N-Acyl Homoserine Lactone Production by Klebsiella pneumoniae Isolated from Human Tongue Surface



Abstract
: Bacteria communicate by producing quorum sensing molecules called autoinducers, which include autoinducer-1, an N-hexanoyl homoserine lactone (AHL), and autoinducer-2. Bacteria present in the human oral cavity have been shown to produce autoinducer-2, but not AHL. Here, we report the isolation of two AHL-producing Klebsiella pneumoniae strains from the posterior dorsal surface of the tongue of a healthy individual. Spent culture supernatant extracts from K. pneumoniae activated the biosensors Agrobacterium tumefaciens NTL4(pZLR4) and Escherichia coli [pSB401], suggesting the presence of both long and short chain AHLs. High resolution mass spectrometry analyses of these extracts confirmed that both K. pneumoniae isolates produced N-octanoylhomoserine lactone and N-3-dodecanoyl-l-homoserine lactone. To the best of our knowledge, this is the first report of the isolation of K. pneumoniae from the posterior dorsal surface of the human tongue and the production of these AHLs by this bacterium.1. Introduction
The human oral cavity contains more than 700 species of microorganisms [1]. This plethora of bacteria lives in harmony in a healthy individual, therefore it is logical to visualise a dynamic interaction between the host environment and interaction among the oral bacterial consortium. While streptococci represent the major genus, other bacteria, including Actinomyces spp., Capnocytophaga spp., Eikenella spp., Haemophilus spp., Prevotella spp., Propionibacterium spp., Veillonella spp. and Fusobacterium spp., are commonly found in the oral cavity [2]. Hence the oral cavity provides a unique environment for microbial interaction and communication. Most oral bacteria, which include Prevotella intermedia, Porphyromonas gingivalis, Streptococcus gordonii and Streptococcus mutans, produce autoinducer-2 (AI-2) [3–7].
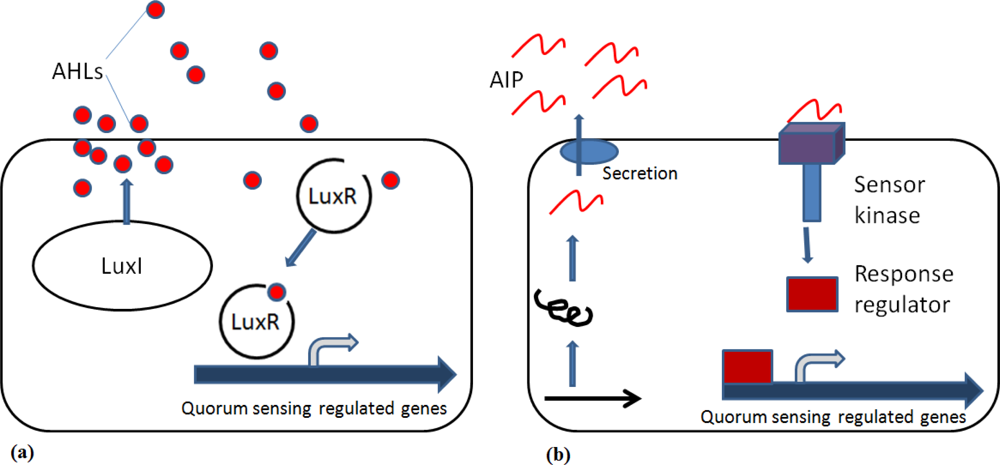
Gram-negative and Gram-positive bacteria communicate by a process termed quorum sensing (QS), which typically involves the production and sensing of QS molecules called autoinducers to link the surrounding cell density with activation of appropriate compensatory regulation of cell function (Figure 1). In Gram-negative bacteria, N-acylhomoserine lactones (AHLs) are arguably the most studied QS signalling molecules. AHLs are usually produced by a protein homologous to LuxI and they will in turn bind to LuxR protein [8]. Once the threshold level of an AHL is detected, transduction leads to the induction of genes that control a variety of survival functions, including the production of antimicrobial substances and protection against the host’s defence mechanisms [9,10].
In Gram-positive bacteria the signalling molecules are post-translationally modified peptides, known as autoinducer peptides (AIP) (Figure 1). Thus far, the only QS mechanism shared by both Gram-positive and Gram-negative bacteria involves the production of AI-2 by the enzyme LuxS. AI-2 appears to be an important signal molecule among different species in a mixed-species community [11] and it differs from AHL, which regulates gene expression in genetically identical cells.
Both Gram-positive and Gram-negative bacteria have been implicated in several systemic infections like bacterial endocarditis, aspiration pneumonia, osteomyelitis in children, preterm low birth weight, cardiovascular disease and dental diseases like caries, periodontal disease and dentoalveolar abscess [1]. Recently, we have analysed the pathogenic Aeromonas sp. from various parts of the human body and found that AHL production is very common in this bacterium [12]. Hence, it is of clinical importance to study the role of autoinducers produced by oral pathogens in determining the health of an individual. Hitherto, there has been no report of AHL production by oral bacteria [2,6,13]. Here we report the isolation of two strains of AHL-producing Klebsiella pneumoniae from the tongue surface of a healthy individual and the characterization of the AHLs produced.
