
Analysis of Cooperative Perception in Ant Traffic and Its Effects on Transportation System by Using a Congestion-Free Ant-Trail Model


Abstract
:1. Introduction
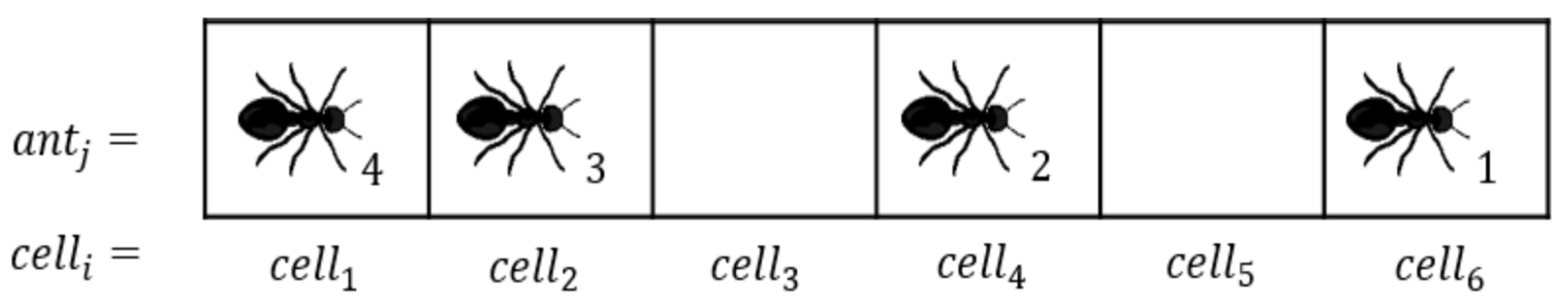
2. Model and Simulation Scenario
- is a binary variable, which either can be or 1 depending on whether the cell is empty () or occupied () by an ant at time step .
- (t) is a numerical variable, which represents the pheromone concentration in the given cell. ranges from 0 to , where means that there is no pheromone at time step , whereas means that the cell is saturated with pheromone at that time step. In a real-life AT study, pheromone concentration is measured in the number of molecules per cubic centimeter (). Whereas in ATM, pheromone concentration is measured in units of pheromone per cell ().
- is the instantaneous velocity of ant at time step measured in cells per time step (). is continuous and ranges from zero to one.
- is the position of ant on the trail at time step and ranges from zero to . Similar to is also continuous.
2.1. Stage I: Ant Motion
2.2. Stage II: Pheromone Updating
- Evaporation:
- Accumulation:
2.3. Simulation Scenarios
3. Analysis of Pheromone Concentration and Its Implications for Cooperative Perception in the ATM
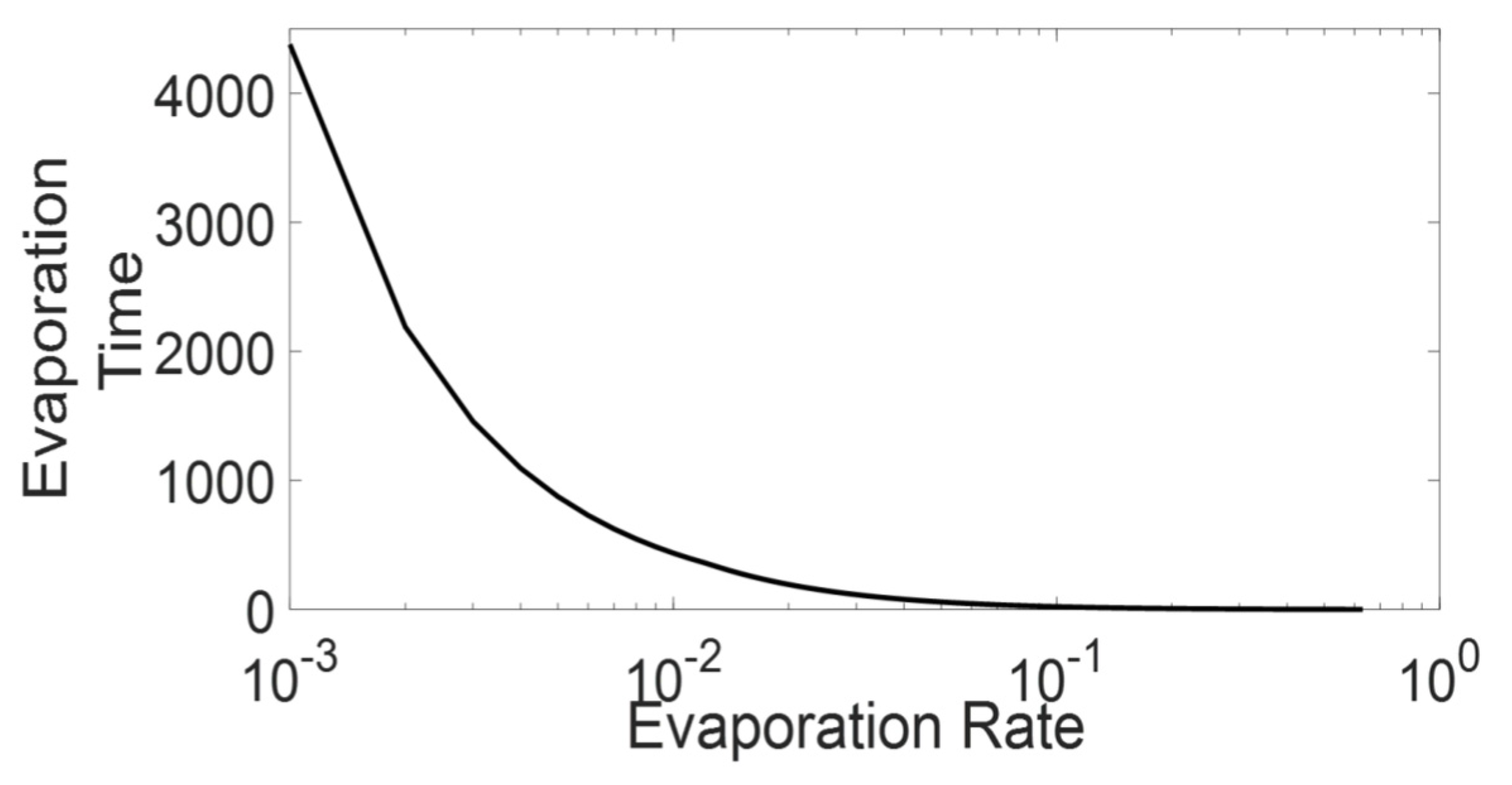
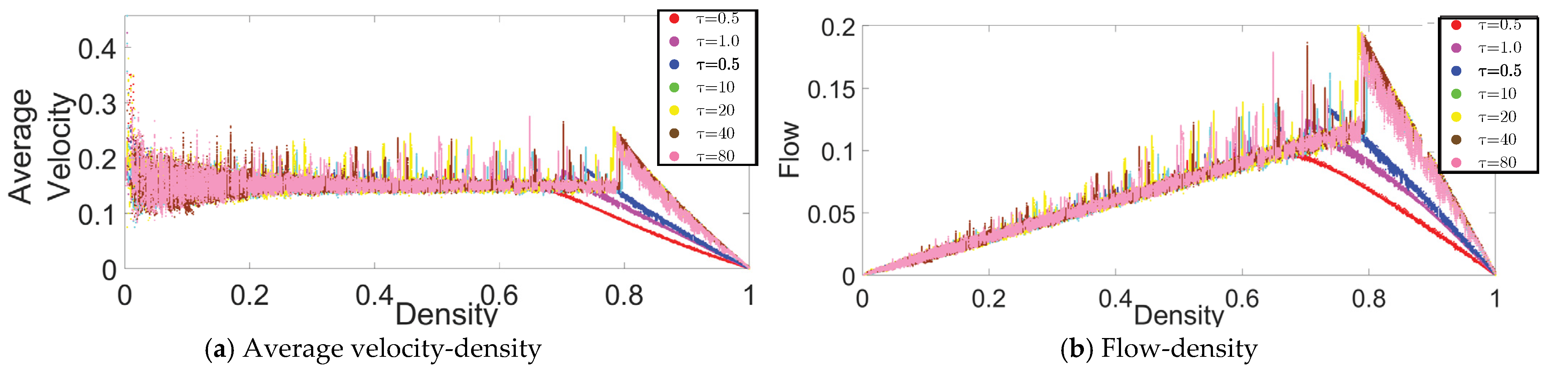
3.1. Evaporation Rate and Fundamental Diagrams
3.1.1. High-Medium Evaporation Rate (
3.1.2. Meager Evaporation Rate ()
3.1.3. Low Evaporation Rate (
3.2. Pheromone Concentration and the Corresponding States of the CP&C in AT
3.2.1. Minimal Pheromone State
3.2.2. Inactive State
3.2.3. Active State
3.3. Pheromone Concentration and Cooperative Perception in AT
4. Analysis of Pheromone Dynamics in Cooperative Perceptions of AT
4.1. Aggregation of Pheromone
4.2. Depletion of Pheromone
- Minimal state:
- Active state:
- Inactive state:
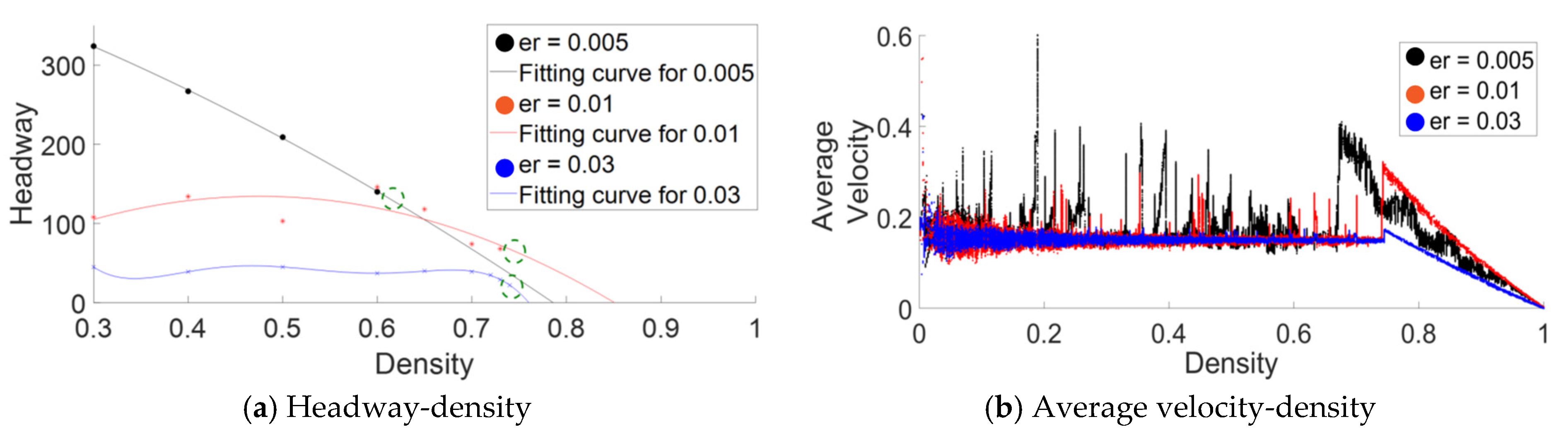
5. Analysis of Pheromone Emission Rate and Its Effect on CP&C System
6. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A. Validation of Analysis by Verifying the Non-Monotonic Behavior in ATM

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (Equation (9)) | (Equation (12)) | ||
|---|---|---|---|
| 0.005 | 80.00 | 874.21 | 131.13 |
| 0.01 | 80.00 | 436.00 | 65.40 |
