Assessing Understory Complexity in Beech-dominated Forests (Fagus sylvatica L.) in Central Europe—From Managed to Primary Forests

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
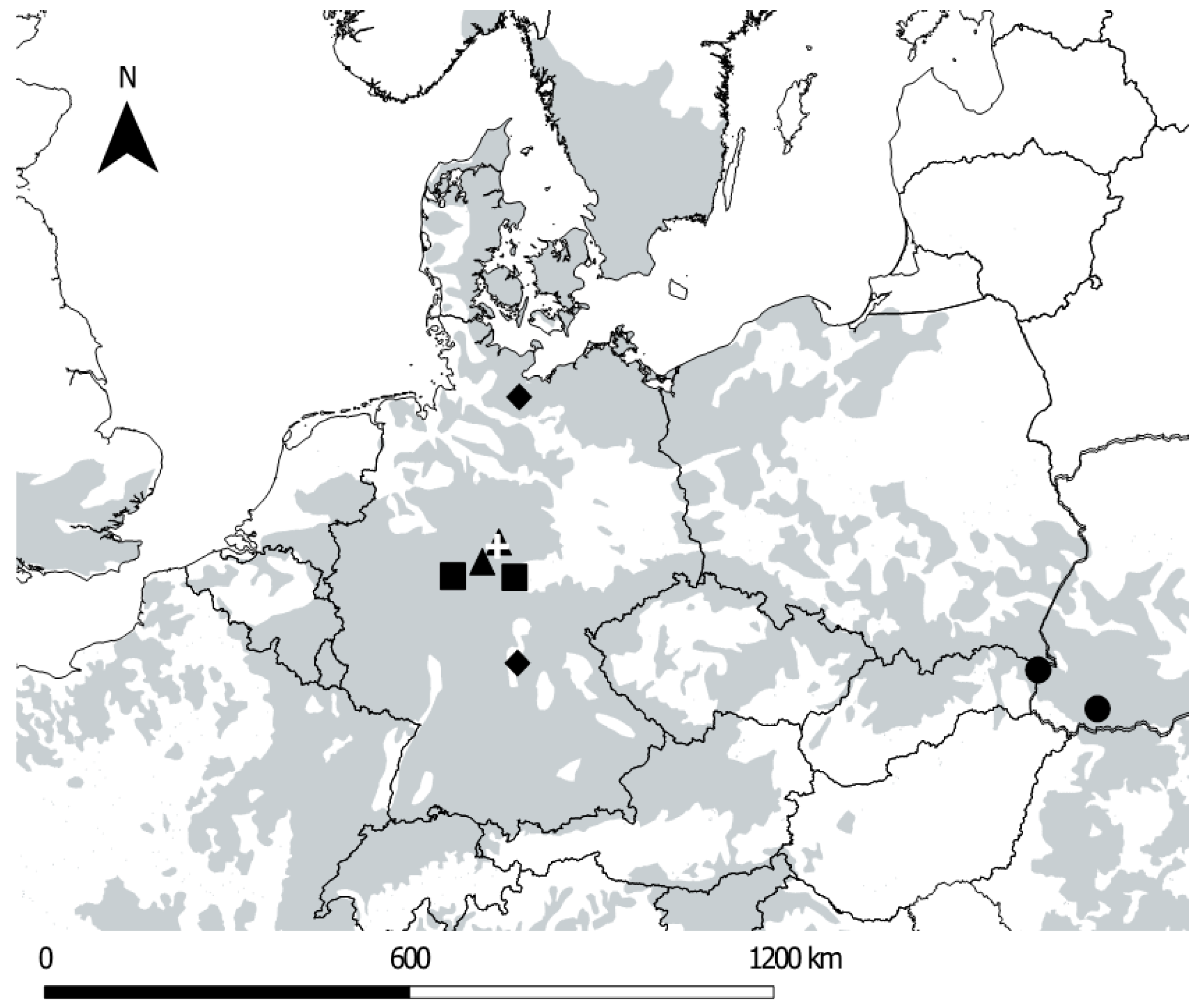
2.1. Study Sites
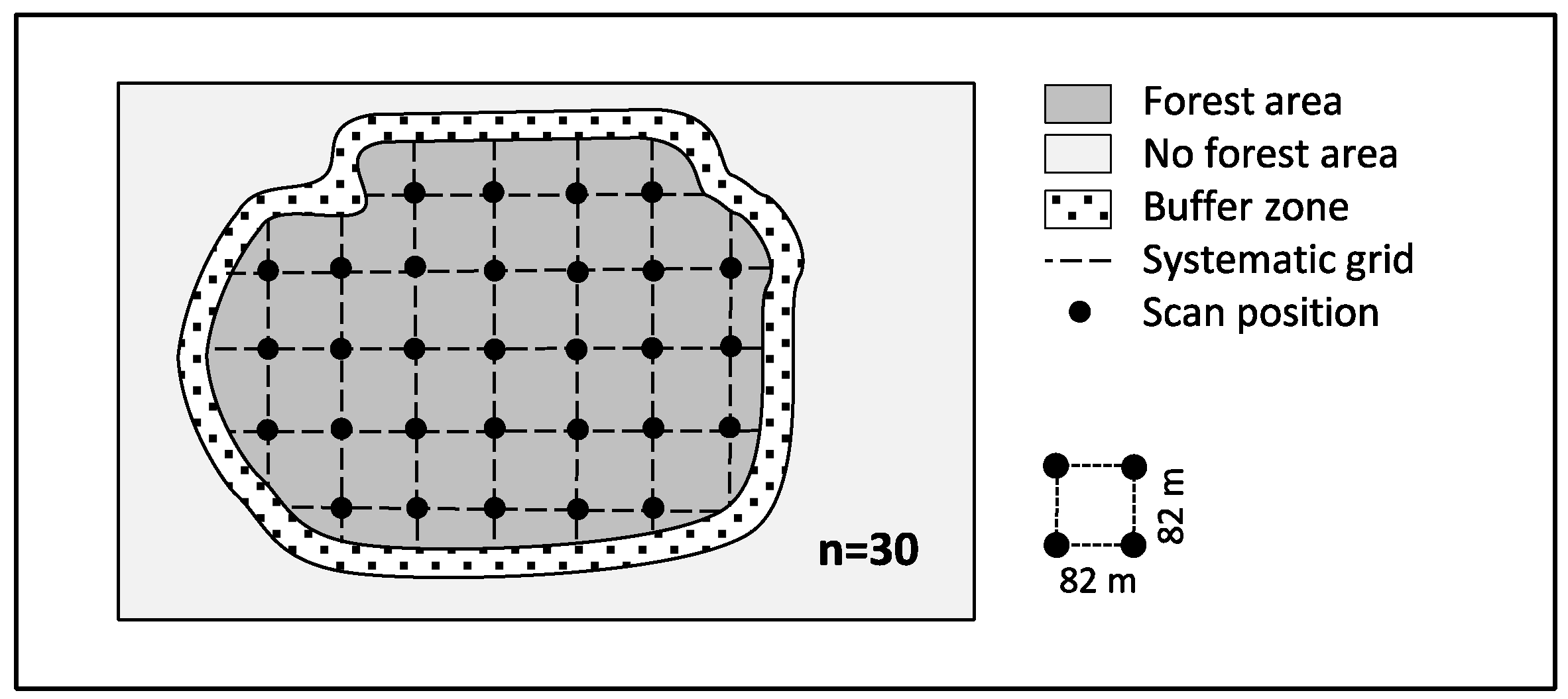
2.2. Terrestrial Laser Scanning and Sampling Design
2.3. Construction of An Understory Complexity Index
2.4. Statistics
3. Results
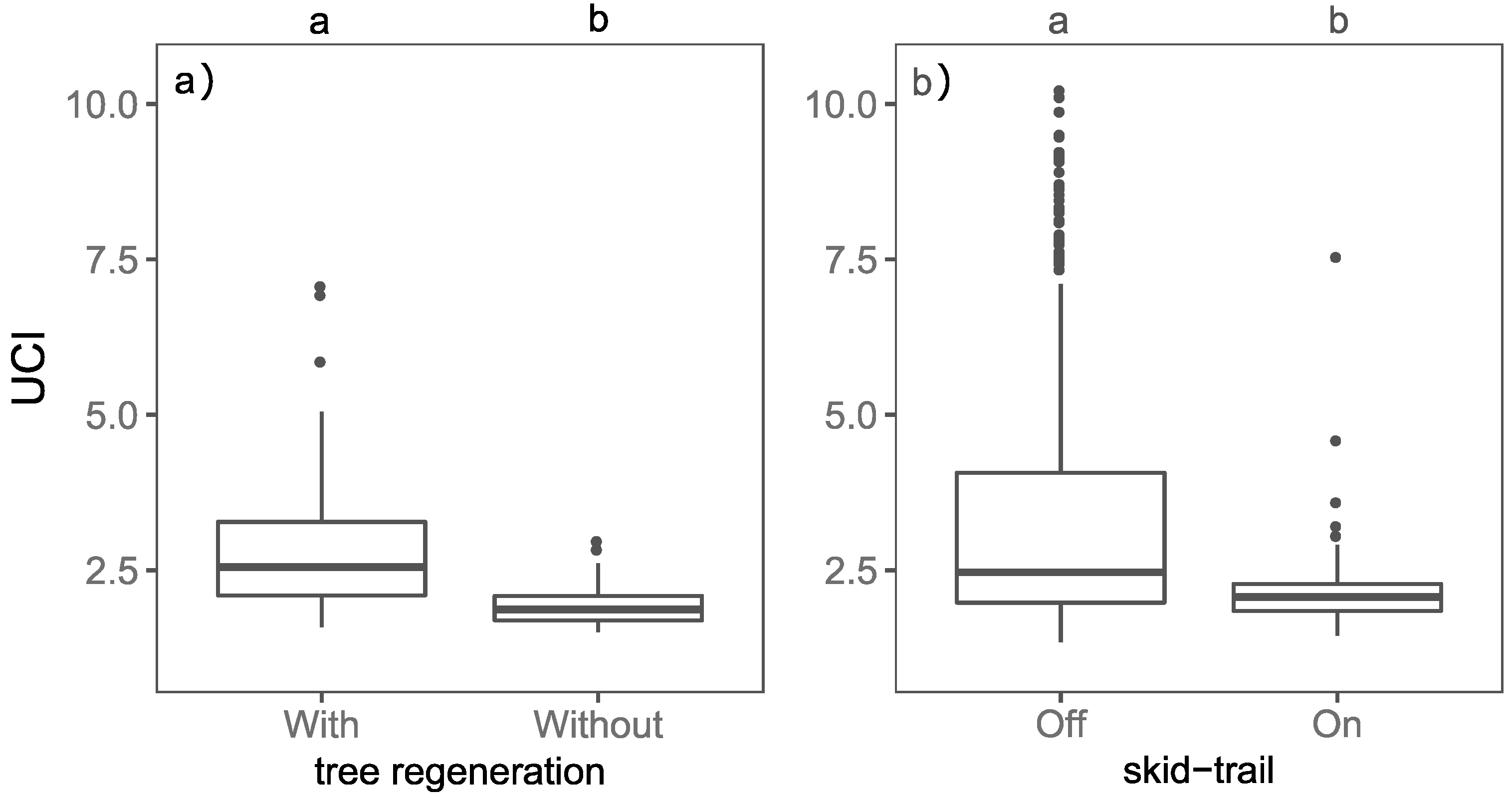
3.1. Effect of Tree Regeneration and Skid-Trails on the UCI
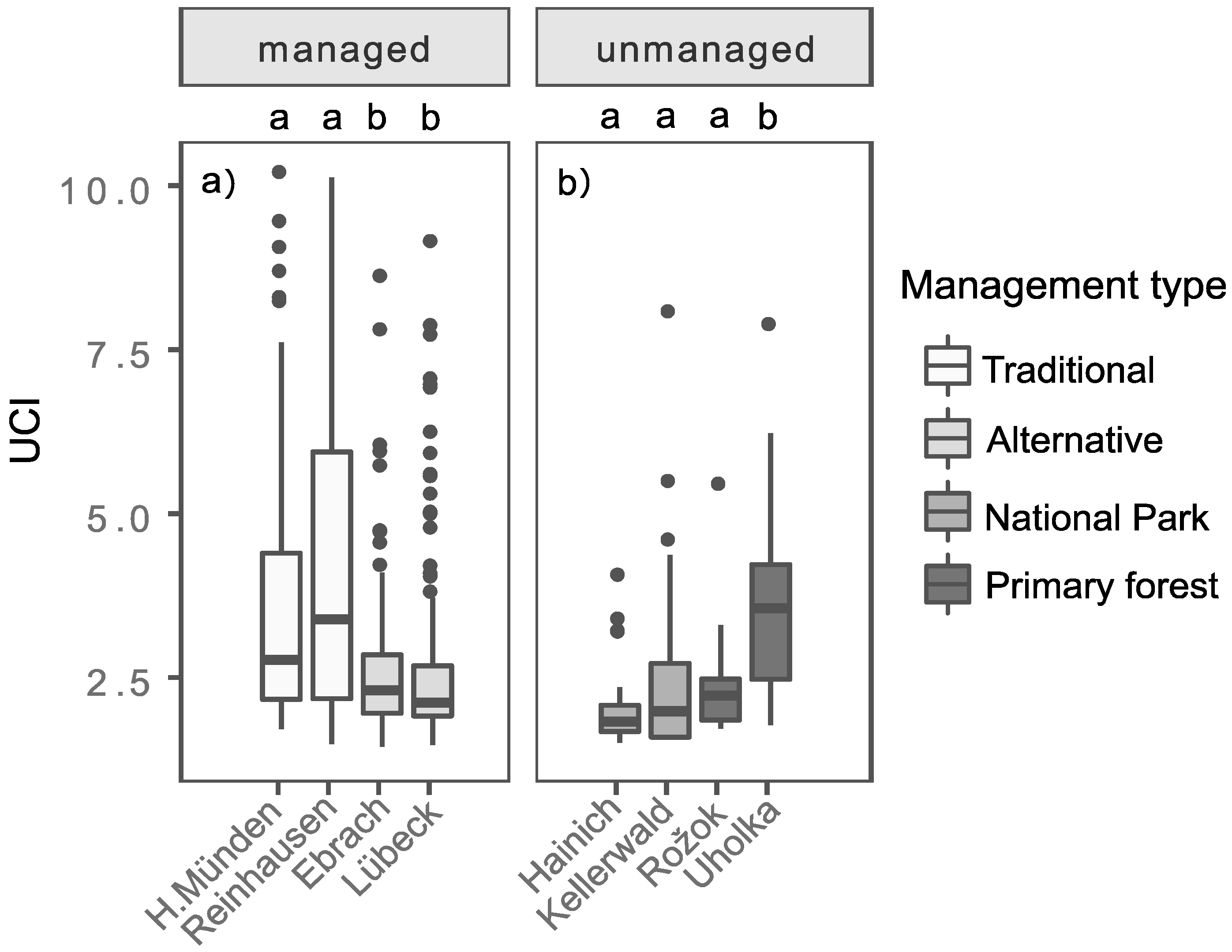
3.2. UCI of Beech Stands with Regard to Different Management Types
4. Discussion
4.1. Important Drivers of Understory Complexity
4.2. Effects of Different Management Intensities on the UCI
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Helms, J.A. The Dictionary of Forestry; Society of American Foresters: Bethesda, MD, USA, 1998; p. 192. [Google Scholar]
- Kimmins, J.P. The Biogeochemical Cycle: Nutrient Cycling Within Ecosystems. In Forest Ecology: A Foundation for Sustainable Forest Management and Environmental Ethics in Forestry, 3rd ed.; Prentice Hall: Upper Saddle River, NJ, USA, 2004; pp. 103–104. [Google Scholar]
- Thomas, S.C.; Halpern, C.B.; Falk, D.A.; Liguori, D.A.; Austin, K.A. Plant diversity in managed forests: Understory responses to thinning and fertilization. Ecol. Appl. 1999, 9, 864–879. [Google Scholar] [CrossRef]
- Augusto, L.; Dupouey, J.-L.; Ranger, J. Effects of tree species on understory vegetation and environmental conditions in temperate forests. Ann. For. Sci. 2003, 60, 823–831. [Google Scholar] [CrossRef] [Green Version]
- Antos, J.A. Understory Plants in Temperate Forests. For. For. Plants 2009, I, 262–279. [Google Scholar]
- Bauhus, J.; Puettmann, K.; Messier, C. Silviculture for old-growth attributes. For. Ecol. Manag. 2009, 258, 525–537. [Google Scholar] [CrossRef] [Green Version]
