Quorum Sensing and Expression of Virulence in Pectobacteria

Abstract
: Quorum sensing (QS) is a population density-dependent regulatory mechanism in which gene expression is coupled to the accumulation of a chemical signaling molecule. QS systems are widespread among the plant soft-rotting bacteria. In Pectobacterium carotovorum, at least two QS systems exist being specified by the nature of chemical signals involved. QS in Pectobacterium carotovorum uses N-acylhomoserine lactone (AHL) based, as well as autoinducer-2 (AI-2) dependent signaling systems. This review will address the importance of the QS in production of virulence factors and interaction of QS with other regulatory systems in Pectobacterium carotovorum.1. Introduction
Pectobacterium carotovorum subsp. carotovorum (Pcc) [ex Erwinia carotovora subsp. carotovora] and Pectobacterium atrosepticum (Pba) [ex Erwinia carotovora subsp. atroseptica] are plant pathogens which cause soft rot in a wide range of plant species, including many crops of economic importance such as vegetables, ornamentals and also the model plant Arabidopsis. Pectobacteria are often described as brute-force pathogens because their virulence strategy relies on plant cell-wall degrading enzymes (PCWDE) which disrupt host cell integrity and thus promote rotting [1,2]. Symptom progression depends on aggressiveness of the bacterial strain and susceptibility of the host plant, but also on environmental conditions, among which temperature and humidity are particularly critical [3]. Coordination of virulence factor synthesis is crucial for the pathogenicity. Regulation of the virulence in Pcc is coordinated by quorum sensing (QS) [4,5] through a complex set of transcription factors and posttranscriptional regulators [6–13]. A highly complex manner of regulation of PCWDE production may reflect a demand for very precise regulation of certain genes in response to both environmental stimuli and population density. The finely tuned regulation enables a certain group of bacteria to launch a unified coordinated response to environmental stimuli and to accomplish tasks which would be difficult or even impossible to be achieved by a single bacterium.
Though QS plays a key role in regulation of a battery of PCWDE in pectobacteria, it is important to note that it is only one component of an extremely complicated regulatory network that modulates expression of virulence factors. During the infection, PCWDE are not synthesized until the bacterial population density exceeds a certain number [14]. While the QS model proposes that the cells use signal molecules to sense population density, the diffusion sensing model suggests that signal molecules are used by cells to evaluate mass-transfer properties of the environment [15]. According to the latter hypothesis, pectobacteria may use QS to make sure that gene products (e.g., PCWDE) are produced only then if they will have a beneficial effect for the bacterium. One may envisage that accumulation of signal molecules may enable the cells to discriminate between certain environments such as host plant tissue and soil (plant pathogens) or light organ tissue and sea water Vibrio fischeri (V. fischeri). The PCWDE secreted by pectobacteria are potential activators of the plant response to infection. So, if bacterial population is very small, it would make sense to postpone enzyme production until they are absolutely required [16]. On the other hand, synthesis of PCWDE may also be just a response of pectobacteria to nutrient limitation as the population size increases. In this case, sensing of population increase will trigger transcription of the genes for PCWDE degradation and further growth will be supported by liberation of additional nutrients from the host plant [17]. In this way pathogens might be able to put off expression of PCWDE for as long as possible to prevent PCWDE production until a suitable population density has been achieved at which it can overcome the host defence system.
2. AHL-Based Quorum Sensing in Pectobacteria
2.1. General Aspects of AHL-Based Quorum Sensing
Quorum sensing was first described as a regulator of bioluminescence in V. fischeri and Vibrio harveyi (V. harveyi) [18,19] in which acyl-homoserine lactons (AHLs) act as signaling molecules. Since then, in many Gram-negative bacteria several kinds of AHLs (called autoinducers) have been identified as signal compounds for this mechanism [20,21]. Bacteria that produce AHLs can respond to a local concentration of signaling molecules, and high population densities (1010 cells mL−1) foster accumulation of inducing levels of AHLs. The chain length (C4-C18) and the oxidative status of the acyl side chain of AHLs vary and reflect the observed species-specificity of this communication system [22]. QS allows the bacteria to respond to fluctuation in their numbers and enables a synchronous regulation of target genes when living in a community. Clinically important and commercially relevant pathogens often use QS to control various cellular processes [23]. AHLs are central to the regulation and have been shown to control processes as diverse as bioluminescence in marine bacteria V. fischeri [24], biofilm formation and virulence in an opportunistic pathogen Pseudomonas aeruginosa [25], conjugal transfer of the Ti plasmid in Agrobacterium tumefaciens [26,27], production of the exopolysaccharide stewartan acting as a virulence factor in a maize pathogen Pantoea stewartii [28,29] and production of carbapenem antibiotics as well as PCWDE in a plant pathogen Pectobacterium carotovorum [4,5]. Although the QS systems are used by many plant pathogenic bacteria to regulate the virulence, they are not essential for their survival under laboratory conditions. This indicates that the need for QS is more urgent in the natural habitat, where bacterial population density fluctuates in response to environmental conditions.
The luciferase operon in V. fischeri is regulated by two proteins, LuxI (I-protein), which is responsible for the production of the AHL autoinducer, and LuxR (R-protein), which is activated by this autoinducer to increase the transcription from the luciferase operon [24]. Although the LuxR-type proteins have similar structures, their operating mechanisms can be different. So, in contrast to LuxR, a LuxR-type protein EsaR of the plant-pathogen Pantoea stewartii [30] binds the target DNA in the absence of AHL and represses the transcription, whereas binding of cognate AHL is thought to be needed for the release of EsaR from the DNA to enable derepression of the target gene [31,32].
The LuxI type proteins are AHL synthases that use S-adenosylmethionine (SAM) to synthesize the homoserine lactone ring, and the acyl chains come from lipid metabolism provided by various acyl-carrier proteins [32,33]. Several studies have revealed a conserved region with a number of specific residues required for LuxI activity. This region comprising residues 25–104 of the 193-aa LuxI polypeptide has been proposed to represent the active site for formation of the amide bond between the acyl group and the amino donor, SAM [34,35]. Acyl chains vary in number of carbons from 4 to 18 and are sufficiently diverse to ensure specific recognition of AHLs by different LuxR-type proteins.
The LuxR type proteins are transcription factors, which upon binding the AHL signal molecule modulate transcription of their target genes [36,37]. While most QS systems conform to this pattern, there are examples of the binding of LuxR-like proteins to the DNA in their native state [30]. The LuxR-type proteins share an end-to-end sequence identity of 18–28%. The AHL-interacting region (66–38 aa) and a DNA binding motif (183–229 aa) are defined by two clusters of stronger sequence conservation [38]. Only after the AHL concentration reaches a critical threshold level corresponding to a certain quorum of the bacteria, the AHL interacts with the cognate R-protein. The LuxR-type proteins act as QS regulators. They distinguish between different AHLs by binding only a cognate AHL suggesting their involvement in mainly intraspecies signaling [39]. In different Pectobacterium strains there are examples of LuxR-type proteins which can recognize AHLs with different side chains [40–42]. Many LuxR-type activators, including CarR, LuxR and TraR, the latter being the only crystallized LuxR-type protein so far [37,43,44] form dimers or multimers upon binding the AHLs [42,45]. Originally, the DNA sequence element required for LuxR-AHL binding was called the lux operator, but now mostly the lux box [46]. The lux box is a 20-base pair inverted repeat sequence located upstream of the lux operon transcriptional start site and centered at position −42.5. Additionally, both arms of the dyad repeat are required for activation [38].
2.2. ExpI/ExpR Quorum Sensing in Pectobacterium
Within the Pectobacterium genus, a particular bacterial species, and depending on the strain, normally synthesizes only one or two major AHL signals, sometimes along with some minor AHL products. Pcc and Pba strains have been divided into two classes on the basis of produced AHL. Class I strains, such as Pcc EC153 and SCC3193 synthesize predominantly N-3-oxooctanoyl-l-homoserine lactone (3-oxo-C8-AHL), along with lower amount of 3-oxohexanoyl-l-homoserine lactone (3-oxo-C6-AHL). In contrast, class II strains which include Pcc ATCC39048 (and its derivatives, e.g., GS101, ATTn10, and MS1), Pcc 71, Pcc SCC1 and Pba SCRI1043 produce predominantly 3-oxo-C6-AHL, whereas little or none of 3-oxo-C8-AHL [47]. In these class I and class II strains of Pectobacterium, a single LuxI homologue has been described for each strain being responsible for the production of all AHLs produced by the strain (Table 1). The AHL biosynthetic specificity mainly resides with AHL synthases [47] and availability of the precursors [40,48]. LuxI homologues of different strains have different names. So, the LuxI counterpart in Pcc 3193 and Pba SCRI1043 is called ExpI [4,49], in Pcc ATCC39048 CarI [23], in Pcc EC153 AhlI [48] and in Pcc 71 AhlI [50] or HslI [6]. Protein alignment revealed that AhlI of Pcc 71 (accession no. L40174), ExpI of Pba SCRI1043 (accession no. CAG73025) share 93 to 99% identity, whereas the identity of these proteins with respective homologs from Pcc SCC3193 (accession no. X80457), Pcc EC153 (accession no. DQ093124) and Pba CFBP6272 (accession no. AJ580600) is only 70%.
Brader et al. [40] have studied the specificity of AHL signaling in Pcc and demonstrated the molecular basis for the substrate chain length specificity of the AHL synthase ExpI of Pcc SCC1 and SCC3193. The Pcc SCC1 and SCC3193 produce 3-oxo-C6-AHL and 3-oxo-C8-AHL as their main autoinducers, respectively. The difference in chain length specificity of AHL production is accompanied by a higher sensitivity toward 3-oxo-C8-AHL in Pcc SCC3193 and towards 3-oxo-C6-AHL in Pcc SCC1. The type of AHL produced is dependent on corresponding I-protein [33,50]. So, the expI mutant of SCC3193 which produces no detectable amount of AHLs will produce either 3-oxo-C6-AHL or 3-oxo-C8-AHL after introduction of the plasmids harboring expISCC1 or expISCC3193, respectively. Mutagenesis of the AHL synthase gene expISCC1 identified amino acid replacements M127T and F69L in ExpISCC1 that altered the chain length of the produced autoinducer—a 3-oxo-C6-HSL producer was converted to a 3-oxo-C8-AHL producer. These substitutions increased the size of a putative acyl moiety binding catalytic pocket creating space for the larger C8 side chain. This observation suggests that acyl chains of different length may be accommodated by coordinated sequence differences in and near the binding pocket of the ExpI protein.