2. Experimental Section
2.1. Bacterial Strains
Besides the oral K. pneumoniae isolated in this work, Escherichia coli DH5α was used as a host for DNA manipulations. Bioreporters E. coli [pSB401] [14] and Agrobacterium tumefaciens NTL4(pZLR4) [15] were used. In the lux-based plasmid-bearing AHL biosensor E. coli [pSB401], the Photorhabdus luminescens luxCDABE cassette has been fused to the Vibrio fischeri luxI promoter and contains the V. fischeri luxR gene [14]. Activation of LuxR by exogenous N-3-oxohexanoyl-l-homoserine lactone (3-oxo-C6-HSL) or a related AHL leads to the expression of luxCDABE and the emission of light. This biosensor is versatile and able to report the structural differences inherent in the AHL family since it responds differentially to AHL molecules with variable acyl side chain lengths. A. tumefaciens NTL4(pZLR4) is an NT1 derivative carrying a traG::lacZ reporter fusion. This strain does not produce its own AHLs, but the traG::lacZ reporter gene is induced only when its transcription activator TraR detects a cognate exogenous AHL. Induction of the reporter gene, leading to production of β-galactosidase enzyme, is measured by using X-gal, a β-galactosidase substrate, for colorimetric (blue pigmentation) detection. A. tumefaciens NTL4(pZLR4) was cultured in AB medium or agar (solidified with bacto-agar at 15 g/L), supplemented with gentamicin (150 μg/mL) and glucose (0.5%, w/v) [15]. For AHL detection with A. tumefaciens NTL4(pZLR4), AB agar was supplemented with X-gal (60 μg/mL, final concentration). All other bacteria were routinely cultured in Lysogeny Broth (LB) medium (in grams per litre: tryptone, 10; yeast extract, 5; and NaCl, 5), without or with bacto-agar (15 g/L), buffered with 50 mM 3-[N-morpholino] propanesulfonic acid (MOPS) to pH 5.5. When necessary, growth media were supplemented with ampicillin (100 μg/mL). A. tumefaciens NTL4(pZLR4) was grown at 28 °C, whereas E. coli and oral bacteria were grown at 37 °C.
2.2. Enrichment of Bacteria from Tongue Surface Debris
This study was approved by the Ethics Committee of the Faculty of Dentistry, University of Malaya. Tongue surface debris was collected from an individual with healthy oral condition in 2008 at the Faculty of Dentistry, University of Malaya. A sterile stainless steel tongue scraper was used to gently scrape the tongue surface to collect the specimen. The debris on the scraper was suspended in 300 μL of sterile saline. To isolate bacteria from the tongue debris, we first evaporated 5 μL of N-heptanoyl-l-homoserine lactone (C7-HSL, 0.1 M), dissolved in absolute ethanol, to dryness and then added the 300 μL of tongue debris to rehydrate the dried C7-HSL. After mixing, 1 μL of NH4Cl (0.3 g/L) was added and the total volume was brought to 3 mL with KG medium [16]. The mixture was incubated at 37 °C (220 rpm) for 48 hours and transferred to fresh NH4Cl and C7-HSL supplemented KG medium at 48 hours interval. Aliquots were withdrawn at the fourth enrichment and spread on LB agar. Pure colonies were obtained by repeated streaking on LB agar. We used C7-HSL, rather than the commonly used even-numbered AHLs [16,17], in this study because we wished to check if any novel bacteria from the tongue surface could be isolated using C7-HSL as the sole carbon source.
2.3. Strain Identification
All DNA manipulations and PCR amplification of 16S rDNA genes with the universal primer pairs 27F and 1525R were carried out as described [18,19]. T7 and SP6 universal primers and internal primers previously designed to anneal to internal target regions of the 16S rDNA were used [20]. Nucleotide sequence alignments and phylogenetic analysis were carried out using LASERGENE software package (DNASTAR, Inc.) and MEGA software version 4.0 [21], respectively. Trees were generated using the Neighbour-Joining algorithm on aligned 16S rDNA gene sequences. Bootstrap analyses for 1,000 resamplings were used to ensure the robustness and reliability of trees constructed.
2.4. Extraction of AHLs from Spent Bacterial Culture Supernatants
An overnight bacterial culture was inoculated into 100 mL of LB broth. Cells were grown to OD600 of 1.0 and the spent culture supernatant was extracted twice with an equal volume of ethyl acetate. The organic layer was collected in a separation funnel, dried over anhydrous magnesium sulphate, filtered, and evaporated to dryness. Residues were dissolved in 100 μL of acetonitrile and stored at −20 °C. AHL extracts were analysed with a luminometer–spectrophotometer used in conjunction with lux-based biosensor E. coli [pSB401], thin layer chromatography and liquid chromatography mass spectrometry.