- Commarmot, B. Structures of virgin and managed beech forests in Uholka (Ukraine) and Sihlwald (Switzerland): A comparative study. For. Snow Landsc. Res. 2005, 79, 45–56. [Google Scholar]
- Kucbel, S.; Jaloviar, P.; Saniga, M.; Vencurik, J.; Klimaš, V. Canopy gaps in an old-growth fir-beech forest remnant of Western Carpathians. Eur. J. For. Res. 2010, 129, 249–259. [Google Scholar] [CrossRef]
- Trotsiuk, V.; Hobi, M.L.; Commarmot, B. Age structure and disturbance dynamics of the relic virgin beech forest Uholka (Ukrainian Carpathians). For. Ecol. Manag. 2012, 265, 181–190. [Google Scholar] [CrossRef]
- Ehbrecht, M.; Schall, P.; Ammer, C.; Seidel, D. Quantifying stand structural complexity and its relationship with forest management, tree species diversity and microclimate. Agric. For. Meteorol. 2017, 242, 1–9. [Google Scholar] [CrossRef]
- Gustafsson, L.; Baker, S.C.; Bauhus, J.; Beese, W.J.; Brodie, A.; Kouki, J.; Lindenmayer, D.B.; Lõhmus, A.; Pastur, G.M.; Messier, C.; et al. Retention Forestry to Maintain Multifunctional Forests: A World Perspective. BioScience 2012, 62, 633–645. [Google Scholar] [CrossRef] [Green Version]
- Nagel, T.A.; Zenner, E.K.; Brang, P. Research in old-growth forests and forest reserves: Implications for integrated forest management. In Integrative Approaches as An Opportunity for the Conservation of Forest Biodiversity; European Forest Institute: Freiburg, Germany, 2013; pp. 44–50. [Google Scholar]
- Brang, P. Virgin forests as a knowledge source for central European silviculture: Reality or myth? For. Snow Landsc. Res. 2005, 79, 19–32. [Google Scholar]
- Feldmann, E.; Drößler, L.; Hauck, M.; Kucbel, S.; Pichler, V.; Leuschner, C. Canopy gap dynamics and tree understory release in a virgin beech forest, Slovakian Carpathians. For. Ecol. Manag. 2018, 415–416, 38–46. [Google Scholar] [CrossRef]
