Comparison and Combination of Thermal, Fluorescence, and Hyperspectral Imaging for Monitoring Fusarium Head Blight of Wheat on Spikelet Scale

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Plants
2.2. Inoculum and Inoculation
2.3. Disease Severity of FHB
2.4. Infrared Thermal Imaging (IRT)
2.5. Chlorophyll Fluorescence Imaging (CFI)
2.6. Hyperspectral Imaging (HSI)
2.7. Realization of Measurements
2.8. Statistical Analysis
3. Results
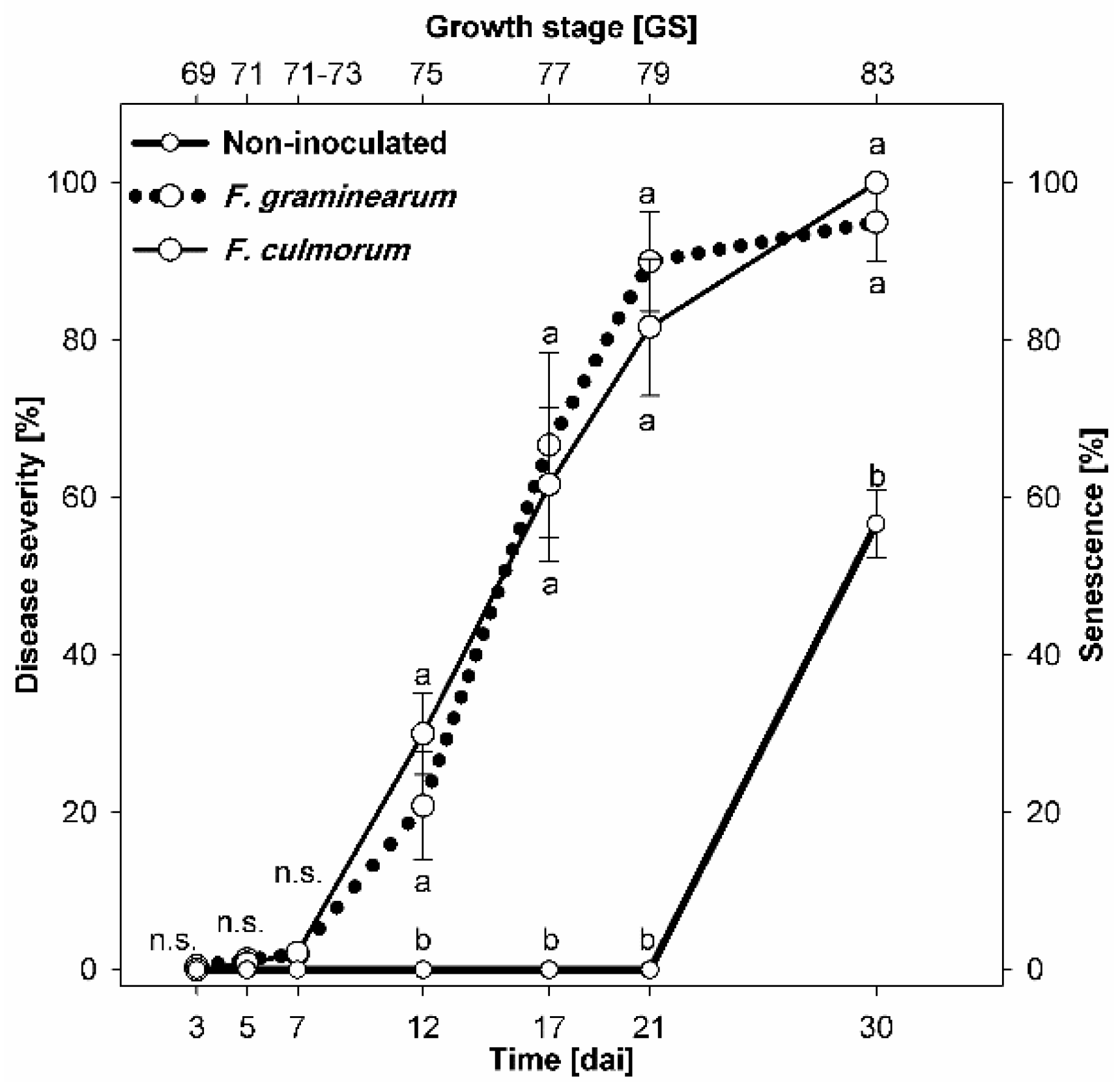
3.1. Disease Development
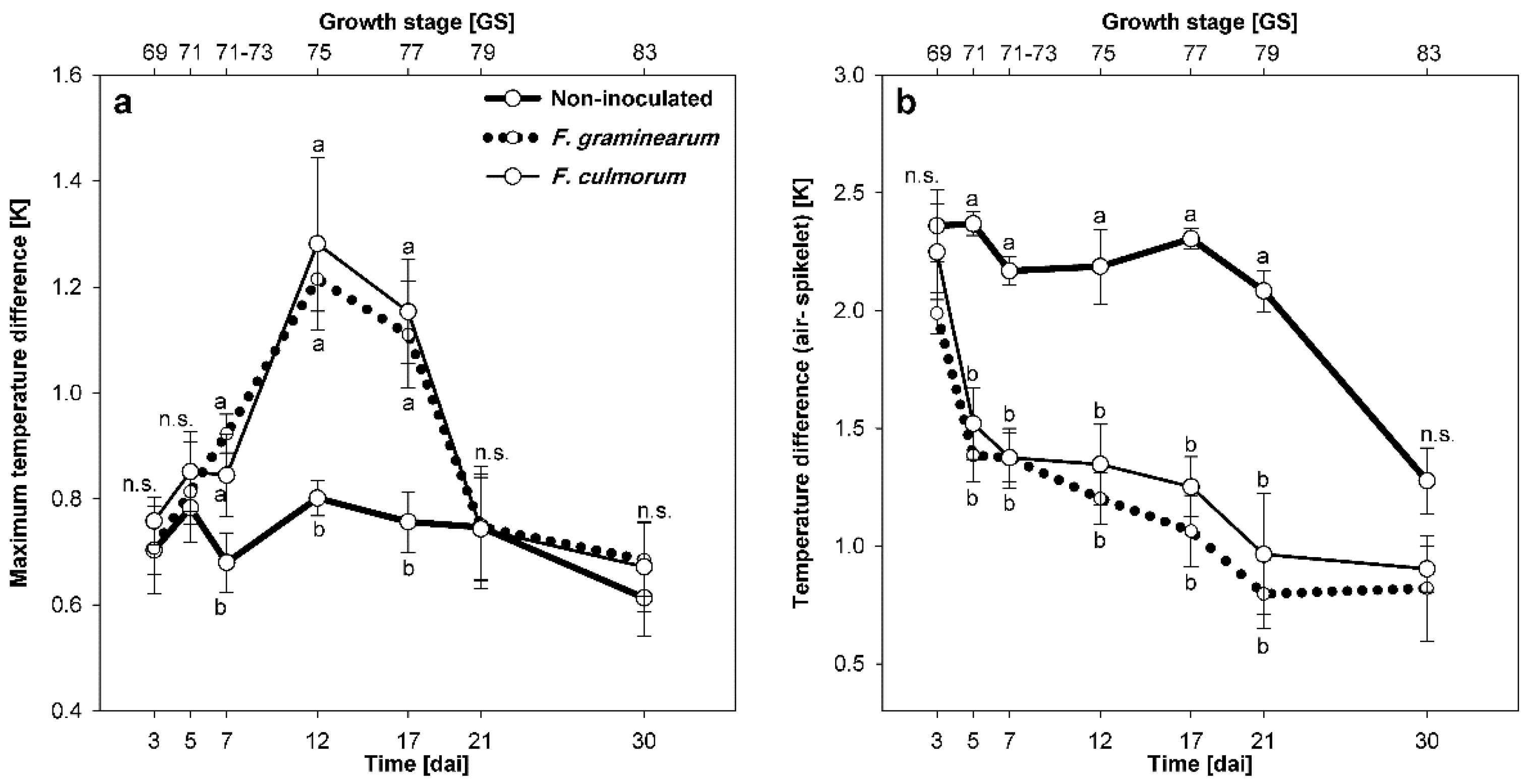
3.2. Effect of Fusarium Infection on Spikelet Temperature
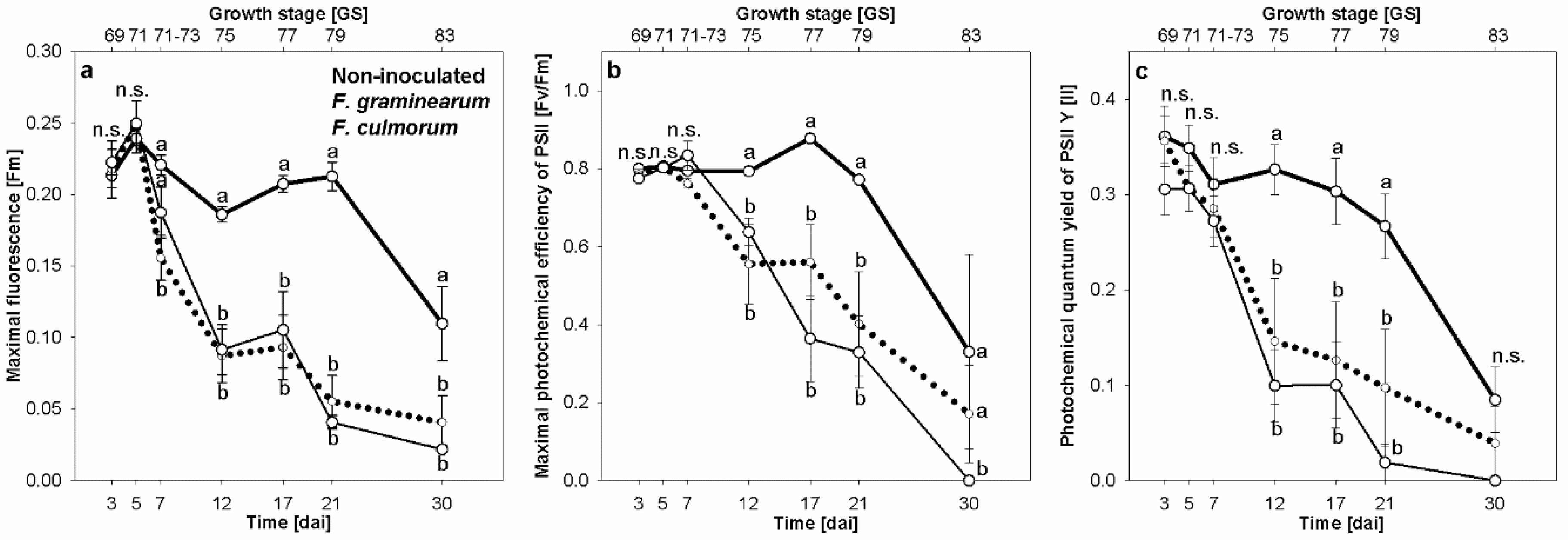
3.3. Effect of Fusarium Infection on Chlorophyll Fluorescence
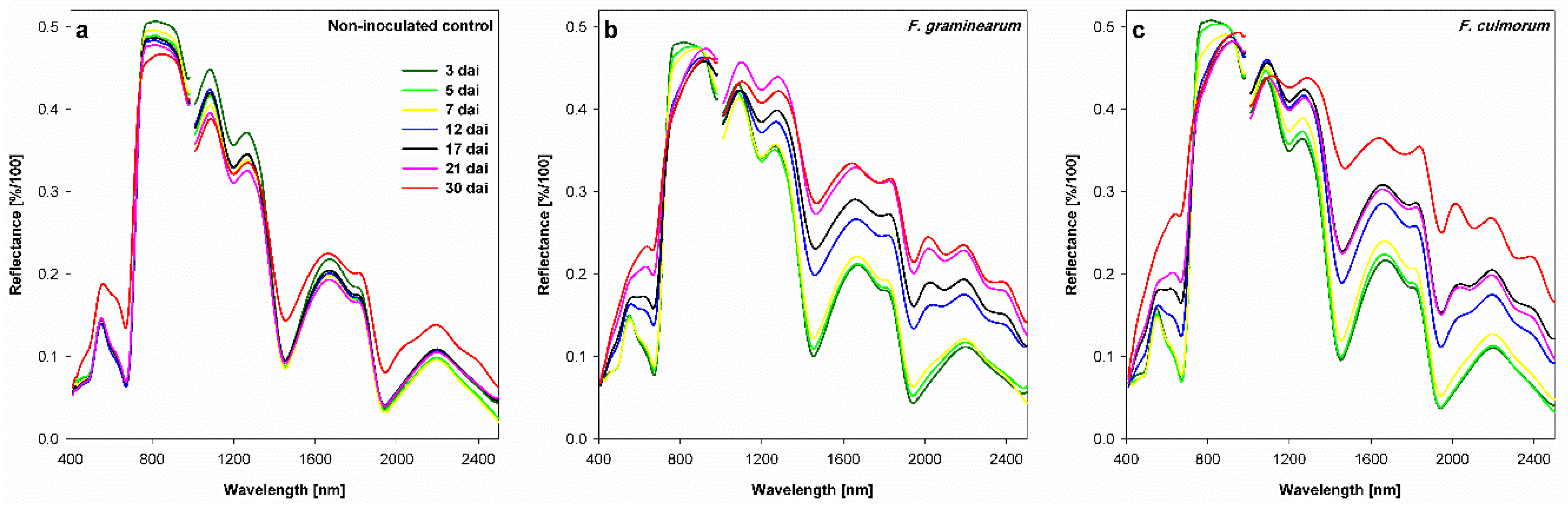
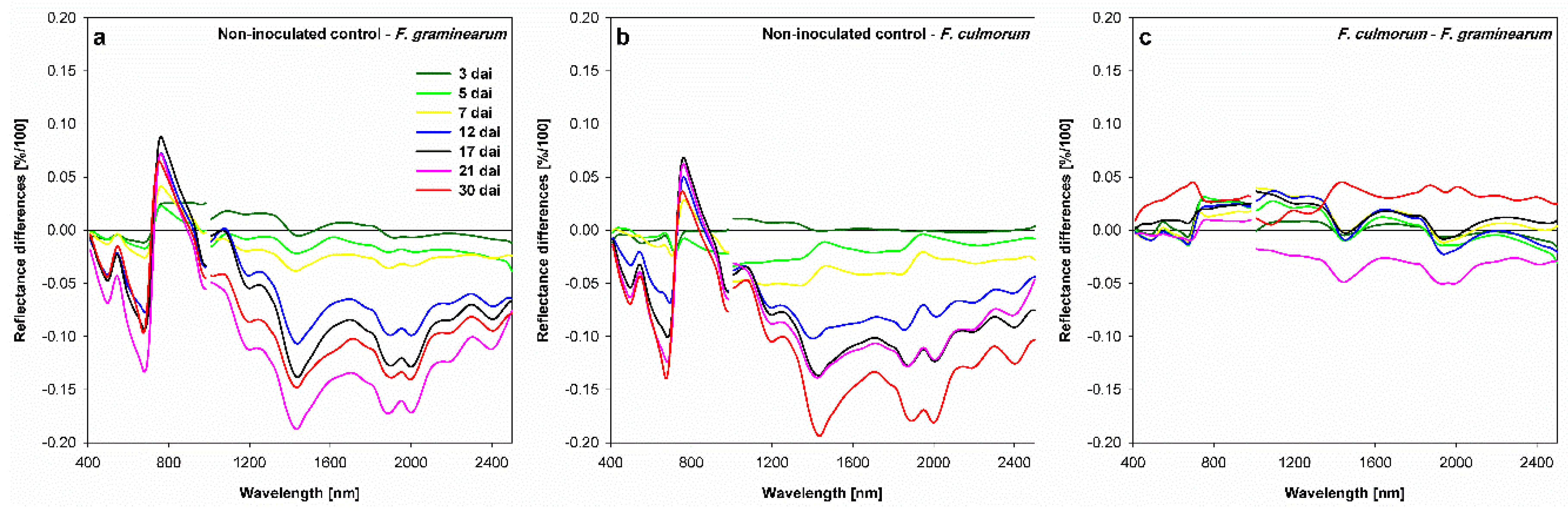
3.4. Effect of Fusarium Infection on Spectral Signature of Spikelets
3.5. Correlation between Parameters Derived from Different Sensors
3.6. Spatio-Temporal Dynamics of Fusarium Head Blight
3.7. Support Vector Machine Classification of Infected and Non-Infected Spikelets at Different Pathogenesis Stages
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Stack, R.W. History of Fusarium head blight with emphasis on North America. In Fusarium Head Blight of Wheat and Barley; Leonard, K.J., Bushnell, W.R., Eds.; American Phytopathological Society Press: St. Paul, MN, USA, 2003; pp. 1–34. [Google Scholar]
- Bauriegel, E.; Herppich, W. Hyperspectral and chlorophyll fluorescence imaging for early detection of plant diseases, with special reference to Fusarium spec. infections on wheat. Agriculture 2014, 4, 32–57. [Google Scholar] [CrossRef]
- Madgwick, J.W.; West, J.S.; White, R.P.; Semenov, M.; Townsend, J.A.; Turner, J.A.; Fitt, B.D. Impacts of climate change on wheat anthesis and Fusarium ear blight in the UK. Eur. J. Plant Pathol. 2011, 130, 117–131. [Google Scholar] [CrossRef]
- Simpson, D.R.; Weston, G.E.; Turner, J.A.; Jennings, P.; Nicholson, P. Differential control of head blight pathogens of wheat by fungicides and consequences for mycotoxin contamination of grain. Eur. J. Plant Pathol. 2001, 107, 421–431. [Google Scholar] [CrossRef]
- Bottalico, A.; Perrone, G. Toxigenic Fusarium species and mycotoxins associated with head blight in small-grain cereals in Europe. Eur. J. Plant Pathol. 2002, 108, 611–624. [Google Scholar] [CrossRef]
- Goswami, R.S.; Kistler, H.C. Heading for disaster: Fusarium graminearum on cereal crops. Mol. Plant Pathol. 2004, 5, 515–525. [Google Scholar] [CrossRef]
