Species Richness and Relative Abundance of Reef-Building Corals in the Indo-West Pacific

Abstract
:1. Introduction
- Diffusive—typically sparsely distributed across a broad geographic range;
- Geo-suffusive—geographically limited (occurring in a relatively small geographic area, e.g., northern Red Sea);
- Habitat-suffusive—habitat limited (confined to one or few habitats, e.g., reef flat, mesophotic zone).
2. Methods
2.1. Coral Identification
2.2. Assessing Abundance of Massive Porites
2.3. Analysis

3. Results
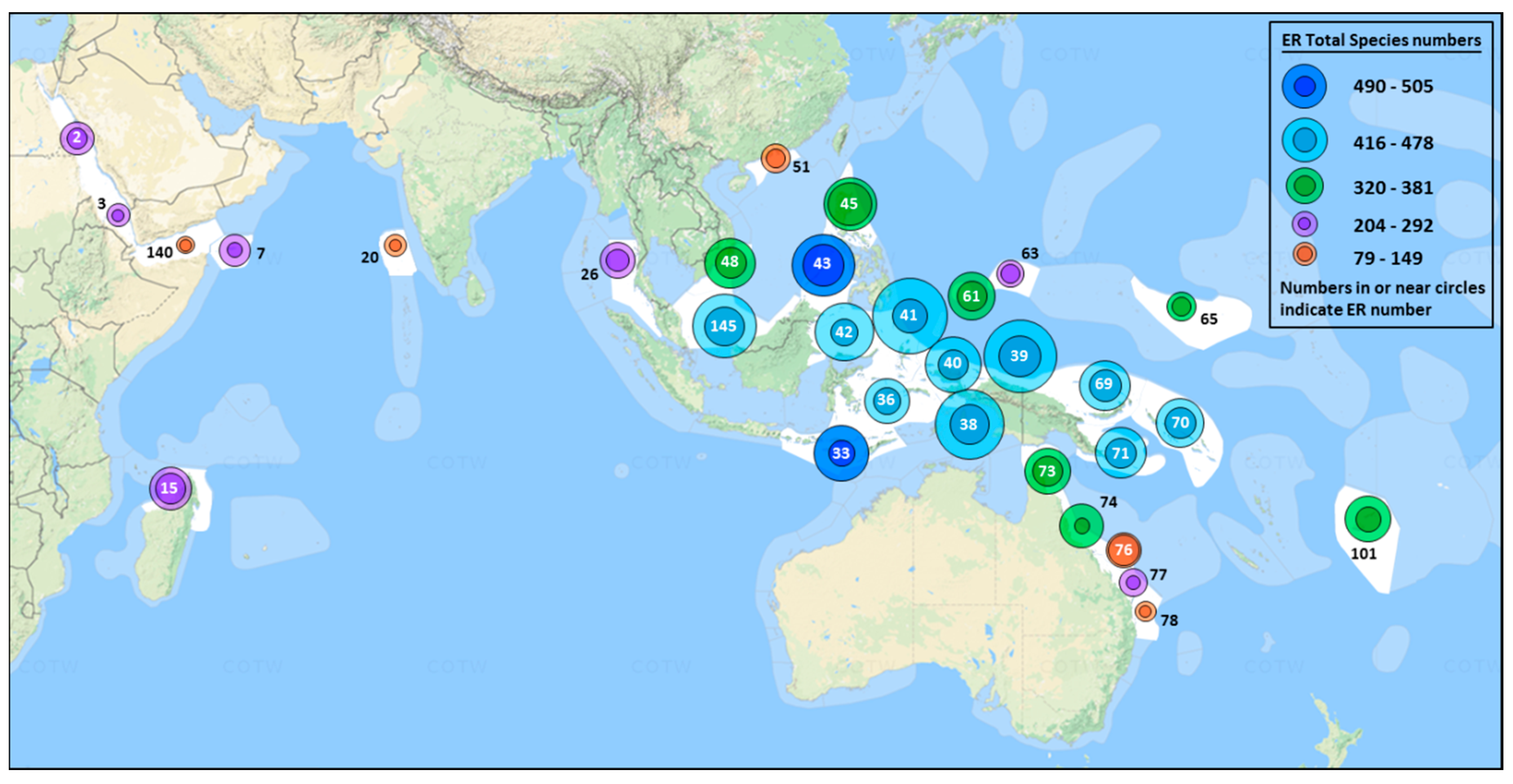
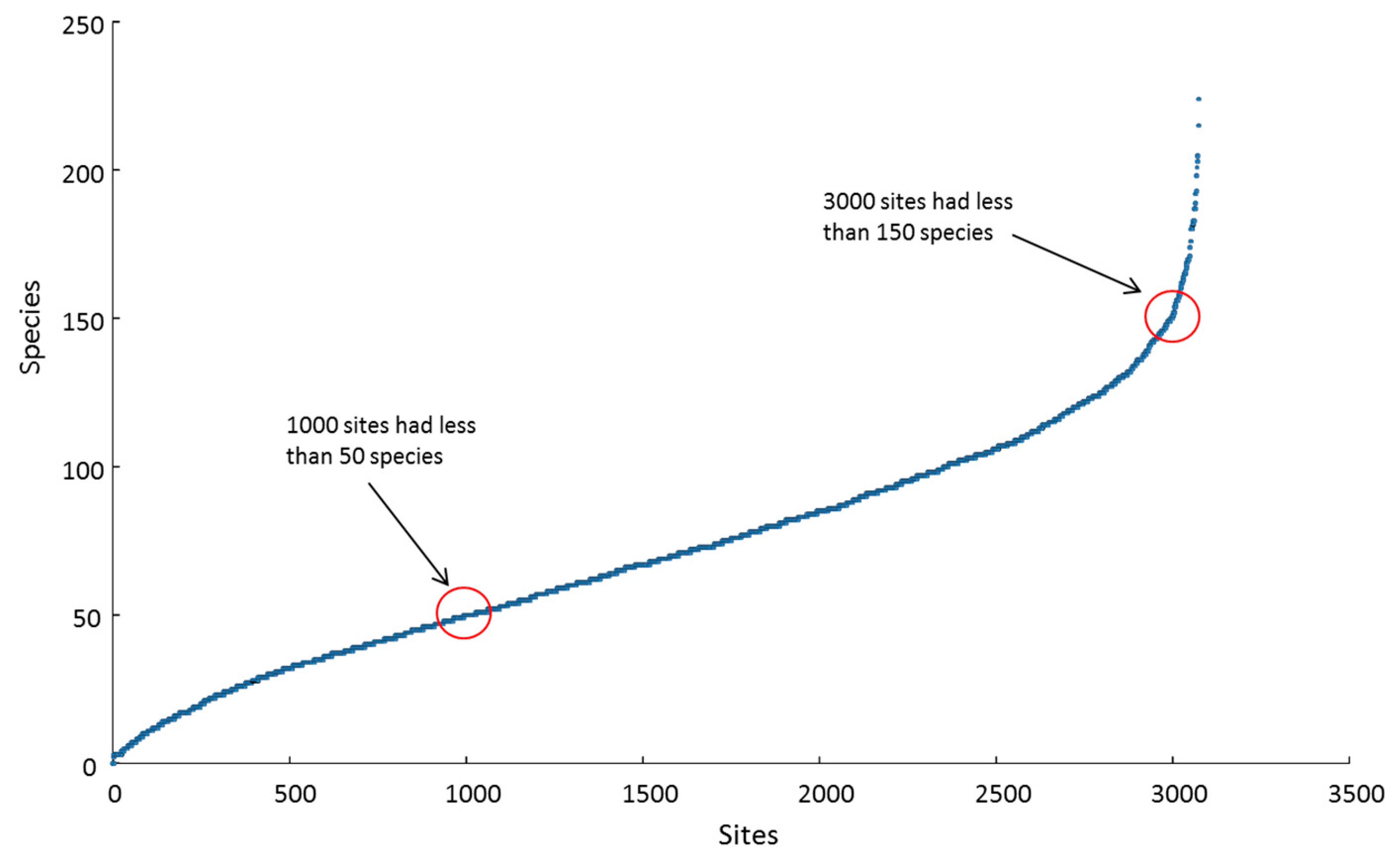
3.1. Species Distributions
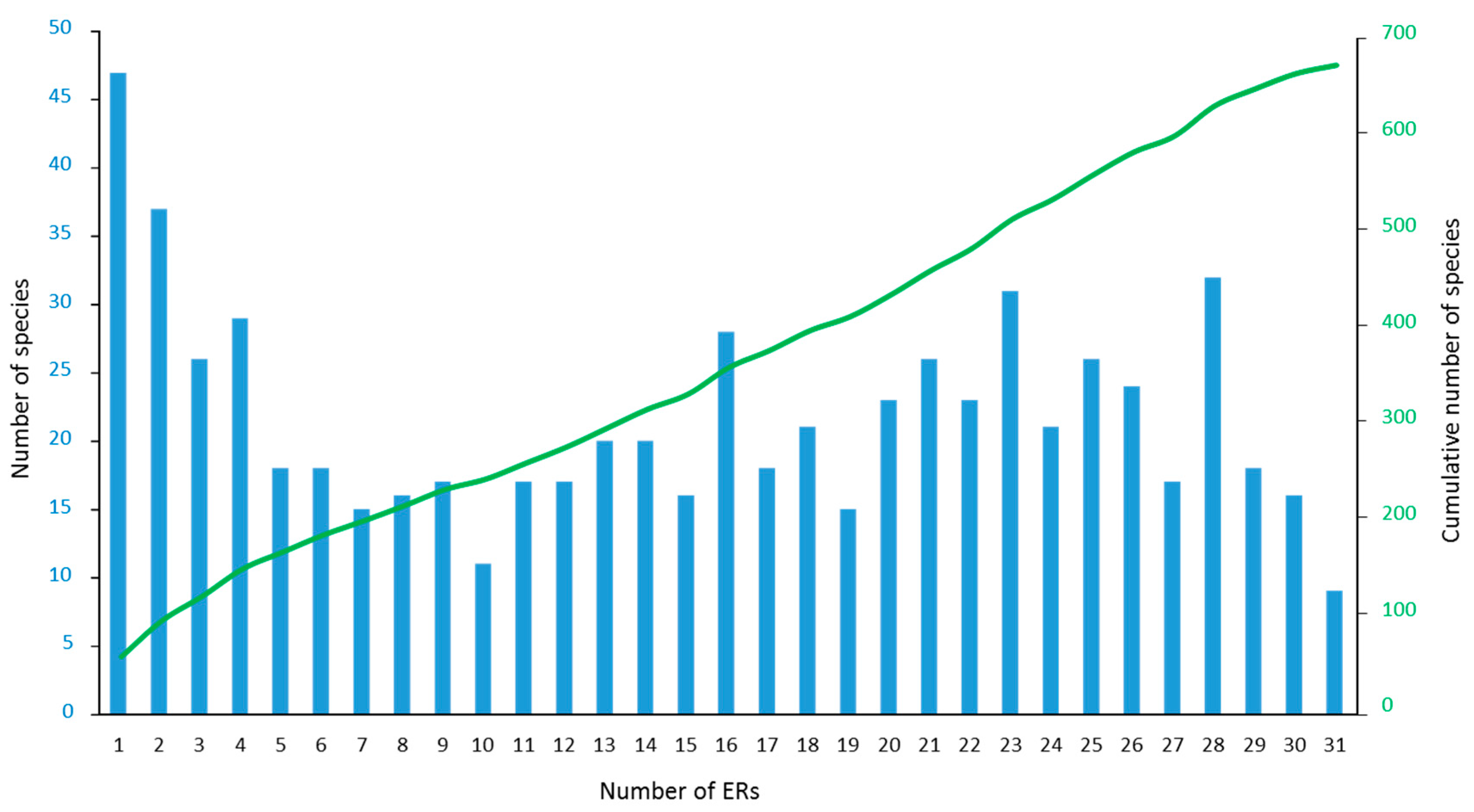
3.2. Species Richness
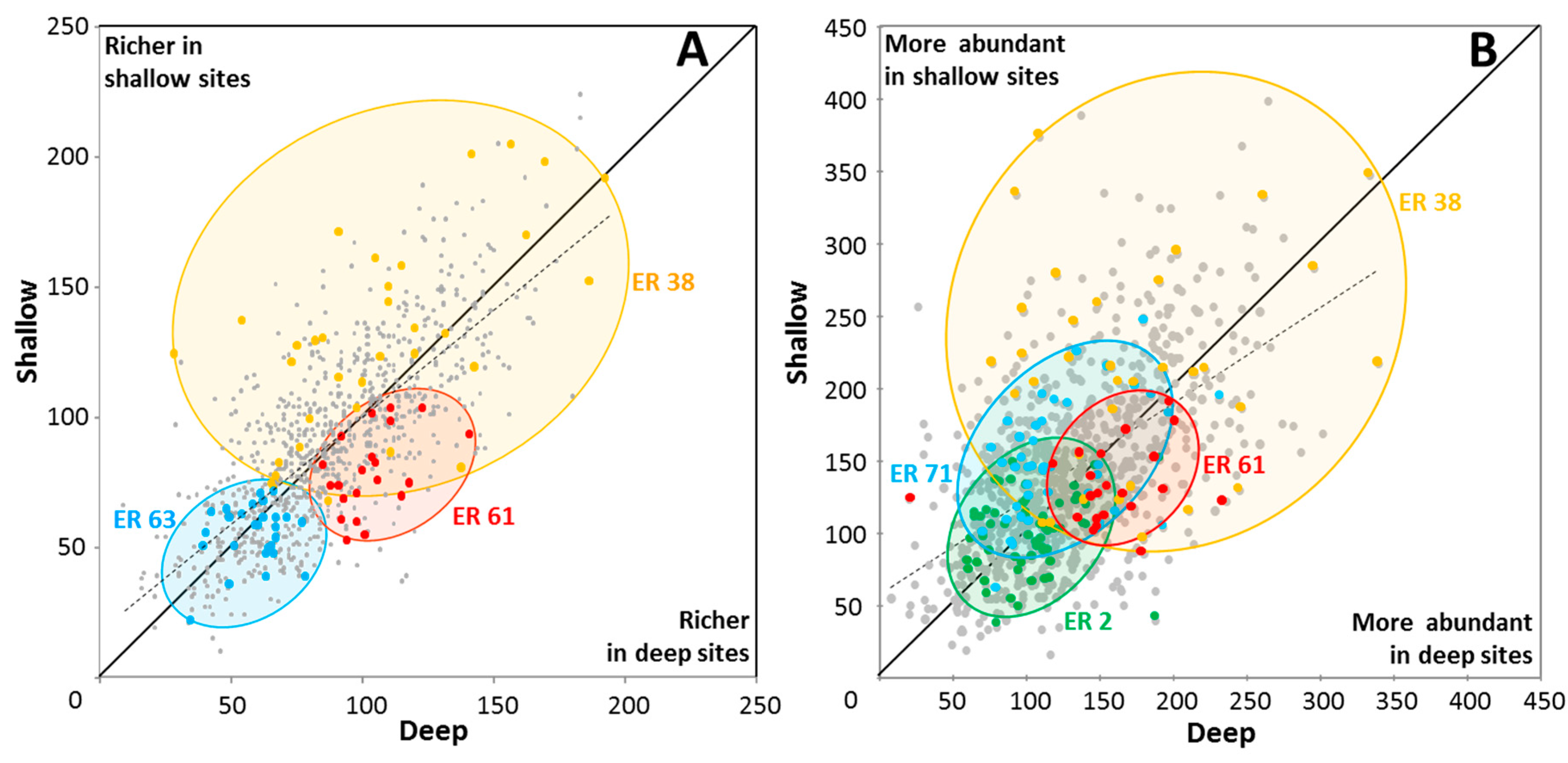
3.3. Marginal ERs
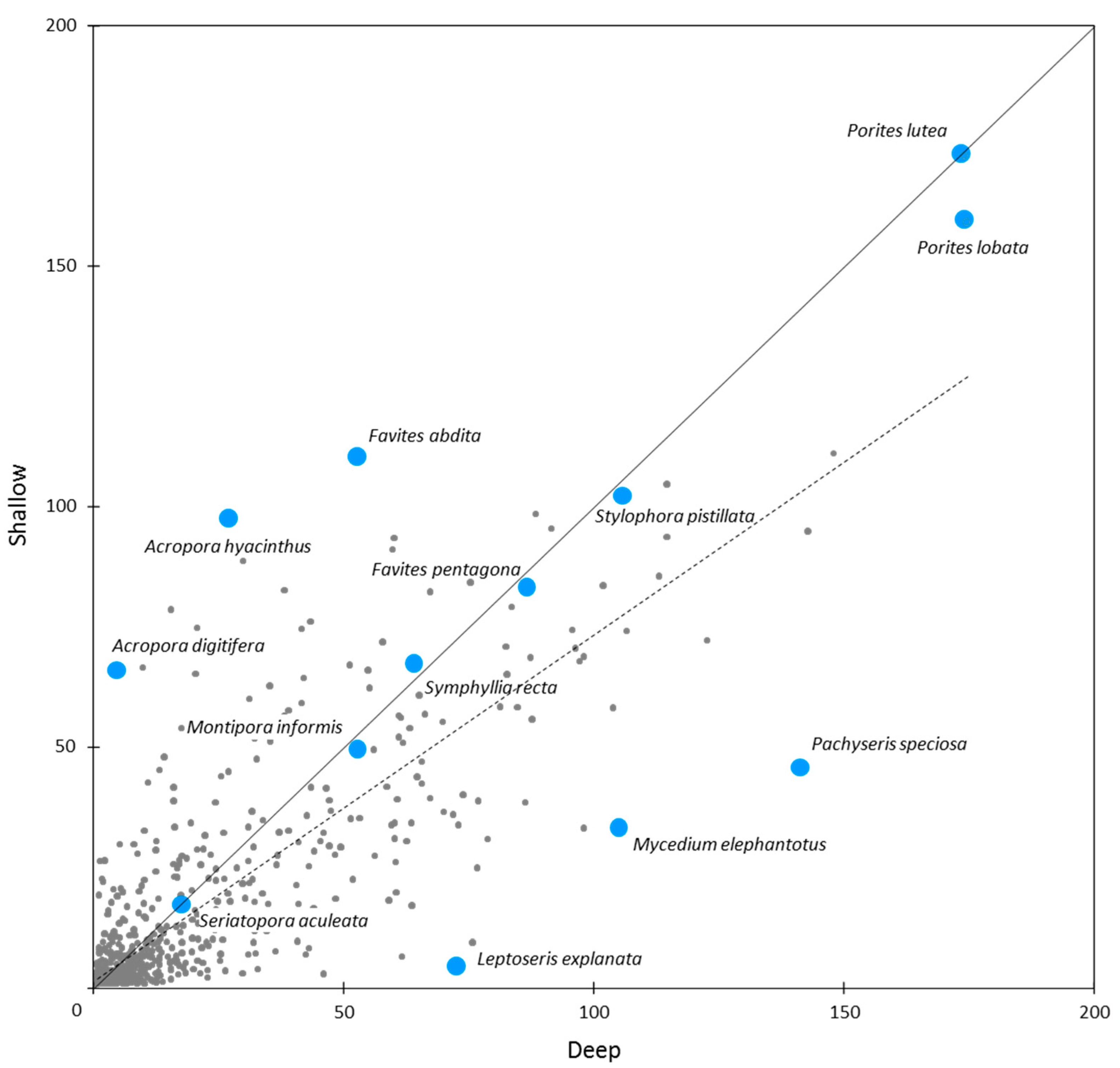