- Berger, A.L.; Puettmann, K. Overstory Composition and Stand Structure Influence Herbaceous Plant Diversity in the Mixed Aspen Forest of Northern Minnesota. Am. Nat. 2000, 143, 111–125. [Google Scholar] [CrossRef]
- Spies, T.A.; Franklin, J.F. The Structure of Natural Young, Mature, and Old-Growth Douglas-Fir Forests in Oregon and Washington. In Wildlife and Vegetation of Unmanaged Douglas-fir Forests; USDA Forest Service: Asheville, NC, USA, 1991; pp. 91–111. [Google Scholar]
- Drößler, L. Struktur und Dynamik von zwei Buchenurwäldern in der Slowakei. Ph.D. Thesis, Georg-August-University, Göttingen, Germany, 2006. [Google Scholar]
- Sullivan, T.P.; Sullivan, D.S.; Lindgren, P.M. Stand structure and small mammals in young lodgepole pine forest: 10-year results after thinning. Ecol. Appl. 2001, 11, 1151–1173. [Google Scholar] [CrossRef]
- Hinsley, S.A.; Hill, R.A.; Fuller, R.J.; Bellamy, P.E.; Rothery, P. Bird species distributions across woodland canopy structure gradients. Community Ecol. 2009, 10, 99–110. [Google Scholar] [CrossRef]
- Anderson, C.S.; Meikle, D.B. Annual changes in structural complexity of understory vegetation and relative abundance of Peromyscus leucopus in fragmented habitats. Acta Theriol. 2006, 51, 43–51. [Google Scholar] [CrossRef]
- Seidel, D.; Ehbrecht, M.; Puettmann, K. Assessing different components of three-dimensional forest structure with single-scan terrestrial laser scanning: A case study. For. Ecol. Manag. 2016, 381, 196–208. [Google Scholar] [CrossRef]
- Palace, M.; Sullivan, F.B.; Ducey, M.; Herrick, C. Estimating Tropical Forest Structure Using a Terrestrial Lidar. PLoS ONE 2016, 11, e0154115. [Google Scholar] [CrossRef] [PubMed]