- O’Donnell, K.; Ward, T.J.; Geiser, D.M.; Kistler, H.C.; Aoki, T. Genealogical concordance between the mating type locus and seven other nuclear genes supports formal recognition of nine phylogenetically distinct species within the Fusarium graminearum clade. Fungal Genet. Biol. 2004, 41, 600–623. [Google Scholar] [CrossRef]
- Desjardin, A.E. Fusarium Mycotoxins: Chemistry, Genetics, and Biology; American Phytopathological Society Press: St. Paul, MN, USA, 2006; p. 268. [Google Scholar]
- Mesterházy, Á.; Tóth, B.; Varga, M.; Bartók, T.; Szabó-Hevér, Á.; Farády, L.; Lehoczki-Krsjak, S. Role of fungicides, application of nozzle types, and the resistance level of wheat varieties in the control of Fusarium Head Blight and Deoxynivalenol. Toxins 2011, 3, 1453–1483. [Google Scholar] [CrossRef]
- Wegulo, S.N.; Bockus, W.W.; Nopsa, J.H.; De Wolf, E.D.; Eskridge, K.M.; Peiris, K.H.S.; Dowell, F.E. Effects of integrating cultivar resistance and fungicide application on Fusarium head blight and deoxynivalenol in winter wheat. Plant Dis. 2011, 95, 554–560. [Google Scholar] [CrossRef] [PubMed]
- Amarasinghe, C.C.; Tamburic-Ilincic, L.; Gilbert, J.; Brûlé-Babel, A.L.; Dilantha Fernando, W.G. Evaluation of different fungicides for control of Fusarium head blight in wheat inoculated with 3ADON and 15ADON chemotypes of Fusarium graminearum in Canada. Can. J. Plant Pathol. 2013, 35, 200–208. [Google Scholar] [CrossRef]
- Mahlein, A.K.; Oerke, E.C.; Steiner, U.; Dehne, H.W. Recent advances in sensing plant diseases for precision crop protection. Eur. J. Plant Pathol. 2012, 133, 197–209. [Google Scholar] [CrossRef]
- Oerke, E.C.; Mahlein, A.K.; Steiner, U. Proximal sensing of plant diseases. In Detection and Diagnostics of Plant Pathogens; Gullino, M.L., Bonants, P.J.M., Eds.; Springer: Dordrecht, Netherlands, 2014; pp. 55–68. [Google Scholar]
- Mahlein, A.K. Plant disease detection by imaging sensors—Parallels and specific demands for precision agriculture and plant phenotyping. Plant Dis. 2016, 100, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Alisaac, E.; Behmann, J.; Kuska, M.T.; Dehne, H.W.; Mahlein, A.K. Hyperspectral quantification of wheat resistance to Fusarium head blight: comparison of two Fusarium species. Eur. J. Plant Pathol. 2018, 152, 869–884. [Google Scholar] [CrossRef]
- Lindenthal, M.; Steiner, U.; Dehne, H.W.; Oerke, E.C. Effect of downy mildew development on transpiration of cucumber leaves visualized by digital infrared thermography. Phytopathology 2005, 95, 233–240. [Google Scholar] [CrossRef]