3.4. Effects of Depth on Richness and Abundance
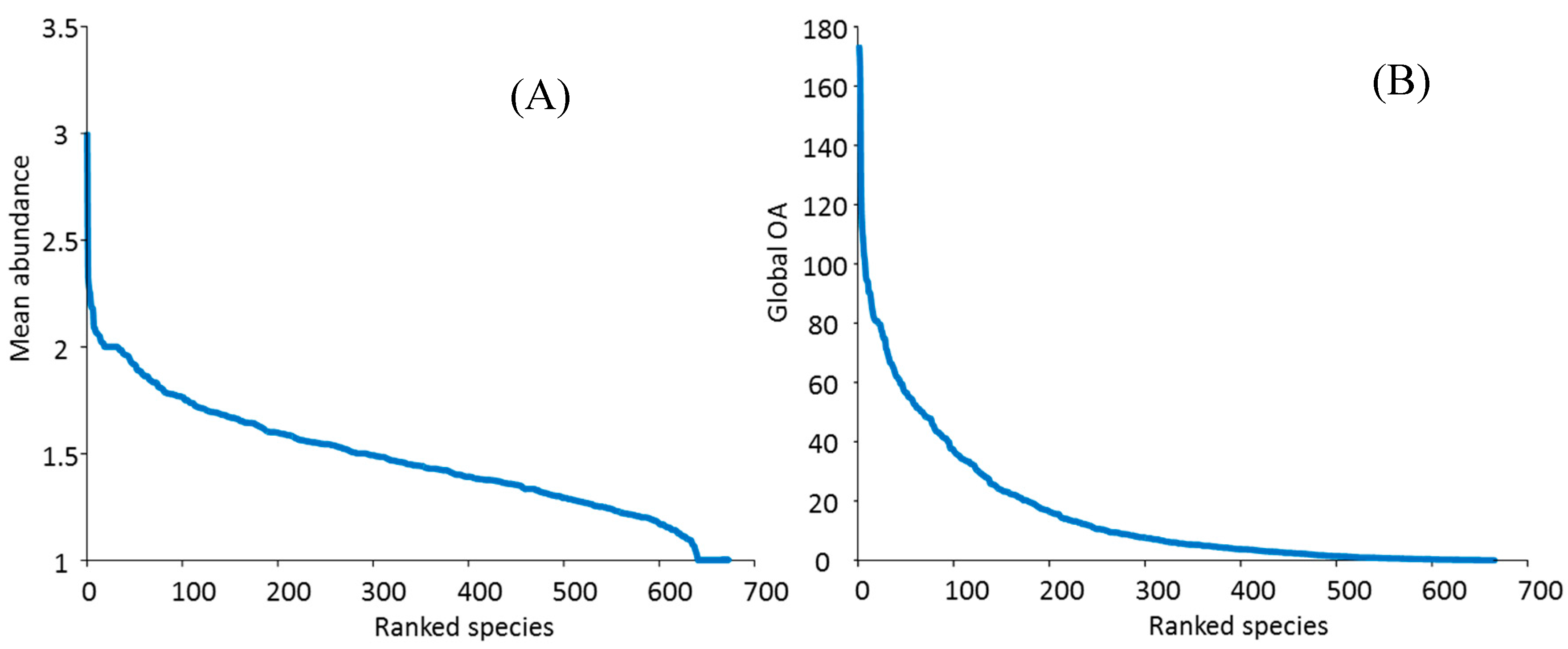
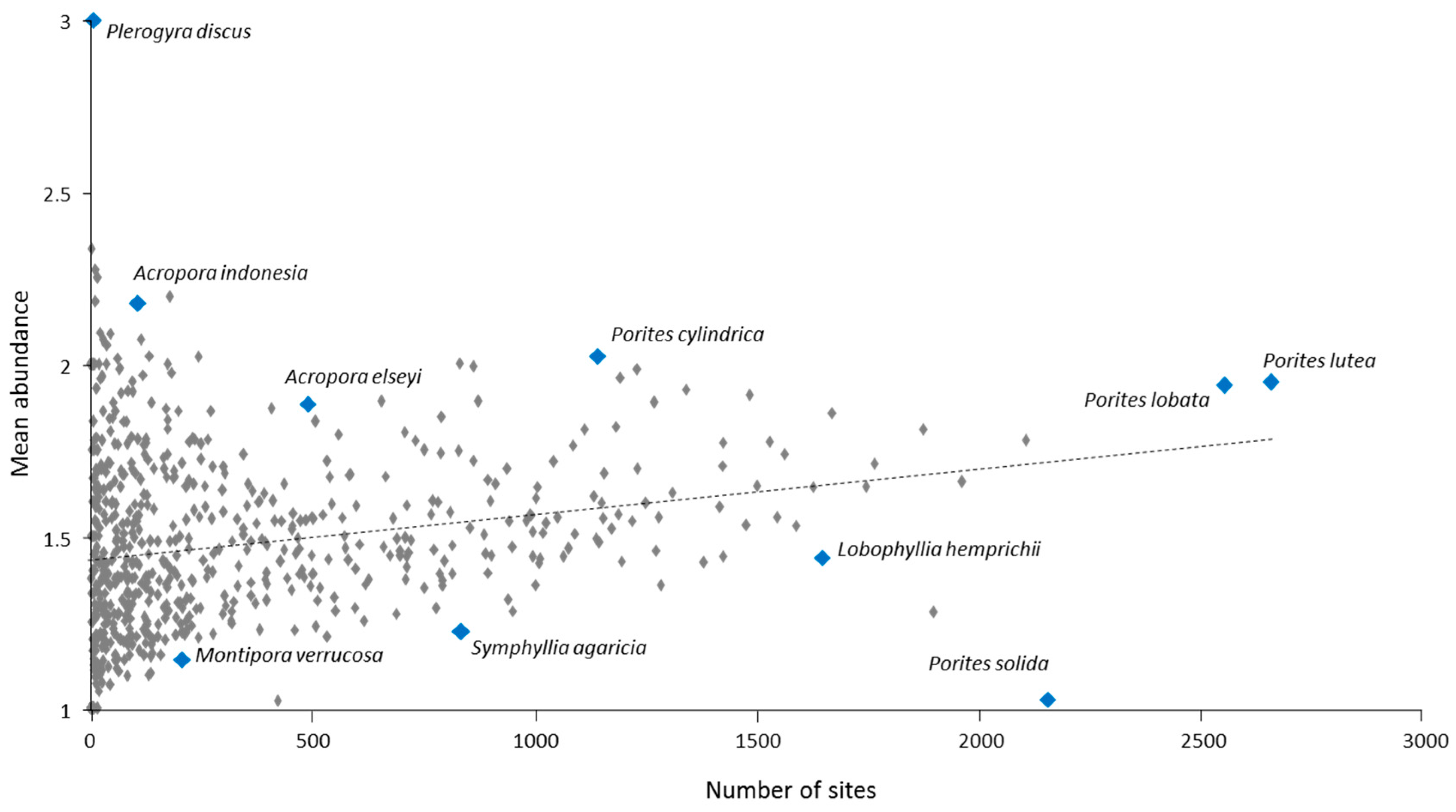
3.5 Relative Abundance
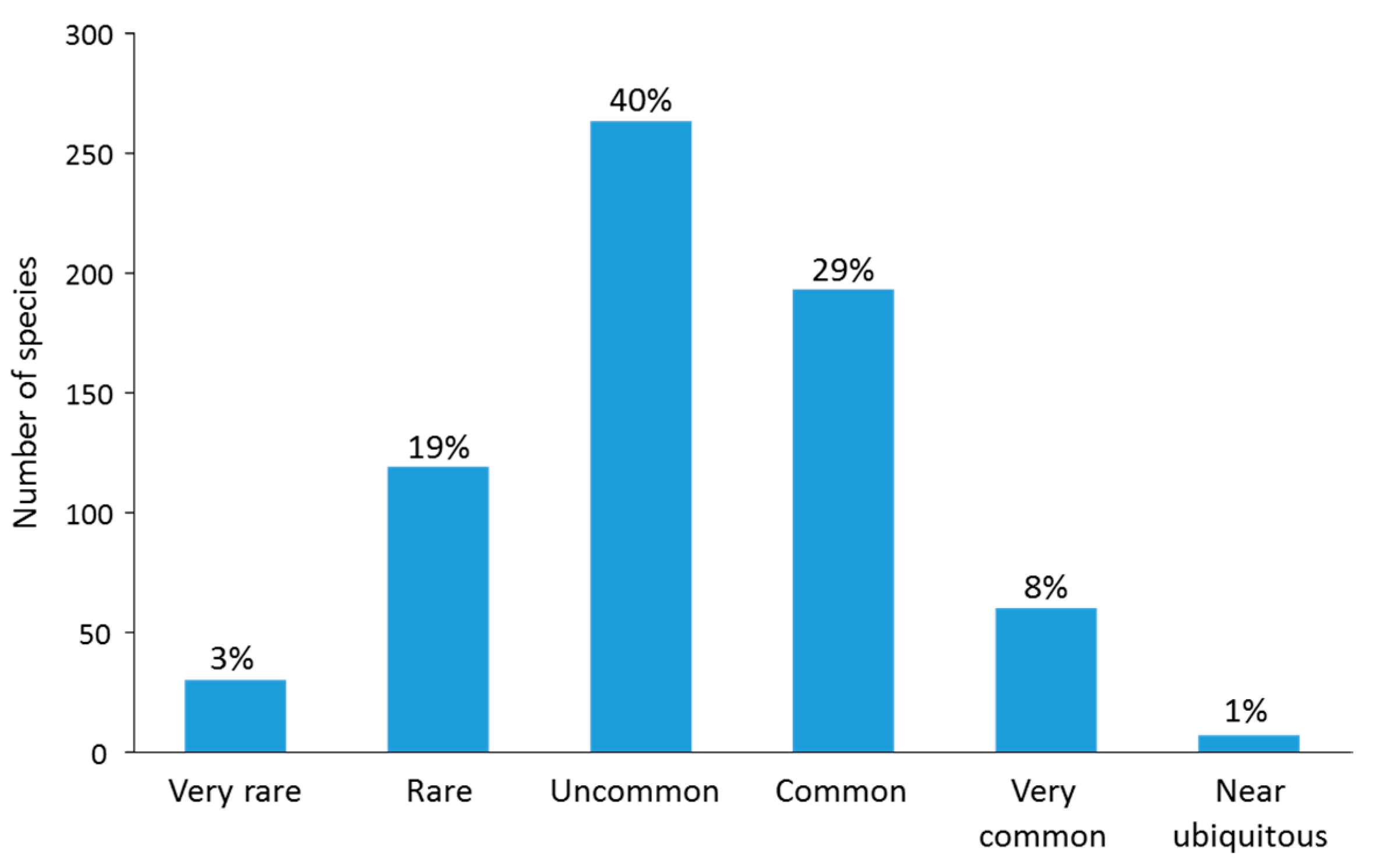
3.6. Rare Taxa
3.7. Centres of Abundance
4. Discussion
4.1. Species Richness
4.2. Relative Abundance
4.3. Different Forms of Rarity
4.4. Centres of Abundance
4.5. Refugia
4.6. Species Vulnerability
- our surveys, or indeed any work to date, have not sampled the entire ranges of many species, not even the main distribution ranges of some, with the most accurate and up-to-date compilation of such data being shown in the distribution maps of “Corals of the World” [1].
- most corals have the potential for widespread dispersal as planktonic larvae or via rafting [75] on the decadal time scales relevant to assessing extinction risk, both within and among ecoregions.
- some species prefer habitats not well sampled, by us or others, including the mesophotic zone (e.g., Craterastrea levis) or deep sandy areas (e.g., Heterocyathus aequicostatus and Heteropsammia cochlea), or live cryptically under boulders on shallow reef flats (e.g., Stylaraea punctata), limiting assessment of their true abundance.
4.7. Extinction Risk
Supplementary Materials
Acknowledgments
Author contributions
Conflicts of Interest
References
- Veron, J.E.N.; Stafford-Smith, M.G.; Turak, E.; DeVantier, L.M. Corals of the World; Version 0.01 Beta. Available online: http://www.coralsoftheworld.org (accessed on 11 April 2017).
- Veron, J.E.N. Corals of the World; Australian Institute of Marine Science: Townsville, Australia, 2000; Volume 3, p. 1410.