- Brolly, G.; Király, G.; Czimber, K. Mapping Forest Regeneration from Terrestrial Laser Scans. Acta Silv. Lignaria Hung. 2013, 9, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Stiers, M.; Willim, K.; Seidel, D.; Ehbrecht, M.; Kabal, M.; Ammer, C.; Annighöfer, P. A quantitative comparison of the structural complexity of managed, lately unmanaged and primary European beech (Fagus sylvatica L.) forests. For. Ecol. Manag. 2018, 430, 357–365. [Google Scholar] [CrossRef]
- Sabatini, F.M.; Burrascano, S.; Keeton, W.S.; Levers, C.; Lindner, M.; Pötzschner, F.; Verkerk, P.J.; Bauhus, J.; Buchwald, E.; Chaskovsky, O.; et al. Where are Europe’s last primary forests? Divers. Distrib. 2018, 24, 1426–1439. [Google Scholar] [CrossRef]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World Map of the Köppen-Geiger climate classification updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- Entscheidungshilfen zur Behandlung und Entwicklung von Buchenbeständen. Available online: https://www.nw-fva.de/fileadmin/user_upload/Verwaltung/Publikationen/Merkblaetter/Bu_Nds_Entscheidungshilfen_zur_Behandlung_und_Entwicklung_von_Buchenbestaenden.pdf (accessed on 30 June 2018).
- Kucbel, S.; Saniga, M.; Jaloviar, P.; Vencurik, J. Stand structure and temporal variability in old-growth beech-dominated forests of the northwestern Carpathians: A 40-years perspective. For. Ecol. Manag. 2012, 264, 125–133. [Google Scholar] [CrossRef]
- Hobi, M.L.; Commarmot, B.; Bugmann, H. Pattern and process in the largest primeval beech forest of Europe (Ukrainian Carpathians). J. Veg. Sci. 2015, 26, 323–336. [Google Scholar] [CrossRef]
- Korpel, Š.; Korpel’, Š. Eigenschaften, grundlegende Merkmale und Entwicklungsgesetzmäßigkeiten des Urwaldes. In Die Urwälder der Westkarpaten: 39 Tabellen; Fischer: Stuttgart, Germany, 1995; pp. 6–13. [Google Scholar]
- Cifuentes, R.; van der Zande, D.; Farifteh, J.; Salas, C.; Coppin, P. Effects of voxel size and sampling setup on the estimation of forest canopy gap fraction from terrestrial laser scanning data. Agric. For. Meteorol. 2014, 194, 230–240. [Google Scholar] [CrossRef]