- Oerke, E.C.; Steiner, U.; Dehne, H.W.; Lindenthal, M. Thermal imaging of cucumber leaves affected by downy mildew and environmental conditions. J. Exp. Bot. 2006, 57, 2121–2132. [Google Scholar] [CrossRef] [Green Version]
- Stoll, M.; Schultz, H.R.; Baecker, G.; Berkelmann-Loehnertz, B. Early pathogen detection under different water status and the assessment of spray application in vineyards through the use of thermal imagery. Precis. Agric. 2008, 9, 407–417. [Google Scholar] [CrossRef]
- Calderón, R.; Navas-Cortés, J.A.; Zarco-Tejada, P.J. Early detection and quantification of Verticillium wilt in olive using hyperspectral and thermal imagery over large areas. Remote Sens. 2015, 7, 5584–5610. [Google Scholar] [CrossRef]
- Joalland, S.; Screpanti, C.; Liebisch, F.; Varella, H.V.; Gaume, A.; Walter, A. Comparison of visible imaging, thermography and spectrometry methods to evaluate the effect of Heterodera schachtii inoculation on sugar beets. Plant Methods 2017, 13, 73. [Google Scholar] [CrossRef]
- Oerke, E.C.; Steiner, U. Potential of digital thermography for disease control. In Precision Crop Protection-the Challenge and Use of Heterogeneity; Oerke, E.C., Gerhards, R., Menz, G., Sikora, R.A., Eds.; Springer: Dordrecht, Netherlands, 2010; pp. 167–182. [Google Scholar]
- Al Masri, A.; Hau, B.; Dehne, H.W.; Mahlein, A.K.; Oerke, E.C. Impact of primary infection site of Fusarium species on head blight development in wheat spikes evaluated by IR-thermography. Eur. J. Plant Pathol. 2017, 147, 855–868. [Google Scholar] [CrossRef]
- Bürling, K. Potential of Fluorescence Techniques with Special Reference to Fluorescence Lifetime Determination for Sensing and Differentiating Biotic and Abiotic Stresses in Triticum aestivum L. Ph.D. Thesis, University of Bonn, Bonn, Germany, 2011. [Google Scholar]
- Kuckenberg, J.; Tartachnyk, I.; Noga, G. Temporal and spatial changes of chlorophyll fluorescence as a basis for early and precise detection of leaf rust and powdery mildew infections in wheat leaves. Precis. Agric. 2009, 10, 34–44. [Google Scholar] [CrossRef]
- Brugger, A.; Kuska, M.T.; Mahlein, A.K. Impact of compatible and incompatible barley—Blumeria graminis f.sp. hordei interactions on chlorophyll fluorescence parameters. J. Plant Dis. Prot. 2017, 125, 177–186. [Google Scholar] [CrossRef]
- Chaerle, L.; Valcke, R.; Van Der Straeten, D. Imaging techniques in plant physiology and agronomy: From simple to multispectral approaches. In Plant Physiology and Plant Molecular Biology in the New Millennium; Hemantaranjan, A., Ed.; Scientific Publisher: Jodhpur, India, 2003; Volume 5, pp. 135–155. [Google Scholar]