- Veron, J.E.N.; Stafford-Smith, M.G.; DeVantier, L.M.; Turak, E. Overview of distribution patterns of zooxanthellate Scleractinia. Front. Mar. Sci. 2015, 1, 81. [Google Scholar] [CrossRef]
- Obura, D.O. An Indian Ocean centre of origin revisited: Palaeogene and Neogene influences defining a biogeographic realm. J. Biogeogr. 2015, 43, 229–242. [Google Scholar] [CrossRef]
- Ginsburg, R.N.; Smith, F.G.W. Proceedings of the colloquium on global aspects of coral reefs: Health, hazards, and history, June 1993. Oceanogr. Lit. Rev. 1995, 42, 673. [Google Scholar]
- Jackson, J.B.C. Reefs since Columbus. Coral Reefs 1997, 16, S23–S32. [Google Scholar] [CrossRef]
- Birkeland, C.; Randall, R.S.; Amesbury, S.S. Coral and Coral-Fish Assessment of the Fagatele Bay National Marine Sanctuary; Report to the National Oceanographic and Atmospheric Administration; US Department of Commerce: Washington, DC, USA, 1994; p. 126.
- Done, T.J.; Turak, E.; Wakeford, M.; DeVantier, L.M.; McDonald, A.; Fisk, D. Decadal changes in turbid-water coral communities at Pandora Reef: Loss of resilience or too soon to tell? Coral Reefs 2007, 26, 789–805. [Google Scholar] [CrossRef]
- Loya, Y. Changes in a Red Sea coral community structure: A long-term case history study. In The Earth in Transition: Patterns and Processes of Biotic Impoverishment; Woodwell, G.M., Ed.; Cambridge University Press: Cambridge, UK, 1990; pp. 369–384. [Google Scholar]
- Connell, J.H.; Hughes, T.P.; Wallace, C.C. A 30-year study of coral abundance, recruitment, and disturbance at several scales in space and time. Ecol. Monogr. 1997, 67, 461–488. [Google Scholar] [CrossRef]
- Global Coral Reef Monitoring Network. Available online: http://www.icriforum.org/gcrmn (accessed on 13 June 2017).
- Ninio, R.; Meekan, M. Spatial patterns in benthic communities and the dynamics of a mosaic ecosystem on the Great Barrier Reef, Australia. Coral Reefs 2002, 21, 95–104. [Google Scholar] [CrossRef]
- De’ath, G.; Fabricius, K.E.; Sweatman, H.; Puotinen, M. The 27–year decline of coral cover on the Great Barrier Reef and its causes. Proc. Natl. Acad. Sci. USA 2012, 109, 17995–17999. [Google Scholar] [CrossRef] [PubMed]
- Gardner, T.A.; Côté, I.M.; Gill, J.A.; Grant, A.; Watkinson, A.R. Long-term region-wide declines in Caribbean corals. Science 2003, 301, 958–960. [Google Scholar] [CrossRef] [PubMed]
- Bruno, J.F.; Selig, E.R. Regional decline of coral cover in the Indo-Pacific: Timing, extent, and subregional comparisons. PLoS ONE 2007, 2, e711. [Google Scholar] [CrossRef] [PubMed]
- Hoegh-Guldberg, O.; Mumby, P.J.; Hooten, A.J.; Steneck, R.S.; Greenfield, P.; Gomez, E.; Harvell, C.D.; Sale, P.F.; Edwards, A.J.; Caldeira, K.; et al. Coral reefs under rapid climate change and ocean acidification. Science 2007, 318, 1737–1742. [Google Scholar] [CrossRef] [PubMed]
- McClanahan, T.; Ateweberhan, M.; Graham, N.; Wilson, S.; Sebastian, C.; Guillaume, M.M.; Bruggemann, J. Western Indian Ocean coral communities: Bleaching responses and susceptibility to extinction. Mar. Ecol. Prog. Ser. 2007, 337, 1–13. [Google Scholar] [CrossRef]
- Veron, J.E.N. A Reef in Time: The Great Barrier Reef from Beginning to End; Harvard University Press: Cambridge, MA, USA, 2008; p. 289. [Google Scholar]
- Carpenter, K.E.; Abrar, M.; Aeby, G.; Aronson, R.B.; Banks, S.; Bruckner, A.; Chiriboga, A.; Cortés, J.; Delbeek, J.C.; DeVantier, L.; et al. One-third of reef-building corals face elevated extinction risk from climate change and local impacts. Science 2008, 321, 560–563. [Google Scholar] [CrossRef] [PubMed]
- Van Woesik, R.; Franklin, E.C.; O’Leary, J.; McClanahan, T.R.; Klaus, J.S.; Budd, A.F. Hosts of the Plio-Pleistocene past reflect modern-day coral vulnerability. Proc. R. Soc. B Biol. Sci. 2012, 279, 2448–2456. [Google Scholar] [CrossRef] [PubMed]
- Huang, D. Threatened reef corals of the world. PLoS ONE 2012, 7, e34459. [Google Scholar] [CrossRef] [PubMed]
- Foden, W.B.; Butchart, S.H.M.; Stuart, S.N.; Vié, J.-C.; Akçakaya, H.R.; Angulo, A.; DeVantier, L.M.; Gutsche, A.; Turak, E.; Cao, L.; et al. Identifying the world’s most climate change vulnerable species: A systematic trait-based assessment of all birds, amphibians and corals. PLoS ONE 2013, 8, e65427. [Google Scholar] [CrossRef] [PubMed]
- Richards, Z.; Beger, M. Regional conservation status of scleractinian coral biodiversity in the Republic of the Marshall Islands. Diversity 2013, 5, 522–540. [Google Scholar] [CrossRef]
- Done, T.J. Patterns in the distribution of coral communities across the central Great Barrier Reef. Coral Reefs 1982, 1, 95–107. [Google Scholar] [CrossRef]
- Veron, J.E.N. Corals of Australia and the Indo-Pacific; Hawaii University Press: Honolulu, HI, USA, 1986; p. 642. [Google Scholar]
- DeVantier, L.M.; De’Ath, G.; Done, T.J.; Turak, E. Ecological assessment of a complex natural system: A case study from the Great Barrier Reef. Ecol. Appl. 1998, 8, 480–496. [Google Scholar] [CrossRef]
- DeVantier, L.; De’ath, G.; Klaus, R.; Al-Moghrabi, S.; Abdulaziz, M.; Reinicke, G.B.; Cheung, C. Reef-building corals and coral communities of the Socotra Archipelago, a zoogeographic ‘crossroads’ in the Arabian Sea. Fauna Arab. 2004, 20, 117–168. [Google Scholar]
- DeVantier, L.M.; De’ath, G.; Turak, E.; Done, T.J.; Fabricius, K. Species richness and community structure of reef-building corals on the nearshore Great Barrier Reef. Coral Reefs 2006, 25, 329–340. [Google Scholar] [CrossRef]
- DeVantier, L.M.; Turak, E.; Skelton, P. Ecological Assessment of the Coral Communities of Bunaken National Park: Indicators of Management Effectiveness. In Proceedings of the 10th International Coral Reef Symposium, Naha, Okinawa, Japan, 28 June–2 July 2004; Suzuki, Y., Nakamori, T., Hidaka, M., Kayanne, H., Casareto, B.E., Nadaoka, K., Yamano, H., Tsuchiya, M., Eds.; Japanese Coral Reef Society: Okinawa, Japan, 2005; pp. 175–187. [Google Scholar]
- Karlson, R.H.; Cornell, H.V.; Hughes, T.P. Coral communities are regionally enriched along an oceanic biodiversity gradient. Nature 2004, 429, 867–870. [Google Scholar] [CrossRef] [PubMed]
- Turak, E.; Brodie, J.; DeVantier, L. Reef-building corals and coral communities of the Yemen Red Sea. Fauna Arab. 2007, 23, 1–40. [Google Scholar]
- Cornell, H.V.; Karlson, R.H.; Hughes, T.P. Local-regional species richness relationships are linear at very small to large scales in west-central Pacific corals. Coral Reefs 2008, 27, 145–151. [Google Scholar] [CrossRef]
- Richards, Z.T.; van Oppen, M.J.H.; Wallace, C.C.; Willis, B.L.; Miller, D.J. Some rare Indo-Pacific coral species are probable hybrids. PLoS ONE 2008, 3, e3240. [Google Scholar] [CrossRef] [PubMed]
- Done, T.J.; DeVantier, L.M.; Turak, E.; Fisk, D.A.; Wakeford, M.; van Woesik, R. Coral growth on three reefs: Development of recovery benchmarks using a space for time approach. Coral Reefs 2010, 29, 815–833. [Google Scholar] [CrossRef]
- Fenner, D.P. Reef Corals of the Eastern Lagoon (Touho-Ponerihouen) of Grande-Terre, New Caledonia; Conservation International: Arlington, VA, USA, 2013; pp. 141–160. [Google Scholar]
- Jackson, J.; Donovan, M.; Cramer, K.; Lam, V. Status and Trends of Caribbean Coral Reefs; Global Coral Reef Monitoring Network, IUCN: Gland, Switzerland, 2014; p. 304. [Google Scholar]
- Veron, J.E.N.; Devantier, L.M.; Turak, E.; Green, A.L.; Kininmonth, S.; Stafford-Smith, M.; Peterson, N. Delineating the coral triangle. Galaxea 2009, 11, 91–100. [Google Scholar] [CrossRef]
- Briggs, J.C. Coral reefs: Conserving the evolutionary sources. Biol. Conserv. 2005, 126, 297–305. [Google Scholar] [CrossRef]
- Allen, G.R. Conservation hotspots of biodiversity and endemism for Indo-Pacific coral reef fishes. Aquat. Conserv. Mar. Freshw. Ecosyst. 2008, 18, 541–556. [Google Scholar] [CrossRef]
- Hoeksema, B.W. Delineation of the Indo-Malayan centre of maximum marine biodiversity: The Coral Triangle. In Biogeography, Time, and Place: Distributions, Barriers, and Islands; Renema, W., Ed.; Springer: Dodrecht, The Netherlands, 2007; Volume 29, pp. 117–178. [Google Scholar]
- Wilkinson, C.; Linden, O.; Cesar, H.; Hodgson, G.; Rubens, J.; Strong, A.E. Ecological and socioeconomic impacts of 1998 coral mortality in the Indian Ocean: An ENSO impact and a warning of future change? Ambio 1999, 28, 188–196. [Google Scholar]
- Veron, J.E.N.; Pichon, M. Scleractinia of Eastern Australia. Part I. Families Thamnasteriidae, Astrocoeniidae, Pocilloporidae; Australian Institute of Marine Science: Townsville, Australia, 1976; Volume 1, pp. 1–86.
- Veron, J.E.N.; Pichon, M.; Wijsman-Best, M. Scleractinia of Eastern Australia. Part II. Families Faviidae, Trachyphylliidae; Australian Institute of Marine Science: Townsville, Australia, 1977; Volume 3, pp. 1–233.
