
Bioluminescence Assays for Monitoring Chondrogenic Differentiation and Cartilage Regeneration

Abstract
:1. Introduction
2. Fundamentals of Bioluminescence Assays
3. Real-Time Monitoring of Gene Expression
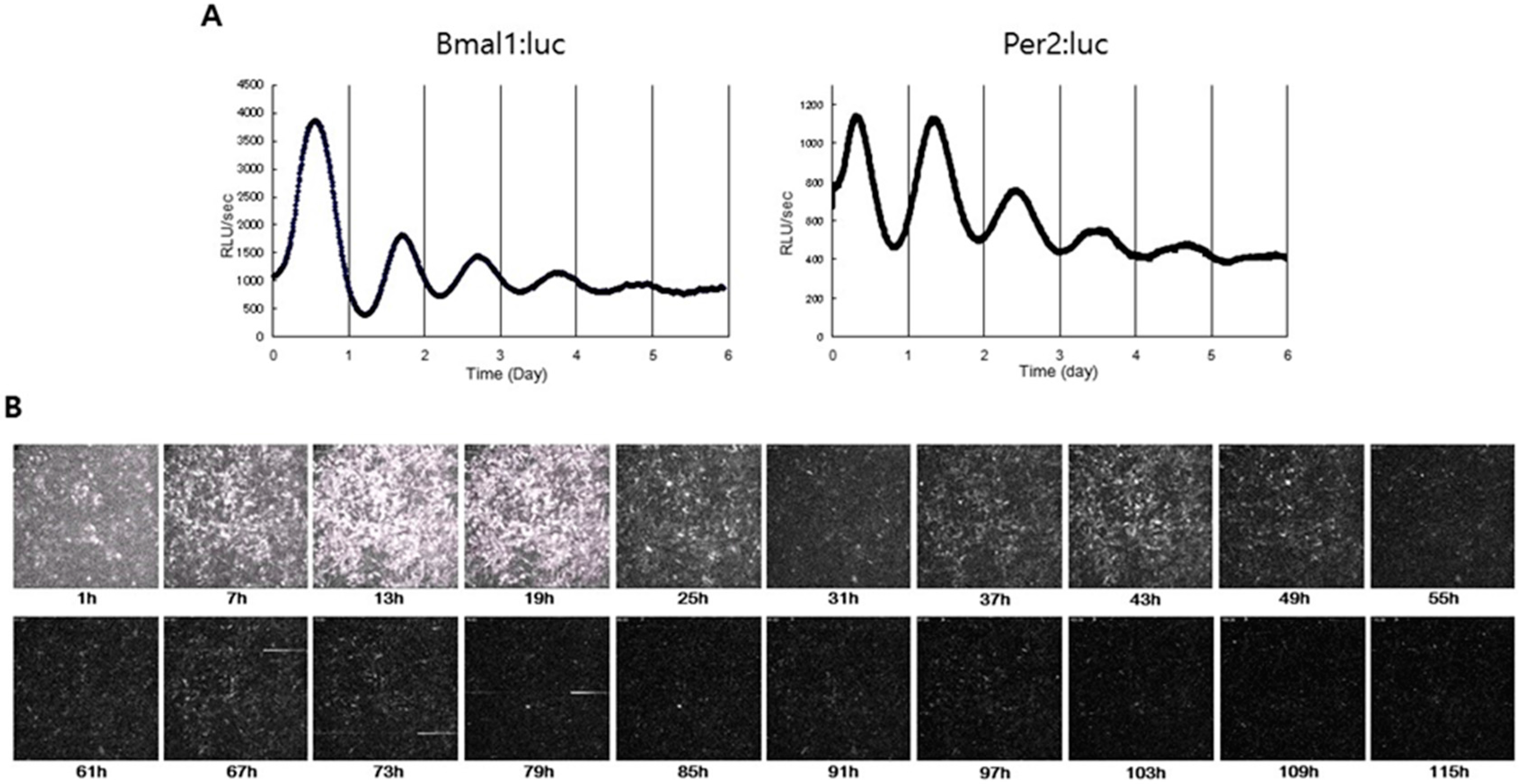
3.1. Principles
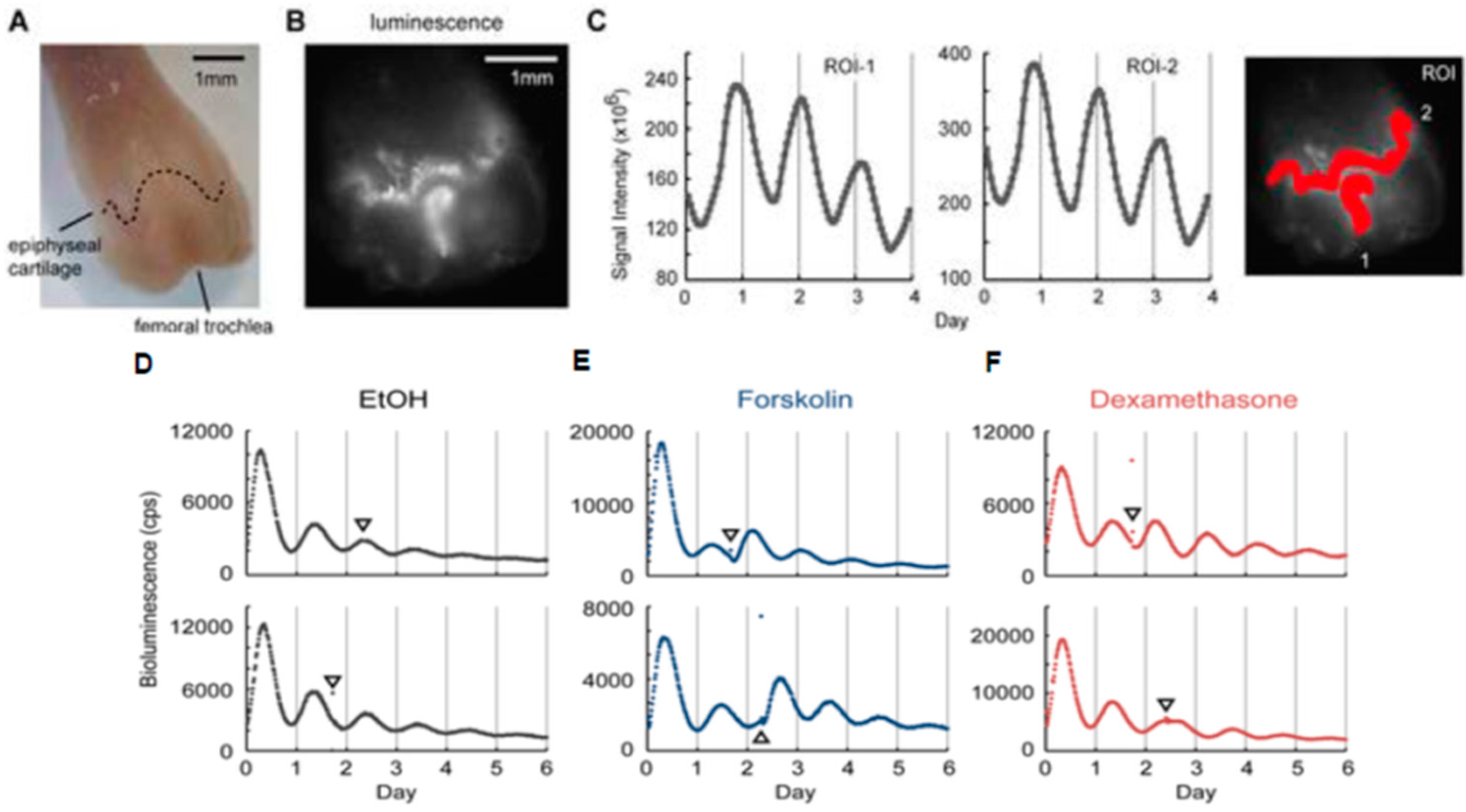
3.2. Application into Cartilage Researches
4. Realtime Monitoring of Biomolecules
4.1. Principles
4.2. Application to Cartilage Research
5. Real-Time Monitoring of Exocytotic Activity
5.1. Principles
5.2. Application to Cartilage Research
6. In Vivo Imaging for Transplanted Cells
6.1. Principles
6.2. Application to Cartilage Research
7. Potential Applications of Bioluminescence Assays for Cartilage Regeneration
8. Conclusions
Acknowledgments
Conflicts of Interest
Abbreviations
| OA | osteoarthritis |
| CCCD | cooled charged coupled device |
| Fluc | Photinus pyralis (firefly) luciferase |
| Rluc | the sea pansy Renilla reniformis luciferase |
| Gluc | the marine copepod Gaussia princeps luciferase |
| PxRe | Phrixothrix hirtus red luciferase |
| Cluc | Cypridina luciferase |
| lux | the bacterial luciferase gene cassette |
| AAV | adeno-associated virus |
| FRET | fluorescence resonance energy transfer |
| BRET | bioluminescence resonance energy transfer |
References
- O’Driscoll, S.W. Current concepts review—The healing and regeneration of articular cartilage. JBJS 1998, 80, 1795–1812. [Google Scholar] [CrossRef]
- Mandelbaum, B.R.; Browne, J.E.; Fu, F.; Micheli, L.; Mosely, J.B.; Erggelet, C.; Minas, T.; Peterson, L. Articular cartilage lesions of the knee. Am. J. Sports Med. 1998, 26, 853–861. [Google Scholar] [PubMed]
- Zhang, J.; Dong, S.; Sivak, W.; Sun, H.B.; Tao, K. Stem cells in cartilage regeneration. Stem Cells Int. 2017, 7034726. [Google Scholar] [CrossRef] [PubMed]
- Massoud, T.F.; Gambhir, S.S. Molecular imaging in living subjects: Seeing fundamental biological processes in a new light. Genes Dev. 2003, 17, 545–580. [Google Scholar] [CrossRef] [PubMed]
- Stephens, D.J.; Allan, V.J. Light microscopy techniques for live cell imaging. Science 2003, 300, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Badr, C.E.; Tannous, B.A. Bioluminescence imaging: Progress and applications. Trends Biotechnol. 2011, 29, 624–633. [Google Scholar] [CrossRef] [PubMed]
- Marques, S.M.; Esteves da Silva, J.C. Firefly bioluminescence: A mechanistic approach of luciferase catalyzed reactions. IUBMB Life 2009, 61, 6–17. [Google Scholar] [CrossRef] [PubMed]
- Greer, L.F.; Szalay, A.A. Imaging of light emission from the expression of luciferases in living cells and organisms: A review. Luminescence 2002, 17, 43–74. [Google Scholar] [CrossRef] [PubMed]
