
Spatial Memory in a Spiking Neural Network with Robot Embodiment

 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
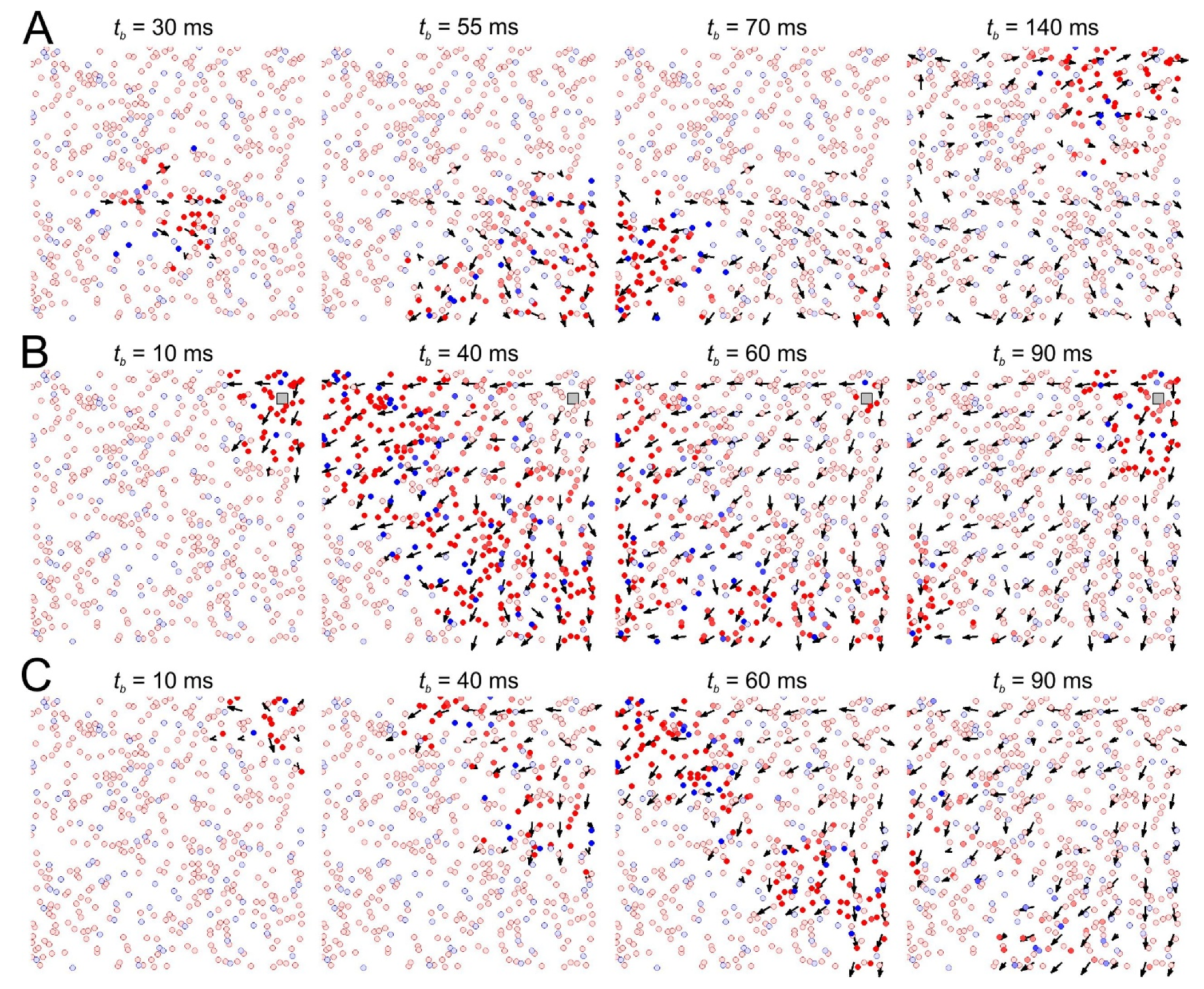
3.1. Synaptic Memory at Network Scale: Pathways of Spike Patches Match Potentiated Neural Couplings
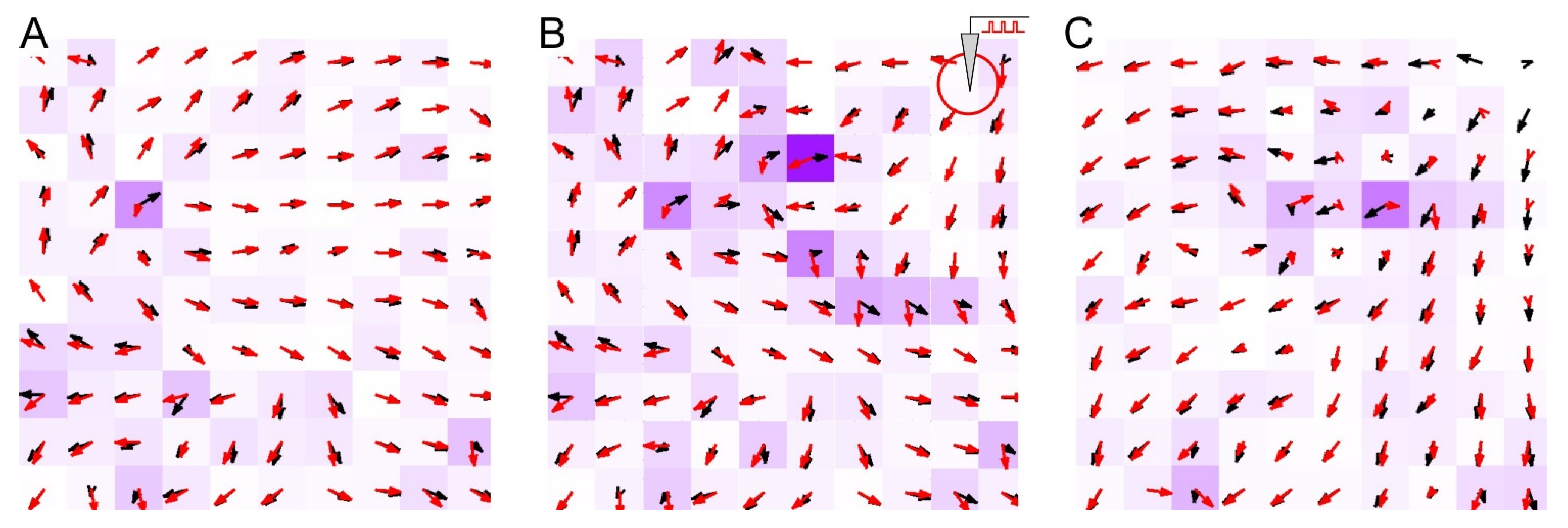
3.2. Stimulation of SNN Is an Iteration of Recording into Network Memory
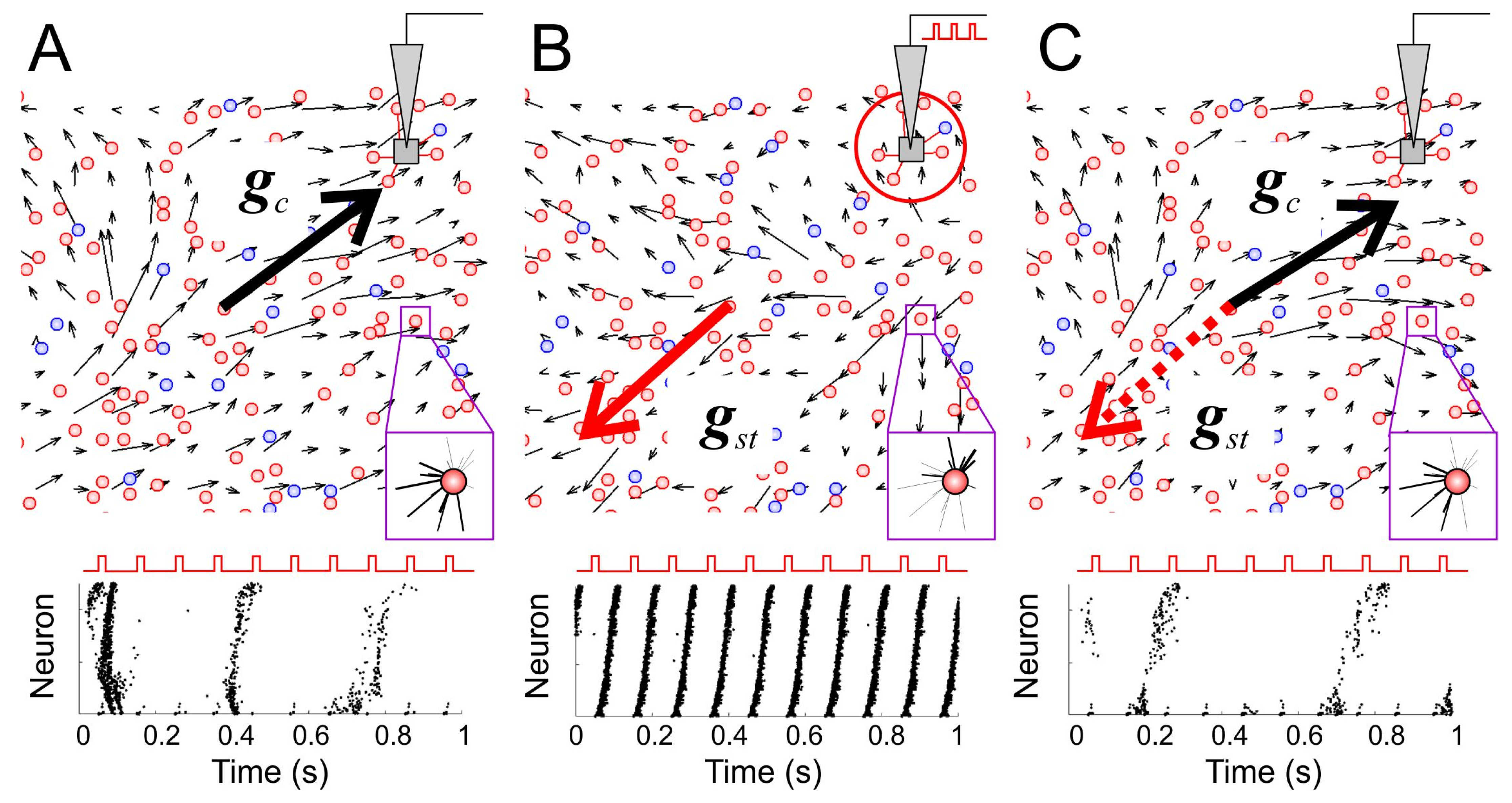
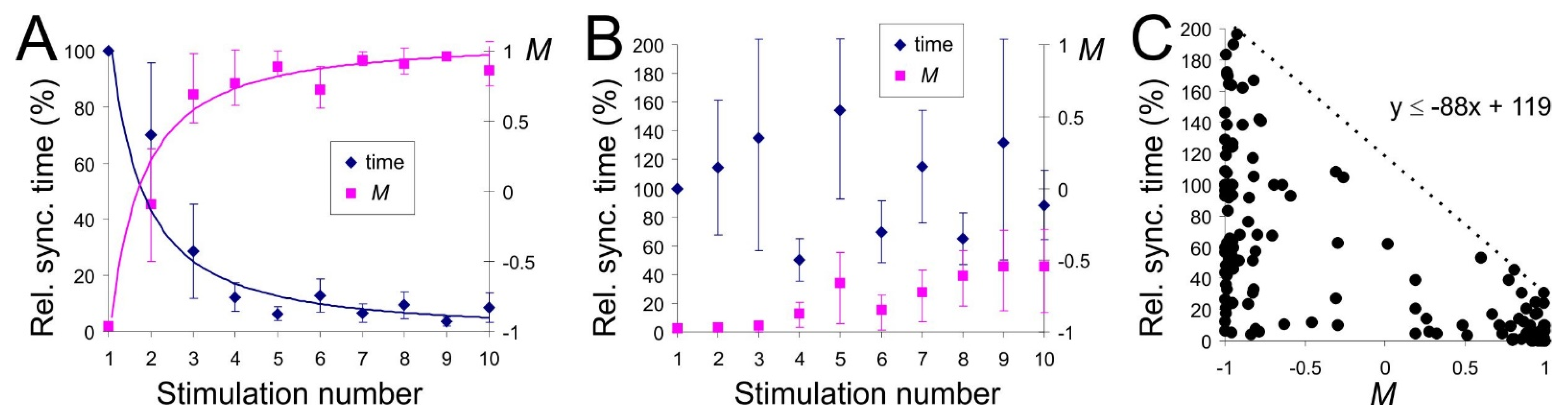
3.3. Learning and Forgetting Spatial Stimuli
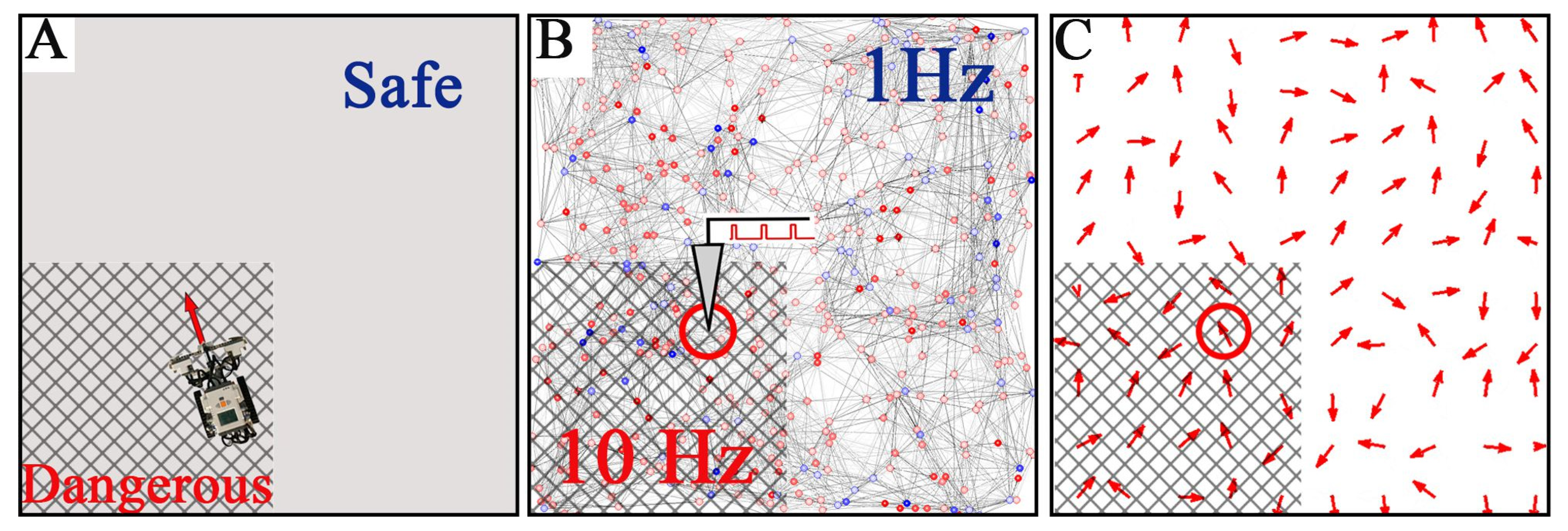
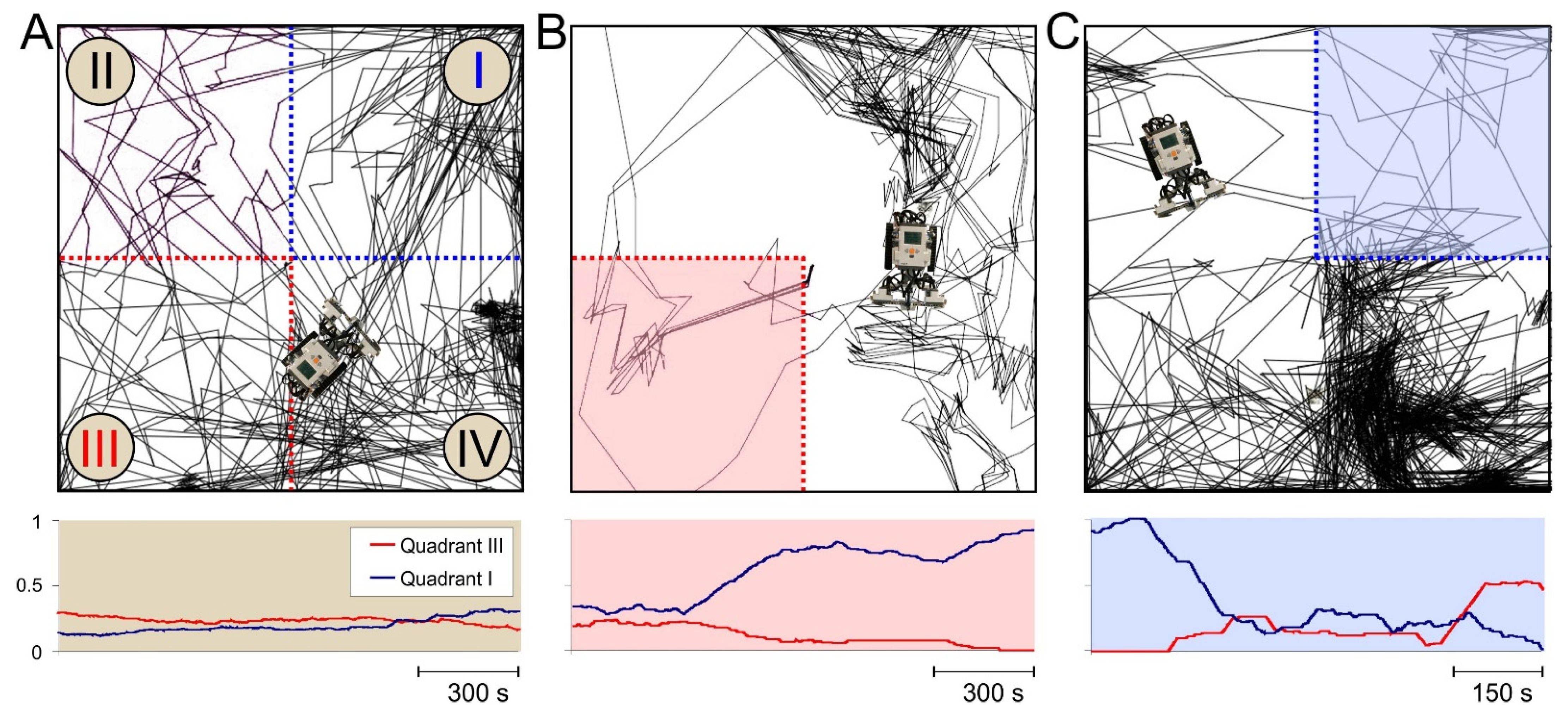
3.4. Spatial Memory with Negative Reinforcement: Embodiment of the SNN in a Robot
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Anokhin, K.V. The brain and memory: The biology of traces of time past. Her. Russ. Acad. Sci. 2010, 80, 237–242. [Google Scholar] [CrossRef]
- Frankland, P.W.; Bontempi, B. The organization of recent and remote memories. Nat. Rev. Neurosci. 2005, 6, 119–130. [Google Scholar] [CrossRef]
- Eichenbaum, H. Memory: Organization and control. Annu. Rev. Psychol. 2017, 68, 19–45. [Google Scholar] [CrossRef] [Green Version]
- Snoddy, G.S. Learning and stability: A psychophysiological analysis of a case of motor learning with clinical applications. J. Appl. Psychol. 1926, 10, 1–36. [Google Scholar] [CrossRef]
- Crossman, E.R.F.W. A theory of the acquisition of speed-skill. Ergonomics 1959, 2, 153–166. [Google Scholar] [CrossRef]
- Ebbinghaus, H. Memory: A Contribution to Experimental Psychology; Teachers College, Columbia University: New York, NY, USA, 1885. [Google Scholar]