- Veron, J.E.N.; Pichon, M. Scleractinia of Eastern Australia. Part III. Families Agariciidae, Siderastreidae, Fungiidae, Oculinidae, Merulinidae, Mussidae, Pectiniidae, Caryophylliidae, Dendrophylliidae; Australian Institute of Marine Science: Townsville, Australia, 1980; Volume 4, pp. 1–422.
- Veron, J.E.N.; Pichon, M. Scleractinia of Eastern Australia. Part IV. Family Poritidae; Australian Institute of Marine Science: Townsville, Australia, 1982; Volume 5, pp. 1–159.
- Veron, J.E.N.; Wallace, C. Scleractinia of Eastern Australia. Part V. Family Acroporidae; Australian Institute of Marine Science: Townsville, Australia, 1984; Volume 6, pp. 1–485.
- Best, M.B.; Hoeksema, B.W.; Moka, W.; Moll, H.; Sutarna, I.N. Recent scleractinian coral species collected during the Snellius-II expedition in Eastern Indonesia. Neth. J. Sea Res. 1989, 23, 107–115. [Google Scholar] [CrossRef]
- Hoeksema, B.W. Taxonomy, phylogeny and biogeography of mushroom corals (Scleractinia: Fungiidae). Zool. Verh. 1989, 254, 1–295. [Google Scholar]
- Sheppard, C.R.; Sheppard, A.L. Corals and coral communities of Arabia. Fauna Arab. 1991, 12, 1–170. [Google Scholar]
- Wallace, C.C.; Wolstenholme, J. Revision of the coral genus Acropora (Scleractinia: Astrocoeniina: Acroporidae) in Indonesia. Zool. J. Linn. Soc. 1998, 123, 199–384. [Google Scholar] [CrossRef]
- Wallace, C.C. Staghorn Corals of the World. A Revision of the Genus Acropora; CSIRO Publishing: Collingwood, Australia, 1999; p. 422. [Google Scholar]
- Veron, J.E.N.; Stafford-Smith, M.G. Coral ID; Australian Institute of Marine Science: Townsville, Australia, 2002.
- Benzoni, F.; Stefani, F.; Pichon, M.; Galli, P. The name game: Morpho-molecular species boundaries in the genus Psammocora (Cnidaria, Scleractinia). Zool. J. Linn. Soc. 2010, 160, 421–456. [Google Scholar] [CrossRef]
- Turak, E.; DeVantier, L. Field Guide to Reef-Building Corals of Brunei Darussalam; Ministry of Industry and Primary Resources: Bandar Seri Begawan, Brunei Darussalam, 2011; p. 256.
- Huang, D.; Benzoni, F.; Arrigoni, R.; Baird, A.H.; Berumen, M.L.; Bouwmeester, J.; Chou, L.M.; Fukami, H.; Licuanan, W.Y.; Lovell, E.R. Towards a phylogenetic classification of reef corals: The Indo-Pacific genera Merulina, Goniastrea and Scapophyllia (Scleractinia, Merulinidae). Zool. Scr. 2014, 43, 531–548. [Google Scholar] [CrossRef]
- Van der Maarel, E. Transformation of cover-abundance values in phytosociology and its effects on community similarity. In Data-Processing in Phytosociology: Report on the Activities of the Working-Group for Data-Processing in Phytosociology of the International Society for Vegetation Science, 1969–1978; van der Maarel, E., Orlóci, L., Pignatti, S., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 1980; pp. 133–150. [Google Scholar]
- Barkman, J.J.; Doing, H.; Segal, S. Kritische Bemerkungen und Vorschläge zur Quantitativen Vegetationsanalyse. Acta Bot. Neerlandica 1964, 13, 394–419. [Google Scholar] [CrossRef]
- Jongman, R.H.G.; ter Braak, C.J.F.; van Tongereren, O.F.R. Data Analysis in Community and Landscape Ecology; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Wallace, C.C.; Turak, E.; DeVantier, L. Novel characters in a conservative coral genus: Three new species of Astreopora (Scleractinia: Acroporidae) from West Papua. J. Nat. Hist. 2011, 45, 1905–1924. [Google Scholar] [CrossRef]
- Turak, E.; DeVantier, L.M.; Erdmann, M. Euphyllia baliensis sp. nov. (Cnidaria: Anthozoa: Scleractinia: Euphyllidae): A new species of reef coral from Indonesia. Zootaxa 2012, 3422, 52–61. [Google Scholar]
- Fukami, H.; Chen, C.A.; Budd, A.F.; Collins, A.; Wallace, C.; Chuang, Y.-Y.; Chen, C.; Dai, C.-F.; Iwao, K.; Sheppard, C. Mitochondrial and nuclear genes suggest that stony corals are monophyletic but most families of stony corals are not (Order Scleractinia, Class Anthozoa, Phylum Cnidaria). PLoS ONE 2008, 3, e3222. [Google Scholar] [CrossRef] [PubMed]
- Benzoni, F.; Stefani, F.; Stolarski, J.; Pichon, M.; Mitta, G.; Galli, P. Debating phylogenetic relationships of the scleractinian Psammocora: Molecular and morphological evidences. Contrib. Zool. 2007, 76, 35–54. [Google Scholar]
- Schmidt-Roach, S.; Lundgren, P.; Miller, K.J.; Gerlach, G.; Noreen, A.M.; Andreakis, N. Assessing hidden species diversity in the coral Pocillopora damicornis from eastern Australia. Coral Reefs 2013, 32, 161–172. [Google Scholar] [CrossRef]
- Benzoni, F.; Arrigoni, R.; Stefani, F.; Reijnen, B.T.; Montano, S. Phylogenetic position and taxonomy of Cycloseris explanulata and C. wellsi (Scleractinia: Fungiidae): Lost mushroom corals find their way home. Contrib. Zool. 2012, 81, 125–146. [Google Scholar]
- Budd, A.F.; Fukami, H.; Smith, N.D.; Knowlton, N. Taxonomic classification of the reef coral family Mussidae (Cnidaria: Anthozoa: Scleractinia). Zool. J. Linn. Soc. 2012, 166, 465–529. [Google Scholar] [CrossRef]
- Veron, J.E.N. Overview of the taxonomy of zooxanthellate Scleractinia. Zool. J. Linn. Soc. 2013, 169, 485–508. [Google Scholar] [CrossRef] [PubMed]
- Potts, D.C.; Done, T.J.; Isdale, P.J.; Fisk, D.A. Dominance of a coral community by the genus Porites (Scleractinia). Mar. Ecol. Prog Ser. 1985, 23, 79–84. [Google Scholar] [CrossRef]
- Colwell, R.K. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples. Available online: http://purl.oclc.org/estimates (accessed on 4 May 2017).
- DeVantier, L.; Williamson, D.; Willan, R. Coastal Marine Biodiversity of the Sunshine Coast, South-East Queensland: Inventory of Corals, Fishes and Molluscs July 2010; Baseline Survey Report to the Noosa Integrated Catchment Association: Tewantin, Australia, 2010; pp. 1–111. [Google Scholar]
- IUCN. The IUCN Red List of Threatened Species. Available online: http://www.iucnredlist.org/technical-documents/categories-and-criteria/2001-categories-criteria (accessed on 12 April 2017).
- Head, S.M. An undescribed species of Merulina and a new genus and species of siderastreid coral from the Red Sea. J. Nat. Hist. 1983, 17, 419–435. [Google Scholar] [CrossRef]
- Benzoni, F.; Arrigoni, R.; Stefani, F.; Stolarski, J. Systematics of the coral genus Craterastrea (Cnidaria, Anthozoa, Scleractinia) and description of a new family through combined morphological and molecular analyses. Syst. Biodiver. 2012, 10, 417–433. [Google Scholar] [CrossRef]
- Connolly, S.R.; Hughes, T.P.; Bellwood, D.R.; Karlson, R.H. Community structure of corals and reef fishes at multiple scales. Science 2005, 309, 1363–1365. [Google Scholar] [CrossRef] [PubMed]
- Evans, S.M.; McKenna, C.; Simpson, S.D.; Tournois, J.; Genner, M.J. Patterns of species range evolution in Indo-Pacific reef assemblages reveal the Coral Triangle as a net source of transoceanic diversity. Biol. Lett. 2016, 12. [Google Scholar] [CrossRef] [PubMed]
- Harrison, P.; Wallace, C. Reproduction, dispersal and recruitment of scleractinian corals. Ecosyst. World 1990, 25, 133–207. [Google Scholar]
- Babcock, R.C. Comparative demography of three species of scleractinian corals using age- and size-dependent classifications. Ecol. Monogr. 1991, 61, 225–244. [Google Scholar] [CrossRef]
- LaJeunesse, T.; Bhagooli, R.; Hidaka, M.; DeVantier, L.M.; Done, T.J.; Schmidt, G.W.; Fitt, W.K.; Hoegh-Guldberg, O. Closely related Symbiodinium spp. differ in relative dominance in coral reef host communities across environmental, latitudinal and biogeographic gradients. Mar. Ecol. Progr. Ser. 2004, 284, 147–161. [Google Scholar] [CrossRef]
- Anthony, K.R.N.; Fabricius, K.E. Shifting roles of heterotrophy and autotrophy in coral energetics under varying turbidity. J. Exp. Mar. Biol. Ecol. 2000, 252, 221–253. [Google Scholar] [CrossRef]
- Bridge, T.C.L.; Fabricius, K.E.; Bongaerts, P.; Wallace, C.C.; Muir, P.R.; Done, T.J.; Webster, J.M. Diversity of Scleractinia and Octocorallia in the mesophotic zone of the Great Barrier Reef, Australia. Coral Reefs 2012, 31, 179–189. [Google Scholar] [CrossRef]
- Renema, W.; Bellwood, D.R.; Braga, J.C.; Bromfield, K.; Hall, R.; Johnson, K.G.; Lunt, P.; Meyer, C.P.; McMonagle, L.B.; Morley, R.J.; et al. Hopping hotspots: Global shifts in marine biodiversity. Science 2008, 321, 654–657. [Google Scholar] [CrossRef] [PubMed]
- Aronson, R.B.; Precht, W.F.; Macintyre, I.G. Extrinsic control of species replacement on a Holocene reef in Belize: The role of coral disease. Coral Reefs 1998, 17, 223–230. [Google Scholar] [CrossRef]
- Cacciapaglia, C.; van Woesik, R. Reef-coral refugia in a rapidly changing ocean. Glob. Chang. Biol. 2015, 21, 2272–2282. [Google Scholar] [CrossRef] [PubMed]
- Van Hooidonk, R.; Maynard, J.A.; Planes, S. Temporary refugia for coral reefs in a warming world. Nat. Clim. Chang. 2013, 3, 508–511. [Google Scholar] [CrossRef]
- Kleypas, J.A. Modeled estimates of global reef habitat and carbonate production since the Last Glacial Maximum. Paleoceanography 1997, 12, 533–545. [Google Scholar] [CrossRef]
- Muir, P.R.; Wallace, C.C.; Done, T.J.; Aguirre, J.D. Limited scope for latitudinal extension of reef corals. Science 2015, 348, 1135–1138. [Google Scholar] [CrossRef] [PubMed]
- Van Oppen, M.J.H.; Bongaerts, P.I.M.; Underwood, J.N.; Peplow, L.M.; Cooper, T.F. The role of deep reefs in shallow reef recovery: An assessment of vertical connectivity in a brooding coral from west and east Australia. Mol. Ecol. 2011, 20, 1647–1660. [Google Scholar] [CrossRef] [PubMed]
- Hubbell, S.P. A unified theory of biogeography and relative species abundance and its application to tropical rain forests and coral reefs. Coral Reefs 1997, 16, S9–S21. [Google Scholar] [CrossRef]
- Rosindell, J.; Hubbell, S.P.; Etienne, R.S. The Unified Neutral Theory of Biodiversity and Biogeography at Age Ten. Trends Ecol. Evol. 2011, 26, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Brainard, R.E.; Birkeland, C.; Eakin, C.M.; McElhany, P.; Miller, M.W.; Patterson, M.; Piniak, G.A. Status Review Report of 82 Candidate Coral Species Petitioned Under the U.S. Endangered Species Act; NOAA Technical Memorandum, NOAA-TM-NMFS-PIFSC-27; National Oceanic and Atmospheric Administration: Washington, DC, USA, 2011; pp. 530, A-1.