Though the pattern of produced AHLs is mainly dependent on the I-protein, it can also be modulated by external milieu [47]. In Pcc, different growth media or growth conditions result either in very complex AHL profile containing more than 10 different AHLs (in case of LB medium) or a very simple one with only few AHLs (in case of M9 minimal medium and potato). It seems that under stringent in planta growth, only few AHLs have a role in determining the virulence as indicated by a poor AHL profile of Pcc in the host plant [40].
In pectobacteria, the AHL-based QS system relies on a transcriptional regulator protein ExpR which belongs to LuxR family of proteins. ExpR detects the AHL signal and transduces it into cellular response. There are similar inverted repeat elements ranging from 18 to 22 bp, generally called exp-type boxes, associated with the promoters of genes regulated by ExpR-type proteins in different Pcc and Pba strains [48]. Chatterjee et al. [48] demonstrated that several members of Pectobacterium subspecies which are otherwise related possess two types of structurally and functionally distinct ExpR proteins. The ExpR proteins of class I strains represented by AY894424 of Pcc strain EC153, X80457 of Pcc SCC3193 and AY 80600 of Pba strain CFBP 6272share over 95% identity with each other. In contrast to that, the ExpR protein (AY894425) of the class II strain Pcc 71 shares only ca 60% of homology with ExpR proteins of class I strains [48]. Major differences between the ExpR proteins of these two groups of strains rely in their N-terminal AHL-binding domains. The ExpR proteins of class I strains mostly interact with 3-oxo-C8-AHL whereas those of the class II strains bind mostly 3-oxo-C6-AHL.
In Pba SCRI1043, in addition to luxR homologue expR (renamed as expR1) found adjacent to its cognate luxI gene, a second luxR homologue which has been termed virR appears to be present in the genome. Sequence analysis revealed the presence of VirR homologue ExpR2 in Pcc strains 71 and SCC3193 [13,52]. Sequence analysis revealed that ExpR2 of Pcc strain 71 exhibits a high level of identity (93%) with VirR of SCRI1043. The ExpR2 of SCC3193 shows 94% aa identity (97% similarity) to VirR of Pba SCRI1043 [13]. Based on genetic and functional homology, it was concluded that VirR is genetically identical or very similar to ExpR2 [52].
Pcc strain ATCC39048 has also a third luxR gene homologue called carR, which is apparently mediating QS control of carbapenem antibiotic production in this strain [53]. Unlike expR1 and virR/expR2, carR functions as an activator which positively regulates carbapenem production in Pcc [53].
2.3. Role of ExpI/ExpR in Production of PCWDE
The regulation of PCWDE production in Pcc and Pba is a well-studied example of AHL-dependent gene expression [4,5]. Inactivation of expI has led to decreased production of PCWDE and decreased virulence. The simplest way for activation of virulence gene expression by ExpR is its direct binding to promoter regions of the genes to be transcribed. The initial studies with ExpR (renamed as ExpR1) of Pcc SCC3193 did not establish that clear role for the AHL receptor protein in PCWDE production. Inactivation on expR (renamed as expR1) had very little effect on extracellular pectinolytic enzyme production and also on AHL synthesis. However, multiple copies of expR1 had some inhibitory effect on PCWDE production [54]. ExpR1-like other members of the LuxR family of regulators have a DNA binding domain and they are believed to function as transcription factors. Cui et al. documented that in Pcc 71 ExpR1 has no direct effect on pel1 (pectate lyase) or ohlI (luxI homologue) transcription [50,55]. Interestingly, studies with Erwinia chrysanthemi (E. chr) ExpI/ExpR system presented a very different picture. Inactivation of expI had only little effect on pectinases synthesis in E. chr as expression of only two of the pectate lyase genes, pelA and pelB was decreased. E. chr expR− mutants still produced AHLs and pectinases. However, gel shift and DNase I footprintig experiments showed that ExpR protein binds specifically to promoter regions of the five major pel genes. The ExpR–DNA band-shift profiles changed in the presence of AHL [56,57]. Based on these results it is clear that operation mode of the QS systems is fundamentally different in these two groups of soft-rotting bacteria.
It has been proposed that there is a connection between QS- and Rsm system-mediated regulation of PCWDE synthesis. The Rsm system involves RsmA, a protein that is able to complex with mRNAs of PCWDE genes to trigger their degradation by RNAses, and a small noncoding rsmB sRNA. The rsmB sRNA regulates gene expression by binding to RsmA protein impeding its activity [6,50,55,58]. Early studies have revealed a negative regulatory role of AHL on rsmA, for example: (1) inactivation of expI causes an increase in the level of rsmA transcript; (2) addition of exogenous AHL to the expI− mutant reduces the level of rsmA RNA to that detected in the parent strain and restores the production of PCWDE [10,11]. These observations suggested that at least part of the regulatory role of QS in Pcc strains 3193 and 71 is mediated by its effect on rsmA expression.
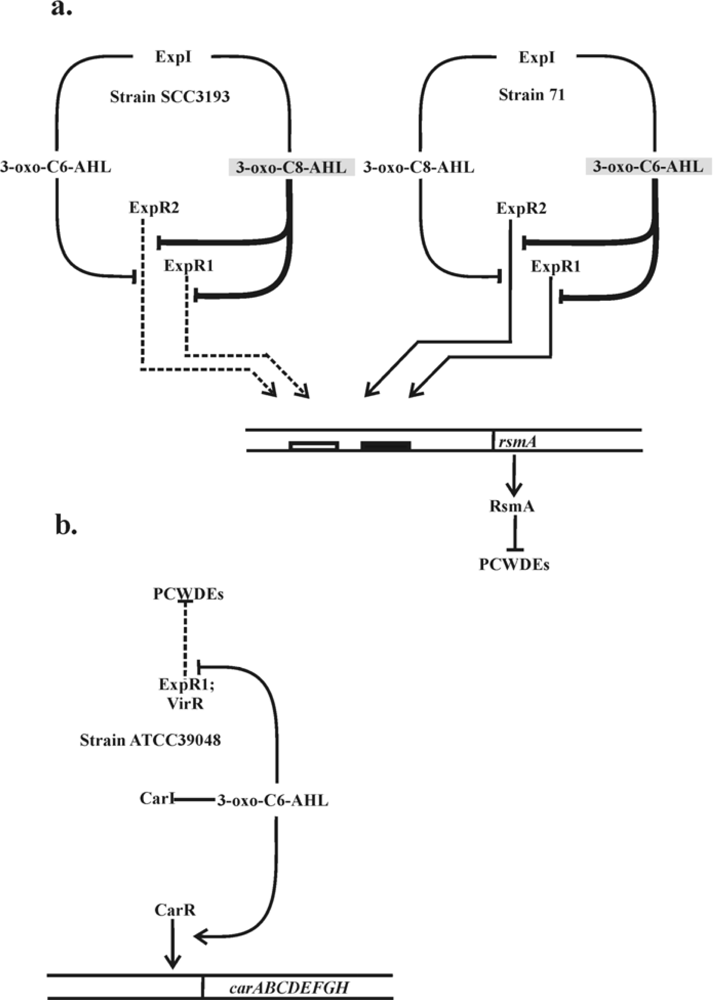
The studies by Chatterjee and associates provided further confirmation for the connection between the QS and RsmA protein through discovery that ExpR1 activates the rsmA transcription [48,50]. These studies demonstrated that the ExpR1-RsmA system is different from a typical LuxI-LuxR regulatory system since ExpR1, but not the ExpR1-AHL complex activates the transcription of rsmA. The ExpR1 protein in Pcc 71 was shown to bind in vitro the rsmA promoter in the absence of AHL and this binding was abolished by the addition of the AHL signal, 3-oxo-C6-AHL (but not 3-oxo-C8-AHL) (Figure 1(a)). In the absence of AHL signals, the receptors ExpR1 and ExpR2 activate the transcription of rsmA that encodes a global regulatory RNA-binding protein which reduces the half-life of PCWDE mRNAs. The action of ExpRs is neutralized by their interaction with AHL. In both strains the ExpR1 has strict specificity for the signal molecule: in strain SCC3193 ExpR1 is released from rsmA only in the presence of 3-oxo-C8-AHL whereas in strain 71 only in the presence of 3-oxo-C6-AHL. The activator function of ExpR2 is neutralized by both signal molecules. The nucleotide sequence of the ExpR1 binding site was similar to lux-box, designated in Pcc as the expR-box [48]. The fact that ExpR1 acts as an activator only in the absence of the AHL signal explains why it was not previously isolated in genetics screens for QS-dependent regulation of PCWDE production. These observations clearly explain why components of the ExpR1/ExpR2 QS system have opposite effects on PCWDE in Pcc.
As a mutation in expR1 gene in the AHL+ background of Pcc 71 had almost no effect on levels of PCWDE and RsmA, and level of PCWDE production in an expR1− ahlI− mutant was higher than that of in expR1+ ahlI− strain, but still lower than in Pcc71, it referred to the possibility that there is more than one functional expR gene in Pcc71. Support for this suggestion came from the analysis of the whole genome sequence of Pba SCRI1043 [49], which disclosed two open reading frames corresponding to AHL binding proteins (putative ExpR homologues).
