
Development and Experimental Validation of a Dry Non-Invasive Multi-Channel Mouse Scalp EEG Sensor through Visual Evoked Potential Recordings


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
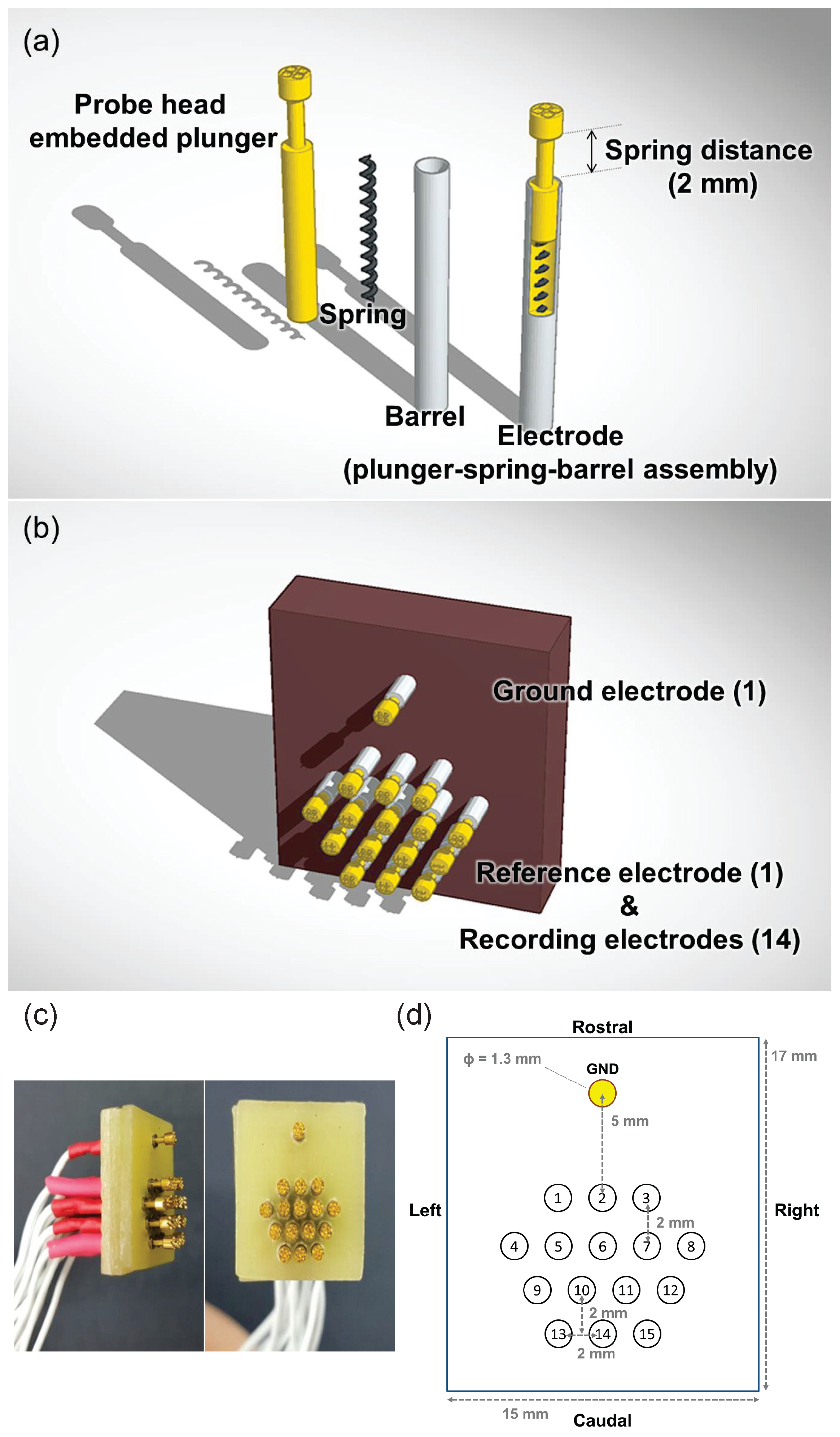
2. Development of the Mouse EEG Sensor
2.1. Components
2.2. Structure
2.3. Instructions
3. In Vivo Test: Visual Evoked Potential Experiment
3.1. Experimental Environments and Apparatus
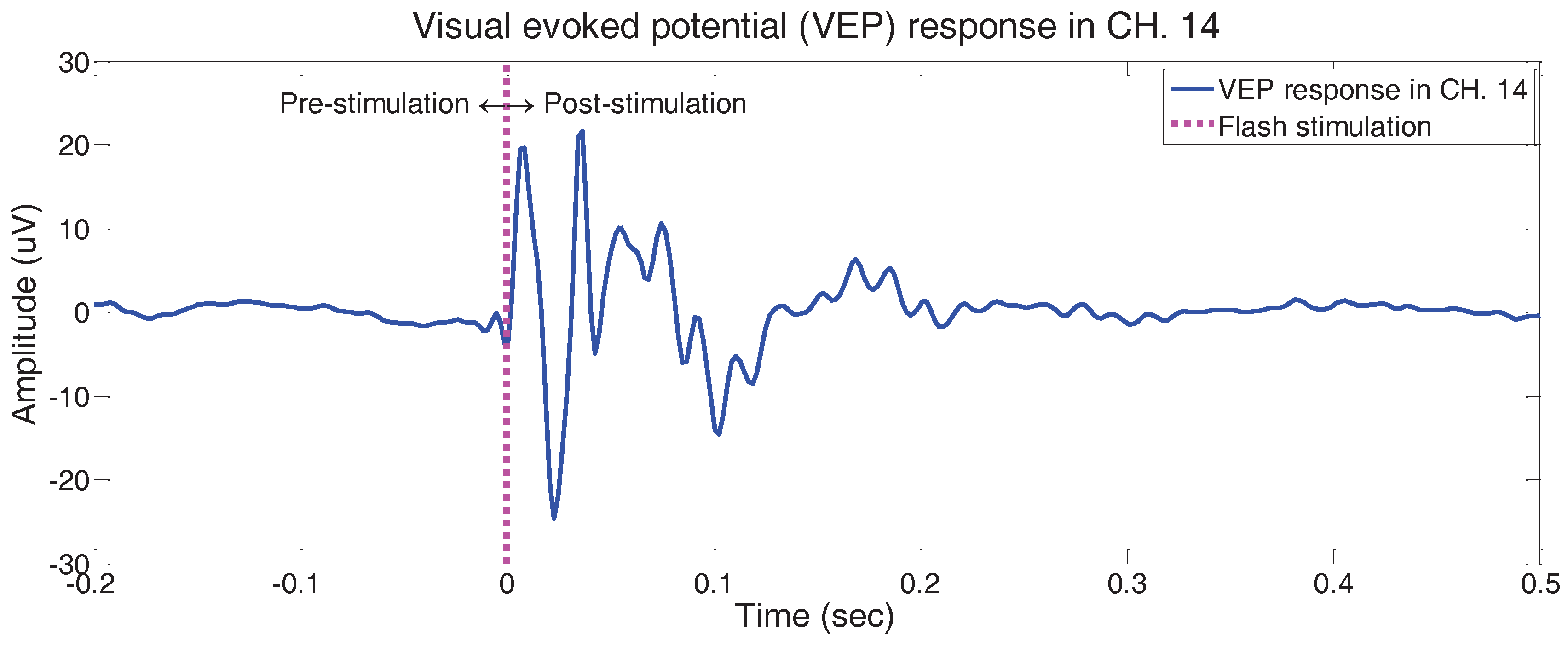
3.2. Experimental Results
3.2.1. Comparison with Existing Results in Literature
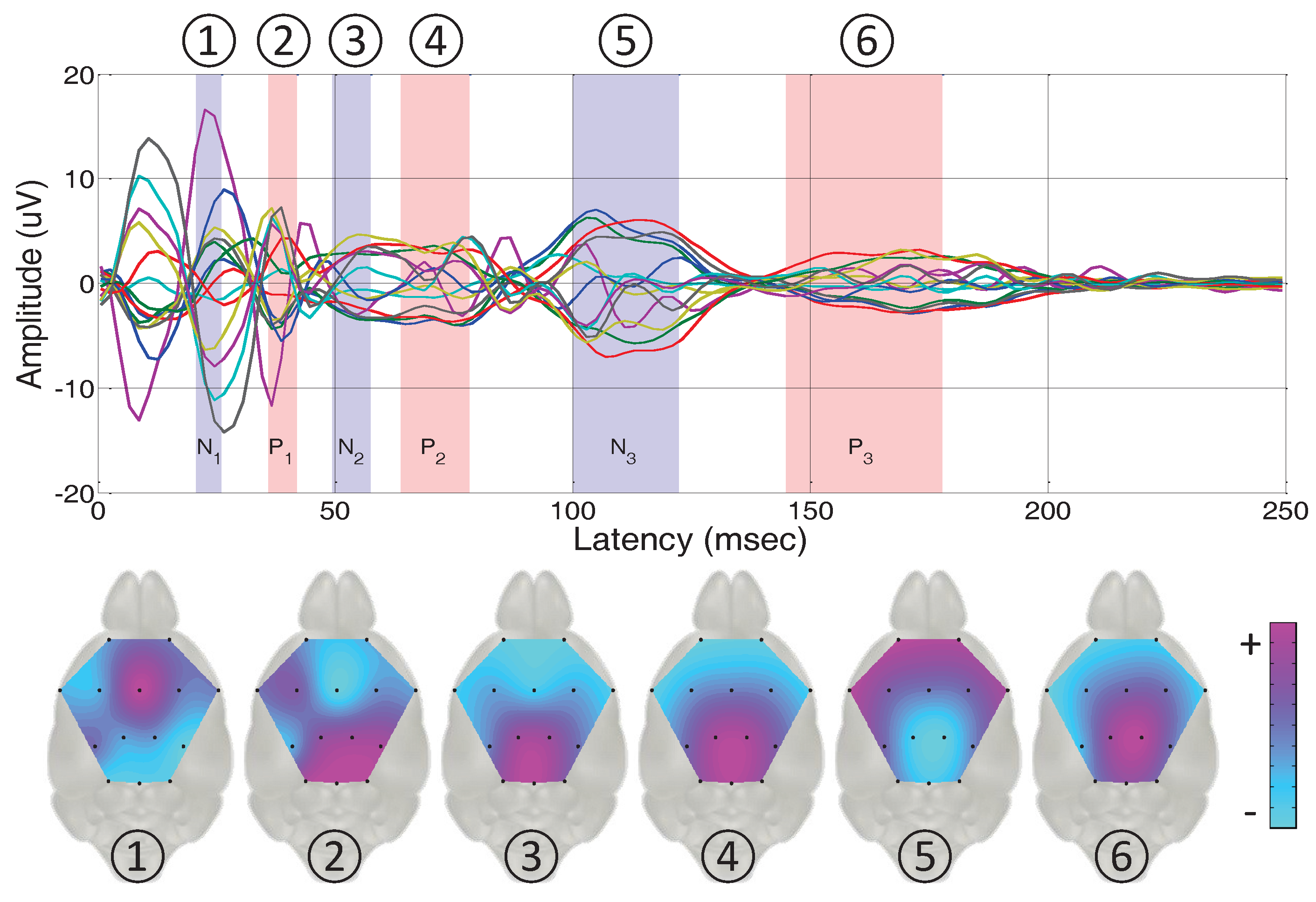
3.2.2. Topographical Analysis Based on the Peak Latencies
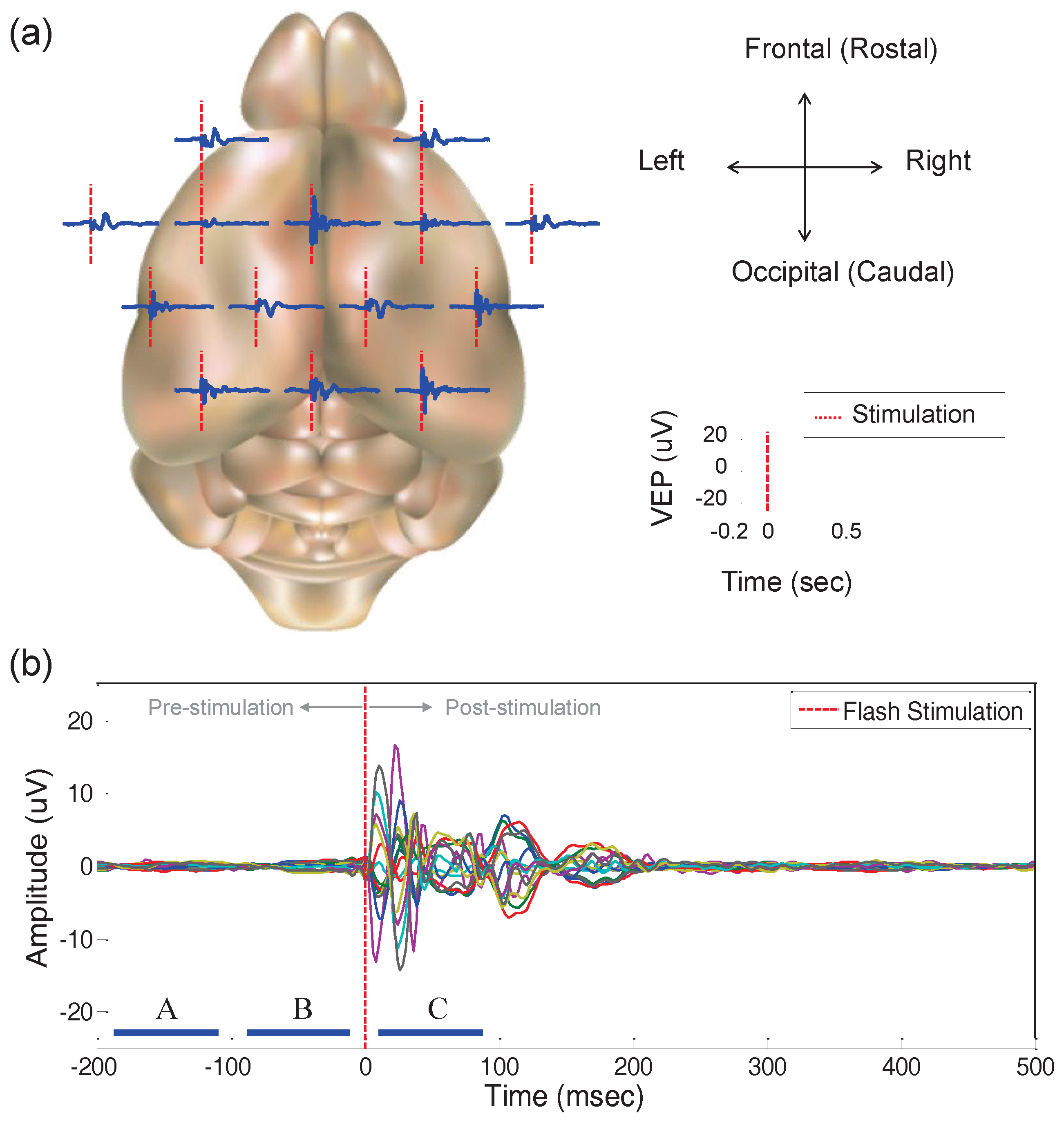
3.2.3. Multi-Channel VEP Responses in Spatio-Temporal Aspects
4. Analysis of VEP Responses in the Time-Frequency Domain
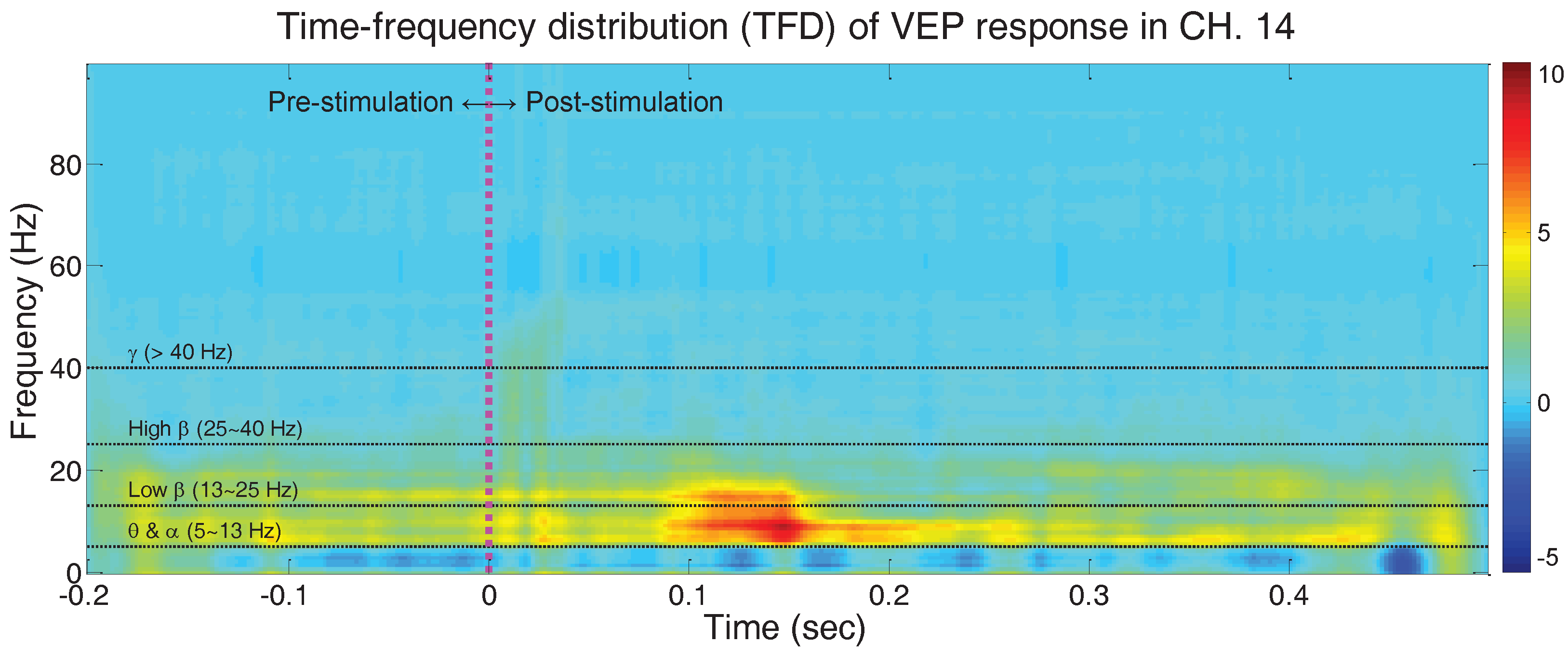
4.1. Time-Frequency Distribution (TFD) Analysis
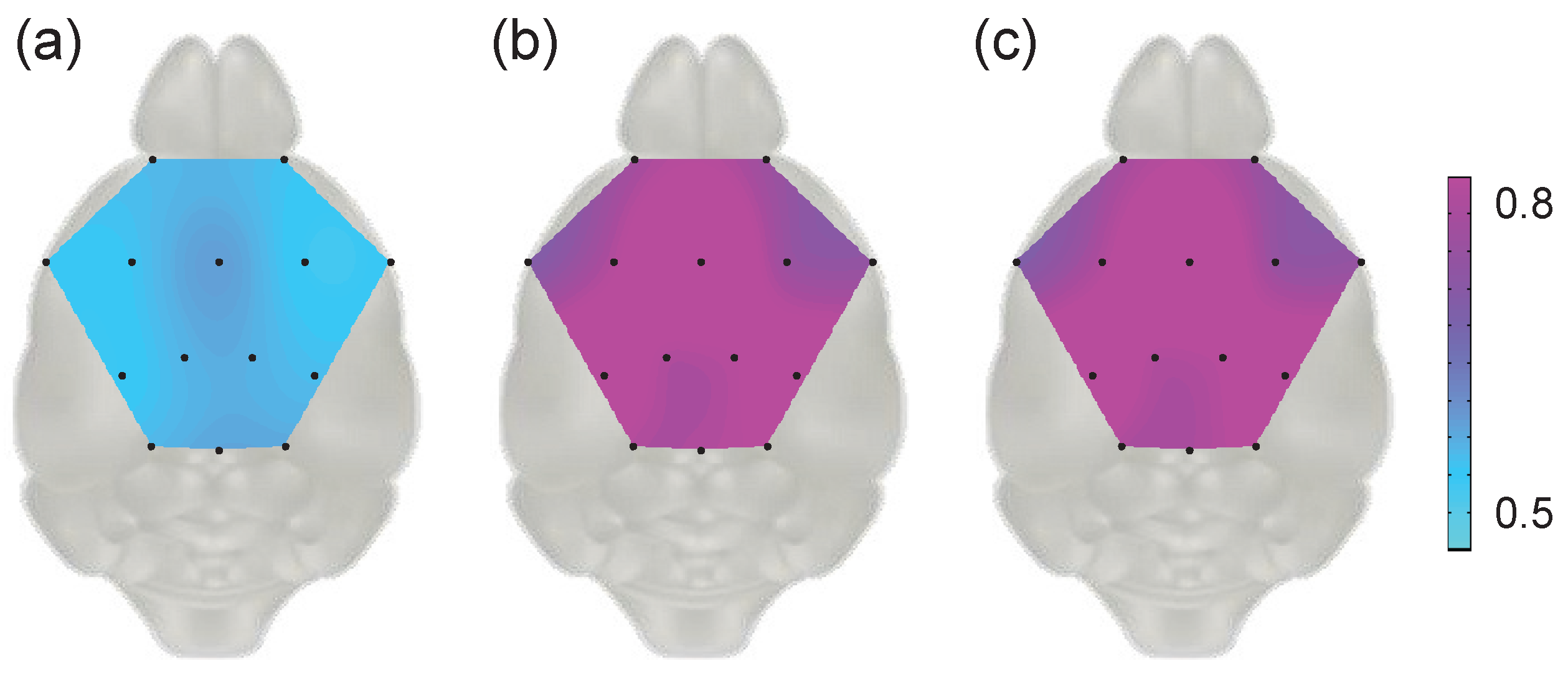
4.2. Topographical Analysis Using TFD-Based Distance
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| BJD | Born–Jordan distribution |