- Prescher, J.A.; Contag, C.H. Guided by the light: Visualizing biomolecular processes in living animals with bioluminescence. Curr. Opin. Chem. Biol. 2010, 14, 80–89. [Google Scholar] [CrossRef] [PubMed]
- LI, J.; Chen, L.; Du, L.; Li, M. Cage the firefly luciferin!—A strategy for developing bioluminescent probes. Chem. Soc. Rev. 2013, 42, 662–676. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, O.; Teranishi, K. Light-emitters involved in the luminescence of coelenterazine. Luminescence 2000, 15, 51–58. [Google Scholar] [CrossRef]
- Badr, C.E.; Hewett, J.W.; Breakefield, X.O.; Tannous, B.A. A highly sensitive assay for monitoring the secretory pathway and ER stress. PLoS ONE 2007, 2, e571. [Google Scholar] [CrossRef] [PubMed]
- Wurdinger, T.; Badr, C.; Pike, L.; De Kleine, R.; Weissleder, R.; Breakefield, X.O.; Tannous, B.A. A secreted luciferase for ex vivo monitoring of in vivo processes. Nat. Methods 2008, 5, 171–173. [Google Scholar] [CrossRef] [PubMed]
- Villalobos, V.; Naik, S.; Bruinsma, M.; Dothager, R.S.; Pan, M.H.; Samrakandi, M.; Moss, B.; Elhammali, A.; Piwnica-Worms, D. Dual-color click beetle luciferase heteroprotein fragment complementation assays. Chem. Biol. 2010, 17, 1018–1029. [Google Scholar] [CrossRef] [PubMed]
- Caysa, H.; Jacob, R.; Müther, N.; Branchini, B.; Messerle, M.; Söling, A. A redshifted codon-optimized firefly luciferase is a sensitive reporter for bioluminescence imaging. Photochem. Photobiol. Sci. 2009, 8, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Branchini, B.R.; Southworth, T.L.; DeAngelis, J.P.; Roda, A.; Michelini, E. Luciferase from the Italian firefly Luciola italica: Molecular cloning and expression. Comp. Biochem. Physiol. Biochem. Mol. Biol. 2006, 145, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, Y.; Kimura, T.; Sugata, K.; Enomoto, T.; Asakawa, A.; Kubota, H.; Ikeda, M.; Ohmiya, Y. Multicolor luciferase assay system: One-step monitoring of multiple gene expressions with a single substrate. Biotechniques 2005, 38, 891–894. [Google Scholar] [CrossRef] [PubMed]
- Close, D.M.; Patterson, S.S.; Ripp, S.; Baek, S.J.; Sanseverino, J.; Sayler, G.S. Autonomous bioluminescent expression of the bacterial luciferase gene cassette (lux) in a mammalian cell line. PLoS ONE 2010, 5, e12441. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, Y.; Ohmiya, Y. Bioluminescence assays: Multicolor luciferase assay, secreted luciferase assay and imaging luciferase assay. Expert Opin. Drug Discov. 2010, 5, 835–849. [Google Scholar] [CrossRef] [PubMed]