The data presented by Cui et al. [51] confirmed also the presence of another functional expR homolog, expR2, in the genome of Pcc71. Burr et al. reported the presence of a second LuxR homolog, VirR, which negatively regulates the production of PCWDE and virulence in the absence of AHL in Pba SCRI1043. Based on genetic homology, it was concluded that virR is genetically identical or very similar to expR2 in strain 71. The virR (expR2) gene appears to be present in a number of Pectobacterium strains including Pcc ATCC39048, SCC3193, 71 and Pba SCRI1093 [52]. Comparative analysis of Ecc 71 ExpR2 with previously characterized ExpR1 revealed that they have 62% of identity and 81% of similarity. Both ExpR variants contain autoinducer-binding domain at the N-terminus and helix-turn-helix motifs at the C-terminus [51]. ExpR1 and ExpR2 also differ in their AHL-interaction pattern as ExpR2 only binds 3-oxo-C8-AHL (this can be explained by differences between the AHL binding sites in ExpR1 and ExpR2). Examination of the sequences of ExpR1 and ExpR2 revealed significant differences (46 of 150 residues) between the N-terminal autoinducer-binding domains of these two proteins. This diversity is responsible for significant differences in the binding affinities of ExpR variants for AHL analogs [51].
Gel mobility shift data demonstrated that both, ExpR1 and ExpR2 bind the rsmA promoter region. However, the DNA binding properties of these two ExpR variants are different. The ExpR2 binds poorly to the 20-mer expR-box upstream of the rsmA which is sufficient for the binding of ExpR1. Thus, in addition to the 20-mer expR box, a second 16-mer binding site is necessary for ExpR2-rsmA binding (Figure 1(a)). Sequence analysis revealed that the two ExpR variants both contain motifs for DNA binding at the C-termini of the protein and differ by 3 (Leu196Pro, Gly203Asp, and Thr206Ile). out of 56 amino acid residues. The three-residue difference may explain the requirement for the second binding site of ExpR2 in Pcc 71 [51].
In Pcc 71 ExpI− ExpR2− mutant background reduced level of RsmA and high levels of exoenzymes were detected. This contrasts with almost complete inhibition of exoenzyme production and a high level of RsmA in an ExpI− ExpR1− mutant. These observations establish ExpR2 as the major regulator of exoprotein production and indicate that ExpR1 plays an ancillary role in exoprotein synthesis regulation primarily modulating the level of the RsmA protein [51]. Burr et al. [52] demonstrated that virR mutation restored the PCWDE production and virulence in the AhlI− mutant of SCRI1043; thus, these phenotypes closely resemble those resulting from inactivation of expR2 in Pcc 71.
However, in Pcc SCC3193 ExpR1 and ExpR2 act synergistically to repress the production of PCWDE and thereby also the virulence in the absence of AHL. By inactivating the QS system of SCC3193, that includes the expI, expR1 and expR2 genes, it was shown that a triple mutant was able to produce wild-type levels of PCWDE without the addition of any AHLs or macerated plant tissue similarly or even better than the wild-type. The presence of either ExpR1 or ExpR2 in expI− mutant background essentially abolished the production of PCWDE in the absence of AHLs. Intriguingly, in expI− expR1− double mutant background the transcription of rsmA can be prevented with either 3-oxo-C6-AHL or 3-oxo-C8-AHL, leading to subsequent production of PCWDE. In contrast, expression of rsmA is abolished only by the addition of 3-oxo-C8-AHL in the expI− expR2− mutant. These data show that QS controls rsmA expression and suggest that QS regulation of PCWDE can be mediated by RsmA. Interestingly, addition of the cognate, but not of the non-cognate AHL to the expI− rsmA− double mutant resulted in 30% increase in cellulase activity. So, it cannot rule out the possibility that the ExpR proteins of Pcc SCC3193 have a dual role by functioning both in an AHL-bound and in a ligand-free states. These results suggest that though the QS control of PCWDE production is largely mediated by RsmA, the cognate 3-oxo-C8-AHL can trigger an additional RsmA-independent pathway to fine tune the PCWDE production.
The ability of ExpR2 to respond to several different AHLs raises a question of its role. Sjöblom et al. speculated that ExpR2 may allow interspecies communication between a Pcc strain SCC3193 and neighboring bacteria [13]. On the other hand, variation in environmental and physiological conditions may affect the availability of precursors with different acyl chain length in the acyl-ACP pool which may explain the varied profile of AHLs produced by the cells under different growth conditions. ExpR (ExpR2) with wider substrate specificity may help the bacteria to modulate their virulence factors production according to current situation.
An open question remains whether the ExpR1 and ExpR2 (in SCC3193) bind the same promoter region or several possible ExpR binding sites exist in the rsmA promoter. In the rsmA promoter of Pcc 71 two ExpR sites were recently identified and the binding of ExpR to this sites were verified by DNase I footprint analysis [51]. A similar DNA element was identified in the rsmA promoter of SCC3193 and also in Pba SCRI1043, but binding of either ExpR to this site was not verified.
RsmA plays at least partially a role in inhibition of exoenzyme production at elevated temperatures. The production of PCWDE is reduced when Pcc 71 is cultured at 34.5 °C compared to 28 °C and this trend was correlated with reduction in the concentration of oxo-C6-HSL signal. Decrease of AHL concentration enhanced the rsmA expression. It was observed that levels of the rsmA transcript and the RsmA protein were increased at elevated temperature, suggesting a potential link between the thermoregulation and the Rsm system. The elevated levels of RsmA at 34.5 °C promote RNA decay and consequently reduce the PCWDE production and virulence. One exception is strain EC153 that produces more AHL at 34.5 °C than at 28 °C, and this contrasts with the responses of other Pcc strains, including Ecc71 [59].
The fact that Pcc strains lacking the QS system (expI− expR1− and virR−/expR2− triple mutant) are still able to grow and macerate plant tissue similarly to wild-type cells under laboratory conditions indicates that the QS system is biologically relevant mainly in natural habitat, where densities and the composition of bacterial populations fluctuate in response to environmental cues. An ecological study would be essential to elucidate the significance of QS in success of Pcc in the environment [1,60,61].
2.4. Role of Quorum Sensing in the Production of other Virulence Factors
For a long time only little was known about the extent of QS control within a single microorganism. So, previous studies showed that AHL is required in a Pc subspecies for the expression of PCWDE genes [4,6,54]. To identify new virulence genes in Pba that may be controlled by QS, Pemberton et al. [62] carried out a genome-wide screen using transposon mutagenesis. They isolated seven new genes controlled by QS signal molecule AHL and demonstrated that at least one of these genes, nip, is involved in pathogenesis on potato and can induce necrosis in tobacco.
Corbett et al. [63] used the proteomic technique of two-dimentional difference gel electrophoresis analysis of the wild-type and expI− negative mutant of Pba SCRI1043 to identify QS-dependent proteins in the secretome. This study identified three novel QS-dependent secreted enzymes: Svx, a putative proteoglycan hydrolase and a putative cellulase, in addition to previously known QS-dependent secreted PCWDE. Although such proteomic analysis provides valuable information in identifying and comparing secreted proteins from wild-type pectobacteria and defined mutants, many proteins may be missed because they are poorly soluble, present in very low amount or expressed only in planta.
Analysis of the full genome sequence of Pba SCRI1043 has revealed several other putative virulence factors [49]. Importantly, for many years the role of QS in pathogenesis with regard to PCWDE regulation was studied intensively only in vitro. In 2008, a whole genome microarray approach was used by Liu et al. for an expI− mutant Pba1043 grown in planta to determine global effects of QS on gene regulation during potato infection, with particular emphasis on the relationship between PCWDE production and possible stealth mechanisms [2]. According to transcriptomic analysis by Liu et al., approximately 26% of all Pba genes are QS-regulated, suggesting that QS may more strongly contribute to pathogenesis than previously thought. Given that many important secreted PCWDE are produced by Pectobacterium in a QS-dependent fashion, it is also important to consider how these secreted proteins are moved out of the bacterial cell. Most interestingly, QS in Pba was shown also to modulate type I (T1SS) and type II (T2SS) secretion systems which are considered “accessory virulence factors” responsible for the delivery of PCWDE. T1SS and T2SS genes exhibited reduced transcription in the expI mutant. It was also observed that type III (T3SS) secretion system structural, putative effector, and helper genes are also downregulated in the expI mutant. The finding that QS regulates the entire T3SS indicates that coordinated physical (PCWDE) and stealth (T3SS) attack may be necessary for successful infection and disease development. The QS regulon also included a novel Type VI secretion system (T6SS), its predicted substrates Hcp and VgrG and over 70 known or putative regulators, some of which have been demonstrated to control pathogenicity. The complexity of QS circuit indicates that it is governing multiple other regulators that in turn precisely regulate the destructive arsenal of PCWDE during the infection [2].