| CAR | Common average reference |
| EEG | Electroencephalography |
| ERP | Event related potential |
| RID | Reduced interference distribution |
| ROI | Region of interest |
| SE | Standard error |
| TFD | Time-frequency distribution |
| VEP | Visual evoked potentia |
References
- Roelfsema, P.R.; Treue, S. Basic neuroscience research with nonhuman primates: A small but indispensable component of biomedical research. Neuron 2014, 82, 1200–1204. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Wais, M.; Sheppya, E.; Campob, M. D.; Zhanga, L. A glue-based, screw-free method for implantation of intra-cranial electrodes in young mice. J. Neurosci. Methods 2008, 171, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Mègevand, P.; Quairiaux, C.; Lascano, A.M.; Kiss, J.Z.; Michel, C.M. A mouse model for studying large-scale neuronal networks using EEG mapping techniques. NeuroImage 2008, 42, 591–602. [Google Scholar] [CrossRef] [PubMed]
- Daszuta, A.; Gambarelli, F. Early postnatal development of EEG and sleep-waking cycle in two inbred mouse strains. Brain Res. 1985, 354, 39–47. [Google Scholar] [CrossRef]
- Quairiaux, C.; Mègevand, P.; Kiss, J.Z.; Michel, C.M. Functional development of large-scale sensorimotor cortical networks in the brain. J. Neurosci. 2011, 31, 9574–9584. [Google Scholar] [CrossRef] [PubMed]
- Wisor, J.P.; Edgar, D.M.; Yesavage, J.; Ryan, H.S.; McCormick, C.M.; Lapustea, N.; Murphy, G.M. Sleep and circadian abnormalities in a transgenic mouse model of Alzheimer’s disease: A role for cholinergic transmission. Neuroscience 2005, 131, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Bergstrom, R.A.; Choi, J.H.; Manduca, A.; Shin, H.S.; Worrell, G.A.; Howe, C.L. Automated identification of multiple seizure-related and interictal epileptiform event types in the EEG of mice. Sci. Rep. 2013, 3, 1483. [Google Scholar] [CrossRef] [PubMed]
- Frank, H.Y.; Mantegazza, M.; Westenbroek, R.E.; Robbins, C.A.; Kalume, F.; Burton, K.A.; Spain, W.J.; McKnight, G.S.; Scheuer, T.; Catterall, W.A. Reduced sodium current in GABAergic interneurons in a mouse model of severe myoclonic epilepsy in infancy. Nat. Neurosci. 2006, 9, 1142–1149. [Google Scholar]