- Bauriegel, E.; Giebel, A.; Herppich, W.B. Hyperspectral and chlorophyll fluorescence imaging to analyse the impact of Fusarium culmorum on the photosynthetic integrity of infected wheat ears. Sensors 2011, 11, 3765–3779. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Lamb, D.W.; Niu, Z.; Zhang, Y.; Liu, L.; Wang, J. Identification of yellow rust in wheat using in-situ spectral reflectance measurements and airborne hyperspectral imaging. Precis. Agric. 2007, 8, 187–197. [Google Scholar] [CrossRef]
- Kuska, M.; Wahabzada, M.; Leucker, M.; Dehne, H.W.; Kersting, K.; Oerke, E.C.; Steiner, U.; Mahlein, A.K. Hyperspectral phenotyping on the microscopic scale: Towards automated characterization of plant-pathogen interactions. Plant Methods 2015, 11, 28. [Google Scholar] [CrossRef]
- Leucker, M.; Mahlein, A.K.; Steiner, U.; Oerke, E.C. Improvement of lesion phenotyping in Cercospora beticola-sugar beet interaction by hyperspectral imaging. Phytopathology 2016, 106, 177–184. [Google Scholar] [CrossRef]
- Mahlein, A.K.; Steiner, U.; Dehne, H.W.; Oerke, E.C. Spectral signatures of sugar beet leaves for the detection and differentiation of diseases. Precis. Agric. 2010, 11, 413–431. [Google Scholar] [CrossRef]
- Mahlein, A.K.; Rumpf, T.; Welke, P.; Dehne, H.W.; Plümer, L.; Steiner, U.; Oerke, E.C. Development of spectral indices for detecting and identifying plant diseases. Remote Sens. Environ. 2013, 128, 21–30. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, M.; Zhu, J.; Geng, S. Spectral prediction of Phytophthora infestans infection on tomatoes using artificial neural network (ANN). Int. J. Remote Sens. 2008, 29, 1693–1706. [Google Scholar] [CrossRef]
- Bock, C.H.; Poole, G.H.; Parker, P.E.; Gottwald, T.R. Plant disease severity estimated visually, by digital photography and image analysis, and by hyperspectral imaging. Crit. Rev. Plant Sci. 2010, 29, 59–107. [Google Scholar] [CrossRef]
- Berdugo, C.A.; Zito, R.; Paulus, S.; Mahlein, A.K. Fusion of sensor data for the detection and differentiation of plant diseases in cucumber. Plant Pathol. 2014, 63, 1344–1356. [Google Scholar] [CrossRef]
- Anonymous. Descriptive List of Varieties, Bundessortenamt, Germany. Getreide, Mais, Öl- und Faserpflanzen, Leguminosen, Rüben, Zwischenfrüchte, Bundessortenamt: Hannover, Deutschland, Germany. 2013. Available online: https://www.bundessortenamt.de/bsa/media/Files/BSL/bsl_getreide_2013.pdf (accessed on 15 May 2019).
- Moradi, G.M. Microbiological and Molecular Assessment of Interactions among the Major Fusarium Head Blight Pathogens on Wheat Ear. Ph.D. Thesis, University of Bonn, Bonn, Germany, 2008. [Google Scholar]