2.5. Role of Quorum Sensing in Carbapenem Production
A few strains of Pcc from geographically diverse sources produce low levels of a β-lactam antibiotic, carbapenem [64]. The genes for carbapenem production are clustered in an operon of eight genes (carA-H) [65] and are controlled in a QS manner in response to accumulation of AHL [5,66,67]. The car gene cluster of Pcc ATCC39048 was the first among Pcc strains to be sequenced and both functionally and transcriptionally defined [53]. QS in Pcc ATCC39048 involves the production of the diffusible signaling molecule 3-oxo-C6-AHL, which is synthesized by a LuxI family member CarI (an ExpI homologue). CarR, a member of the LuxR family transcription factors, is a DNA-binding transcriptional activator of carA–H that functions in the presence of AHL. Interestingly, cryptic car gene clusters have been identified in a number of Pectobacterium strains [64]. Antibiotic production can be induced in some of these strains by the provision of carR from ATCC39048 in trans, suggesting that lack of carbapenem production in these bacteria is due to defects in their CarR proteins or defects in CarR production [64]. The CarR–AHL complex binds to the carA promoter in the carR–A intergenic region (Figure 1(b)). QS depends on a diffusible signal molecule 3-oxo-C6-AHL produced by CarI synthase. 3-oxo-C6-AHL binds to the regulatory protein CarR. The active CarR-AHL complex binds directly to the carA promoter and activates transcription of the carA-H operon. This strain has two additional luxR homologues, expR (renamed as expR1) and virR which are involved only in regulation of PCWDE production. In rsmA promoter the expR box (filled rectangle) and the 2nd binding site for ExpR1 and ExpR2 proteins (empty rectangle) are designated. No lux box-like elements have been identified in this region [41,68]. The CarR-mediated AHL-dependent expression of the carbapenem operon results not only in production of carbapenem antibiotic encoded by carA-E genes and also expression of carbapenem resistance functions encoded by carFG [68]. Disruption of carI and/or carR abolishes carbapenem production and transcription from the QS-dependent carA promoter [68]. However, inactivation of either expR or virR has no obvious effect on carbapenem production, either in the presence or absence of a functional carI gene. Therefore, it appears that CarR is the only LuxR homologue regulating carbapenem production in ATCC39084. It should be noted that neither CarR nor CarR-AHL complex controls PCWDE production (Figure 1(b)). Like many LuxI/LuxR systems, the Pcc carI gene has an overlap with the luxR gene that encodes a second luxR gene expR (renamed as expR1) involved in regulation of PCWDE [23]. It is clear that carbapenem-producing Pcc strains have a single AHL synthase gene CarI, and two different LuxR homologues that recognize the same AHL but control the expression of different genes [41]. Taken together these data clearly demonstrate that Pcc has two different LuxR homologues for the control of carbapenem and PCWDE production (Figure 1(b)) [65,69,70]. Coregulation of these two processes, synthesis of the antibiotic and PCWDE via AHL-dependent QS implies that a high cell population density of pectobacteria, having generated a substantial food resource through degradation of plant cell wall material will also promote carbapenem production to eliminate the competing bacteria from the infection site in planta.
2.6. Environmental Impact of AHL Stability in Quorum Sensing
An earlier report showed that AHL levels rise during the log phase of growth and then decline during the stationary phase when bacteria are cultured in rich synthetic media [64]. Under alkaline conditions, AHLs are hydrolyzed and thereby are unable to function as signal molecules for LuxR-type receptor proteins. AHL become unstable over a narrow pH range (pH from 7 to 8). The sensitivity of AHL to extreme pH (pH of >8.2) may be important during infection in planta. So, one of the first recognizable responses of plants to Pectobacterium infection is the pH increase of the apoplastic fluid around the infection site [71]. This is achieved by activating a very rapid H+ influx into plant cells during the early stages of infection (prior to any signs of tissue maceration) which increases the intercellular pH from 6.4 to 8.3 [72]. The most likely mechanisms to break down AHLs are either hydrolysis of the amide bond between the lactone ring and the acyl chain or hydrolysis of the ester bond within the lactone ring. The latter bond is more labile and delactonization of AHL has been shown to occur in vitro at pH 12 [73]. Instability of AHL also increases at high temperature. Although in Pcc strain ATTn10 grown at 37 °C carI transcription is not affected, the concentration of 3-oxo-C6-AHL is reduced below the amount shown to be required for carbapenem production induction. So, degradation of 3-oxo-C6-AHL at 37 °C may be responsible for the effect of temperature on carbapenem production [68].
Oxygen availability also affects AHL stability. This effect can probably be explained by different pH values of the growth environment resulting from either aerobic or anaerobic metabolism. Aerobic metabolism in complex medium (e.g., L-broth) is known to result in alkalinization of the environment due to the preferential use of weak acids as carbon sources which explains degradation of AHL in stationary phase cultures [74]. Anaerobic metabolism does the opposite, and results in net production of weak acids that may account for the persistence of AHL during stationary phase, and explain why soft rot occurs much more readily when oxygen is limiting [3].
Recently, many different bacteria belonging to various genera were reported to express AHL-degrading activity [75]. Two groups of AHL-degrading enzymes have been identified so far: AHL lactonases [76] and AHL acylases (AiiA) [77]. AHL lactonases hydrolyze the lactone ring in the homoserine moiety of AHLs without affecting the rest of the molecule structure. AiiA from Bacillus sp. 24B1 was one of the first AHL lactonases characterized [76]. During recent years more bacteria possessing AHL lactonase activity have been described [37,78,79]. AHL acylases hydrolyze the amide bond between the acyl side chain and the homoserine lactone in the AHL molecule [80]. To the present, nine AHL acylases from various groups of bacteria have been reported and five of them described in detail [77,81,82]. Most of AHL-utilizing bacterial strains were reported as soil or rhizosphere inhabitants [83,84]. The ability to inactivate AHLs might be useful in controlling virulence of many plant pathogenic bacteria [85–87].
The data reviewed here clearly demonstrate that loss of bacterial communication system does not always result from mutational inactivation of AHL synthase (expI), but may also be caused by enzymatic or nonenzymatic degradation of AHL under certain environmental conditions.
3. LuxS/AI-2-Based Quorum Sensing in Pectobacteria
3.1. General Aspects of LuxS/AI-2 Quorum Sensing
A different QS sensing system has been described in several bacteria. This system is based on AI-2 and it was first identified in a Gram-negative bacterium V. harveyii [88]. In this QS system, the first signal molecule identified to bind the AI-2 receptor was a furanosyl borate diester. The synthesis of AI-2 is dependent on the enzyme LuxS [89]. In V. harveyii at high cell density, AI-2 binds LuxP (a receptor protein), initiating a dephosphorylation cascade which results in dephosphorylation and inactivation of the response regulator LuxO; inactivation of LuxO allows expression of LuxRVh which in turn activates the expression of the bioluminescence operon [90–92]. Numerous Gram-negative and Gram-positive bacteria in addition to V. harveyii have a luxS homologue and synthesize compounds capable of activating the AI-2 biosensor [91].
LuxS protein, the product of the luxS gene which is widely conserved throughout the bacterial kingdom, is responsible for AI-2 biosynthesis. LuxS synthesizes 4,5-dihydroxy-2,3-pentanedione (DPD), which undergoes spontaneous rearrangements to form a variety of DPD derivatives that interconvert comprising the AI-2 pool. Cyclization of DPD generates compounds (2R,4S)-and (2S,4S)-2,4-dihydroxy-2-methyldihydrofuranone R- and S-DHMF, respectively and boronoate ester formation from DPD occurs if enough borate is present in solution. Because DPD exists in equilibrium with other chemical species in the solution, AI-2 is actually not only a furanosyl borate diester, but this term is used to collectively designate multiple chemical derivatives of DPD [91]. AI-2 responses in different bacterial species can be triggered by different members of the AI-2 pool. Because the chemical nature of the active signaling molecule from this pool varies between the species [92], it is unsurprising that the nature of the AI-2 receptor for these signals is also variable. To date, only three proteins that bind AI-2 signaling components have been characterized [93–95].
The biosynthetic pathway of AI-2 has disclosed a metabolic role for LuxS, involvement in S-adenosylmethionine-utilization pathway [96]. LuxS converts S-ribosylhomocysteine (produced by detoxification of 5-adenosylhomocysteine) to homocysteine (which is recycled back to methionine) and AI-2. Since LuxS has a metabolic role, it is currently unclear how many of the phenotypes described for luxS− mutant actually result from a signaling defect due to the absence of AI-2, and how many are caused by hampered methionine recycling.
3.2. LuxS/AI-2-Based Quorum Sensing in Pectobacterium carotovorum
A luxS homologue from a Pectobacterium was first reported in a derivative of Pcc ATTn10 and in Pba SCRI1043 [97,98]. Then, Coulthurst et al. [99] used the proteomic approach to identify LuxS-dependent proteins in the secretome of Pcc ATTn10 and Pba SCRI1043. Production of secreted virulence factors, particularly a multitude of secreted pectinases and other PCWDE was studied. Protein spots corresponding to pectate lyases, cellulase and protease were all present in reduced amounts in SCC2 (luxS− mutant) if compared with the wild-type Pcc ATTn10. Specifically, the proteins with expression affected in SCC2 were identified as cellulase V (CelV), the major cellulase in Pcc and an important virulence factor [97,100], and the secreted metalloprotease, protease W (PrtW), which participates in disease progression [101]. Also, decreased expression was detected for pectate lyase I, II and III (PelA, B and C) isoforms [49,102]. Pectinases are primary virulence determinants of Pc and pectate lyases are the most important among the pectinases [103]. The corresponding secreted enzyme activities were also measured for the wild-type Pba SCRI1043 and the luxS− mutant, SCC10 grown to stationary phase in phosphate minimal medium. Similarly to Pcc ATTn10, activities of secreted pectate lyase, cellulase and protease were reduced in the luxS− mutant of Pba SCRI0143 compared to the wild-type. These results demonstrate that full production of secreted virulence factors pectate lyase, cellulase and protease is dependent on functional luxS gene in both Pba SCRI1043 and Pcc ATTn10, at least under the conditions applied.
The luxS− mutants of Pba SCRI1043 and SCC10 exhibited modestly decreased swimming motility compared to the wild-type. luxS mutants of several other bacteria, including E. coli were also affected in motility [104,105]. In Pectobacterium, motility enhances the invasion and infection of potato plants [3,106] suggesting that SCC10 may be disadvantaged compared with the wild-type during the initial stages of natural infection.
Reduced level of secreted enzymes in the luxS− mutant and reduced motility of the mutant, if reproduced in planta, are expected to result in decreased virulence. However, in SCC10 (a luxS− mutant) the observed decrease in motility and secreted enzyme production did not translate into a significant decrease in virulence according to the test models applied. In Pcc ATTn10, a potato tuber rotting assay was used to compare the effect of luxS mutation on virulence. SCC2 (a luxS− mutant) exhibited reduced virulence if compared with the wild-type in this test. By contrast, although SCC10 exhibited a slightly lower mean rot than the wild-type Pba SCRI1043 in a tuber rotting test, the difference was not statistically significant. The virulence of SCC10 was also compared with that of the wild-type using a potato stem infection assay [63]. SCC10 failed to show a statistically significant reduction in lesion formation over the length of the assay compared with wild-type Pba SCRI1043. It is likely that the impact of luxS inactivation is dependent on environmental conditions and the nature of the assay used. The reduction in motility of the strain may not have a significant impact in these assays because the bacterial cells were injected directly into the plant tissue, overcoming the need to gain access to the interior of the plant during a ‘real-life’ infection.
