Tuning the Sensitivity of the PDR5 Promoter-Based Detection of Diclofenac in Yeast Biosensors

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains
2.2. Growth Media and Culture Conditions
2.3. Plasmid Construction and Manipulation
2.4. Response of the Yeast Cell Sensors to Diclofenac
2.5. Databases
3. Results
3.1. TGFP Reporter Constructs with the Native 5′URS of PDR5 Allow Detection of up to 1 μM Diclofenac
3.2. Diclofenac-Mediated Induction of PDR5 Depends on the Transcription Factors Pdr1p and Pdr3p, the Master Regulators of the Pleiotropic Drug Response
3.3. The Number of Pleiotropic Drug Response Elements (PDRE) in the WT 5′URS of the PDR5 Gene Influences the Response to Diclofenac
3.4. The Promoter Sequence −306 to −375 Harbours Binding Sites for Mot3p, Pdr8p, Stb5p and Yrr1p
3.4.1. Role of Mot3p for the Response of the PDR5 Promoter Sequence −306 to −375 to Diclofenac
3.4.2. In the Absence of Stb5p the Response to Diclofenac Is Delayed
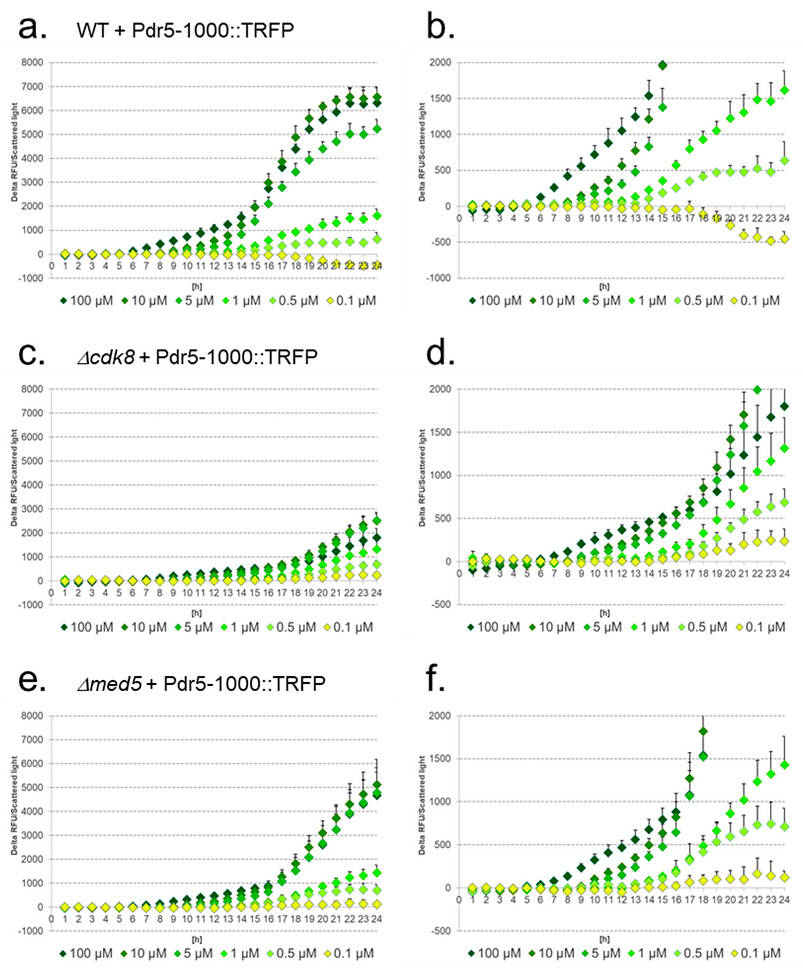
3.5. Deletion of Mediator Subunits Differentially Influence the Response Strength to Diclofenac
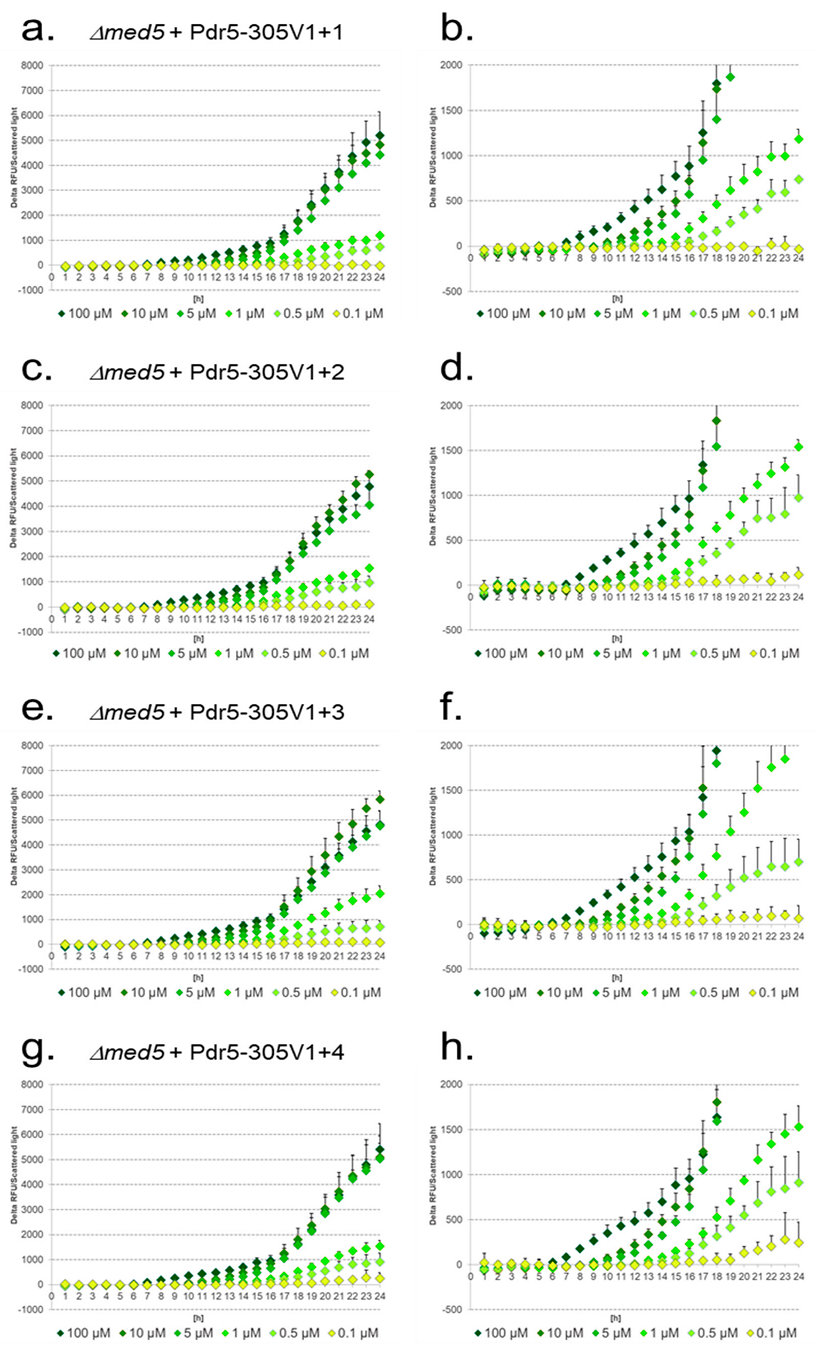
3.6. Design of Artificial Promoter Constructs with Additional PDREs
3.7. Response of Native and Artificial Promoter Constructs in WT, Δcdk8, and Δmed5 Strains
4. Discussion
Supplementary Materials
Acknowledgements
Author Contributions
Conflicts of Interest
Abbreviations
| PDR | Pleiotropic Drug Response |
| PDRE | Pleiotropic Drug Response Element |
| TF | Transcription Factor |
| URS | Upstream Regulatory Sequence |
References
- European Parliament, Council of the European Union. Directive 2013/39/EU of the European Parliament and of the Council of 12 August 2013 amending Directives 2000/60/EC and 2008/105/EC as regards priority substances in the field of water policy. Off. J. Eur. Union 2013, L 226, 1–17. [Google Scholar]
- Reemtsma, T.; Weis, S.; Mueller, J.; Petrovic, M.; Gonzáles, S.; Barcelo, D.; Ventura, F.; Knepper, T.P. Polar pollutants entry into the water cycle by municipal wastewater: A european perspective. Environ. Sci. Technol. 2006, 40, 5451–5458. [Google Scholar] [CrossRef] [PubMed]