- Lancashire, P.D.; Bleiholder, H.; Van den Boom, T.; Langeluddecke, P.; Stauss, R.; Weber, E.; Witzenberger, A. A uniform decimal code for growth stages of crops and weeds. Ann. Appl. Biol. 1991, 119, 561–601. [Google Scholar] [CrossRef]
- Kitajima, M.; Butler, W.L. Fluorescence quenching in photosystem II of chloroplasts. Biochim. Biophys. Acta 1975, 376, 116–125. [Google Scholar]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring vegetation systems in the Great Plains with ERTS. In Proceedings of the 3th Earth Resources Technology Satellite-1 Symposium, Goddard Space Flight Center, NASA, Washington, DC, USA, 10–14 December 1973; pp. 309–317. [Google Scholar]
- Gamon, J.A.; Peñuelas, J.; Field, C.B. A narrow-waveband spectral index that tracks diurnal changes in photosynthetic efficiency. Remote Sens. Environ. 1992, 44, 35–44. [Google Scholar] [CrossRef]
- Blackburn, G.A. Quantifying chlorophylls and carotenoids at leaf and canopy Scales. Remote Sens. Environ. 1998, 66, 273–285. [Google Scholar] [CrossRef]
- Peñuelas, J.; Pinol, J.; Ogaya, R.; Filella, I. Estimation of plant water concentration by the reflectance Water Index WI (R900/R970). Int. J. Remote Sens. 1997, 18, 2869–2875. [Google Scholar] [CrossRef]
- Gebbers, R.; Adamchuk, V.I. Precision agriculture and food security. Science 2010, 327, 828–831. [Google Scholar] [CrossRef]
- Mahlein, A.K.; Kuska, M.T.; Behmann, J.; Polder, G.; Walter, A. Hyperspectral sensors and imaging technologies in phytopathology: State of the art. Annu. Rev. Phytopathol. 2018, 56, 535–558. [Google Scholar] [CrossRef] [PubMed]
- Lucas, J.A. Advances in plant disease and pest management. J. Agric. Sci. 2010, 149, 91–114. [Google Scholar] [CrossRef]
- Lemmens, M.; Haim, K.; Lew, H.; Ruckenbauer, P. The effect of nitrogen fertilization on Fusarium head blight development and deoxynivalenol contamination in wheat. J. Phytopathol. 2004, 152, 1–8. [Google Scholar] [CrossRef]
- Oerke, E.C.; Fröhling, P.; Steiner, U. Thermographic assessment of scab disease on apple leaves. Precis. Agric. 2010, 12, 699–715. [Google Scholar] [CrossRef]
- Gomez, S. Infection and Spread of Peronospora sparsa on Rosa sp. (Berk.)―A Microscopic and a Thermographic Approach. Ph.D. Thesis, University of Bonn, Bonn, Germany, 2014. [Google Scholar]
- Chaerle, L.; Van Caeneghem, W.; Messens, E.; Lambers, H.; Van Montagu, M.; Van Der Straeten, D. Presymptomatic visualization of plant-virus interactions by thermography. Nat. Biotechnol. 1999, 17, 813–816. [Google Scholar] [CrossRef]