In Pcc strain SCC3193 inactivation of the luxSPcc gene is sufficient to partly repress the production of pectinolytic enzymes during early exponential growth phase (6–8 h). These observations and the pattern of AI-2 production demonstrate that there is a correlation between AI-2 level and production of pectinolytic enzymes. Indeed, the maximum difference in pectinolytic enzyme production between the SCC3193 and luxS−Pcc strains coincide with the maximum level of AI-2 in the parental strain SCC3193 which is in the end of exponential growth phase. The fact that luxS−Pcc mutant had an impaired, but not abolished pectinolytic enzyme production capacity, suggests that AI-2 was not solely required for, but rather to contribute in maximization of pectinolytic enzymes production at low cell densities [107]. It should be noted that receptors for AI-2 have not yet been identified in Pectobacterium strains.
The dramatic consequence of the absence of the AHL type QS signal molecules is reduction of the maceration capacity of Pcc [4,5]. The defect in AI-2 production in the luxS−Pcc of SCC3193 caused 50% less tissue maceration in the potato tuber test already by first 36 h after the inoculation. The reduced maceration of plant tissue in case of luxS−Pcc mutant is not related to alteration of the ability of the mutant to synthesize AHL or grow in planta, but rather to the defect in AI-2 production. In agreement with this, the luxS−Pcc mutant was able to cause consequent maceration of plant tissue, and by 72 h of the assay no significant differences in tissue maceration between the parental and luxSPcc mutant strain were detected [107].
This observation provides evidence for the view that in QS regulation in Pcc during later stages of plant infection, the accumulation of AHL signal is sufficient for efficient PCWDE production even in the absence of AI-2 signal. Brader et al. [40] have shown that Pcc strain SCC3193 produces at least six different AHLs depending on growth conditions. It is tempting to speculate that the roles of AHL species and AI-2 type QS signals may vary at different stages of infection and in different host plants. However, regulation of PCWDE production by two QS systems is certainly clearly different. The one comprising ExpI/AHL, affects PCWDE production through modulation of the rsmA expression [10,11]. The other regulatory system comprising LuxS/AI-2 controls PCWDE production by a still unknown mechanism.
It was of interest to determine whether a link can be detected between the luxS/AI-2 and the AHL QS sensing systems of Pcc and Pba. Production of the QS signal molecule AHL throughout the growth was found to be unaffected in both SCC2 and SCC10 mutants compared with their respective wild-type strains [97]. Similarly, production of AI-2 was not affected in the carI mutant of Pcc ATTn10, strain JBC1, which cannot produce AHL [74]. The production of AHL and AI-2 signal molecules also does not depend on each other in SCC3193 [107].
The mechanism(s) by which luxS is ‘regulating’ these phenotypes remain to be determined and several possibilities exist. AI-2 may act as an extracellular signaling molecule. Such a signaling role could involve reporting on the bacterial population density (i.e., quorum sensing) as proposed previously [89], but it may also involve sensing of environmental conditions, presence and metabolic status of other cells. It is also possible that AI-2 could act as an intracellular regulatory molecule. Finally, any phenotype, including alteration of the abundance of a given protein in a luxS mutant could be the indirect result of a metabolic perturbation due to the loss of LuxS activity in the activated methyl cycle [108]. This final possibility could be excluded by complementation of luxS-dependent phenotypes with the addition of synthetic AI-2. The effect of a luxS mutation could be mediated differently for different proteins and the molecular basis of luxS-dependent regulation in Pectobacterium carotovorum remains to be determined.
4. Concluding Remarks
Many Gram-negative plant pathogenic bacteria use some variant of QS system, either the ExpR-I type or the LuxS/AI-2 type or both to assess the environment and regulate production of virulence factors. Initially, regulation through ExpR-I type QS system was perceived as a relatively simple model involving an AHL synthase, an AHL signal and an ExpR-type regulator activating specific genes at a high cell density. This model is valid only for carbapenem production regulation by AHL/CarR. The overall picture for virulence regulation is far more complex. So, the AHL/ExpR QS system of Pcc governing the production of PCWDE feeds into a post-transcriptional regulatory system which monitor the pool size of RsmA protein which in turn modulates the stability of PCWDE mRNAs. The LuxS/AI-2 type QS plays a strain-dependent role in virulence of different Pectobacterium strains. The fact that luxS− mutant has impaired but not abolished pectinolytic enzyme production suggests that AI-2 does not solely determine but rather contribute to maximize PCWDE production during the infection. The advantage of having two different QS systems is still not entirely clear. It can be that with the aid of two different QS systems, the genes can be regulated also at low cell population densities and under diverse environmental conditions, allowing fine-tuning of QS-regulated traits. A rich diversity of chemical structures among QS signaling molecules used by a pathogen most probably enables population density-dependent modulation of virulence gene expression even under conditions when certain types of signal molecules will be degraded.
Acknowledgments
This research was supported by the Estonian Science Foundation grants GLOMR9072 and SF0180088s08.
References
- Toth, I.K.; Birch, P.R. Rotting softly and stealthily. Curr. Opin. Plant Biol 2005, 8, 424–429. [Google Scholar]
- Liu, H.; Coulthurst, S.J.; Pritchard, L.; Hedley, P.E.; Ravensdale, M.; Humphris, S.; Burr, T.; Takle, G.; Brurberg, M.B.; Birch, P.R.; et al. Quorum sensing coordinates brute force and stealth modes of infection in the plant pathogen Pectobacterium atrosepticum. PLoS. Pathog 2008, 20, e1000093. [Google Scholar]
- Perombelon, M.C.M.; Kelman, A. Ecology of the soft rot erwinias. Annu. Rev. Phytopathol 1980, 18, 361–387. [Google Scholar]
- Pirhonen, M.; Flego, D.; Heikinheimo, R.; Palva, E.T. A small diffusible signal molecule is responsible for the global control of virulence and exoenzyme production in the plant pathogen Erwinia carotovora. EMBO J 1993, 12, 2467–2476. [Google Scholar]
- Jones, S.; Yu, B.; Bainton, N.J.; Birdsall, M.; Bycroft, B.W.; Chhabra, S.R.; Cox, A.J.; Golby, P.; Reeves, J.; Stephens, S.; et al. The lux autoinducer regulates the production of exoenzyme virulence determinants in Erwinia carotovora and Pseudomonas aeruginosa. EMBO J 1993, 12, 2477–2482. [Google Scholar]
- Chatterjee, A.; Cui, Y.; Liu, Y.; Dumenyo, C.K.; Chatterjee, A.K. Inactivation of rsmA leads to overproduction of extracellular pectinases, cellulases, and proteases in Erwinia carotovora subsp. carotovora in the absence of the starvation/cell density-sensing signal, N-(3-oxohexanoyl)-L-homoserine lactone. Appl. Environ. Microbiol 1995, 61, 1959–1967. [Google Scholar]
- Mukherjee, A.; Cui, Y.; Ma, W.; Liu, Y.; Chatterjee, A.K. hexA of Erwinia carotovora ssp. carotovora strain Ecc71 negatively regulates production of RpoS and rsmB RNA, a global regulator of extracellular proteins, plant virulence and the quorum-sensing signal, N-(3-oxohexanoyl)-L-homoserine lactone. Environ. Microbiol 2000, 2, 203–215. [Google Scholar]
- Hyytiäinen, H.; Montesano, M.; Palva, E.T. Global regulators ExpA (GacA) and KdgR modulate extracellular enzyme gene expression through the RsmA-rsmB system in Erwinia carotovora subsp. carotovora. Mol. Plant. Microbe Interact 2001, 14, 931–938. [Google Scholar]
- Hyytiäinen, H.; Sjöblom, S.; Palomäki, T.; Tuikkala, A.; Tapio, P.E. The PmrA-PmrB two-component system responding to acidic pH and iron controls virulence in the plant pathogen Erwinia carotovora ssp. carotovora. Mol. Microbiol 2003, 50, 795–807. [Google Scholar]