- Kandel, E.R. The molecular biology of memory storage: A dialogue between genes and synapses. Science 2001, 294, 1030–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakkum, D.J.; Chao, Z.C.; Potter, S.M. Spatio-temporal electrical stimuli shape behavior of an embodied cortical network in a goal-directed learning task. J. Neural Eng. 2008, 5, 310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahaf, G.; Marom, S. Learning in networks of cortical neurons. J. Neurosci. 2001, 21, 8782–8788. [Google Scholar] [CrossRef] [Green Version]
- Pimashkin, A.; Gladkov, A.; Mukhina, I.; Kazantsev, V. Adaptive enhancement of learning protocol in hippocampal cultured networks grown on multielectrode arrays. Front. Neural Circuits 2013, 7, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baruchi, I.; Ben-Jacob, E. Towards neuro-memory-chip: Imprinting multiple memories in cultured neural networks. Phys. Rev. E 2007, 75, 50901. [Google Scholar] [CrossRef]
- Fernandez-Ruiz, A.; Makarov, V.A.; Herreras, O. Sustained increase of spontaneous input and spike transfer in the CA3-CA1 pathway following long-term potentiation in vivo. Front. Neural Circuits 2012, 6, 71. [Google Scholar] [CrossRef] [Green Version]
- Le Feber, J.; Stegenga, J.; Rutten, W.L.C. The Effect of slow electrical stimuli to achieve learning in cultured networks of rat cortical neurons. PLoS ONE 2010, 5, e8871. [Google Scholar] [CrossRef]