- Loos, R.; Carvalho, R.; António, D.C.; Comero, S.; Locoro, G.; Tavazzi, S.; Paracchini, B.; Ghiani, M.; Lettier, T.; Blaha, L.; et al. EU-wide monitoring survey on emerging polar organic contaminants in wastewater treatment plant effluents. Water Res. 2013, 47, 6475–6487. [Google Scholar] [CrossRef] [PubMed]
- Heberer, H. Occurrence, fate, and removal of pharmaceutical residues in the aquatic environment: A review of recent research data. Toxicol. Lett. 2002, 131, 5–17. [Google Scholar] [CrossRef]
- Clara, M.; Strenn, B.; Gans, O.; Martinez, E.; Kreuzinger, N.; Kroiss, H. Removal of selected pharmaceuticals, fragrances and endocrine disrupting compounds in a membrane bioreactor and conventional wastewater treatment plants. Water Res. 2005, 39, 4797–4807. [Google Scholar] [CrossRef] [PubMed]
- Zwiener, C.; Frimmel, F.H. Short-term tests with a pilot sewage plant and biofilm reactors for the biological degradation of the pharmaceutical compounds clofibric acid, ibuprofen, and diclofenac. Sci. Total Environ. 2003, 309, 201–211. [Google Scholar] [CrossRef]
- Adeniran, A.; Sherer, M.; Tyo, K.E.J. Yeast-based biosensors: Design and applications. FEMS Yeast Res. 2015, 15, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, J.S.; Orij, R.; Luttik, M.A.H.; Smits, G.J.; Vermeulen, N.P.E.; Vos, J.C. Subunits Rip1p and Cox9p of the respiratory chain contribute to diclofenac-induced mitochondrial Dysfunction. Microbiology 2011, 157, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, J.S.; Vermeulen, N.P.E.; Vos, J.C. Involvement of the Pleiotropic Drug Resistance Response, Protein Kinase C Signaling, and Altered Zinc Homeostasis in Resistance of Saccharomyces cerevisiae to diclofenac. Appl. Environ. Microbiol. 2011, 77, 5973–5980. [Google Scholar] [CrossRef] [PubMed]
- Katzmann, D.J.; Burnett, P.B.; Golin, J.; Mahe, Y.; Moye-Rowley, W.S. Transcriptional Control of the Yeast PDR5 Gene by the PDR3 Gene Product. Mol. Cell. Biol. 1994, 14, 4653–4661. [Google Scholar] [CrossRef] [PubMed]