- Guinotte, J.M.; Buddemeier, R.W.; Kleypas, J.A. Future coral reef habitat marginality: Temporal and spatial effects of climate change in the Pacific basin. Coral Reefs 2003, 22, 551–558. [Google Scholar] [CrossRef]
- Birkeland, C.; Miller, M.W.; Piniak, G.A.; Eakin, C.M.; Weijerman, M.; McElhany, P.; Dunlap, M.; Brainard, R.E. Safety in numbers? Abundance may not safeguard corals from increasing carbon dioxide. BioScience 2013, 63, 967–974. [Google Scholar]
- Maynard, J.; van Hooidonk, R.; Eakin, C.M.; Puotinen, M.; Garren, M.; Williams, G.; Heron, S.F.; Lamb, J.; Weil, E.; Willis, B.; et al. Projections of climate conditions that increase coral disease susceptibility and pathogen abundance and virulence. Nat. Clim. Chang. 2015, 5, 688–694. [Google Scholar] [CrossRef]
- Kleypas, J.A.; Buddemeier, R.W.; Gattuso, J.-P. The future of coral reefs in an age of global change. Int. J. Earth Sci. 2001, 90, 426–437. [Google Scholar] [CrossRef]
- Brodie, J.; Waterhouse, J. A critical review of environmental management of the ‘not so Great’ Barrier Reef. Estuar. Coast. Shelf Sci. 2012, 104–105, 1–22. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ER Name, Number | Main Survey Locations (and Countries) | Survey Year(s) | No. of Sites | Species Richness: ER Total Number of Spp.; Site Mean (SD) and Range Across Sites |
|---|---|---|---|---|
| Red Sea north-central, ER 2 | Mainland coast and islands from near Jeddah north to Jordan border (Saudi Arabia) | 1997–1999 | 145 | 240; 60 (15) |
| 22–98 | ||||
| Red Sea south, ER 3 | Mainland coast and islands from near Saudi Arabian border south to Bab Al Mandeb (Yemen) | 1994–1995 | 43 | 166; 34 (17) |
| 3–67 | ||||
| Socotra Archipelago, ER 7 | Socotra, Abd al Kuri, Samha and Darsa islands and islets (Yemen) | 1997–1999 | 93 | 254; 44 (18) |
| 8–94 | ||||
| Gulf of Aden, ER 140 | Mainland coast and islands of Bir Ali and Balhaf area and Sikha Isl. (Yemen) | 1998 | 13 | 98; 36 (8) |
| 28–51 | ||||
| Lakshadweep, ER 20 | Kadmat and Amini Islands, Arabian Sea (India) | 2000 | 14 | 119; 36 (18) |
| 0–66 | ||||
| Madagascar north, ER 15 | Nosy Be area, from Nosy Iranja north to Nosy Hao (Madagascar) | 2003 | 36 | 292; 90 (23) |
| 33–125 | ||||
| Andaman Sea, ER 26 | Phuket area, mainland coast and islands from near Palong and Koh Phi Phi north to Similan and Koh Surin (Thailand) | 2005 | 54 | 290; 65 (18) |
| 15–104 | ||||
| Vietnam south, ER 48 | Nha Trang Bay, Nui Chua National Park, Con Dao islands (Vietnam) | 2003–2005 | 97 | 381; 92 (21) |
| 40–148 | ||||
| Sunda Shelf, ER 145 | Patch reefs and coastal islands (Brunei Darussalam), Anambas island group (Indonesia) | 2008–2009, 2012, 2014 | 103 | 467; 115 (29) |
| 21–187 | ||||
| Hong Kong, ER 51 | Hong Kong islands and mainland (China) | 2001 | 240 | 78; 20 (12) |
| 3–45 | ||||
| Philippines north, ER 45 | Batangas Bay – Verde Passage, Luzon (Philippines) | 2007 | 16 | 348; 127 (23) |
| 80–156 | ||||
| Pohnpei and Kosrae, Micronesia, ER 65 | Pohnpei island, Ant and Pakin Atolls (Federated States of Micronesia) | 2005 | 71 | 320; 56 (15) |
| 16–85 | ||||
| Yap Islands, Micronesia, ER 63 | Ngulu and Ulithi Atolls, Yap (Federated States of Micronesia) | 2007 | 54 | 204; 57 (12) |
| 21–78 | ||||
| Palau, ER 61 | North Reef, Velasco and Kayangel Atolls (Palau) | 2009 | 51 | 333; 89 (22) |
| 10–141 | ||||
| Sulu Sea, ER 43 | Derawan, East Kalimantan, including Berau Delta area, from Panjang area south to Kakaban Isl., Melalungan and Muaras Atolls and Seliman Bay (Indonesia), El Nido area and north Palawan mainland and islands (Philippines) | 2004, 2009 | 149 | 505; 123 (28) |
| 43–183 | ||||
| Lesser Sunda Islands and Savu Sea, ER 33 | Bali and Nusa Penida, west Lombok, Komodo, West Timor and Roti (Indonesia), west and north coasts and Atauro Isl. (Timor Leste) | 1995, 2006, 2008, 2011–2012, 2016 | 248 | 490; 75 (32) |
| 2–164 | ||||
| Celebes Sea, ER 42 | Bunaken islands and North Sulawesi coast, Bangka Isl., Lembeh Strait, and Sangihe-Talaud islands including Tahulandang, Siao, Sangihe, Karakelong and the Nanusa group (Indonesia) | 2000, 2004–2005 | 160 | 475; 79 (29) |
| 24–171 | ||||
| Banda Sea and Moluccas, ER 36 | Banda islands including Banda Neira, Pulau Run, Ai and Hatta, and Wakatobi islands including Pulau Wangiwangi and Tomea, Koro Maha and Kaledupa Atolls (Indonesia) | 2003–2004 | 86 | 416; 78 (15) |
| 47–133 | ||||
| Halmahera, ER 41 | Halmahera west, north and east coasts from Ternate to Buli Bay and Morotai (Indonesia) | 1997, 2008 | 89 | 472; 102 (46) |
| 30–224 | ||||
| Raja Ampat, Papua, ER 40 | All main islands and offshore reefs, including Misool, Kofiau, Batanta, Gag, Wayag, Kawe and Waigeo Isl., Papua (Indonesia) | 2002 | 94 | 478; 87 (26) |
| 19–165 | ||||
| Papua south-west coast, ER 38 | Fak Fak – Kaimana and Triton Bay coasts and islands, Papua (Indonesia) | 2006 | 68 | 473; 117 (40) |
| 28–205 | ||||
| Cenderawasih Bay, Papua, ER 39 | Biak, Numfoor and Yapen islands and inner bay (Indonesia) | 2006 | 66 | 475; 122 (32) |
| 70–215 | ||||
| Bismarck Sea, New Guinea, ER 69 | New Britain and New Ireland, including Kimbe and Stettin Bays, Manus Isl., Tigak area and New Hanover Isl. (Papua New Guinea) | 2002, 2004, 2006 | 155 | 465; 95 (24) |
| 41–150 | ||||
| Milne Bay, Papua New Guinea, ER 71 | Mullins Harbour area, Goodenough and Collingwood Bays, d’Entrecasteaux and Amphlett Island groups, Fergusson and Normanby Isl. (Papua New Guinea) | 2000, 2007 | 85 | 432; 89 (21) |
| 43–150 | ||||
| Solomon Islands and Bougainville, ER 70 | Main island groups including Florida, Isabel, Choiseul, Shortlands, New Georgia, Russell, Guadalcanal and Makira Isl. (Solomon Islands) | 2005 | 114 | 442; 91 (20) |
| 42–143 | ||||
| Great Barrier Reef far north and Torres Strait, ER 73 | Princess Charlotte Bay, nearshore islands and reefs (Australia) | 2001, 2011 | 32 | 329; 89 (23) |
| 37–133 | ||||
| Great Barrier Reef north-central, ER 74 | Lizard Island area south to Whitsunday Islands, including nearshore, mid- and outer continental shelf reefs (Australia) | 1994–1997, 2001–2002, 2011 | 582 | 369; 46 (25) |
| 0–129 | ||||
| Great Barrier Reef south-east, Pompey and Swain Reefs, ER 76 | Pompey reefs (Australia) | 2002 | 3 | 148; 92 (10) |
| 83–102 | ||||
| Great Barrier Reef south, ER 77 | Percy Islands south to Keppel Islands, including nearshore and mid continental shelf reefs (Australia) | 1995, 2002 | 43 | 216; 40 (18) |
| 6–84 | ||||
| Moreton Bay, east Australia, ER 78 | Hervey Bay and Sunshine Coast (Australia) | 2008-10 | 25 | 114; 36 (14) |
| 16–59 | ||||
| Fiji, ER 101 | Naitauba Isl., Taveuni Isl., Viti Levu Isl. (Fiji) | 2010, 2012 | 46 | 344; 75 (28) |
| 30–136 |
| Issue | Our Approach |
|---|---|
| Field survey method: estimating local and regional richness | All methods have their advantages and limitations. Ours is based on a long-established approach to rapid ecological assessment, pioneered by terrestrial botanists and adapted to coral reefs [24]. It does not deliver quantitative estimates of species’ abundance, but does provide robust “semi-quantitative” estimates. It is more effective than traditional quantitative methods in estimating local and regional species richness, facilitating the active searching for species at each site, rather than being restricted to a defined quadrat area or transect line. At site level, the present method has regularly returned a two- to three-fold increase in coral species records in comparison with line transects conducted concurrently (e.g., Red Sea, Great Barrier Reef and Vietnam, L.M. DeVantier unpublished; Socotra Islands [27]). |