2.5. Separation and Detection of AHLs by Thin Layer Chromatography (TLC) Analysis
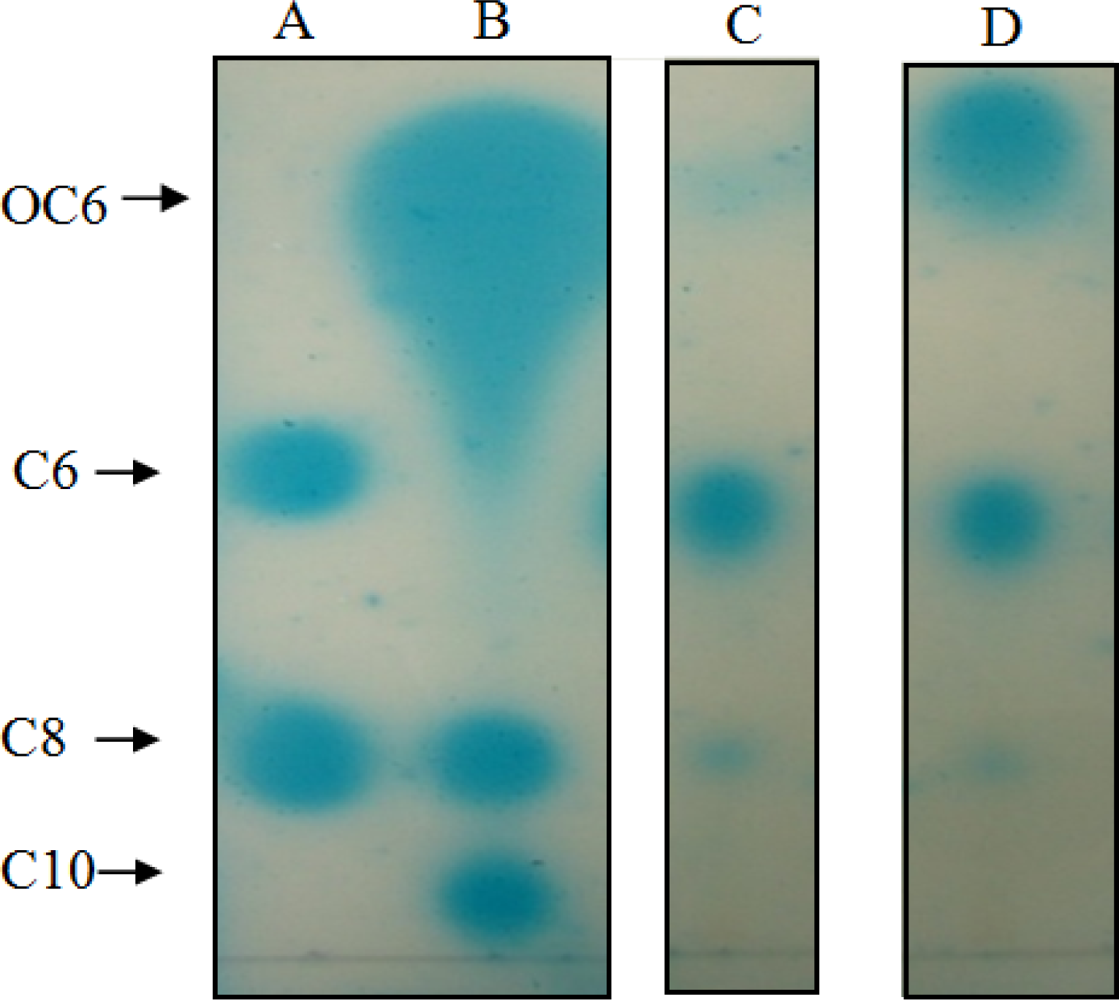
TLC was performed according to the protocol of Shaw et al. [22]. A TLC tank was filled with methanol (60%, v/v) and left for 1 hour to enable saturation. The ethyl acetate extracts (20 μL) and AHL standards (dissolved in acetonitrile) were chromatographed on an RP-18 F254s aluminium plate (20 × 20 cm; Merck, Germany). The AHL standards used were 3-oxo-C6-HSL (0.0008 μg per lane), N-hexanoyl-l-homoserine lactone (C6-HSL) (0.01 μg per lane), N-octanoyl-l-homoserine lactone (C8-HSL) (0.18 μg per lane), and N-decanoyl-l-homoserine lactone (C10-HSL) (0.3 μg per lane), all obtained from Sigma-Aldrich. After chromatography, the plate was air-dried and overlaid with a thin film of X-gal-containing AB agar seeded with overnight-grown A. tumefaciens NTL4(pZLR4). The TLC plate was then incubated for 24 to 48 hours at 28 °C. AHLs were detected by blue pigmentation on the TLC plate, and the result was digitally recorded. Duplicate experiments were carried out.