- Ayeneh, A.; Van Ginkel, M.; Reynolds, M.P.; Ammar, K. Comparison of leaf, spike, peduncle and canopy temperature depression in wheat under heat stress. Field Crops Res. 2002, 79, 173–184. [Google Scholar] [CrossRef]
- Chaerle, L.; Leinonen, I.; Jones, H.G.; Van Der Straeten, D. Monitoring and screening plant populations with combined thermal and chlorophyll fluorescence imaging. J. Exp. Bot. 2007, 58, 773–784. [Google Scholar] [CrossRef]
- Cséfalvay, L.; Di Gaspero, G.; Matouš, K.; Bellin, D.; Ruperti, B.; Olejníčková, J. Pre-symptomatic detection of Plasmopara viticola infection in grapevine leaves using chlorophyll fluorescence imaging. Eur. J. Plant Pathol. 2009, 125, 291–302. [Google Scholar] [CrossRef]
- Scholes, J.; Rolfe, S.A. Chlorophyll fluorescence imaging as tool for understanding the impact of fungal diseases on plant performance: a phenomics perspective. Funct. Plant Biol. 2009, 36, 880–892. [Google Scholar] [CrossRef]
- Bauriegel, E.; Giebel, A.; Geyer, M.; Schmidt, U.; Herppich, W.B. Early detection of Fusarium infection in wheat using hyper-spectral imaging. Comput. Electron. Agric. 2011, 75, 304–312. [Google Scholar] [CrossRef]
- Bauriegel, E.; Giebel, A.; Herppich, W.B. Rapid Fusarium head blight detection on winter wheat spikes using chlorophyll fluorescence imaging. J. Appl. Bot. Food Qual. 2010, 83, 196–203. [Google Scholar]
- Leucker, M.; Wahabzada, M.; Kersting, K.; Peter, M.; Beyer, W.; Steiner, U.; Mahlein, A.K.; Oerke, E.C. Hyperspectral imaging reveals the effect of sugar beet quantitative trait loci on Cercospora leaf spot resistance. Funct. Plant Biol. 2016, 44, 1–9. [Google Scholar] [CrossRef]
- Kuska, M.T.; Brugger, A.; Thomas, S.; Wahabzada, M.; Kersting, K.; Oerke, E.C.; Steiner, U.; Mahlein, A.K. Spectral patterns reveal early resistance reactions of barley against Blumeria graminis f. sp. hordei. Phytopathology 2017, 107, 1388–1398. [Google Scholar] [CrossRef]
- Delalieux, S.; Somers, B.; Verstraeten, W.W.; van Aardt, J.A.N.; Keulemans, W.; Coppin, P. Hyperspectral indices to diagnose leaf biotic stress of apple plants, considering leaf phenology. Int. J. Remote Sens. 2009, 30, 1887–1912. [Google Scholar] [CrossRef]
- Moroni, M.; Lupo, E.; Cenedese, A. Hyperspectral proximal sensing of Salix Alba trees in the Sacco river valley (Latium, Italy). Sensors 2013, 13, 14633–14649. [Google Scholar] [CrossRef] [PubMed]
- Sims, D.A.; Gamon, J.A. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 2002, 81, 337–354. [Google Scholar] [CrossRef]
- Fourty, T.; Baret, F. Vegetation water and dry matter contents estimated from top-of-the-atmosphere reflectance data: a simulation study. Remote Sens. Environ. 1997, 61, 34–45. [Google Scholar]