- van der Zande, D.; Hoet, W.; Jonckheere, I.; van Aardt, J.; Coppin, P. Influence of measurement set-up of ground-based LiDAR for derivation of tree structure. Agric. For. Meteorol. 2006, 141, 147–160. [Google Scholar] [CrossRef]
- McGarigal, K.; Marks, B.J. FRAGSTATS: Spatial Pattern Analysis Program for Quantifying Landscape Structure, version 2; Forest Science Department: Corvallis, OR, USA, 1994; pp. 1–138. [Google Scholar]
- Fuller, R.J. Responses of woodland birds to increasing numbers of deer: A review of evidence and mechanisms. Forestry 2001, 74, 289–298. [Google Scholar] [CrossRef]
- Hinsley, S.A.; Carpenter, J.E.; Broughton, R.K.; Bellamy, P.E.; Rothery, P.; Amar, A.; Hewson, C.M.; Gosler, A.G. Habitat selection by Marsh Tits Poecile palustris in the UK. IBIS 2007, 149, 224–233. [Google Scholar] [CrossRef] [Green Version]
- McElhinny, C.; Gibbons, P.; Brack, C.; Bauhus, J. Forest and woodland stand structural complexity: Its definition and measurement. For. Ecol. Manag. 2005, 218, 1–24. [Google Scholar] [CrossRef]
- Daume, S.; Robertson, D. A heuristic approach to modelling thinnings. Silva Fenn. 2000, 34, 237–249. [Google Scholar] [CrossRef]
- Schall, P.; Gossner, M.M.; Heinrichs, S.; Fischer, M.; Boch, S.; Prati, D.; Jung, K.; Baumgartner, V.; Blaser, S.; Böhm, S.; et al. The impact of even-aged and uneven-aged forest management on regional biodiversity of multiple taxa in European beech forests. J. Appl. Ecol. 2018, 109, 267–278. [Google Scholar] [CrossRef]
- Agestam, E.; Ekö, P.-M.; Nilsson, U.; Welander, N. The effects of shelterwood density and site preparation on natural regeneration of Fagus sylvatica in southern Sweden. For. Ecol. Manag. 2003, 176, 61–73. [Google Scholar] [CrossRef]