- Segev, R.; Shapira, Y.; Benveniste, M.; Ben-Jacob, E. Observations and modeling of synchronized bursting in two-dimensional neural networks. Phys. Rev. E 2001, 64, 11920. [Google Scholar] [CrossRef]
- Chao, Z.C.; Bakkum, D.J.; Wagenaar, D.A.; Potter, S.M. Effects of random external background stimulation on network synaptic stability after tetanization: A modeling study. Neuroinformatics 2005, 3, 263–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degterev, A.A.; Burtsev, M.S. Simulation of spontaneous activity in neuronal cultures with long-term plasticity. Math. Biol. Bioinf. 2015, 10, 234–244. [Google Scholar] [CrossRef]
- Bi, G.Q.; Poo, M.M. Synaptic modifications in cultured hippocampal neurons: Dependence on spike timing, synaptic strength, and postsynaptic cell type. J. Neurosci. 1998, 18, 10464–10472. [Google Scholar] [CrossRef] [PubMed]
- Sjöström, P.J.; Turrigiano, G.G.; Nelson, S.B. Rate, timing, and cooperativity jointly determine cortical synaptic plasticity. Neuron 2001, 32, 1149–1164. [Google Scholar] [CrossRef] [Green Version]
- Morrison, A.; Diesmann, M.; Gerstner, W. Phenomenological models of synaptic plasticity based on spike timing. Biol. Cybern. 2008, 98, 459–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, C.C.; Han, V.Z.; Sugawara, Y.; Grant, K. Synaptic plasticity in a cerebellum-like structure depends on temporal order. Nature 1997, 387, 278–281. [Google Scholar] [CrossRef]
- Kampa, B.M.; Letzkus, J.J.; Stuart, G.J. Dendritic mechanisms controlling spike-timing-dependent synaptic plasticity. Trends Neurosci. 2007, 30, 456–463. [Google Scholar] [CrossRef] [Green Version]