- Katzmann, D.J.; Hallstrom, T.C.; Mahe, Y.; Moye-Rowley, W.S. Multiple Pdr1p/Pdr3p binding sites are essential for normal expression of the ATP binding cassette transporter protein-encoding Gene PDR5. J. Biol. Chem. 1996, 271, 23049–23054. [Google Scholar] [CrossRef] [PubMed]
- Balzi, E.; Wang, M.; Leterme, S.; Van Dyck, L.; Goffe, A. PDR5, a Novel Yeast multidrug resistance conferring transporter controlled by the transcription regulator PDR1. J. Biol. Chem. 1994, 269, 2206–2214. [Google Scholar] [PubMed]
- MacPherson, S.; Larochelle, M.; Turcotte, B. A Fungal Family of Transcriptional Regulators: The Zinc Cluster Proteins. Microbiol. Mol. Biol. Rev 2006, 70, 583–604. [Google Scholar] [CrossRef] [PubMed]
- Thakur, J.K.; Arhanari, H.; Yang, F.; Pan, S.J.; Fan, X.; Breger, J.; Frueh, D.; Gulshan, K.; Li, D.K.; Mylonakis, E.; et al. A nuclear receptor-like pathway regulating multidrug resistance in fungi. Nature 2008, 452, 604–609. [Google Scholar] [CrossRef] [PubMed]
- Mumberg, D.; Muller, R.; Funk, M. Yeast vectors for the controlled expression of heterologous proteins in different genetic backgrounds. Gene 1995, 156, 119–122. [Google Scholar] [CrossRef]
- Ausubel, F.M.; Brent, R.; Kingston, R.E.; Moore, D.D.; Seidman, J.G.; Smith, J.A.; Struhl, K. (Eds.) Current Protocols in Molecular Biology; Wiley: New York, NY, USA, 1987. [Google Scholar]
- Saccharomyces Genome Database. Available online: http://www.yeastgenome.org/ (accessed on 25 June 2017).
- YEASTRACT. Available online: http://www.yeastract.com/ (accessed on 25 June 2017).
- YPA (Yeast Promoter Atlas). Available online: http://ypa.csbb.ntu.edu.tw/ (accessed on 25 June 2017).
- Teixeira, M.C.; Monteiro, P.T.; Guerreiro, J.F.; Gonc, J.P.; Mira, N.P.; Costa dos Santos, S.; Cabrito, T.R.; Palma, M.; Costa, C.; Francisco, A.P.; et al. The YEASTRACT database: An upgraded information system for the analysis of gene and genomic transcription regulation in Saccharomyces cerevisiae. Nucleic Acids Res. 2014, 42, D161–D166. [Google Scholar] [CrossRef] [PubMed]