- Modrý, M.; Hubený, D.; Rejšek, K. Differential response of naturally regenerated European shade tolerant tree species to soil type and light availability. For. Ecol. Manag. 2004, 188, 185–195. [Google Scholar] [CrossRef]
- Ammer, C. Konkurrenz um Licht-Zur Entwicklung der Naturverjüngung im Bergmischwald; Forstliche Forschungsberichte München: Freising, Germany, 1996; p. 196. [Google Scholar]
- Vockenhuber, E.A.; Scherber, C.; Langenbruch, C.; Meißner, M.; Seidel, D.; Tscharntke, T. Tree diversity and environmental context predict herb species richness and cover in Germany’s largest connected deciduous forest. Perspect. Plant Ecol. Evol. Syst. 2011, 13, 111–119. [Google Scholar] [CrossRef]
- Holzwarth, F.; Kahl, A.; Bauhus, J.; Wirth, C. Many ways to die–partitioning tree mortality dynamics in a near-natural mixed deciduous forest. J. Ecol. 2013, 101, 220–230. [Google Scholar] [CrossRef]
- Meyer, P.; Vath, T.; Burghard, V.L. Die Struktur albanischer Rotbuchen-Urwälder-Ableitungen für eine naturnahe Buchenwirtschaft. Forstwiss. Cent. 2003, 122, 47–58. [Google Scholar] [CrossRef]
- Firm, D.; Nagel, T.A.; Diaci, J. Disturbance history and dynamics of an old-growth mixed species mountain forest in the Slovenian Alps. For. Ecol. Manag. 2009, 257, 1893–1901. [Google Scholar] [CrossRef]
- Rugani, T.; Diaci, J.; Hladnik, D. Gap Dynamics and Structure of Two Old-Growth Beech Forest Remnants in Slovenia. PLoS ONE 2012, 8, e52641. [Google Scholar] [CrossRef]
- Hobi, M.L.; Ginzler, C.; Commarmot, B.; Bugmann, H. Gap pattern of the largest primeval beech forest of Europe revealed by remote sensing. Ecosphere 2015, 6, 1–15. [Google Scholar] [CrossRef]
- Nagel, T.A.; Svoboda, M.; Rugani, T.; Diaci, J. Gap regeneration and replacement patterns in an old-growth Fagus–Abies forest of Bosnia–Herzegovina. Plant Ecol. 2010, 208, 307–318. [Google Scholar] [CrossRef]