- Mahlein, A.K.; Steiner, U.; Hillnhütter, C.; Dehne, H.W.; Oerke, E.C. Hyperspectral imaging for small-scale analysis of symptoms caused by different sugar beet diseases. Plant Methods 2012, 8, 3. [Google Scholar] [CrossRef] [PubMed]
- Moshou, D.; Bravo, C.; Oberti, R.; West, J.; Bodria, L.; McCartney, A.; Ramon, H. Plant disease detection based on data fusion of hyper-spectral and multi-spectral fluorescence imaging using Kohonen maps. Real-Time Imaging 2005, 11, 75–83. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sensor | Index | Equation | Indicator |
|---|---|---|---|
| Thermography (IRT) | Maximum temperature difference (MTD) | MTD = maximum − minimum temperature within spikelets | Biotic stresses in early stage [17] |
| Average temperature difference (ΔT) | ΔT = average air temperature − average spikelets temperature | Biotic stresses in early and late stages [17] | |
| Chlorophyll fluorescence imaging (CFI) | Maximal fluorescence yields | Fm | Fast chlorophyll fluorescence kinetics [24] |
| Maximal PSII quantum yield (Fv/Fm) | Fv/Fm = (Fm − F0)/Fm | Maximal photochemical efficacy of photosynthesis II [39] | |
| Effective PSII quantum yield (Y [II]) | Y [II] = (Fm’ − F)/Fm’ | Photochemical quantum yields at steady state [40] | |
| Hyperspectral imaging (HSI) | Normalized differences vegetation index (NDVI) | NDVI = (R800 − R670)/(R800 + R670) | Biomass, leaf area [41] |
| Photochemical reflection index (PRI) | PRI = (R531 − R570)/(R531 + R570) | Epoxidation state xanthophyll’s cycle; pigments and photosynthetic radiation use efficiency [42] | |
| Pigment-specific simple ratio (PSSR) | PSSRa = R800/R680 | Chlorophyll a [43] | |
| PSSRb = R800/R635 | Chlorophyll b [43] | ||
| PSSRc = R800/R470 | Carotenoid [43] | ||
| Water index (WI) | WI = R900/R970 | Water content [44] |
| Index | Treatment | Time [dai] | ||||||
|---|---|---|---|---|---|---|---|---|
| 3 | 5 | 7 | 12 | 17 | 21 | 30 | ||
| NDVI | Non-inoculated control F. graminearum F. culmorum | 0.77 a 0.72 a 0.75 a | 0.76 a 0.70 a 0.75 a | 0.77 a 0.68 a 0.72 a | 0.77 a 0.52 b 0.55 b | 0.76 a 0.47 b 0.46 b | 0.74 a 0.38 b 0.40 b | 0.55 a 0.30 b 0.24 b |
| PRI | Non-inoculated control F. graminearum F. culmorum | −0.01 a −0.02 a −0.02 a | −0.01 a −0.02 a −0.02 a | −0.02 a −0.03 b −0.03 b | −0.02 a −0.04 b −0.04 b | −0.02 a −0.05 b −0.05 b | −0.03 a −0.06 b −0.06 b | −0.06 a −0.06 a −0.06 a |