- Robertson, J.B.; Johnson, C.H. Luminescence as a continuous real-time reporter of promoter activity in yeast undergoing respiratory oscillations or cell division rhythms. Yeast Genet. Netw. Methods Protoc. 2011, 734, 63–79. [Google Scholar]
- Bronstein, I.; Fortin, J.; Stanley, P.E.; Stewart, G.S.; Kricka, L.J. Chemiluminescent and bioluminescent reporter gene assays. Anal. Biochem. 1994, 219, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, H.; Nakajima, Y.; Ohmiya, Y. Luciferase-YFP fusion tag with enhanced emission for single-cell luminescence imaging. Nat. Methods 2007, 4, 637–639. [Google Scholar] [CrossRef] [PubMed]
- Yasunaga, M.; Nakajima, Y.; Ohmiya, Y. Dual-color bioluminescence imaging assay using green- and red-emitting beetle luciferases at subcellular resolution. Anal. Bioanal. Chem. 2014, 406, 5735–5742. [Google Scholar] [CrossRef] [PubMed]
- Welsh, D.K; Kay, S.A. Bioluminescence imaging in living organisms. Curr. Opin. Biotechnol. 2005, 16, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.; Enomoto, T.; Shimogawara, M.; Yasuda, K.; Nakajima, Y.; Ohmiya, Y. Bioluminescence imaging of dual gene expression at the single-cell level. BioTechniques 2010, 48, 460–462. [Google Scholar] [CrossRef] [PubMed]
- Yoo, SH.; Yamazaki, S.; Lowrey, P.L.; Shimomura, K.; Ko, C.H.; Buhr, E.D.; Siepka, S.M.; Hong, H.K.; Oh, W.J.; Yoo, O.J.; et al. PERIOD2: LUCIFERASE real-time reporting of circadian dynamics reveals persistent circadian oscillations in mouse peripheral tissues. Proc. Natl. Acad. Sci. USA 2004, 101, 5339–5346. [Google Scholar] [CrossRef] [PubMed]
- Gossan, N.; Zeef, L.; Hensman, J.; Hughes, A.; Bateman, J.F.; Rowley, L.; Little, C.B.; Piggins, H.D.; Rattray, M.; Boot-Handford, R.P. The circadian clock in murine chondrocytes regulates genes controlling key aspects of cartilage homeostasis. Arthritis Rheum. 2013, 65, 2334–2345. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, Y.; Yamazaki, T.; Nishii, S.; Noguchi, T.; Hoshino, H.; Niwa, K.; Viviani, V.R.; Ohmiya, Y. Enhanced beetle luciferase for high-resolution bioluminescence imaging. PLoS ONE 2010, 5, e10011. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, T.; Tsuchiya, Y.; Okubo, N.; Kunimoto, T.; Minami, Y.; Fujiwara, H.; Umemura, Y.; Koike, N.; Kubo, T; Yagita, K. Robust Circadian Rhythm and Parathyroid Hormone-Induced Resetting during Hypertrophic Differentiation in ATDC5 Chondroprogenitor Cells. Acta Histochem. Cytochem. 2015, 48, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Okubo, N.; Minami, Y.; Fujiwara, H.; Umemura, Y.; Tsuchiya, Y.; Shirai, T.; Oda, R.; Inokawa, H.; Kubo, T.; Yagita, K. Prolonged bioluminescence monitoring in mouse ex vivo bone culture revealed persistent circadian rhythms in articular cartilages and growth plates. PLoS ONE 2013, 8, e78306. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, K.; Sandell, L.J. Extracellular matrix gene regulation. Clin. Orthop. Relat. Res. 2004, 427, S123–S128. [Google Scholar] [CrossRef]