- Kõiv, V.; Mäe, A. Quorum sensing controls the synthesis of virulence factors by modulating rsmA gene expression in Erwinia carotovora subsp. carotovora. Mol. Genet. Genomics 2001, 265, 287–292. [Google Scholar]
- Chatterjee, A.; Cui, Y.; Chatterjee, A.K. RsmA and the quorum-sensing signal, N-[3-oxohexanoyl]-L-homoserine lactone, control the levels of rsmB RNA in Erwinia carotovora subsp. carotovora by affecting its stability. J. Bacteriol 2002, 184, 4089–4095. [Google Scholar]
- Laasik, E.; Ojarand, M.; Pajunen, M.; Savilahti, H.; Mäe, A. Novel mutants of Erwinia carotovora subsp. carotovora defective in the production of plant cell wall degrading enzymes generated by Mu transpososome-mediated insertion mutagenesis. FEMS Microbiol. Lett 2005, 243, 93–99. [Google Scholar]
- Sjöblom, S.; Brader, G.; Koch, G.; Palva, E.T. Cooperation of two distinct ExpR regulators controls quorum sensing specificity and virulence in the plant pathogen Erwinia carotovora. Mol. Microbiol 2006, 60, 1474–1489. [Google Scholar]
- Barnard, A.M.; Salmond, G.P. Quorum sensing in Erwinia species. Anal. Bioanal. Chem 2007, 387, 415–423. [Google Scholar]
- Redfield, J.R. Is quorum sensing a side effect diffusion sensing? Trends Microbiol 2002, 10, 365–310. [Google Scholar]
- Mäe, A.; Montesano, M.; Koiv, V.; Palva, E.T. Transgenic plants producing the bacterial pheromone N-acyl-homoserine lactone exhibit enhanced resistance to the bacterial phytopathogen Erwinia carotovora. Mol. Plant Microbe Interact 2001, 14, 1035–1042. [Google Scholar]
- Barnard, A.M.; Bowden, S.D.; Burr, T.; Coulthurst, S.J.; Monson, R.E.; Salmond, G.P. Quorum sensing, virulence and secondary metabolite production in plant soft-rotting bacteria. Philos. Trans. R. Soc. Lond. B. Biol. Sci 2007, 362, 1165–1183. [Google Scholar]
- Nealson, K.H.; Platt, T.; Hastings, J.W. Cellular control of the synthesis and activity of the bacterial luminescent system. J. Bacteriol 1970, 104, 313–322. [Google Scholar]
- Nealson, K.H.; Hastings, J.W. Bacterial bioluminescence: Its control and ecological significance. Microbiol. Rev 1979, 43, 496–518. [Google Scholar]
- de Kievit, T.R.; Iglewski, B.H. Bacterial quorum sensing in pathogenic relationships. Infect. Immun 2000, 68, 4839–4849. [Google Scholar]
- Stevens, A.M.; Greenberg, E.P. Quorum sensing in Vibrio fischeri: Essential elements for activation of the luminescence genes. J. Bacteriol 1997, 179, 557–562. [Google Scholar]
- Kuo, A.; Blough, N.V.; Dunlap, P.V. Multiple N-acyl-L-homoserine lactone autoinducers of luminescence in the marine symbiotic bacterium Vibrio fischeri. J. Bacteriol 1994, 176, 7558–7565. [Google Scholar]
- Swift, S.; Downie, J.A.; Whitehead, N.A.; Barnard, A.M.; Salmond, G.P.; Williams, P. Quorum sensing as a population-density-dependent determinant of bacterial physiology. Adv. Microb. Physiol 2001, 45, 199–270. [Google Scholar]
- Engebrecht, J.; Nealson, K.; Silverman, M. Bacterial bioluminescence: Isolation and genetic analysis of functions from Vibrio fischeri. Cell 1983, 32, 773–781. [Google Scholar]
- Singh, P.K.; Schaefer, A.L.; Parsek, M.R.; Moninger, T.O.; Welsh, M.J.; Greenberg, E.P. Quorum-sensing signals indicate that cystic fibrosis lungs are infected with bacterial biofilms. Nature 2000, 407, 762–764. [Google Scholar]
- Piper, K.R.; von Bodman, S.; Farrand, S.K. Conjugation factor of Agrobacterium tumefaciens regulates Ti plasmid transfer by autoinduction. Nature 1993, 362, 448–450. [Google Scholar]
- Zhang, L.; Murphy, P.J.; Kerr, A.; Tate, M.E. Agrobacterium conjugation and gene regulation by N-acyl-L-homoserine lactones. Nature 1993, 362, 446–448. [Google Scholar]
- von Bodman, S.B.; Majerczak, D.R.; Coplin, D.L. A negative regulator mediates quorum-sensing control of exopolysaccharide production in Pantoea stewartii subsp. stewartii. Proc. Natl. Acad. Sci. USA 1998, 95, 7687–7692. [Google Scholar]
- von Bodman, S.B.; Ball, J.K.; Faini, M.A.; Herrera, C.M.; Minogue, T.D.; Urbanowski, M.L.; Stevens, A.M. The quorum sensing negative regulators EsaR and ExpR(Ecc), homologues within the LuxR family, retain the ability to function as activators of transcription. J. Bacteriol 2003, 185, 7001–7007. [Google Scholar]
- von Bodman, S.; Farrand, S.K. Capsular polysaccharide biosynthesis and pathogenicity in Erwinia stewartii require induction by an N-acylhomoserine lactone autoinducer. J. Bacteriol 1995, 177, 5000–5008. [Google Scholar]
- Minogue, T.D.; Wehland-von Trebra, M.; Bernhard, F.; von Bodman, S.B. The autoregulatory role of EsaR, a quorum-sensing regulator in Pantoea stewartii ssp. stewartii: Evidence for a repressor function. Mol. Microbiol 2002, 44, 1625–1635. [Google Scholar]
- Moré, M.I.; Finger, L.D.; Stryker, J.L.; Fuqua, C.; Eberhard, A.; Winans, S.C. Enzymatic synthesis of a quorum-sensing autoinducer through use of defined substrates. Science 1996, 272, 1655–1658. [Google Scholar]
- Parsek, M.R.; Val, D.L.; Hanzelka, B.L.; Cronan, J.E.; Greenberg, E.P. Acyl homoserine-lactone quorum-sensing signal generation. Proc. Natl. Acad. Sci. USA 1999, 96, 4360–4365. [Google Scholar]
- Parsek, M.R.; Schaefer, A.L.; Greenberg, E.P. Analysis of random and site-directed mutations in rhII, a Pseudomonas aeruginosa gene encoding an acylhomoserine lactone synthase. Mol. Microbiol 1997, 26, 301–310. [Google Scholar]
- Hanzelka, B.L.; Stevens, A.M.; Parsek, M.R.; Crone, T.J.; Greenberg, E.P. Mutational analysis of the Vibrio fischeri LuxI polypeptide: Critical regions of an autoinducer synthase. J. Bacteriol 1997, 179, 4882–4887. [Google Scholar]
- Zhu, J.; Winanas, S.C. The quorum-sensing transcriptional regulator TraR requires its cognate signaling ligand for protein folding, protease resistance, and dimerization. Proc. Natl. Acad. Sci. USA 2001, 98, 1507–1512. [Google Scholar]
- Zhang, H.B.; Wang, L.H.; Zhang, L.H. Genetic control of quorum-sensing signal turnover in Agrobacterium tumefaciens. Proc. Natl. Acad. Sci. USA 2002, 99, 4638–4643. [Google Scholar]
- Egland, K.A.; Greenberg, E.P. Quorum sensing in Vibrio fischeri: Elements of the luxl promoter. Mol. Microbiol 1999, 31, 1197–1204. [Google Scholar]
- Lazdunski, A.M.; Ventre, I.; Sturgis, J.N. Regulatory circuits and communication in Gram-negative bacteria. Nat. Rev. Microbiol 2004, 2, 581–592. [Google Scholar]
- Brader, G.; Sjöblom, S.; Hyytiäinen, H.; Sims-Huopaniemi, K.; Palva, E.T. Altering substrate chain length specificity of an acylhomoserine lactone synthase in bacterial communication. J. Biol. Chem 2005, 280, 10403–10409. [Google Scholar]
- Welch, M.; Todd, D.E.; Whitehead, N.A.; McGowan, S.J.; Bycroft, B.W.; Salmond, G.P. N-acyl homoserine lactone binding to the CarR receptor determines quorum-sensing specificity in Erwinia. EMBO J 2000, 15, 631–641. [Google Scholar]
- Whitehead, N.A.; Barnard, A.M.; Slater, H.; Simpson, N.J.; Salmond, G.P. Quorum-sensing in Gram-negative bacteria. FEMS Microbiol. Rev 2001, 25, 365–404. [Google Scholar]
- Qin, Y.; Luo, Z.Q.; Smyth, A.J.; Gao, P.; von Bodman, S.; Farrand, S.K. Quorum-sensing signal binding results in dimerization of TraR and its release from membranes into the cytoplasm. EMBO J 2000, 19, 5212–5221. [Google Scholar]
- Vannini, A.; Volpari, C.; Di Marco, S. Crystal structure of the quorum-sensing protein TraM and its interaction with the transcriptional regulator TraR. J. Biol. Chem 2004, 279, 24291–24296. [Google Scholar]
- Pappas, K.M.; Weingart, C.L.; Winans, S.C. Chemical communication in proteobacteria: Biochemical and structural studies of signal synthases and receptors required for intercellular signalling. Mol. Microbiol 2004, 53, 755–769. [Google Scholar]
- Gray, K.M.; Passador, L.; Iglewski, B.H.; Greenberg, E.P. Interchangeability and specificity of components from the quorum-sensing regulatory systems of Vibrio fischeri and Pseudomonas aeruginosa. J. Bacteriol 1994, 176, 3076–3080. [Google Scholar]
- Watson, W.T.; Minogue, T.D.; Val, D.L.; von Bodman, S.B.; Churchill, M.E. Structural basis and specificity of acyl-homoserine lactone signal production in bacterial quorum sensing. Mol. Cell 2002, 9, 685–694. [Google Scholar]
- Chatterjee, A.; Cui, Y.; Hasegawa, H.; Leigh, N.; Dixit, V.; Chatterjee, A.K. Comparative analysis of two classes of quorum-sensing signaling systems that control production of extracellular proteins and secondary metabolites in Erwinia carotovora subspecies. J. Bacteriol 2005, 187, 8026–8038. [Google Scholar]