| 0.03 | 32.33 | 114.12 | 17.12 |

References
- Rauch, A.; Klanner, F.; Dietmayer, K. Analysis of V2X communication parameters for the development of a fusion architecture for cooperative perception systems. In Proceedings of the 2011 IEEE Intelligent Vehicles Symposium, Baden-Baden, Germany, 5–9 June 2011; pp. 685–690. [Google Scholar]
- Tsukada, M.; Oi, T.; Kitazawa, M.; Esaki, H. Networked Roadside Perception Units for Autonomous Driving. Sensors 2020, 20, 5320. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Qin, B.; Chong, Z.J.; Shen, X.; Liu, W.; Ang, M.H.; Frazzoli, E.; Rus, D. Multivehicle Cooperative Driving Using Cooperative Perception: Design and Experimental Validation. IEEE Trans. Intell. Transp. Syst. 2015, 16, 663–680. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Y.; Chen, L.; Ji, J. Multi-Vehicle Cooperative Target Tracking with Time-Varying Localization Uncertainty via Recursive Variational Bayesian Inference. Sensors 2020, 20, 6487. [Google Scholar] [CrossRef] [PubMed]
- Shan, M.; Narula, K.; Wong, Y.F.; Worrall, S.; Khan, M.; Alexander, P.; Nebot, E. Demonstrations of Cooperative Perception: Safety and Robustness in Connected and Automated Vehicle Operations. Sensors 2020, 21, 200. [Google Scholar] [CrossRef]
- Schiegg, F.A.; Llatser, I.; Bischoff, D.; Volk, G. Collective Perception: A Safety Perspective. Sensors 2020, 21, 159. [Google Scholar] [CrossRef]
- Naujoks, F.; Grattenthaler, H.; Neukum, A.; Weidl, G.; Petrich, D. Effectiveness of advisory warnings based on cooperative perception. IET Intell. Transp. Syst. 2015, 9, 606–617. [Google Scholar] [CrossRef] [Green Version]
- Lenné, M.G.; Triggs, T.J.; Mulvihill, C.M.; Regan, M.A.; Corben, B.F. undefined Detection of emergency vehicles: Driver responses to advance warning in a driving simulator. Hum. Factors 2008, 50, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Seeliger, F.; Weidl, G.; Petrich, D.; Naujoks, F.; Breuel, G.; Neukum, A.; Dietmayer, K. Advisory warnings based on cooperative perception. In Proceedings of the 2014 IEEE Intelligent Vehicles Symposium, Dearborn, MI, USA, 8–11 June 2014; pp. 246–252. [Google Scholar]
- Thangaraj, M.; Sangam, R.S. Study of Self-Organizing Coordination for Multi-UAV Systems. In Managing Resources for Futuristic Wireless Networks; IGI Global: Hershey, PA, USA, 2020; pp. 56–69. [Google Scholar]