- Lefebvre, V.; Huang, W.; Harley, V.R.; Goodfellow, P.N.; De Crombrugghe, B. SOX9 is a potent activator of the chondrocyte-specific enhancer of the pro alpha1 (II) collagen gene. Mol. Cell. Biol. 1997, 17, 2336–2346. [Google Scholar] [CrossRef] [PubMed]
- Vilalta, M.; Jorgensen, C.; Dégano, I.R.; Chernajovsky, Y.; Gould, D.; Noël, D.; Andrades, J.A.; Becerra, J.; Rubio, N.; Blanco, J. Dual luciferase labelling for non-invasive bioluminescence imaging of mesenchymal stromal cell chondrogenic differentiation in demineralized bone matrix scaffolds. Biomaterials 2009, 30, 4986–4995. [Google Scholar] [CrossRef] [PubMed]
- Mailhiot, S.E.; Zignego, D.L.; Prigge, J.R.; Wardwell, E.R.; Schmidt, E.E.; June, R.K. Non-invasive quantification of cartilage using a novel in vivo bioluminescent reporter mouse. PLoS ONE 2015, 10, e0130564. [Google Scholar] [CrossRef] [PubMed]
- Bridger, W.A.; Henderson, J.F. Cell ATP; John Wiley & Sons: Hoboken, NJ, USA, 1983. [Google Scholar]
- Gordon, J.L. Extracellular ATP: Effects, sources and fate. Biochem. J. 1986, 233, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Del Puerto, A.; Wandosell, F.G.; Garrido, J.J. Neuronal and glial purinergic receptors functions in neuron development and brain disease. Front. Cell. Neurosci. 2013, 7, 197. [Google Scholar] [CrossRef] [PubMed]
- Coddou, C.; Yan, Z.; Obsil, T.; Huidobro-Toro, J.P.; Stojilkovic, S.S. Activation and regulation of purinergic P2X receptor channels. Pharmacol. Rev. 2011, 63, 641–683. [Google Scholar] [CrossRef] [PubMed]
- Lundin, A. Use of firefly luciferase in ATP-related assays of biomass, enzymes, and metabolites. Methods Enzymol. 2000, 305, 346–370. [Google Scholar] [PubMed]
- Crouch, S.P.M.; Kozlowski, R.; Slater, K.J.; Fletcher, J. The use of ATP bioluminescence as a measure of cell proliferation and cytotoxicity. J. Immunol. Methods 1993, 160, 81–88. [Google Scholar] [CrossRef]
- Dexter, S.J.; Cámara, M.; Davies, M.; Shakesheff, K.M. Development of a bioluminescent ATP assay to quantify mammalian and bacterial cell number from a mixed population. Biomaterials 2003, 24, 27–34. [Google Scholar] [CrossRef]
- Ainscow, E.K.; Rutter, G.A. Glucose-stimulated oscillations in free cytosolic ATP concentration imaged in single islet β-cells. Diabetes 2002, 51, S162–S170. [Google Scholar] [CrossRef]