- Roberts, P.; Leen, T. Anti-hebbian spike-timing-dependent plasticity and adaptive sensory processing. Front. Comput. Neurosci. 2010, 4, 156. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, F.; Stiber, M. A simple model of cortical culture growth: Burst property dependence on network composition and activity. Biol. Cybern. 2014, 108, 423–443. [Google Scholar] [CrossRef]
- Chao, Z.C.; Bakkum, D.J.; Potter, S.M. Region-specific network plasticity in simulated and living cortical networks: Comparison of the center of activity trajectory (CAT) with other statistics. J. Neural Eng. 2007, 4, 294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gritsun, T.A.; le Feber, J.; Rutten, W.L.C. Growth dynamics explain the development of spatiotemporal burst activity of young cultured neuronal networks in detail. PLoS ONE 2012, 7, e43352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolman, E.C. Cognitive maps in rats and men. Psychol. Rev. 1948, 55, 189–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villacorta-Atienza, J.A.; Calvo Tapia, C.; Diez-Hermano, S.; Sanchez-Jimenez, A.; Lobov, S.; Krilova, N.; Murciano, A.; Lopez-Tolsa, G.E.; Pellon, R.; Makarov, V.A. Static internal representation of dynamic situations reveals time compaction in human cognition. J. Adv. Res. 2021, 28, 111–125. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, J.; Dostrovsky, J. The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat. Brain Res. 1971, 34, 171–175. [Google Scholar] [CrossRef]