- Diaci, J.; Adamic, T.; Rozman, A. Gap recruitment and partitioning in an old-growth beech forest of the Dinaric Mountains: Influences of light regime, herb competition and browsing. For. Ecol. Manag. 2012, 285, 20–28. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country | Management Type | Study Sites | Mean Temperature (°C) | Precipitation (mm y−1) | Elevation (m a.s.l.) | Age Class (Years) |
|---|---|---|---|---|---|---|
| Germany | Traditional | Waake (inventory site) | 7.5 | 750 | 220–400 | 0–20, 21–40, 41–80, 81–120, 121–190 |
| Germany | Traditional | Hann. Münden Reinhausen | 6.5–7.5 8 | 750–1050 740 | 270–410 190–310 | 0–20, 21–40, 41–80, 81–120 |
| Alternative | Ebrach Lübeck | 7–8 8–8.5 | 850 625–725 | 320–480 40–90 | 0–20, 21–40, 41–80, 81–120 | |
| National Park | Kellerwald Hainich | 6–8 7–8 | 600–800 600–800 | 540–635 330–380 | ~180 ~180 | |
| Slovakia | Primary forest | Rožok | 6–7 | 780 | 580–745 | ~220 |
| Ukraine | Uholka | 7 | 1407 | 700–840 | ~350 |
| Management Type | Degree of Intervention |
|---|---|
| Traditional | Yield-orientated with thinning cycles of 5 to 10 years and removal of up to 3 competitors per intervention; target-diameter harvest |
| Alternative | Compared to traditional forestry, lower thinning and harvesting frequencies and intensities + additional management goals |
| National Park | Unmanaged for 20–30 years |
| Primary forest | Unmanaged, no or minimal human impact |
| Management Type | Age Class | Mean | Median | Minimum | Maximum | Standard Deviation | Variance | Coefficient of Variation (%) |
|---|---|---|---|---|---|---|---|---|
| Traditional | all | 3.89 | 2.91 | 1.49 | 10.23 | 2.24 | 5.02 | 57.53 |
| 0–20 e | 6.05 | 6.47 | 1.76 | 10.23 | 2.51 | 6.31 | 41.54 | |
| 21–40 a | 4.06 | 3.93 | 2.11 | 8.67 | 1.62 | 2.63 | 39.88 | |
| 41–80 bc | 2.25 | 2.10 | 1.67 | 7.64 | 0.77 | 0.59 | 34.17 | |
| 81–120 ba | 3.25 | 2.47 | 1.49 | 9.52 | 1.73 | 2.99 | 53.30 | |