- Koop, A.; Cobbold, P.H. Continuous bioluminescent monitoring of cytoplasmic ATP in single isolated rat hepatocytes during metabolic poisoning. Biochem. J. 1993, 295, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Turner, D.E.; Daugherity, E.K.; Altier, C.; Maurer, K.J. Efficacy and limitations of an ATP-based monitoring system. J. Am. Assoc. Lab. Anim. Sci. 2010, 49, 190–195. [Google Scholar] [PubMed]
- Clapham, D.E. Calcium signaling. Cell 2007, 131, 1047–1058. [Google Scholar] [CrossRef] [PubMed]
- Rizzuto, R.; Pozzan, T. Microdomains of intracellular Ca2+: Molecular determinants and functional consequences. Physiol. Rev. 2006, 86, 369–408. [Google Scholar] [CrossRef] [PubMed]
- Llinás, R.; Blinks, JR.; Nicholson, C. Calcium transient in presynaptic terminal of squid giant synapse: Detection with aequorin. Science 1972, 176, 1127–1129. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, O. Bioluminescence in the sea: Photoprotein systems. Symp. Soc. Exp. Biol. 1985, 39, 351–372. [Google Scholar] [PubMed]
- Tsuji, F.I.; Ohmiya, Y.; Fagan, T.F.; Toh, H.; Inouye, S. Molecular evolution of the Ca2+ binding photoproteins of the hydrozoa. Photochem. Photobiol. 1995, 62, 657–661. [Google Scholar] [CrossRef] [PubMed]
- Hofer, A.M.; Machen, T.E. Technique for in situ measurement of calcium in intracellular inositol 1,4,5-trisphosphate-sensitive stores using the fluorescent indicator mag-fura-2. Proc. Natl. Acad. Sci. USA 1993, 90, 2598–2602. [Google Scholar] [CrossRef] [PubMed]
- Inouye, S.; Aoyama, S.; Miyata, T.; Tsuji, F.I.; Sakaki, Y. Overexpression and purification of the recombinant Ca2+-binding protein, apoaequorin. J. Biochem. 1989, 105, 473–477. [Google Scholar] [CrossRef] [PubMed]
- Allen, D.G.; Prendergast, F.G. Aequorin luminescence: Relation of light emission to calcium concentration—A calcium-independent component. Science 1977, 195, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Rizzuto, R.; Simpson, A.W.; Brini, M.; Pozzan, T. Rapid changes of mitochondrial Ca2+ revealed by specifically targeted recombinant aequorin. Nature 1992, 358, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Montero, M.; Brini, M.; Marsault, R.; Alvarez, J.; Sitia, R.; Pozzan, T.; Rizzuto, R. Monitoring dynamic changes in free Ca2+ concentration in the endoplasmic reticulum of intact cells. EMBO J. 1995, 14, 5467–5475. [Google Scholar] [PubMed]
- Brini, M.; Murgia, M.; Pasti, L.; Picard, D.; Pozzan, T.; Rizzuto, R. Nuclear Ca2+ concentration measured with specifically targeted recombinant aequorin. EMBO J. 1993, 12, 4813–4819. [Google Scholar] [PubMed]