- Hafting, T.; Fyhn, M.; Molden, S.; Moser, M.-B.; Moser, E.I. Microstructure of a spatial map in the entorhinal cortex. Nature 2005, 436, 801–806. [Google Scholar] [CrossRef] [PubMed]
- Krichmar, J.L.; Seth, A.K.; Nitz, D.A.; Fleischer, J.G.; Edelman, G.M. Spatial navigation and causal analysis in a brain-based device modeling cortical-hippocampal interactions. Neuroinformatics 2005, 3, 197–221. [Google Scholar] [CrossRef]
- Villacorta-Atienza, J.A.; Velarde, M.G.; Makarov, V.A. Compact internal representation of dynamic situations: Neural network implementing the causality principle. Biol. Cybern. 2010, 103, 285–297. [Google Scholar] [CrossRef]
- Borisyuk, R.; Chik, D.; Kazanovich, Y.; da Silva Gomes, J. Spiking neural network model for memorizing sequences with forward and backward recall. Biosystems 2013, 112, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Villacorta-Atienza, J.A.; Makarov, V.A. Neural network architecture for cognitive navigation in dynamic environments. IEEE Trans. Neural Netw. Learn. Syst. 2013, 24, 2075–2087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponulak, F.; Hopfield, J. Rapid, parallel path planning by propagating wavefronts of spiking neural activity. Front. Comput. Neurosci. 2013, 7, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobov, S.; Simonov, A.; Kastalskiy, I.; Kazantsev, V. Network response synchronization enhanced by synaptic plasticity. Eur. Phys. J. Spec. Top. 2016, 225, 29–39. [Google Scholar] [CrossRef]