- Parmentier, R.; Ohtsu, H.; Djebbara-Hannas, Z.; Valatx, J.L.; Watanabe, T.; Lin, J.S. Anatomical, physiological, and pharmacological characteristics of histidine decarboxylase knock-out mice: Evidence for the role of brain histamine in behavioral and sleep-wake control. J. Neurosci. 2002, 22, 7695–7711. [Google Scholar] [PubMed]
- Handforth, A.; DeLorey, T.M.; Homanics, G.E.; Olsen, R.W. Pharmacologic Evidence for Abnormal Thalamocortical Functioning in GABAA Receptor β3 Subunit-Deficient Mice, a Model of Angelman Syndrome. Epilepsia 2005, 46, 1860–1870. [Google Scholar] [CrossRef] [PubMed]
- Buzsáki, G.; Anastassiou, C.A.; Koch, C. The origin of extracellular fields and currents—EEG, ECoG, LFP and spikes. Nat. Rev. Neurosci. 2012, 13, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Wais, M.; Sheppya, E.; Campob, M.D.; Zhanga, L. An improved screw-free method for electrode implantation and intracranial electroencephalographic recordings in mice. Behav. Res. Methods 2009, 41, 736–741. [Google Scholar] [CrossRef] [PubMed]
- Jeantet, Y.; Cho, Y.H. Design of a twin tetrode microdrive and headstage for hippocampal single unit recordings in behaving mice. J. Neurosci. Methods 2003, 129, 129–134. [Google Scholar] [CrossRef]
- Buzsáki, G.; Buhl, D.L.; Harris, K.D.; Csicsvari, J.; Czeh, B.; Morozov, A. Hippocampal network patterns of activity in the mouse. Neuroscience 2003, 116, 201–211. [Google Scholar] [CrossRef]
- Korshunov, V.A. Miniature microdrive–headstage assembly for extracellular recording of neuronal activity with high-impedance electrodes in freely moving mice. J. Neurosci. Methods 2006, 158, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Hollenberg, B.A.; Richards, C.D.; Richards, R.; Bahr, D.F.; Rector, D.M. A MEMS fabricated flexible electrode array for recording surface field potentials. J. Neurosci. Methods 2006, 153, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Pinto-Duarte, A.; Behrens, M.M.; Zhou, X.; Sejnowski, T.J. Characterization of spatio-temporal epidural event-related potentials for mouse models of psychiatric disorders. Sci. Rep. 2015, 5, 14964. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Song, I.; Lee, S.M.; Choi, H.S.; Byeon, H.; Kim, I.; Lee, S.H. An electroplating-free and minimal noise polyimide microelectrode for recording auditory evoked potentials from the epicranius. IEEE Trans. Biomed. Eng. 2013, 60, 3425–3431. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Kim, D.; Shin, H.S.; Sung, H.G.; Choi, J.H. High-density EEG recordings of the freely moving mice using polyimide-based microelectrode. J. Vis. Exp. 2011, 47, e2562. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Koch, K.P.; Poppendieck, W.; Lee, M.; Shin, H.S. High resolution electroencephalography in freely moving mice. J. Neurophysiol. 2010, 104, 1825–1834. [Google Scholar] [CrossRef] [PubMed]
- Baek, D.H.; Moon, J.H.; Choi, Y.Y.; Lee, M.; Choi, J.H.; Pak, J.J.; Lee, S.H. A dry release of polyimide electrodes using Kapton film and application to EEG signal measurements. Microsyst. Technol. 2011, 17, 7–14. [Google Scholar] [CrossRef]
- Troncoso, E.; Muller, D.; Czellar, S.; Kiss, J.Z. Epicranial sensory evoked potential recordings for repeated assessment of cortical functions in mice. J. Neurosci. Methods 2000, 97, 51–58. [Google Scholar] [CrossRef]
- Garner, J.P. The significance of meaning: why do over 90% of behavioral neuroscience results fail to translate to humans, and what can we do to fix it? ILAR J. 2014, 55, 438–456. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Gordo, M.A.; Sanchez-Morillo, D.; Pelayo Valle, F. Dry EEG electrodes. Sensors 2014, 14, 12847–12870. [Google Scholar] [CrossRef] [PubMed]
- Liao, L.D.; Wang, I.J.; Chen, S.F.; Chang, J.Y.; Lin, C.T. Design, fabrication and experimental validation of a novel dry-contact sensor for measuring electroencephalography signals without skin preparation. Sensors 2011, 11, 5819–5834. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Yeon, C.; Chung, E.; Kim, K. A non-invasive flexible multi-channel electrode for in vivo mouse EEG recoding. In Proceedings of the IEEE Sensors Conference, Valencia, Spain, 2–5 November 2014; pp. 1111–1114.