- Rodriguez-Garcia, A.; Rojo-Ruiz, J.; Navas-Navarro, P.; Aulestia, F.J.; Gallego-Sandin, S.; Garcia-Sancho, J.; Alonso, M.T. GAP, an aequorin-based fluorescent indicator for imaging Ca2+ in organelles. Proc. Natl. Acad. Sci. USA 2014, 111, 2584–2589. [Google Scholar] [CrossRef] [PubMed]
- McKnight, G.S. Cyclic AMP second messenger systems. Curr. Opin. Cell Biol. 1991, 3, 213–217. [Google Scholar] [CrossRef]
- Takeuchi, M.; Nagaoka, Y.; Yamada, T.; Takakura, H.; Ozawa, T. Ratiometric bioluminescence indicators for monitoring cyclic adenosine 3′,5′-monophosphate in live cells based on luciferase-fragment complementation. Anal. Chem. 2010, 82, 9306–9313. [Google Scholar] [CrossRef] [PubMed]
- Fan, F.; Binkowski, B.F.; Butler, B.L.; Stecha, P.F.; Lewis, M.K.; Wood, K.V. Novel genetically encoded biosensors using firefly luciferase. ACS Chem. Biol. 2008, 3, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Binkowski, B.F.; Butler, B.L.; Stecha, P.F.; Eggers, C.T.; Otto, P.; Zimmerman, K.; Vidugiris, G.; Wood, M.G.; Encell, L.P.; Fan, F.; et al. A luminescent biosensor with increased dynamic range for intracellular cAMP. ACS Chem. Biol. 2011, 6, 1193–1197. [Google Scholar] [CrossRef] [PubMed]
- Olsen, B.R.; Reginato, A.M.; Wang, W. Bone development. Annu. Rev. Cell Dev. Biol. 2000, 16, 191–220. [Google Scholar] [CrossRef] [PubMed]
- Mariani, F.V.; Martin, G.R. Deciphering skeletal patterning: Clues from the limb. Nature 2003, 423, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Martin, T.F. Stages of regulated exocytosis. Trends Cell Biol. 1997, 7, 271–276. [Google Scholar] [CrossRef]
- Jones, P.M.; Persaud, S.J. Protein Kinases, protein Phosphorylation, and the regulation of insulin secretion from pancreatic β-Cells. Endocr. Rev 1998, 19, 429–461. [Google Scholar] [PubMed]
- Kwon, H.J.; Ohmiya, Y.; Honma, K.I.; Honma, S.; Nagai, T.; Saito, K.; Yasuda, K. Synchronized ATP oscillations have a critical role in prechondrogenic condensation during chondrogenesis. Cell Death Dis. 2012, 3, e278. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.J.; Ohmiya, Y.; Yasuda, K. Simultaneous monitoring of intracellular ATP and oxygen levels in chondrogenic differentiation using a dual-color bioluminescence reporter. Luminescence 2014, 29, 1194–1198. [Google Scholar] [CrossRef] [PubMed]