- Bell, K.S.; Sebaihia, M.; Pritchard, L.; Holden, M.T.; Hyman, L.J.; Holeva, M.C.; Thomson, N.R.; Bentley, S.D.; Churcher, L.J.; et al. Genome sequence of the enterobacterial phytopathogen Erwinia carotovora subsp. atroseptica and characterization of virulence factors. Proc. Natl. Acad. Sci. USA 2004, 101, 11105–11110. [Google Scholar]
- Cui, Y.; Chatterjee, A.; Hasegawa, H.; Dixit, V.; Leigh, N.; Chatterjee, A.K. ExpR, a LuxR homolog of Erwinia carotovora subsp. carotovora, activates transcription of rsmA, which specifies a global regulatory RNA-binding protein. J. Bacteriol 2005, 187, 4792–4803. [Google Scholar]
- Cui, Y.; Chatterjee, A.; Hasegawa, H.; Chatterjee, A.K. Erwinia carotovora subspecies produce duplicate variants of ExpR, LuxR homologs that activate rsmA transcription but differ in their interactions with N-acylhomoserine lactone signals. J. Bacteriol 2006, 188, 4715–4726. [Google Scholar]
- Burr, T.; Barnard, A.M.; Corbett, M.J.; Pemberton, C.L.; Simpson, N.J.; Salmond, G.P. Identification of the central quorum sensing regulator of virulence in the enteric phytopathogen, Erwinia carotovora: The VirR repressor. Mol. Microbiol 2006, 59, 113–125. [Google Scholar]
- McGowan, S.; Sebaihia, M.; Jones, S.; Yu, B.; Bainton, N.; Chan, P.F.; Bycroft, B.; Stewart, G.S.; Williams, P.; Salmond, G.P. Carbapenem antibiotic production in Erwinia carotovora is regulated by CarR, a homologue of the LuxR transcriptional activator. Microbiology 1995, 141, 541–550. [Google Scholar]
- Andersson, R.A.; Eriksson, A.R.; Heikinheimo, R.; Mäe, A.; Pirhonen, M.; Kõiv, V.; Hyytiäinen, H.; Tuikkala, A.; Palva, E.T. Quorum sensing in the plant pathogen Erwinia carotovora subsp. carotovora: The role of expR(Ecc). Mol. Plant Microbe Interact 2000, 13, 384–393. [Google Scholar]
- Cui, Y.; Chatterjee, A.; Liu, Y.; Dumenyo, C.K.; Chatterjee, A.K. Identification of a global repressor gene, rsmA, of Erwinia carotovora subsp. carotovora that controls extracellular enzymes, N-(3-oxohexanoyl)-L-homoserine lactone, and pathogenicity in soft-rotting Erwinia spp. J. Bacteriol 1995, 177, 5108–5115. [Google Scholar]
- Nasser, W.; Bouillant, M.L.; Salmond, G.P.; Reverchon, S. Characterization of the Erwinia chrysanthemi expI-expR locus directing the synthesis of two N-acyl-homoserine lactone signal molecules. Mol. Microbiol 1998, 29, 1391–1405. [Google Scholar]
- Reverchon, S.; Bouillant, M.L.; Salmond, G.P.; Nasser, W. Integration of the quorum-sensing system in the regulatory networks controlling virulence factor synthesis in Erwinia chrysanthemi. Mol. Microbiol 1998, 29, 1407–1418. [Google Scholar]
- Liu, Y.; Cui, Y.; Mukherjee, A.; Chatterjee, A.K. Characterization of a novel RNA regulator of Erwinia carotovora ssp. carotovora that controls production of extracellular enzymes and secondary metabolites. Mol. Microbiol 1998, 29, 219–234. [Google Scholar]
- Hasegawa, H.; Chatterjee, A.; Cui, Y.; Chatterjee, A.K. Elevated temperature enhances virulence of Erwinia carotovora subsp. carotovora strain EC153 to plants and stimulates production of the quorum sensing signal, N-acyl homoserine lactone, and extracellular proteins. Appl. Environ. Microbiol 2005, 71, 4655–4663. [Google Scholar]
- Manefield, M.; Turner, S.L. Quorum sensing in context: Out of molecular biology and into microbial ecology. Microbiology 2002, 148, 3762–3764. [Google Scholar]
- Keller, L.; Surette, M.G. Communication in bacteria: An ecological and evolutionary perspective. Nat. Rev. Microbiol 2006, 4, 49–58. [Google Scholar]
- Pemberton, C.L.; Whitehead, N.A.; Sebaihia, M.; Bell, K.S.; Hyman, L.J.; Harris, S.J.; Matlin, A.J.; Robson, N.D.; Birch, P.R.; Carr, J.P.; et al. Novel quorum-sensing-controlled genes in Erwinia carotovora subsp. carotovora: Identification of a fungal elicitor homologue in a soft-rotting bacterium. Mol. Plant Microbe Interact 2005, 18, 343–353. [Google Scholar]
- Corbett, M.; Virtue, S.; Bell, K.; Birch, P.; Burr, T.; Hyman, L.; Lilley, K.; Poock, S.; Toth, I.; Salmond, G.P. Identification of a new quorum-sensing-controlled virulence factor in Erwinia carotovora subsp. atroseptica secreted via the type II targeting pathway. Mol. Plant Microbe Interact 2005, 18, 334–342. [Google Scholar]
- Holden, M.T.; McGowan, S.J.; Bycroft, B.W.; Stewart, G.S.; Williams, P.; Salmond, G.P. Cryptic carbapenem antibiotic production genes are widespread in Erwinia carotovora: Facile trans activation by the carR transcriptional regulator. Microbiology 1998, 144, 1495–1550. [Google Scholar]
- McGowan, S.J.; Sebaihia, M.; O’Leary, S.; Hardie, K.R.; Williams, P.; Stewart, G.S.; Bycroft, B.W.; Salmond, G.P. Analysis of the carbapenem gene cluster of Erwinia carotovora: Definition of the antibiotic biosynthetic genes and evidence for a novel beta-lactam resistance mechanism. Mol. Microbiol 1997, 26, 545–556. [Google Scholar]
- Bainton, N.J.; Stead, P.; Chhabra, S.R.; Bycroft, B.W.; Salmond, G.P.; Stewart, G.S.; Williams, P. N-(3-oxohexanoyl)-L-homoserine lactone regulates carbapenem antibiotic production in Erwinia carotovora. Biochem. J 1992, 288, 997–1004. [Google Scholar]
- Bainton, N.J.; Bycroft., B.W.; Chhabra, S.R.; Stead, P.; Gledhill, L.; Hill, P.J.; Rees, C.E.; Winson, M.K.; Salmond, G.P.; Stewart, G.S.; et al. A general role for the lux autoinducer in bacterial cell signalling: Control of antibiotic biosynthesis in Erwinia. Gene 1992, 116, 87–91. [Google Scholar]
- McGowan, S.J.; Barnard, A.M.; Bosgelmez, G.; Sebaihia, M.; Simpson, N.J.; Thomson, N.R.; Todd, D.E.; Welch, M.; Whitehead, N.A.; Salmond, G.P. Carbapenem antibiotic biosynthesis in Erwinia carotovora is regulated by physiological and genetic factors modulating the quorum sensing-dependent control pathway. Mol Microbiol 2005, 55, 526–545. [Google Scholar]
- McGowan, S.J.; Sebaihia, M.; Porter, L.E.; Stewart, G.S.; Williams, P.; Bycroft, B.W.; Salmond, G.P. Analysis of bacterial carbapenem antibiotic production genes reveals a novel beta-lactam biosynthesis pathway. Mol. Microbiol 1996, 22, 415–422. [Google Scholar]
- Whitehead, N.A; Byers, J.T; Commander, P.; Corbett, M.J.; Coulthurst, S.J.; Everson, L.; Harris, A.K.; Pemberton, C.L.; Simpson, N.J.; Slater, H.; et al. The regulation of virulence in phytopathogenic Erwinia species: Quorum sensing, antibiotics and ecological considerations. Antonie Van Leeuwenhoek 2002, 81, 223–231. [Google Scholar]
- Nachin, L.; Barras, F. External pH: An environmental signal that helps to rationalize pel gene duplication in Erwinia chrysanthemi. Mol. Plant Microbe Interact 2000, 13, 882–886. [Google Scholar]
- Baker, C.J.; Mock, N.; Ainson, M.M.; Hutcheson, S.W. Inhibition of the hypersensitive response in tobacco by pectate lyase digests of cell wall and polygalacturonic acid. Physiol. Mol. Plant Pathol 1990, 37, 133–167. [Google Scholar]
- Eberhard, A.; Burlingame, A.L.; Eberhard, C.; Kenyon, G.L.; Nealson, K.H.; Oppenheimer, N.J. Structural identification of autoinducer of Photobacterium fischeri luciferase. Biochemistry 1981, 20, 2444–2449. [Google Scholar]
- Byers, J.T.; Lucas, C.; Salmond, G.P.; Welch, M. Nonenzymatic turnover of an Erwinia carotovora quorum-sensing signaling molecule. J. Bacteriol 2002, 184, 163–171. [Google Scholar]
- Wang, L.H.; Weng, L.X.; Dong, Y.H.; Zhang, L.H. Specificity and enzyme kinetics of the quorum-quenching N-Acyl homoserine lactone lactonase (AHL-lactonase). J. Biol. Chem 2004, 279, 13645–13651. [Google Scholar]
- Dong, Y.H.; Xu, J.L.; Li, X.Z.; Zhang, L.H. AiiA, an enzyme that inactivates the acylhomoserine lactone quorum-sensing signal and attenuates the virulence of Erwinia carotovora. Proc. Natl. Acad. Sci. USA 2000, 97, 3526–3531. [Google Scholar]
- Lin, Y.H.; Xu, J.L.; Hu, J.; Wang, L.H.; Ong, S.L.; Leadbetter, J.R.; Zhang, L.H. Acyl-homoserine lactone acylase from Ralstonia strain XJ12B represents a novel and potent class of quorum-quenching enzymes. Mol. Microbiol 2003, 47, 849–860. [Google Scholar]
- Park, S.Y.; Lee, S.J.; Oh, T.K.; Oh, J.W.; Koo, B.T.; Yum, D.Y.; Lee, J.K. AhlD, an N-acylhomoserine lactonase in Arthrobacter sp., and predicted homologues in other bacteria. Microbiology 2003, 149, 541–550. [Google Scholar]