- Lobov, S.A.; Zhuravlev, M.O.; Makarov, V.A.; Kazantsev, V.B. Noise enhanced signaling in STDP driven spiking-neuron network. Math. Model. Nat. Phenom. 2017, 12, 109–124. [Google Scholar] [CrossRef]
- Lobov, S.A. Generalized memory of STDP-driven spiking neural network. Math. Biol. Bioinform. 2019, 14, 649–664. [Google Scholar] [CrossRef] [Green Version]
- Izhikevich, E.M. Simple model of spiking neurons. IEEE Trans. Neural Netw. 2003, 14, 1569–1572. [Google Scholar] [CrossRef] [Green Version]
- Izhikevich, E.M. Which model to use for cortical spiking neurons? IEEE Trans. Neural Netw. 2004, 15, 1063–1070. [Google Scholar] [CrossRef]
- Tsodyks, M.; Pawelzik, K.; Markram, H. Neural networks with dynamic synapses. Neural Comput. 1998, 10, 821–835. [Google Scholar] [CrossRef]
- Song, S.; Miller, K.D.; Abbott, L.F. Competitive Hebbian learning through spike-timing-dependent synaptic plasticity. Nat. Neurosci. 2000, 3, 919. [Google Scholar] [CrossRef]
- Gong, P.; Van Leeuwen, C. Distributed dynamical computation in neural circuits with propagating coherent activity patterns. PLoS Comput. Biol. 2009, 5, e1000611. [Google Scholar] [CrossRef]
- Villacorta-Atienza, J.A.; Makarov, V.A. Wave-processing of long-scale information by neuronal chains. PLoS ONE 2013, 8, e57440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, J.H.C.; Gong, P. Associative learning of classical conditioning as an emergent property of spatially extended spiking neural circuits with synaptic plasticity. Front. Comput. Neurosci. 2014, 8, 79. [Google Scholar] [CrossRef] [Green Version]