- Chowdhury, D.; Schadschneider, A.; Nishinari, K. Physics of transport and traffic phenomena in biology: From molecular motors and cells to organisms. Phys. Life Rev. 2005, 2, 318–352. [Google Scholar] [CrossRef] [Green Version]
- Couzin, I.D.; Franks, N.R. Self-organized lane formation and optimized traffic flow in army ants. Proc. R. Soc. Lond. B Biol. Sci. 2003, 270, 139–146. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, D.; Guttal, V.; Nishinari, K.; Schadschneider, A. A cellular-automata model of flow in ant trails: Non-monotonic variation of speed with density. J. Phys. A. Math. Gen. 2002, 35, L573. [Google Scholar] [CrossRef] [Green Version]
- Kasture, P.; Nishimura, H. Platoon Definitions and Analysis of Correlation Between Number of Platoons and Jamming in Traffic System. IEEE Trans. Intell. Transp. Syst. 2019, 22, 319–328. [Google Scholar] [CrossRef]
- John, A.; Schadschneider, A.; Chowdhury, D.; Nishinari, K. Collective effects in traffic on bi-directional ant trails. J. Theor. Biol. 2004, 231, 279–285. [Google Scholar] [CrossRef] [Green Version]
- Nishinari, K.; Sugawara, K.; Kazama, T.; Schadschneider, A.; Chowdhury, D. Modelling of self-driven particles: Foraging ants and pedestrians. Phys. A Stat. Mech. Appl. 2006, 372, 132–141. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, D.; Santen, L.; Schadschneider, A. Statistical physics of vehicular traffic and some related systems. Phys. Rep. 2000, 329, 199–329. [Google Scholar] [CrossRef] [Green Version]
- Hölldobler, B.; Wilson, E.O. The Ants; Harvard University Press: Cambridge, MA, USA, 1990. [Google Scholar]
- Wilson, E.O. The Insect Societies; Harvard University Press: Cambridge, MA, USA, 1971. [Google Scholar]
- Camazine, S.; Deneubourg, J.; Franks, N.; Sneyd, J. Self-Organization in Biological Systems; Princeton University Press: Princeton, NJ, USA, 2003. [Google Scholar]
- John, A.; Schadschneider, A.; Chowdhury, D.; Nishinari, K. Trafficlike collective movement of ants on trails: Absence of a jammed phase. Phys. Rev. Lett. 2009, 102, 108001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Song, W.; Zhang, J.; Wang, S.; Wu, C.; Lo, S. Understanding single-file movement with ant experiments and a multi-grid CA model. Phys. A Stat. Mech. Appl. 2019, 513, 1–13. [Google Scholar] [CrossRef]