| Alternative | all | 2.67 | 2.22 | 1.44 | 9.18 | 1.35 | 1.82 | 50.40 |
| 0–20 a | 4.18 | 3.73 | 1.91 | 9.18 | 1.85 | 3.42 | 44.25 | |
| 21–40 b | 2.34 | 2.31 | 1.78 | 3.41 | 0.33 | 0.11 | 14.16 | |
| 41–80 cd | 2.13 | 2.02 | 1.48 | 3.52 | 0.41 | 0.17 | 19.24 | |
| 81–120 cd | 2.05 | 1.84 | 1.44 | 6.10 | 0.74 | 0.55 | 36.07 | |
| National Park | ~180 cd | 2.26 | 1.86 | 1.34 | 8.11 | 1.15 | 1.33 | 50.93 |
| Primary Forest | all | 2.96 | 2.47 | 1.72 | 7.92 | 1.31 | 1.71 | 44.18 |
| ~220 bcd | 2.34 | 2.22 | 1.72 | 5.48 | 0.73 | 0.53 | 31.19 | |
| ~350 a | 3.60 | 3.56 | 1.77 | 7.92 | 1.47 | 2.16 | 40.78 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Willim, K.; Stiers, M.; Annighöfer, P.; Ammer, C.; Ehbrecht, M.; Kabal, M.; Stillhard, J.; Seidel, D. Assessing Understory Complexity in Beech-dominated Forests (Fagus sylvatica L.) in Central Europe—From Managed to Primary Forests. Sensors 2019, 19, 1684. https://doi.org/10.3390/s19071684
Willim K, Stiers M, Annighöfer P, Ammer C, Ehbrecht M, Kabal M, Stillhard J, Seidel D. Assessing Understory Complexity in Beech-dominated Forests (Fagus sylvatica L.) in Central Europe—From Managed to Primary Forests. Sensors. 2019; 19(7):1684. https://doi.org/10.3390/s19071684
Chicago/Turabian StyleWillim, Katharina, Melissa Stiers, Peter Annighöfer, Christian Ammer, Martin Ehbrecht, Myroslav Kabal, Jonas Stillhard, and Dominik Seidel. 2019. "Assessing Understory Complexity in Beech-dominated Forests (Fagus sylvatica L.) in Central Europe—From Managed to Primary Forests" Sensors 19, no. 7: 1684. https://doi.org/10.3390/s19071684
APA StyleWillim, K., Stiers, M., Annighöfer, P., Ammer, C., Ehbrecht, M., Kabal, M., Stillhard, J., & Seidel, D. (2019). Assessing Understory Complexity in Beech-dominated Forests (Fagus sylvatica L.) in Central Europe—From Managed to Primary Forests. Sensors, 19(7), 1684. https://doi.org/10.3390/s19071684