- Herst, P.M.; Tan, A.S.; Scarlett, D.J.G.; Berridge, M.V. Cell surface oxygen consumption by mitochondrial gene knockout cells. BBA Bioenerg. 2004, 1656, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.J.; Ohmiya, Y.; Yasuda, K. Dual-color system for simultaneously monitoring intracellular Ca2+ and ATP dynamics. Anal. Biochem. 2012, 430, 45–47. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.K.; Kauri, L.M.; Qian, W.J.; Kennedy, R.T. Correlated oscillations in glucose consumption, oxygen consumption, and intracellular free Ca2+ in single islets of Langerhans. J. Biol. Chem. 2000, 275, 6642–6650. [Google Scholar] [CrossRef] [PubMed]
- Magnus, G.; Keizer, J. Model of β-cell mitochondrial calcium handling and electrical activity. II. Mitochondrial variables. Am. J. Physiol. Cell Physiol. 1998, 274, C1174–C1184. [Google Scholar]
- Detimary, P.; Gilon, P.; Henquin, J.C. Interplay between cytoplasmic Ca2+ and the ATP/ADP ratio: A feedback control mechanism in mouse pancreatic islets. Biochem. J. 1998, 333, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Burchfield, J.G.; Lopez, J.A.; Mele, K.; Vallotton, P.; Hughes, W.E. Exocytotic vesicle behaviour assessed by total internal reflection fluorescence microscopy. Traffic 2010, 11, 429–439. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Kasai, H. Exocytic process analyzed with two-photon excitation imaging in endocrine pancreas. Endocr. J. 2007, 54, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Inouye, S.; Ohmiya, Y.; Toya, Y.; Tsuji, F.I. Imaging of luciferase secretion from transformed Chinese hamster ovary cells. Proc. Natl. Acad. Sci. USA 1992, 89, 9584–9587. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Usuda, S.; Ichinose, H.; Inouye, S. Real-time bioluminescence imaging of a protein secretory pathway in living mammalian cells using Gaussia luciferase. FEBS Lett. 2007, 581, 4551–4556. [Google Scholar] [CrossRef]
- Miesenböck, G.; Rothman, J.E. Patterns of synaptic activity in neural networks recorded by light emission from synaptolucins. Proc. Natl. Acad. Sci. USA 1997, 94, 3402–3407. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.J.; Han, Y. Dual monitoring of secretion and ATP levels during chondrogenesis using perfusion culture-combined bioluminescence monitoring system. BioMed Res. Int. 2015, 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.A.; Wagers, A.J.; Beilhack, A.; Dusich, J.; Bachmann, M.H.; Negrin, R.S.; Weissman, I.L.; Contag, C.H. Shifting foci of hematopoiesis during reconstitution from single stem cells. Proc. Natl. Acad. Sci. USA 2004, 101, 221–226. [Google Scholar] [CrossRef]
- Van der Bogt, K.E.; Sheikh, A.Y.; Schrepfer, S.; Hoyt, G.; Cao, F.; Ransohoff, K.J.; Swijnenburg, R.J.; Pearl, J.; Lee, A.; Fischbein, M.; et al. Comparison of different adult stem cell types for treatment of myocardial ischemia. Circulation 2008, 118, S121–S129. [Google Scholar] [CrossRef] [PubMed]