2.6. Measurement of Bioluminescence
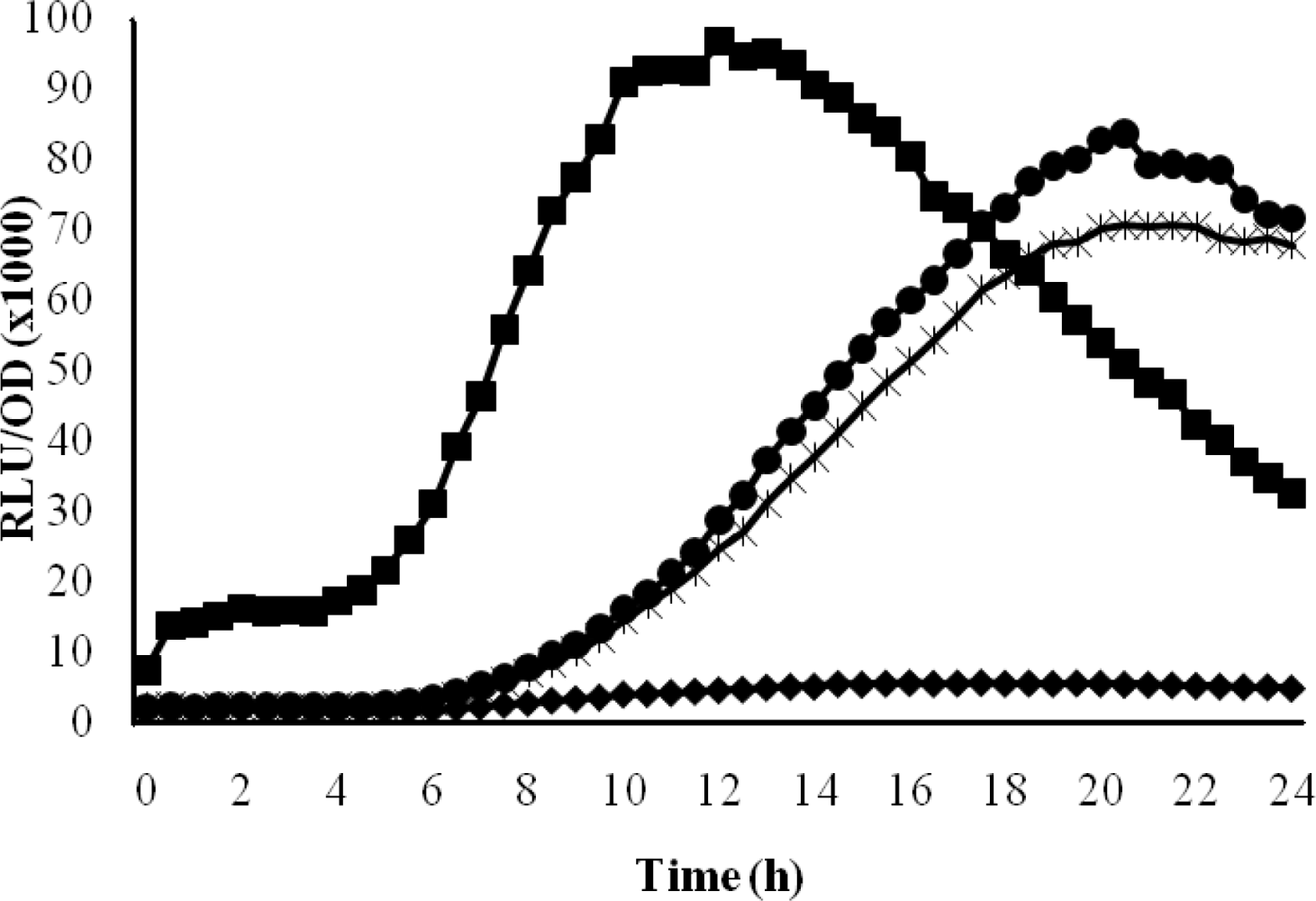
To measure bioluminescence, an overnight culture of E. coli [pSB401] was diluted with LB to OD600 0.01, and 200-μL aliquots of the diluted cells were added to each well of a 96-well optical bottom microtitre plate. AHL extracts from oral bacteria, AHL solvent (ethyl acetate) (control), and N-butanoyl-l-homoserine lactone (C4-HSL) (100 mM, Sigma-Aldrich) (positive control) were added to the wells containing E. coli [pSB401] cells, and incubated at 37 °C for 24 hours in a luminometer-spectrophotometer. Bioluminescence was measured as a function of cell density at 30 minutes interval using a combined automated luminometer–spectrophotometer (Infinite, Tecan). Growth measurement and bioluminescence were the means of triplicate experiments. Data were presented as graph RLU (Relative Light Units)/OD495 nm against time, indicating approximate light output per cell.