- Chaudhuri, D.; Nagar, A. Absence of jamming in ant trails: Feedback control of self-propulsion and noise. Phys. Rev. E 2015, 91, 12706. [Google Scholar] [CrossRef] [Green Version]
- Guo, N.; Hu, M.-B.; Jiang, R.; Ding, J.; Ling, X. Modeling no-jam traffic in ant trails: A pheromone-controlled approach. J. Stat. Mech. Theory Exp. 2018, 2018, 53405. [Google Scholar] [CrossRef]
- Kasture, P.; Nishimura, H. Congestion-Free Ant Traffic: Jam Absorption Mechanism in Multiple Platoons. Appl. Sci. 2019, 9, 2918. [Google Scholar] [CrossRef] [Green Version]
- Nishinari, K.; Chowdhury, D.; Schadschneider, A. Cluster formation and anomalous fundamental diagram in an ant-trail model. Phys. Rev. E 2003, 67, 36120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishinari, K. Jamology: Physics of Self-Driven Particles and Toward Solution of All Jams. In Distributed Autonomous Robotic Systems 8; Springer: Berlin, Germany, 2009; pp. 175–184. [Google Scholar]
- Kunwar, A.; John, A.; Nishinari, K.; Schadschneider, A.; Chowdhury, D. Collective traffic-like movement of ants on a trail: Dynamical phases and phase transitions. J. Phys. Soc. Jpn. 2004, 73, 2979–2985. [Google Scholar] [CrossRef] [Green Version]
- Kasture, P.; Nishimura, H. Analysis of Microscopic Behavior in Ant Traffic to Understand Jam-Free Transportation. In Proceedings of the 2020 4th International Conference on Intelligent Systems, Metaheuristics & Swarm Intelligence, Thimphu, Bhutan, 21–22 March 2020; Association for Computing Machinery: New York, NY, USA, 2020; pp. 59–64. [Google Scholar]
- Kasture, P. Comprehensive Design of Improved Ant Trail Model and Analysis of Jam Absorption Mechanism. Ph.D. Thesis, Graduat e School of System Design and Management, Keio University, Yokohama, Kanagawa, Japan, 2020. [Google Scholar]
- Nishi, R.; Tomoeda, A.; Shimura, K.; Nishinari, K. Theory of jam-absorption driving. Transp. Res. Part B Methodol. 2013, 50, 116–129. [Google Scholar] [CrossRef]
- Taniguchi, Y.; Nishi, R.; Ezaki, T.; Nishinari, K. Jam-absorption driving with a car-following model. Phys. A Stat. Mech. Appl. 2015, 433, 304–315. [Google Scholar] [CrossRef]
- Johnson, S. Emergence: The Connected Lives of Ants, Brains, Cities, and Software; Simon and Schuster: New York, NY, USA, 2002; ISBN 9780684868752. [Google Scholar]