| PSSRa | Non-inoculated control F. graminearum F. culmorum | 7.10 a 5.85 b 6.46 ab | 6.72 a 5.29 b 6.56 a | 7.21 a 4.96 b 5.81 b | 6.99 a 3.29 b 3.48 b | 6.63 a 2.81 b 2.73 b | 6.29 a 2.28 b 2.39 b | 3.50 a 1.86 b 1.57 b |
| PSSRb | Non-inoculated control F. graminearum F. culmorum | 5.53 a 4.74 b 5.02 ab | 5.27 a 4.42 b 5.11 a | 5.58 a 4.24 b 4.65 b | 5.44 a 3.03 b 3.16 b | 5.19 a 2.67 b 2.59 b | 4.94 a 2.23 b 2.33 b | 2.99 a 1.90 b 1.64 b |
| PSSRc | Non-inoculated control F. graminearum F. culmorum | 6.85 a 5.88 a 6.39 a | 6.47 a 5.76 a 6.75 a | 6.96 a 5.68 b 6.50 a | 7.18 a 4.53 b 4.91 b | 6.95 a 4.22 b 4.18 b | 6.94 a 3.67 b 3.96 b | 4.96 a 3.31 b 2.83 b |
| WI | Non-inoculated control F. graminearum F. culmorum | 1.14 a 1.14 a 1.14 a | 1.14 a 1.13 a 1.13 a | 1.15 a 1.11 b 1.10 b | 1.16 a 1.05 b 1.05 b | 1.15 a 1.03 b 1.03 b | 1.15 a 1.02 b 1.02 b | 1.12 a 1.01 b 1.00 b |
| Time [dai] | Accuracy [%] of Two-Class Classification | ||||||
|---|---|---|---|---|---|---|---|
| IRT 1 | CFI 2 | HIS 3 | IRT-CFI | IRT-HSI | CFI-HSI | Multi-Sensor (IRT-CFI-HSI) | |
| 3 | 78 | 56 | 78 | 67 | 67 | 56 | 56 |
| 5 | 100 | 67 | 78 | 67 | 100 | 78 | 100 |
| 7 | 78 | 89 | 89 | 78 | 78 | 78 | 78 |
| 12 | 78 | 89 | 100 | 100 | 100 | 100 | 100 |
| 17 | 100 | 89 | 100 | 100 | 100 | 100 | 100 |
| 21 | 78 | 89 | 100 | 89 | 100 | 100 | 100 |
| 30 | 67 | 78 | 78 | 67 | 89 | 89 | 78 |
| Mean | 82 | 79 | 89 | 81 | 90 | 86 | 87 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahlein, A.-K.; Alisaac, E.; Al Masri, A.; Behmann, J.; Dehne, H.-W.; Oerke, E.-C. Comparison and Combination of Thermal, Fluorescence, and Hyperspectral Imaging for Monitoring Fusarium Head Blight of Wheat on Spikelet Scale. Sensors 2019, 19, 2281. https://doi.org/10.3390/s19102281
Mahlein A-K, Alisaac E, Al Masri A, Behmann J, Dehne H-W, Oerke E-C. Comparison and Combination of Thermal, Fluorescence, and Hyperspectral Imaging for Monitoring Fusarium Head Blight of Wheat on Spikelet Scale. Sensors. 2019; 19(10):2281. https://doi.org/10.3390/s19102281
Chicago/Turabian StyleMahlein, Anne-Katrin, Elias Alisaac, Ali Al Masri, Jan Behmann, Heinz-Wilhelm Dehne, and Erich-Christian Oerke. 2019. "Comparison and Combination of Thermal, Fluorescence, and Hyperspectral Imaging for Monitoring Fusarium Head Blight of Wheat on Spikelet Scale" Sensors 19, no. 10: 2281. https://doi.org/10.3390/s19102281
APA StyleMahlein, A.-K., Alisaac, E., Al Masri, A., Behmann, J., Dehne, H.-W., & Oerke, E.-C. (2019). Comparison and Combination of Thermal, Fluorescence, and Hyperspectral Imaging for Monitoring Fusarium Head Blight of Wheat on Spikelet Scale. Sensors, 19(10), 2281. https://doi.org/10.3390/s19102281