2.7. Mass Spectrometry (MS) Analyses of AHLs
High resolution MS was performed as described [23] using the Agilent RRLC 1200 system coupled with an Agilent ZORBAX Rapid Resolution HT column (100 mm × 2.1 mm, 1.8 μm particle size), carried out at 60 °C, flow rate 0.3 mL/min, with injection volume 20 μL. Mobile phases A and B were 0.1% v/v formic acid in water and 0.1% v/v formic acid in acetonitrile, respectively, and the gradient profile is given in Table 1.
The high resolution ESI-MS and ESI-MS/MS analyses were performed on an Agilent 6500 Q-TOF system, operated in the ESI-positive mode, with probe capillary voltage set at 3,000 V; desolvation temperature 350 °C; sheath gas 11 mL/h; and nebulizer pressure 50 psi. Nitrogen gas was used as the collision gas in the collisionally induced dissociation mode for the MS/MS analysis, with collision energy set at 20 eV. Agilent MassHunter software was used to analyze the MS data.
3. Results and Discussion
3.1. Characterization of Oral K. pneumoniae
The growth medium became turbid within 48 hours after inoculation with the tongue surface debris suggesting bacterial growth. At the end of the enrichment process, cell suspensions were serially diluted and streaked on LB agar. Pure colonies with distinctive morphologies were obtained after several successive streaks. Isolates T2-1-1 and T2-1-2 from the tongue surface debris specimen were selected for further analysis. The partial 16S rDNA sequences of T2-1-1 and T2-1-2, each comprising 1,532 nucleotides, were deposited in the GenBank under accession numbers HQ907955 (T2-1-1) and HQ907956 (T2-1-2). Web-based search and phylogenetic analysis showed that both T2-1-1 and T2-1-2 were K. pneumoniae (Figure 2).
3.2. Detection of AHLs
We succeeded in our initial objective of isolating quorum quenching bacteria from the human tongue surface (data not shown). We then tested these isolates for AHL production. To our surprise, isolates T2-1-1 and T2-1-2 produced AHLs, detected initially with A. tumefaciens NTL4(pZLR4) on agar plates–(data not shown). Subsequently, TLC results of the extracts of spent culture supernatants from T2-1-1 and T2-1-2 revealed three well-resolved spots with relative migration factor (Rf) values corresponding to those of 3-oxo-C6-HSL, C6-HSL, and C8-HSL (Figure 3). Strain T2-1-2 appeared to produce more 3-oxo-C6-HSL than T2-1-1.
The production of AHLs by T2-1-1 and T2-1-2 was confirmed by the activation of bioluminescence of E. coli [pSB401] (Figure 4). However, the light emission was delayed in the presence of AHL extracts from strains T2-1-1 and T2-1-2 as compared to the positive control (Figure 4). This is because in the positive control, C4-HSL was added exogenously in a sufficient amount (100 mM) that triggered bioluminescence earlier.
3.3. MS Analyses of the Extracts of Spent Culture Supernatants from Oral Bacteria
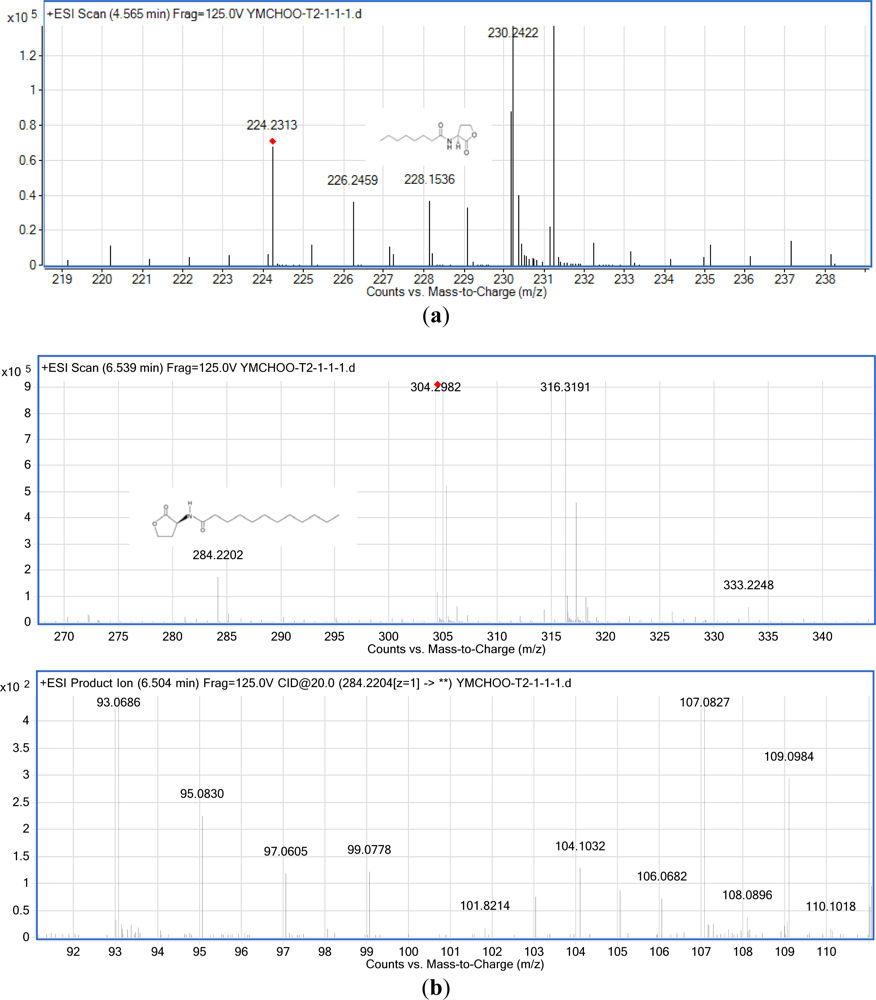
The results of MS (Figure 5) confirmed the presence of C8-HSL (m/z 228.1536) and C12-HSL (m/z 284.2202) in the spent culture supernatant from K. pneumoniae (T2-1-1). The ESI-MS/MS spectrum of C12-HSL shows fragments (m/z 95.0830, 109.0984) typical of lactone-moiety [24]. A similar approach confirmed that K. pneumoniae (T2-1-2) also produced C8-HSL and C12-HSL (the mass spectra for T2-1-2 are not shown). Table 2 summarizes the MS analyses of these AHLs.