- Sage, A.P. System of Systems Engineering: Innovations for the 21st Century; John Wiley & Sons: Hoboken, NJ, USA, 2011; Volume 58. [Google Scholar]
- Andryszak, N.A.; Payne, T.L.; Dickens, J.C.; Moser, J.C.; Fisher, R.W. Antennal olfactory responsiveness of the Texas leaf cutting ant (Hymenoptera: Formicidae) to trail pheromone and its two alarm substances. J. Entomol. Sci. 1990, 25, 593–600. [Google Scholar] [CrossRef]
- Czaczkes, T.J.; Grüter, C.; Ellis, L.; Wood, E.; Ratnieks, F.L.W. Ant foraging on complex trails: Route learning and the role of trail pheromones in Lasius niger. J. Exp. Biol. 2013, 216, 188–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]









| Description | Symbol |
|---|---|
| Unique identity of a cell in the trail | |
| Presence or absence of an ant in the trail at time t | |
| Pheromone concentration in the trail at time t | |
| Pheromone concentration saturation level | |
| Unique identity of an ant in the simulation | |
| Velocity of the at time | |
| Position of the at time | |
| The minimum velocity of an ant towards the cell with no pheromone and no other ant | |
| Number of ants in simulation | |
| Stochastic parameter in velocity reduction scenario | |
| Trail length | |
| Evaporation rate |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kasture, P.; Nishimura, H. Analysis of Cooperative Perception in Ant Traffic and Its Effects on Transportation System by Using a Congestion-Free Ant-Trail Model. Sensors 2021, 21, 2393. https://doi.org/10.3390/s21072393
Kasture P, Nishimura H. Analysis of Cooperative Perception in Ant Traffic and Its Effects on Transportation System by Using a Congestion-Free Ant-Trail Model. Sensors. 2021; 21(7):2393. https://doi.org/10.3390/s21072393
Chicago/Turabian StyleKasture, Prafull, and Hidekazu Nishimura. 2021. "Analysis of Cooperative Perception in Ant Traffic and Its Effects on Transportation System by Using a Congestion-Free Ant-Trail Model" Sensors 21, no. 7: 2393. https://doi.org/10.3390/s21072393
APA StyleKasture, P., & Nishimura, H. (2021). Analysis of Cooperative Perception in Ant Traffic and Its Effects on Transportation System by Using a Congestion-Free Ant-Trail Model. Sensors, 21(7), 2393. https://doi.org/10.3390/s21072393