- Voth, W.P.; Takahata, S.; Nishikawa, J.L.; Metcalfe, B.M.; Näär, A.M.; Stillman, D.J. A Role for FACT in Repopulation of Nucleosomes at Inducible Genes. PLoS ONE 2014, 9, e84092. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Fletcher, A.G.L.; Zhang, L.; Chen, X.; Fischbeck, J.A.; Stargell, L.A. Activation of a Poised RNAPII-Dependent Promoter Requires Both SAGA and Mediator. Genetics 2010, 184, 659–672. [Google Scholar] [CrossRef] [PubMed]
- Plaschka, C.; Nozawa, K.; Cramer, P. Mediator architecture and RNA polymerase II interaction. J. Mol. Biol. 2016, 428, 2569–2574. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Moye-Rowley, W.S. Multidrug resistance in fungi: Regulation of transporter-encoding gene expression. Front. Physiol. 2014, 5, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kolaczkowska, A.; Goffeau, A. Regulation of pleiotropic drug resistance in yeast. Drug Resist. Updates 1999, 2, 403–414. [Google Scholar] [CrossRef] [PubMed]
- DeRisi, J.; van den Hazel, B.; Marc, P.; Balzi, E.; Brown, P.; Jacq, C.; Goffeau, A. Genome microarray analysis of transcriptional activation in multidrug resistance yeast mutants. FEBS Lett. 2000, 470, 156–160. [Google Scholar] [CrossRef]
- Devaux, F.; Carvajal, E.; Moye-Rowley, S.; Jacq, C. Genome-wide studies on the nuclear PDR3-controlled response to mitochondrial dysfunction in yeast. FEBS Lett. 2001, 515, 25–28. [Google Scholar] [CrossRef]
- Hallstrom, T.C.; Moye-Rowley, W.S. Multiple Signals from Dysfunctional Mitochondria Activate the Pleiotropic Drug Resistance Pathway in Saccharomyces cerevisiae. J. Biol. Chem. 2000, 275, 37347–37356. [Google Scholar] [CrossRef] [PubMed]
- Akache, B.; MacPherson, S.; Sylvain, M.A.; Turcotte, B. Complex Interplay among Regulators of Drug Resistance Genes in Saccharomyces cerevisiae. J. Biol. Chem. 2004, 279, 27855–27860. [Google Scholar] [CrossRef] [PubMed]
- Grishin, A.V.; Rothenberg, M.; Downs, M.A.; Blumer, K.J. Mot3, a Zn Finger Transcription Factor That Modulates Gene Expression and Attenuates Mating Pheromone Signaling in Saccharomyces cerevisiae. Genetics 1998, 149, 879–892. [Google Scholar] [PubMed]