Although TLC analysis indicated the presence of C6-HSL and 3-oxo-C6-HSL in the spent culture supernatants from T2-1-1 and T2-1-2, MS did not detect them. The failure to detect short chain AHLs may be due to their instability during MS analysis in our experimental condition or their degradation by T2-1-1 and T2-1-2 during extraction.
Another possible reason is that AHL biosensors, especially A. tumefaciens NTL4(pZLR4), are less reliable although the biosensor-TLC assay method provides a quick and convenient way to detect AHLs, providing the basis for high sensitivity detection by LC-MS/MS and nuclear magnetic resonance spectroscopy. Hence, in this study, we used the high resolution MS analysis, which provides high accuracy of mass determination, to confirm the presence of these AHLs in spent culture supernatants from K. pneumoniae.
K. pneumoniae and K. oxytoca strains have been reported to produce high AHL activities only when grown microaerophilically in LB medium, but insignificant amounts of AHLs when grown in LB medium under aerobic condition [25]. In contrast, in this study K. pneumoniae was shown to produce detectable amounts of AHLs when grown aerobically in LB medium buffered with MOPS. Because AHLs have short half-life in alkali condition [26], the MOPS-buffered LB medium provides protection to AHLs against lactonolysis at basic pH, hence stabilising AHLs and enabling them to be readily detected in our study.
Although AHLs have well defined immune modulatory effects in vitro and are detectable in body fluids (such as sputum from cystic fibrosis patients infected with Pseudomonas aeruginosa) [27], the long term effects of ingesting AHLs into the digestive tracts of human beings remain unknown. We had performed an acid tolerance test on T2-1-1 and T2-1-2, and both isolates were viable when grown for 24 hours in LB buffered at pH 5. However, they did not remain viable when grown in pH > 3 (data not shown). These results led us to speculate that when these AHL-producing bacteria are dislodged from the tongue surface, they may survive gastric juice and persist in human bowels. During the passage from the oral cavity through the digestive tracts, these strains may produce AHLs that could have an impact on the host in vivo.
In this study, K. pneumoniae, which is not commonly found in the oral cavity, was isolated from the posterior dorsal surface of the human tongue. To the best of our knowledge, this is the first report of AHL-producing bacteria on the human tongue surface. Hitherto, oral bacteria have only been reported to produce AI-2, synthesized by LuxS [2,3,5,6]. This study has established the production of AHLs by oral bacteria and has widened the scope of QS research in oral cavity from AI-2 to AHL.
4. Conclusions and Outlook
The tongue surface is a rich source of bacteria including QS bacteria that rely on AHLs as signalling molecules. Here, oral K. pneumoniae isolated from the human tongue surface has been confirmed to produce AHLs by biosensors and high resolution MS.
Acknowledgments
This work was supported by two grants from the University of Malaya, namely a High Impact Research Grant (A000001-50001) to Kok-Gan Chan, and a UMRG grant to Kathivaran Purmal (RG033/09HTM), which are gratefully acknowledged. We thank Yeun-Mun Choo for mass spectrometry analysis and the anonymous reviewers for their constructive comments.