- Van der Bogt, K.E.; Schrepfer, S.; Yu, J.; Sheikh, A.Y.; Hoyt, G.; Govaert, J.A.; Velotta, J.B.; Contag, C.H.; Robbins, R.C.; Wu, J.C. Comparison of transplantation of adipose tissue-and bone marrow-derived mesenchymal stem cells in the infarcted heart. Transplantation 2009, 87, 642. [Google Scholar] [CrossRef] [PubMed]
- Takaku, Y.; Murai, K.; Ukai, T.; Ito, S.; Kokubo, M.; Satoh, M.; Kobayashi, E.; Yamato, M.; Okano, T.; Takeuchi, M.; et al. In vivo cell tracking by bioluminescence imaging after transplantation of bioengineered cell sheets to the knee joint. Biomaterials 2014, 35, 2199–2206. [Google Scholar] [CrossRef] [PubMed]
- Payne, K.A.; Lee, H.H.; Haleem, A.M.; Martins, C.; Yuan, Z.; Qiao, C.; Xiao, X.; Chu, C.R. Single intra-articular injection of adeno-associated virus results in stable and controllable in vivo transgene expression in normal rat knees. Osteoarthr. Cartil. 2011, 19, 1058–1065. [Google Scholar] [CrossRef] [PubMed]
- Mendelsohn, A.R.; Brent, R. Protein interaction methods--toward an endgame. Science 1999, 284, 1948. [Google Scholar] [CrossRef] [PubMed]
- Pfleger, K.D.; Eidne, K.A. Illuminating insights into protein-protein interactions using bioluminescence resonance energy transfer (BRET). Nat. Methods 2006, 3, 165–174. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Advantage | Disadvantage | |
|---|---|---|
| Fluorescence | Superior brightness | Requirement of excitation light |
| Shorter imaging time (miliseconds) | High phototoxicity | |
| High photobleaching | ||
| High background | ||
| Autofluorescence | ||
| Autofluorescence | ||
| Bioluminescence | Low phototoxicity | Low brightness |
| Low background | Longer imaging time (minute) | |
| High sensitivity | Requirement of substrates | |
| Broad linear range |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Je, H.J.; Kim, M.G.; Kwon, H.J. Bioluminescence Assays for Monitoring Chondrogenic Differentiation and Cartilage Regeneration. Sensors 2017, 17, 1306. https://doi.org/10.3390/s17061306
Je HJ, Kim MG, Kwon HJ. Bioluminescence Assays for Monitoring Chondrogenic Differentiation and Cartilage Regeneration. Sensors. 2017; 17(6):1306. https://doi.org/10.3390/s17061306
Chicago/Turabian StyleJe, Hyeon Jeong, Min Gu Kim, and Hyuck Joon Kwon. 2017. "Bioluminescence Assays for Monitoring Chondrogenic Differentiation and Cartilage Regeneration" Sensors 17, no. 6: 1306. https://doi.org/10.3390/s17061306
APA StyleJe, H. J., Kim, M. G., & Kwon, H. J. (2017). Bioluminescence Assays for Monitoring Chondrogenic Differentiation and Cartilage Regeneration. Sensors, 17(6), 1306. https://doi.org/10.3390/s17061306