| Taxonomic framework | Despite recent advances stabilizing coral taxonomy (e.g., taxonomic works cited above), substantial taxonomic uncertainty remains, as demonstrated by various recently published classifications and synonymies [2,53,55,61,62,64,65,66]. This has arisen, in part, because of differences between traditional morphology-based approaches and those employing molecular genetics. These nomenclatural issues are relevant mainly at taxonomic levels higher than species, although "name games” continue, and the placement of certain species within particular genera remains contentious. We have, as far as practicable, included species accepted in the major taxonomic revisions that preceded or were published over the period of this study (i.e., up to 2016), some minor specific nomenclatural differences notwithstanding. With some modification, our framework follows that of Veron et al. [3], as included in the open access website [1]. |
| Species identification and inter-observer variability | Our dataset has been compiled by experienced surveyors, mainly Emre Turak and Lyndon DeVantier, working together or independently, with assistance in particular ERs from Terry Done, Mary Wakeford and Denise McCorry. Use of standard reference works, museum collections, extensive underwater photography and limited collection of specimens has minimized variability as far as practicable. Juvenile corals < 5 cm diameter, particularly in the speciose genera Acropora, Montipora and Porites, pose particular problems, and unless identity was certain, these were not recorded in our dataset. Having adopted a cautious approach to the issue of inter-observer variability, we have not tried to incorporate similar datasets from other researchers in the present study, although we have referred to them for comparison (Table S4). |
| Discrepancies between published ER species lists and abundance data | The abundance data were compiled during our site surveys, and as far as practicable subsequently updated from photography and specimens. Published records for certain species in particular ERs were also based on non-site observations, and hence may not be supported by our site abundance data. |
| Estimating rarity | Because of the taxonomic and survey difficulties noted above (see Table S1), we consider that our abundance estimates for a few species are unreliable. That is, their true abundance may be higher than documented here because of field identification issues and occurrence in cryptic locations or habitats poorly surveyed. |
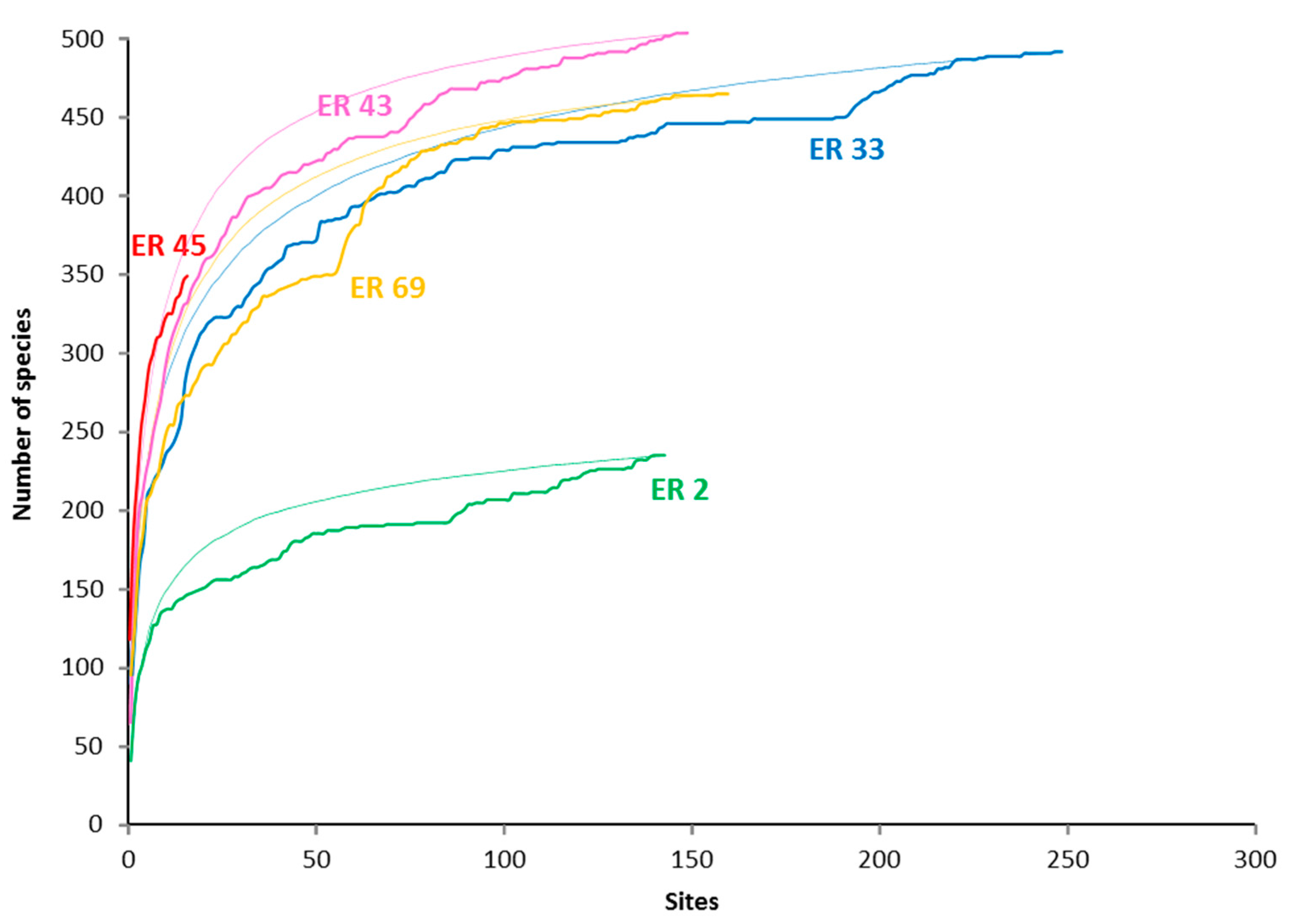
| Sampling effort among ecoregions | With the notable exceptions of North Philippines (16 sites), Lakshadweep (14 sites), Gulf of Aden (13 sites) and the Pompey and Swain Reefs (3 sites) of Australia’s Great Barrier Reef (GBR), sample sizes within ecoregions, although not standardized, were generally sufficient to approach asymptotic species richness (Figure 2). Our dataset is strongly focused on two of the main "centres of diversity” of reef corals, the CT and Arabia, and two nearby regions, notably eastern Australia and the central-western Pacific. Our dataset does not include two regions of major species’ attenuation, be they latitudinal (e.g., Ryukyu Islands—Japan, Western Australia) or longitudinal (e.g., Central – Far Eastern Pacific). These host regional endemics not surveyed in the present study. |
| Ecoregion boundaries | Boundaries were based on the best information available, both published and unpublished [3], but have different levels of support. Our dataset can, however, be interrogated at much smaller geographic scales, an undertaking largely beyond the scope of the present analysis. This is a major objective for future development of the "Corals of the World” website [1]. |
| Distribution ranges | Despite major recent advances in coral biogeography, range boundaries remain "fuzzy”, disjunctions are poorly understood, and ranges are better documented for some species than others. Recently described species typically have highly restricted distributions initially, often only the type locality. Ranges may, or may not, be extended with further work. |
| Sampling effort across habitats | Our surveys were focused on reef slopes, crests and outer-mid reef flats, and typically limited to water depths of less than 50 m or to the base of the reef slope if shallower. We were unable to survey inner reef flats in water depths < 0.5 m. |
| True and false absences | We do not assume that a species missing from our inventories in a particular ER is actually absent there, even though our surveys covered a range of habitats. |
| Estimating individual species abundance ranks for massive Porites | Except where specimens were collected, we were unable to identify the large colony-forming massive Porites to species-level in the field. The approach taken here (Table 3) provides derived estimates at species level, the best that can be achieved from our dataset. |
| Taxon | Site Abundance Score | ||||
|---|---|---|---|---|---|
| Massive Porites | 1 | 2 | 3 | 4 | 5 |
| Porites lobata | 1 | 2 | 2 | 3 | 4 |
| Porites lutea | 1 | 2 | 2 | 3 | 4 |
| Porites australiensis | 0 | 1 | 2 | 2 | 3 |
| Porites myrmidonensis | 0 | 0 | 2 | 3 | 3 |
| Porites solida | 0 | 1 | 1 | 1 | 2 |
| Porites mayeri | 0 | 0 | 1 | 1 | 1 |
| Deep Sites | OA Score | IUCN Red List | Shallow Sites | OA Score | IUCN Red List |
|---|---|---|---|---|---|
| Acanthophyllia deshayesiana *,1 | 0.1 | NT | Acropora bushyensis | 0.23 | LC |
| Acropora fragilis 2 | 0.07 | DD | Acropora pruinosa | 1.27 | DD |
| Acropora natalensis * | 0.13 | DD | Acropora forskali * | 0.26 | DD |
| Acropora pichoni | 0.75 | NT | Acropora rufus * | 0.1 | DD |
| Acropora rongelapensis | 0.26 | DD | Acropora schmitti | 0.03 | DD |
| Acropora roseni | 0.03 | En (A4ce) | Acropora sirikitiae | 0.13 | NA |
| Acropora rudis | 0.16 | En (A4ce) | Acropora spathulata | 2.83 | LC |
| Acropora russelli | 0.52 | Vu (A4c) | Acropora torresiana | 0.62 | DD |
| Acropora simplex | 1.4 | Vu (A4ce) | Acropora tumida | 2.5 | DD |
| Acropora stoddarti | 0.07 | DD | Astreopora cenderawasih | 0.13 | NA |
| Acropora tenella | 0.72 | Vu (A4ce) | Echinopora irregularis | 0.03 | DD |
| Craterastrea levis | 0.16 | NA | Leptoseris kalayaanensis | 0.03 | NA |
| Echinophyllia tarae * | 0.03 | NA | Micromussa diminuta | 0.03 | DD |
| Echinopora robusta | 0.03 | Vu (A4c) | Montipora aspergillus | 0.03 | DD |
| Echinopora tiranensis | 0.03 | DD | Montipora echinata | 0.07 | DD |
| Euphyllia baliensis | 0.07 | NA | Montipora gaimardi | 0.52 | Vu (A4c) |
| Horastrea indica 5 | 0.13 | Vu (A4c) | Montipora pachytuberculata | 0.07 | DD |
| Isopora togianensis | 0.13 | En (A4ce) | Montipora saudii | 0.16 | NT |