References and Notes
- Aas, J.A.; Paster, B.J.; Stokes, L.N.; Olsen, I.; Dewhirst, F.E. Defining the normal bacterial flora of the oral cavity. J. Clin. Microbiol 2005, 43, 5721–5732. [Google Scholar]
- Kolenbrander, P.E.; Andersen, R.N.; Blehert, D.S.; Egland, P.G.; Foster, J.S.; Palmer, R.J., Jr. Communication among oral bacteria. Microbiol. Mol. Biol. Rev 2002, 66, 486–505. [Google Scholar]
- Burgess, N.A.D.; Kirke, D.F.; Williams, P.; Winzer, K.; Hardie, K.R.; Meyers, N.L.; Aduse-Opoku, L.; Curtis, M.A.; Cámara, M. LuxS-dependent quorum sensing in Porphyromonas gingivalis modulates protease and haemagglutinin activities but is not essential for virulence. Microbiology 2002, 148, 763–772. [Google Scholar]
- Chung, W.O.; Park, Y.; Lamont, R.J.; McNab, R.; Barbieri, B.; Demuth, D.R. Signaling system in Porphyromonas gingivalis based on a LuxS protein. J. Bacteriol 2001, 183, 3903–3909. [Google Scholar]
- Fong, K.P.; Chung, W.O.; Lamont, R.J.; Demuth, D.R. Intra- and interspecies regulation of gene expression by Actinobacillus actinomycetemcomitans LuxS. Infect. Immun 2001, 69, 7625–7634. [Google Scholar]
- Frias, J.; Olle, E.; Alsina, M. Periodontal pathogens produce quorum sensing signal molecules. Infect. Immun 2001, 69, 3431–3434. [Google Scholar]
- Merritt, J.; Qi, F.; Goodman, S.D.; Anderson, M.H.; Shi, W. Mutation of luxS affects biofilm formation in Streptococcus mutans. Infect. Immun 2003, 71, 1972–1979. [Google Scholar]
- Fuqua, C.; Winans, S.C.; Greenberg, E.P. Census and consensus in bacterial ecosystems: The LuxR-LuxI family of quorum-sensing transcriptional regulators. Annu. Rev. Microbiol 1996, 50, 727–751. [Google Scholar]
- Fuqua, C.; Parsek, M.R.; Greenberg, E.P. Regulation of gene expression by cell-to-cell communication: Acyl-homoserine lactone quorum sensing. Annu. Rev. Genet 2001, 35, 439–468. [Google Scholar]
- Williams, P.; Winzer, K.; Chan, W.; Cámara, M. Look who’s talking: Communication and quorum sensing in the bacterial world. Philos. Trans. R. Soc. Lond. B. Biol. Sci 2007, 362, 1119–1134. [Google Scholar]
- Schauder, S.; Bassler, B.L. The languages of bacteria. Genes Dev 2001, 15, 1468–1480. [Google Scholar]
- Chan, K.G.; Puthucheary, S.D.; Chan, X.Y.; Yin, W.F.; Wong, C.S.; See Too, W.S.; Chua, K.H. Quorum sensing in Aeromonas species isolated from patients in Malaysia. Curr. Microbiol 2010, 62, 167–172. [Google Scholar]
- Whittaker, C.J.; Klier, C.M.; Kolenbrander, P.E. Mechanisms of adhesion by oral bacteria. Annu. Rev. Microbiol 1996, 50, 513–552. [Google Scholar]
- Winson, M.K.; Swift, S.; Fish, L.; Throup, J.P.; Jorgensen, F.; Chhabra, S.R.; Bycroft, B.W.; Williams, P.; Stewart, G.A.S.B. Construction and analysis of luxCDABE-based plasmid sensors for investigating N-acyl homoserine lactone-mediated quorum sensing. FEMS Microbiol. Lett 1998, 163, 185–192. [Google Scholar]
- Farrand, S.K.; Hwang, I.; Cook, D.M. The tra region of the nopaline-type Ti plasmid is a chimera with elements related to the transfer systems of RSF1010, RP4, and F. J. Bacteriol 1996, 178, 4233–4247. [Google Scholar]
- Chan, K.G.; Yin, W.F.; Sam, C.K.; Koh, C.L. A novel medium for the isolation of N-acylhomoserine lactone-degrading bacteria. J. Ind. Microbiol. Biotechnol 2009, 36, 247–251. [Google Scholar]
- Chan, K.G.; Atkinson, S.; Mathee, K.; Sam, C.K.; Chhabra, S.R.; Cámara, M.; Koh, C.L.; Williams, P. Characterization of N-acylhomoserine lactone-degrading bacteria associated with the Zingiber officinale (ginger) rhizosphere: Co-existence of quorum quenching and quorum sensing in Acinetobacter and Burkholderia. BMC Microbiol 2011, 11, 51. [Google Scholar] [CrossRef]
- Chan, K.G.; Tiew, S.Z.; Ng, C.C. Rapid isolation method of soil bacilli and screening of their quorum quenching activity. As. Pac. J. Mol. Biol. Biotech 2007, 15, 153–156. [Google Scholar]