- Lobov, S.A.; Mikhaylov, A.N.; Shamshin, M.; Makarov, V.A.; Kazantsev, V.B. Spatial properties of STDP in a self-learning spiking neural network enable controlling a mobile robot. Front. Neurosci. 2020, 14, 88. [Google Scholar] [CrossRef]
- Masquelier, T.; Guyonneau, R.; Thorpe, S.J. Spike timing dependent plasticity finds the start of repeating patterns in continuous spike trains. PLoS ONE 2008, 3, e1377. [Google Scholar] [CrossRef] [PubMed]
- Lobov, S.A.; Chernyshov, A.V.; Krilova, N.P.; Shamshin, M.O.; Kazantsev, V.B. Competitive learning in a spiking neural network: Towards an intelligent pattern classifier. Sensors 2020, 20, 500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazhanova, M.V.; Krylova, N.P.; Kazantsev, V.B.; Khramov, A.E.; Lobov, S.A. Synchronization in a network of spiking neural oscillators with plastic connectivity. Radiophys. Quantum Electron. 2020, 63, 298–309. [Google Scholar] [CrossRef]
- Chou, T.-S.; Bucci, L.; Krichmar, J. Learning touch preferences with a tactile robot using dopamine modulated STDP in a model of insular cortex. Front. Neurorobot. 2015, 9, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvo Tapia, C.; Tyukin, I.Y.; Makarov, V.A. Fast social-like learning of complex behaviors based on motor motifs. Phys. Rev. E 2018, 97, 052308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bing, Z.; Meschede, C.; Röhrbein, F.; Huang, K.; Knoll, A.C. A survey of robotics control based on learning-inspired spiking neural networks. Front. Neurorobot. 2018, 12, 35. [Google Scholar] [CrossRef] [Green Version]