- Franklin, K.B.J.; Paxinos, G. The Mouse Brain in Stereotaxic Coordinates; Academic Press: San Diego, CA, USA, 1997. [Google Scholar]
- Goffin, D.; Brodkin, E.S.; Blendy, J.A.; Siegel, S.J.; Zhou, Z. Cellular origins of auditory event-related potential deficits in Rett syndrome. Nat. Neurosci. 2014, 17, 804–806. [Google Scholar] [CrossRef] [PubMed]
- Connolly, P.M.; Maxwell, C.; Liang, Y.; Kahn, J.B.; Kanes, S.J.; Abel, T.; Gur, R.E.; Turetsky, B.I.; Siegel, S.J. The effects of ketamine vary among inbred mouse strains and mimic schizophrenia for the P80, but not P20 or N40 auditory ERP components. Neurochem. Res. 2004, 29, 1179–1188. [Google Scholar] [CrossRef] [PubMed]
- Ridder, W.H.; Nusinowitz, S. The visual evoked potential in the mouse—Origins and response characteristics. Vis. Res. 2006, 46, 902–913. [Google Scholar] [CrossRef] [PubMed]
- Porciatti, V.; Pizzorusso, T.; Maffei, L. The visual physiology of the wild type mouse determined with pattern VEPs. Vis. Res. 1999, 39, 3071–3081. [Google Scholar] [CrossRef]
- Pizzorusso, T.; Fagiolini, M.; Porciatti, V.; Maffei, L. Temporal aspects of contrast visual evoked potentials in the pigmented rat: Effect of dark rearing. Vis. Res. 1997, 37, 389–395. [Google Scholar] [CrossRef]
- Schalk, G.; McFarland, D.J.; Hinterberger, T.; Birbaumer, N.; Wolpaw, J. BCI2000: A general-purpose brain-computer interface (BCI) system. IEEE Trans. Biomed. Eng. 2004, 51, 1034–1043. [Google Scholar] [CrossRef] [PubMed]
- Strain, G.M.; Tedford, B.L. Flash and pattern reversal visual evoked potentials in C57BL/6J and B6CBAF1/J mice. Brain Res. Bull. 1993, 32, 57–63. [Google Scholar] [CrossRef]
- Lopez, L.; Brusa, A.; Fadda, A.; Loizzo, S.; Martinangeli, A.; Sannita, W.G.; Loizzo, A. Modulation of flash stimulation intensity and frequency: Effects on visual evoked potentials and oscillatory potentials recorded in awake, freely moving mice. Behav. Brain Res. 2002, 131, 105–114. [Google Scholar] [CrossRef]
- Dyer, R.S.; Boyes, W.K. Hypothermia and chloropent anesthesia differentially affect the flash evoked potentials of hooded rats. Brain Res. Bull. 1983, 10, 825–831. [Google Scholar] [CrossRef]
- Hetzler, B.E.; Oaklay, K.E. Dose effects of pentobarbital on evoked potentials in visual cortex and superior colliculus of the albino rat. Neuropharmacology 1981, 20, 969–978. [Google Scholar] [CrossRef]
- Tello, C.; De Moraes, C.G.V.; Prata, T.S.; Derr, P.; Patel, J.; Siegfried, J.; Liebmann, J.M.; Ritch, R. Repeatability of short-duration transient visual evoked potentials in normal subjects. Doc. Ophthalmol. 2010, 120, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Altman, D.G.; Bland, J.M. Standard deviations and standard errors. BMJ 2005, 331, 903. [Google Scholar] [CrossRef] [PubMed]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Cohen, L. Time-frequency distributions—A review. Proc. IEEE 1989, 77, 941–981. [Google Scholar] [CrossRef]
- Jeong, J.; Williams, W. Kernel design for reduced interference distributions. IEEE Trans. Signal Process. 1992, 40, 402–412. [Google Scholar] [CrossRef]
- Aviyente, S.; Brakel, L.A.W.; Kushwaha, R.K.; Snodgrass, M.; Shevrin, H.; Willians, W.J. Characterization of event related potentials using information theoretic distances measures. IEEE Trans. Biomed. Eng. 2004, 51, 737–743. [Google Scholar] [CrossRef] [PubMed]







© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.; Yeon, C.; Kim, K. Development and Experimental Validation of a Dry Non-Invasive Multi-Channel Mouse Scalp EEG Sensor through Visual Evoked Potential Recordings. Sensors 2017, 17, 326. https://doi.org/10.3390/s17020326
Kim D, Yeon C, Kim K. Development and Experimental Validation of a Dry Non-Invasive Multi-Channel Mouse Scalp EEG Sensor through Visual Evoked Potential Recordings. Sensors. 2017; 17(2):326. https://doi.org/10.3390/s17020326
Chicago/Turabian StyleKim, Donghyeon, Chanmi Yeon, and Kiseon Kim. 2017. "Development and Experimental Validation of a Dry Non-Invasive Multi-Channel Mouse Scalp EEG Sensor through Visual Evoked Potential Recordings" Sensors 17, no. 2: 326. https://doi.org/10.3390/s17020326
APA StyleKim, D., Yeon, C., & Kim, K. (2017). Development and Experimental Validation of a Dry Non-Invasive Multi-Channel Mouse Scalp EEG Sensor through Visual Evoked Potential Recordings. Sensors, 17(2), 326. https://doi.org/10.3390/s17020326