scholar] [CrossRef]
- Razi, B.A. A contribution towards the study of the dispersal mechanisms in flowering plants of Mysore (south India). Ecology 1950, 31, 282–286. [Google Scholar] [CrossRef]
- Dostine, P.; Morton, S.R. Seasonal abundance and diet of the comb-crested Jacana Irediparra gallinacea in the tropical Northern Territory. Emu-Austral Ornithol. 2000, 100, 299–311. [Google Scholar] [CrossRef]
- Takayama, K.; Kajita, T.; Murata, J.; Tateishi, Y. Phylogeography and genetic structure of Hibiscus tiliaceus—Speciation of a pantropical plant with sea-drifted seeds. Mol. Ecol. 2006, 15, 2871–2881. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, A.H.; McKee, K.L.; Mendelssohn, I.A. The influence of vegetation, salinity, and inundation on seed banks of oligohaline coastal marshes. Am. J. Bot. 1996, 83, 470–479. [Google Scholar] [CrossRef]
- Brookes, M.; Barley, R. Plants Listed in Nursery Catalogues in Victoria 1855–1889; Published for the Ornamental Plant Collections Association Inc., Royal Botanic Gardens: Melbourne, Australia, 1992. [Google Scholar]
- Lorenz’s OK Seeds. Available online: http://www.lorenzsokseedsllc.com/?s=lippia (accessed on 26 February 2017).
- Moldenke, H. Verbenaceae; Amerind Publishing Co. Pvt. Ltd.: New Delhi, India, 1983. [Google Scholar]
- Barker, R.; Telford, I. Oceanic Islands 2. In Flora of Australia; Australian Biological Resources Study and the Australian Government Publishing Service: Canberra, Australia, 1993; Volume 50, p. 415. [Google Scholar]
- Hussey, B.; Keighery, G.; Dodd, J.; Lloyd, S.; Cousens, R. Western Weeds: A Guide to the Weeds of Western Australia; The Plant Protection Society of Western Australia: Victoria Park, Australia, 2007. [Google Scholar]
- HMAS Inverell. Available online: http://www.navy.gov.au/hmas-inverell (accessed on 26 February 2017).
- Townsend, C. Verbenaceae. In Flora of Turkey and the East Aegean Islands; Davis, P., Ed.; Edinburgh University Press: Edinburgh, UK, 1982; pp. 31–35. [Google Scholar]
- Germishuizen, G.; Meyer, N. Plants of Southern Africa: An Annotated Checklist; National Botanical Institute Pretoria: Pretoria, South Africa, 2003. [Google Scholar]
- Saltonstall, K. Cryptic invasion by a non-native genotype of the common reed, Phragmites australis, into North America. Proc. Natl. Acad. Sci. USA 2002, 99, 2445–2449. [Google Scholar] [CrossRef] [PubMed]
- Ellstrand, N.C.; Schierenbeck, K.A. Hybridization as a stimulus for the evolution of invasiveness in plants? Proc. Natl. Acad. Sci. USA 2000, 97, 7043–7050. [Google Scholar] [CrossRef] [PubMed]
- Mason-Gamer, R.J. Reticulate evolution, introgression, and intertribal gene capture in an allohexaploid grass. Syst. Biol. 2004, 53, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Miura, O. Molecular genetic approaches to elucidate the ecological and evolutionary issues associated with biological invasions. Ecol. Res. 2007, 22, 876–883. [Google Scholar] [CrossRef]
- Nishizawa, T. Primer pairs suitable for PCR-SSCP analysis of chloroplast DNA in angiosperms. J. Phytogeogr. Taxon 2000, 48, 63–66. [Google Scholar]
- Shaw, J.; Lickey, E.B.; Schilling, E.E.; Small, R.L. Comparison of whole chloroplast genome sequences to choose noncoding regions for phylogenetic studies in angiosperms: The tortoise and the hare III. Am. J. Bot. 2007, 94, 275–288. [Google Scholar] [CrossRef] [PubMed]
- Taberlet, P.; Gielly, L.; Pautou, G.; Bouvet, J. Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant Mol. Biol. 1991, 17, 1105–1109. [Google Scholar] [CrossRef] [PubMed]





| Criterion | Characteristics Likely to be Associated with a Native Species | Characteristics Likely to be Associated with an Invasive Species | Equivocal Information |
|---|---|---|---|
| 1 | (a) consistently occurs in intact unmodified habitat | (b) species known only from croplands, roadsides and other frequently disturbed sites | |
| 2 | (a) is not persistently invasive in its area of occurrence | (b) persistently invades or encroaches upon natural communities | |
| 3 | (a) is attended by a range of pests or diseases | (b) species that is pest- and disease-free | (c) Damage to herbarium material may have occurred post collecting |
| 4 | (a) displays a range of phenotypic or genetic diversities | (b) phenotypically or genetically uniform populations, probably derived from a single introduction | |
| 5 | (a) does not display any post-settlement expansion of geographical range within the region | (b) species that has a known or inferred expansion in its range over the past 100–150 years | |
| 6 | (a) any discontinuities of distribution of the species within the region are related to climatic and edaphic factors | (b) a species with a patchy distribution correlated with human settlement patterns is probably alien | |
| 7 | (a) a species is probably indigenous if closely related species occur as natives in Australia or nearby (e.g., New Guinea, Timor, Java, New Zealand) | (b) where the closest relatives occur on another continent, the species is likely to be an alien | |
| 8 | (b) a species known to be alien in areas outside the region must be under suspicion of being alien within the region | ||
| 9 | (b) a species is probably not indigenous to the region if the nearest occurrence outside the region represents a major disjunction | ||
| 10 | (b) the initial herbarium record dates well after the first European settlement of the region | (c) alternatively, if the initial herbarium record precedes or is soon after a European settlement, no useful conclusions can be drawn | |
| 11 | (a) plant has an established ethnobotanical use by indigenous peoples | (b) a written record exists of the introduction or importation of a species in a journal, nursery catalogue, or botanic gardens listing | (c) alternatively, if no written record exists, no useful conclusions can be drawn |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gross, C.L.; Fatemi, M.; Julien, M.; McPherson, H.; Van Klinken, R. The Phylogeny and Biogeography of Phyla nodiflora (Verbenaceae) Reveals Native and Invasive Lineages throughout the World. Diversity 2017, 9, 20. https://doi.org/10.3390/d9020020
Gross CL, Fatemi M, Julien M, McPherson H, Van Klinken R. The Phylogeny and Biogeography of Phyla nodiflora (Verbenaceae) Reveals Native and Invasive Lineages throughout the World. Diversity. 2017; 9(2):20. https://doi.org/10.3390/d9020020
Chicago/Turabian StyleGross, Caroline L., Mohammad Fatemi, Mic Julien, Hannah McPherson, and Rieks Van Klinken. 2017. "The Phylogeny and Biogeography of Phyla nodiflora (Verbenaceae) Reveals Native and Invasive Lineages throughout the World" Diversity 9, no. 2: 20. https://doi.org/10.3390/d9020020
APA StyleGross, C. L., Fatemi, M., Julien, M., McPherson, H., & Van Klinken, R. (2017). The Phylogeny and Biogeography of Phyla nodiflora (Verbenaceae) Reveals Native and Invasive Lineages throughout the World. Diversity, 9(2), 20. https://doi.org/10.3390/d9020020