- Martínez-Montañés, F.; Rienzo, A.; Poveda-Huertes, D.; Pascual-Ahuir, A.; Proft, M. Activator and Repressor Functions of the Mot3 Transcription Factor in the Osmostress Response of Saccharomyces cerevisiae. Eukaryot. Cell 2013, 12, 636–647. [Google Scholar] [CrossRef] [PubMed]
- Kołaczkowska, A.; Manente, M.; Kołaczkowski, M.; Laba, J.; Ghislain, M.; Wawrzycka, D. The regulatory inputs controlling pleiotropic drug resistance and hypoxic response in yeast converge at the promoter of the aminocholesterol resistance gene RTA1. FEMS Yeast Res. 2012, 12, 279–292. [Google Scholar] [CrossRef] [PubMed]
- Deaner, M.; Alper, H.S. Promoter and Terminator Discovery and Engineering. Adv. Biochem. Eng. Biotechnol. 2016. [Google Scholar] [CrossRef]
- Blazeck, J.; Liu, L.; Redden, H.; Alper, H. Tuning Gene Expression in Yarrowia lipolytica by a Hybrid Promoter Approach. Appl. Environ. Microbiol. 2011, 77, 7905–7914. [Google Scholar] [CrossRef] [PubMed]
- Blazeck, J.; Reed, B.; Garg, R.; Gerstner, R.; Pan, A.; Agarwal, V.; Alper, H.S. Generalizing a hybrid synthetic promoter approach in Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Rajasärkkä, J.; Virta, M. Characterization of a Bisphenol A Specific Yeast Bioreporter Utilizing the Bisphenol A-Targeted Receptor. Anal. Chem. 2013, 85, 10067–10074. [Google Scholar] [CrossRef] [PubMed]








© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schuller, A.; Rödel, G.; Ostermann, K. Tuning the Sensitivity of the PDR5 Promoter-Based Detection of Diclofenac in Yeast Biosensors. Sensors 2017, 17, 1506. https://doi.org/10.3390/s17071506
Schuller A, Rödel G, Ostermann K. Tuning the Sensitivity of the PDR5 Promoter-Based Detection of Diclofenac in Yeast Biosensors. Sensors. 2017; 17(7):1506. https://doi.org/10.3390/s17071506
Chicago/Turabian StyleSchuller, Astrid, Gerhard Rödel, and Kai Ostermann. 2017. "Tuning the Sensitivity of the PDR5 Promoter-Based Detection of Diclofenac in Yeast Biosensors" Sensors 17, no. 7: 1506. https://doi.org/10.3390/s17071506
APA StyleSchuller, A., Rödel, G., & Ostermann, K. (2017). Tuning the Sensitivity of the PDR5 Promoter-Based Detection of Diclofenac in Yeast Biosensors. Sensors, 17(7), 1506. https://doi.org/10.3390/s17071506