- Chan, K.G.; Wong, C.S.; Yin, W.F.; Sam, C.K.; Koh, C.L. Rapid degradation of N-3-oxo-acylhomoserine lactones by a Bacillus cereus isolate from Malaysian rainforest soil. Antonie van Leeuwenhoek 2010, 98, 299–305. [Google Scholar]
- Lane, D.L.; Pace, B.; Olsen, G.J.; Stahl, D.A.; Sogin, M.L.; Pace, N.R. Rapid determination of 16S ribosomal RNA sequences for phylogenetic analyses. Proc. Natl. Acad. Sci. USA 1985, 82, 6955–6959. [Google Scholar]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol 2007, 24, 1596–1599. [Google Scholar]
- Shaw, P.D.; Ping, G.; Daly, S.L.; Cha, C.; Cronan, J.E., Jr.; Rinehart, K.L.; Farrand, S.K. Detecting and characterizing N-acyl-homoserine lactone signal molecules by thin-layer chromatography. Proc. Natl. Acad. Sci. USA 1997, 94, 6036–6041. [Google Scholar]
- Wong, C.S.; Yin, W.F.; Choo, Y.M.; Sam, C.K.; Koh, C.L.; Chan, K.G. Coexistence of quorum-quenching and quorum-sensing in tropical marine Pseudomonas aeruginosa strain MW3A. World J. Microbiol. Biotechnol 2012, 28, 453–461. [Google Scholar] [CrossRef]
- Ortori, C.A.; Atkinson, S.; Chhabra, S.R.; Cámara, M.; Williams, P.; Barret, D.A. Comprehensive profiling of N-acylhomoserine lactones produced by Yersinia pseudotuberculosis using liquid chromatography coupled to hybrid quadrupole-linear ion trap mass spectrometry. Anal. Bioanal. Chem 2007, 387, 497–511. [Google Scholar]
- Wang, H.; Cai, T.; Weng, M.; Zhou, J.; Cao, H.; Zhong, Z.; Zhu, J. Conditional production of acyl-homoserine lactone-type quorum-sensing signals in clinical isolates of enterobacteria. J. Medical. Microbiol 2006, 55, 1751–1753. [Google Scholar]
- Yates, E.A.; Philipp, B.; Buckley, C.; Atkinson, S.; Chhabra, S.R.; Sockett, R.E.; Goldner, M.; Dessaux, Y.; Cámara, M.; Smith, H.; Williams, P. N-acylhomoserine lactones undergo lactonolysis in a pH-, temperature-, and acyl chain length-dependent manner during growth of Yersinia pseudotuberculosis and Pseudomonas aeruginosa. Infect. Immun 2002, 70, 5635–5646. [Google Scholar]
- Pritchard, D.I. Immune modulation by Pseudomonas aeruginosa quorum-sensing signal molecules. Int. J. Med. Microbiol 2006, 296, 111–116. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (min) | Mobile phase A (0.1% v/v formic acid in water) | Mobile phase B (0.1% v/v formic acid in acetonitrile) |
|---|---|---|
| 0.0 | 60% | 40% |
| 5.0 | 20% | 80% |
| 7.0 | 5% | 95% |
| 10.0 | 5% | 95% |
| 11.0 | 60% | 40% |
| 13.0 | 60% | 40% |
| Bacterial strain | Mass spectra |
|---|---|
| K. pneumoniae (T2-1-1) | C8-HSL (m/z 228.1536) |
| C12-HSL (m/z 284.2202; MS/MS: m/z 95.0830, 109.0984) | |
| K. pneumoniae (T2-1-2), mass spectra not shown | C8-HSL (m/z 228.1541) |
| C12-HSL (m/z 284.2202; MS/MS: m/z 95.0828, 109.0977) | |
© 2012 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yin, W.-F.; Purmal, K.; Chin, S.; Chan, X.-Y.; Koh, C.-L.; Sam, C.-K.; Chan, K.-G. N-Acyl Homoserine Lactone Production by Klebsiella pneumoniae Isolated from Human Tongue Surface. Sensors 2012, 12, 3472-3483. https://doi.org/10.3390/s120303472
Yin W-F, Purmal K, Chin S, Chan X-Y, Koh C-L, Sam C-K, Chan K-G. N-Acyl Homoserine Lactone Production by Klebsiella pneumoniae Isolated from Human Tongue Surface. Sensors. 2012; 12(3):3472-3483. https://doi.org/10.3390/s120303472
Chicago/Turabian StyleYin, Wai-Fong, Kathiravan Purmal, Shenyang Chin, Xin-Yue Chan, Chong-Lek Koh, Choon-Kook Sam, and Kok-Gan Chan. 2012. "N-Acyl Homoserine Lactone Production by Klebsiella pneumoniae Isolated from Human Tongue Surface" Sensors 12, no. 3: 3472-3483. https://doi.org/10.3390/s120303472
APA StyleYin, W. -F., Purmal, K., Chin, S., Chan, X. -Y., Koh, C. -L., Sam, C. -K., & Chan, K. -G. (2012). N-Acyl Homoserine Lactone Production by Klebsiella pneumoniae Isolated from Human Tongue Surface. Sensors, 12(3), 3472-3483. https://doi.org/10.3390/s120303472