| Leptoseris amitoriensis | 0.13 | NT | Pocillopora capitata *,5 | 0.23 | LC |
| Lithophyllon lobata 3 | 0.46 | En (A4c; B2ab (ii,iii)) | Pocillopora fungiformis | 0.39 | En (A4c) |
| Montipora orientalis | 0.03 | Vu (A4c) | Polycyathus chaishanensis * | 0.13 | NA |
| Nemenzophyllia turbida | 0.42 | Vu (A4cd) | Porites cocosensis | 0.16 | Vu (A4cde) |
| Parasimplastrea omanensis 4 | 0.39 | En (A4c) | Porites cumulatus | 1.6 | Vu (A4cde) |
| Plerogyra discus | 0.1 | Vu (A4c) | Porites eridani | 0.7 | En (A4cde) |
| Stylocoeniella cocosensis | 0.1 | Vu (A4c) | Porites ornata * | 0.7 | En (A4cde) |
| Coral Species | Overall Abundance Score | Mean OA Score Across ERs (SD) | Percent of 31 ERs | IUCN Red List |
|---|---|---|---|---|
| Porites lutea | 173.7 | 178.0 (44.3) | 100 | LC |
| Porites lobata | 165.1 | 179.2 (40.9) | 97 | LC |
| Galaxea fascicularis | 125.0 | 141.1 (51.4) | 94 | NT |
| Goniastrea pectinata | 113.3 | 134.3 (58.6) | 97 | LC |
| Platygyra daedalea | 108.6 | 129.5 (49.5) | 97 | LC |
| Stylophora pistillata | 103.5 | 123.0 (54.2) | 94 | NT |
| Cyphastrea serailia | 100.8 | 106.5 (52.9) | 100 | LC |
| Favia favus | 95.8 | 105.4 (42.3) | 100 | LC |
| Pocillopora verrucosa | 94.7 | 111.0 (48.1) | 90 | LC |
| Pocillopora damicornis | 94.3 | 111.0 (44.5) | 97 | LC |
| Fungia fungites | 90.6 | 101.9 (38.7) | 90 | NT |
| Pavona varians | 90.6 | 119.6 (57.3) | 90 | LC |
| Favites abdita | 89.1 | 92.7 (49.0) | 94 | NT |
| Seriatopora hystrix | 86.2 | 97.7 (59.1) | 84 | LC |
| Favites pentagona | 84.2 | 92.9 (51.2) | 100 | LC |
| Favia matthaii | 82.4 | 94.7 (51.8) | 97 | NT |
| Isopora palifera | 81.5 | 95.4 (38.8) | 87 | NT |
| Porites australiensis | 81.0 | 103.8 (30.9) | 84 | LC |
| Favia pallida | 81.0 | 85.8 (44.9) | 100 | LC |
| Pachyseris speciosa | 80.9 | 99.9 (47.2) | 84 | LC |
| Astreopora myriophthalma | 80.2 | 99.2 (42.9) | 94 | LC |
| Acropora muricata * | 79.9 | 87.8 (38.0) | 94 | NT |
| Lobophyllia hemprichii | 79.9 | 96.2 (34.9) | 94 | LC |
| Porites rus | 78.0 | 101.8 (54.2) | 84 | LC |
| Porites cylindrica | 77.2 | 95.4 (36.2) | 74 | NT |
| Hydnophora exesa | 75.5 | 85.6 (43.6) | 100 | NT |
| Merulina ampliata | 75.0 | 103.0 (40.3) | 81 | LC |
| Porites solida | 75.0 | 90.7 (34.3) | 97 | LC |
| Acropora hyacinthus | 71.7 | 76.0 (53.9) | 94 | NT |
| Echinopora lamellosa | 71.1 | 80.6 (37.4) | 90 | LC |
| Species of Pectinia | Occurrence (% of sites) | Mean Abundance | Overall Abundance | IWP OA Category | IUCN Red List |
|---|---|---|---|---|---|
| Pectinia africanus | 0.7 | 1.3 | 0.9 | Rare | Vu (A4c) |
| Pectinia alcicornis | 16.6 | 1.6 | 25.9 | Common | Vu (A4c) |
| Pectinia ayleni | 3.1 | 1.4 | 4.4 | Uncommon | NT |
| Pectinia elongata | 1.0 | 1.6 | 1.6 | Uncommon | NT |
| Pectinia lactuca | 31.7 | 1.5 | 46.5 | Common | Vu (A4cd) |
| Pectinia maxima | 3.9 | 1.5 | 5.7 | Uncommon | En (A4cd) |
| Pectinia paeonia | 15.7 | 1.5 | 23.0 | Common | NT |
| Pectinia pygmaeus | 0.5 | 1.8 | 0.8 | Rare | NT |
| Pectinia teres | 4.2 | 1.8 | 7.5 | Uncommon | NT |
| Species | O (% of sites) | Mean Abn. | OA | Form of Rarity | IUCN Red List Classification | Climate Change Vulnerability | ||
|---|---|---|---|---|---|---|---|---|
| Se | Ua | Ex | ||||||
| Acropora bushyensis | 0.1 | 2.3 | 0.23 | Diffusive | LC | H | L | H |
| Acropora fragilis | 0.07 | 1 | 0.07 | Diffusive | DD (as A. rambleri) | H | L | L |
| Acropora roseni | 0.03 | 1 | 0.03 | Geo-suffusive | En (A4ce) | H | H | H |
| Acropora rudis | 0.13 | 1.25 | 0.16 | Geo-suffusive | En (A4ce) | H | H | L |
| Acropora sirikitiae | 0.07 | 2 | 0.13 | Geo-suffusive | NA | NA | ||
| Acropora teres | 0.03 | 1.5 | 0.1 | Diffusive | DD | H | L | L |
| Alveopora viridis | 0.13 | 1 | 0.13 | Diffusive | NT | H | H | H |
| Anacropora pillai | 0.13 | 1.5 | 0.2 | Diffusive | DD | H | L | H |
| Astreopora cenderawasih | 0.07 | 2 | 0.13 | Geo-suffusive | NA | NA | ||
| Astreopora scabra | 0.13 | 1.25 | 0.16 | Diffusive | LC | H | H | L |
| Craterastrea levis | 0.16 | 1 | 0.16 | Habitat-suffusive | NA | NA | ||
| Duncanopsammia axifuga | 0.16 | 1.2 | 0.2 | Diffusive | NT | H | L | H |
| Echinopora ashmorensis | 0.16 | 1.4 | 0.23 | Diffusive | Vu (A4c) | H | L | H |
| Echinopora irregularis | 0.03 | 1 | 0.03 | Geo-suffusive | DD | H | H | U |
| Echinopora robusta | 0.03 | 1 | 0.03 | Geo-suffusive | Vu (A4c) | H | L | L |
| Echinopora tiranensis | 0.03 | 1 | 0.03 | Geo-suffusive | DD | H | H | U |
| Euphyllia baliensis | 0.03 | 2 | 0.07 | Geo- and habitat-suffusive | NA | NA | ||
| Fungia puishani | 0.16 | 1.6 | 0.26 | Geo-suffusive | DD | H | H | L |
| Goniopora cellulosa * | 0.07 | 1 | 0.07 | Geo-suffusive | Vu (A4c) | H | H | H |
| Goniopora polyformis | 0.13 | 1.5 | 0.2 | Diffusive | Vu (A4c) | H | L | H |
| Goniopora sultani | 0.13 | 1 | 0.13 | Geo-suffusive | LC | H | H | U |
| Horastrea indica * | 0.1 | 1.33 | 0.13 | Geo-suffusive | Vu (A4c) | H | H | L |
| Isopora togianensis | 0.07 | 2 | 0.13 | Geo-suffusive | En (A4ce) | H | L | H |
| Leptoseris amitoriensis | 0.13 | 1 | 0.13 | Diffusive | NT | H | L | H |
| Leptoseris kalayaanensis | 0.03 | 1 | 0.03 | Geo-suffusive | NA | NA | ||
| Micromussa diminuta | 0.03 | 1 | 0.03 | Diffusive | DD | H | H | H |
| Montipora aspergillus | 0.03 | 1 | 0.03 | Geo-suffusive | DD | H | H | U |
| Montipora echinata | 0.07 | 1 | 0.07 | Geo-suffusive | DD | H | H | U |
| Montipora orientalis | 0.03 | 1 | 0.03 | Diffusive | Vu (A4c) | H | H | H |
| Montipora pachytuberculata | 0.07 | 1 | 0.07 | Geo-suffusive | DD | H | H | U |
| Montipora saudii | 0.13 | 1.25 | 0.16 | Geo-suffusive | NT | H | H | U |
| Montipora verrilli * | 0.07 | 1 | 0.07 | Diffusive | DD | H | H | L |
| Plerogyra discus | 0.03 | 3 | 0.1 | Habitat-suffusive | Vu (A4c) | H | L | H |
| Pocillopora capitata * | 0.13 | 1.75 | 0.23 | Range extension to Fiji from Eastern Pacific | LC | H | L | L |
| Porites cocosensis | 0.13 | 1.25 | 0.16 | Diffusive | Vu (A4cde) | H | H | H |
| Porites eridani | 0.07 | 1 | 0.07 | Diffusive | En (A4cde) | H | L | H |
| Porites somaliensis * | 0.07 | 1 | 0.07 | Geo-suffusive | NT | H | H | L |
| Sclerophyllia maxima * | 0.13 | 1.5 | 0.2 | Geo-suffusive | NT (as Acanthastrea maxima) | H | H | U |
| Stylocoeniella cocosensis | 0.1 | 1 | 0.1 | Diffusive | Vu (A4c) | H | L | H |
| Stylophora madagascarensis | 0.16 | 1.4 | 0.23 | Geo-suffusive | En (A4c) | H | H | H |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
DeVantier, L.; Turak, E. Species Richness and Relative Abundance of Reef-Building Corals in the Indo-West Pacific. Diversity 2017, 9, 25. https://doi.org/10.3390/d9030025
DeVantier L, Turak E. Species Richness and Relative Abundance of Reef-Building Corals in the Indo-West Pacific. Diversity. 2017; 9(3):25. https://doi.org/10.3390/d9030025
Chicago/Turabian StyleDeVantier, Lyndon, and Emre Turak. 2017. "Species Richness and Relative Abundance of Reef-Building Corals in the Indo-West Pacific" Diversity 9, no. 3: 25. https://doi.org/10.3390/d9030025
APA StyleDeVantier, L., & Turak, E. (2017). Species Richness and Relative Abundance of Reef-Building Corals in the Indo-West Pacific. Diversity, 9(3), 25. https://doi.org/10.3390/d9030025