- Park, S.Y.; Hwang, B.J.; Shin, M.H.; Kim, J.A.; Kim, H.K.; Lee, J.K. N-acylhomoserine lactonase producing Rhodococcus spp. with different AHL-degrading activities. FEMS Microbiol. Lett 2006, 261, 102–108. [Google Scholar]
- Leadbetter, J.R.; Greenberg, E.P. Metabolism of acyl-homoserine lactone quorum-sensing signals by Variovorax paradoxus. J. Bacteriol 2000, 182, 6921–6926. [Google Scholar]
- Huang, J.J.; Petersen, A.; Whiteley, M.; Leadbetter, J.R. Identification of QuiP, the product of gene PA1032, as the second acyl-homoserine lactone acylase of Pseudomonas aeruginosa PAO1. Appl. Environ. Microbiol 2006, 72, 1190–1197. [Google Scholar]
- Park, S.Y.; Kang, H.O.; Jang, H.S.; Lee, J.K.; Koo, B.T.; Yum, D.Y. Identification of extracellular N-acylhomoserine lactone acylase from a Streptomyces sp. and its application to quorum quenching. Appl. Environ. Microbiol 2005, 71, 2632–2641. [Google Scholar]
- Park, S.J.; Park, S.Y.; Ryu, C.M.; Park, S.H.; Lee, J.K. The role of AiiA, a quorum-quenching enzyme from Bacillus thuringiensis, on the rhizosphere competence. J. Microbiol. Biotechnol 2008, 18, 1518–1521. [Google Scholar]
- Riaz, K.; Elmerich, C.; Moreira, D.; Raffoux, A.; Dessaux, Y.; Faure, D. A metagenomic analysis of soil bacteria extends the diversity of quorum-quenching lactonases. Environ. Microbiol 2008, 10, 560–570. [Google Scholar]
- Uroz, S.; D’Angelo-Picard, C.; Carlier, A.; Elasri, M.; Sicot, C.; Petit, A.; Oger, P.; Faure, D.; Dessaux, Y. Novel bacteria degrading N-acylhomoserine lactones and their use as quenchers of quorum-sensing-regulated functions of plant-pathogenic bacteria. Microbiology 2003, 149, 1981–1989. [Google Scholar]
- Dong, Y.H.; Zhang, X.F.; Xu, J.L.; Zhang, L.H. Insecticidal Bacillus thuringiensis silences Erwinia carotovora virulence by a new form of microbial antagonism, signal interference. Appl. Environ. Microbiol 2004, 70, 954–960. [Google Scholar]
- Jafra, S.; Przysowa, J.; Czajkowski, R.; Michta, A.; Garbeva, P.; van der Wolf, J.M. Detection and characterization of bacteria from the potato rhizosphere degrading N-acyl-homoserine lactone. Can. J. Microbiol 2006, 52, 1006–1015. [Google Scholar]
- Xavier, K.B.; Bassler, B.L. Regulation of uptake and processing of the quorum-sensing autoinducer AI-2 in Escherichia coli. J. Bacteriol 2005, 187, 238–248. [Google Scholar]
- Schauder, S.; Shokat, K.; Surette, M.G.; Bassler, B.L. The LuxS family of bacterial autoinducers: Biosynthesis of a novel quorum-sensing signal molecule. Mol. Microbiol 2001, 41, 463–476. [Google Scholar]
- Schauder, S.; Bassler, B.L. The languages of bacteria. Genes Dev 2001, 15, 1468–1480. [Google Scholar]
- Henke, J.M.; Bassler, B.L. Three parallel quorum-sensing systems regulate gene expression in Vibrio harveyii. J. Bacteriol 2004, 186, 6902–6914. [Google Scholar]
- Miller, S.T.; Xavier, K.B.; Campagna, S.R.; Taga, M.E.; Semmelhack, M.F.; Bassler, B.L.; Hughson, F.M. Salmonella typhimurium recognizes a chemically distinct form of the bacterial quorum-sensing signal AI-2. Mol. Cell 2004, 15, 677–687. [Google Scholar]
- Zhu, J.; Miller, M.B.; Vance, R.E.; Dziejman, M.; Bassle, B.L.; Mekalanos, J.J. Quorum-sensing regulators control virulence gene expression in Vibrio cholerae. Proc. Natl. Acad. Sci. USA 2002, 99, 3129–3134. [Google Scholar]
- Chen, X.; Schauder, S.; Portier, N.; van Dorsselaer, A.; Pelczer, I.; Bassler, B.L.; Hughson, F.M. Structural indentification of a bacterial quorum-sensing signal containing boron. Nature 2002, 415, 545–549. [Google Scholar]
- Miller, S.T.; Xavier, K.B.; Campagna, S.R.; Taga, M.E.; Semmelhack, M.F.; Bassler, B.L.; Hughson, F.M. Salmonella typhimurium recognizes a chemical distinct form of the bacterial quorum-sensing signal AI-2. Mol. Cell 2004, 15, 677–687. [Google Scholar]
- Winzer, K.; Hardie, K.R.; Burgess, N.; Doherty, N.; Kirke, D.; Holden, M.T.; Linforth, R.; Cornell, K.A.; Taylor, A.J.; Hill, P.J.; et al. LuxS: Its role in central metabolism and the in vitro synthesis of 4-hydroxy-5-methyl-3(2H)-furanone. Microbiology 2002, 148, 909–922. [Google Scholar]
- Coulthurst, S.J.; Lilley, K.S.; Salmond, G.P. Genetic and proteomic analysis of the role of luxS in the enteric phytopathogen, Erwinia carotovora. Mol. Plant Pathol 2006, 7, 31–45. [Google Scholar]
- Coulthurst, S.J.; Kurz, C.L.; Salmond, G.P. luxS mutants of Serratia defective in autoinducer-2-dependent ‘quorum sensing’ show strain-dependent impacts on virulence and production of carbapenem and prodigiosin. Microbiology 2004, 150, 901–910. [Google Scholar]
- Cooper, V.J.; Salmond, G.P. Molecular analysis of the major cellulase (CelV) of Erwinia carotovora: Evidence for an evolutionary “mix-and-match” of enzyme domains. Mol. Gen. Genet 1993, 241, 341–350. [Google Scholar]
- Walter, D.; Reeves, P.J.; Salmond, G.P. The major secreted cellulase, CelV, of Erwinia carotovora subsp. carotovora is an important virulence factor. Mol. Plant Microbe Interact 1994, 7, 425–431. [Google Scholar]
- Marits, R.; Kõiv, V.; Laasik, E.; Mäe, A. Isolation of an extracellular protease gene of Erwinia carotovora subsp. carotovora strain SCC3193 by transposon mutagenesis and the role of protease in phytopathogenicity. Microbiology 1999, 145, 1959–1966. [Google Scholar]
- Thomson, N.R.; Nasser, W.; McGowan, S.; Sebaihia, M.; Salmond, G.P. Erwinia carotovora has two KdgR-like proteins belonging to the IciR family of transcriptional regulators: Identification and characterization of the RexZ activator and the KdgR repressor of pathogenesis. Microbiology 1999, 145, 1531–1545. [Google Scholar]
- Toth, I.K.; Bell, K.S.; Holeva, M.C.; Birch, P.R. Soft rot erwiniae: From genes to genomes. Mol. Plant. Pathol 2003, 4, 17–30. [Google Scholar]
- Elvers, K.T.; Park, S.F. Quorum sensing in Campylobacter jejuni: Detection of a luxS encoded signaling molecule. Microbiology 2002, 148, 1475–1481. [Google Scholar]
- Sperandio, V.; Torres, A.G.; Girón, J.A.; Kaper, J.B. Quorum sensing is a global regulatory mechanism in enterohemorrhagic Escherichia coli O157:H7. J. Bacteriol 2001, 183, 5187–5197. [Google Scholar]
- Mulholland, V.; Hinton, J.C.; Sidebotham, J.; Toth, I.K.; Hyman, L.J.; Perombelon, M.C.; Reeves, P.J.; Salmond, G.P. A pleiotropic reduced virulence (Rvi-) mutant of Erwinia carotovora subspecies atroseptica is defective in flagella assembly proteins that are conserved in plant and animal bacterial pathogens. Mol. Microbiol 1993, 9, 343–356. [Google Scholar]
- Laasik, E.; Andresen, L.; Mäe, A. Type II quorum sensing regulates virulence in Erwinia carotovora ssp. carotovora. FEMS Microbiol. Lett 2006, 258, 227–234. [Google Scholar]
- Winzer, K.; Sun, Y.H.; Green, A.; Delory, M.; Blackley, D.; Hardie, K.R.; Baldwin, T.J.; Tang, C.M. Role of Neisseria meningitidis luxS in cell-to-cell signaling and bacteremic infection. Infect. Immun 2002, 70, 2245–2248. [Google Scholar]


{kind=link}
| Strain | Major AHL | LuxI/R homologue(s) | QS regulated phenotype(s) | Reference |
|---|---|---|---|---|
| Class I strains | ||||
| Pcc SCC3193 | 3-oxo-C8-AHL | ExpI/ExpR1/ExpR2 | production of PCWDE; virulence | [4,13] |
| 3-oxo-C6-AHL | ||||
| Pcc EC153 | 3-oxo-C8-AHL | AhlI/ExpR | production of PCWDE; virulence | [48,49] |
| Class II strains | ||||
| Pcc 71 | 3-oxo-C6-AHL | AhlI/ExpR1/ExpR2 | production of PCWDE; virulence | |
| 3-oxo-C8-AHL | [50–52] | |||
| Pcc SCRI193 | 3-oxo-C6-AHL | ExpI/ExpR1/ExpR2 | production of PCWDE; virulence | [5] |
| Pcc ATCC390048 | 3-oxo-C6-AHL | CarI/CarR ExpR1/VirR | production of carbapenem and PCWDE; virulence | [52] |
| Pba SCRI1043 | 3-oxo-C6-AHL | ExpI/ExpR/VirR | production of PCWDE, Nip | [49,52] |
© 2012 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Põllumaa, L.; Alamäe, T.; Mäe, A. Quorum Sensing and Expression of Virulence in Pectobacteria. Sensors 2012, 12, 3327-3349. https://doi.org/10.3390/s120303327
Põllumaa L, Alamäe T, Mäe A. Quorum Sensing and Expression of Virulence in Pectobacteria. Sensors. 2012; 12(3):3327-3349. https://doi.org/10.3390/s120303327
Chicago/Turabian StylePõllumaa, Lee, Tiina Alamäe, and Andres Mäe. 2012. "Quorum Sensing and Expression of Virulence in Pectobacteria" Sensors 12, no. 3: 3327-3349. https://doi.org/10.3390/s120303327
APA StylePõllumaa, L., Alamäe, T., & Mäe, A. (2012). Quorum Sensing and Expression of Virulence in Pectobacteria. Sensors, 12(3), 3327-3349. https://doi.org/10.3390/s120303327