- Calvo Tapia, C.; Villacorta-Atienza, J.A.; Diez-Hermano, S.; Khoruzhko, M.; Lobov, S.A.; Potapov, I.; Sanchez-Jimenez, A.; Makarov, V.A. Semantic knowledge representation for strategic interactions in dynamic situations. Front. Neurorobot. 2020, 14, 4. [Google Scholar] [CrossRef] [PubMed]
- Bing, Z.; Meschede, C.; Chen, G.; Knoll, A.; Huang, K. Indirect and direct training of spiking neural networks for end-to-end control of a lane-keeping vehicle. Neural Netw. 2020, 121, 21–36. [Google Scholar] [CrossRef] [PubMed]
- Quiroga, Q.R.; Panzeri, S. Principles of Neural Coding; CRC Press: Boca Raton, FL, USA, 2013. [Google Scholar]
- O’Keefe, J.; Nadel, L. Précis of O’Keefe & Nadel’s the hippocampus as a cognitive map. Behav. Brain Sci. 1979, 2, 487–494. [Google Scholar]
- Rosenbloom, P.S. A cognitive odyssey: From the power law of practice to a general learning mechanism and beyond. Tutor. Quant. Methods Psychol. 2006, 2, 43–51. [Google Scholar] [CrossRef]
- Anderson, R.B. The power law as an emergent property. Mem. Cognit. 2001, 29, 1061–1068. [Google Scholar] [CrossRef] [Green Version]
- Murre, J.M.J.; Chessa, A.G. Power laws from individual differences in learning and forgetting: Mathematical analyses. Psychon. Bull. Rev. 2011, 18, 592–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lobov, S.A.; Zharinov, A.I.; Makarov, V.A.; Kazantsev, V.B. Spatial Memory in a Spiking Neural Network with Robot Embodiment. Sensors 2021, 21, 2678. https://doi.org/10.3390/s21082678
Lobov SA, Zharinov AI, Makarov VA, Kazantsev VB. Spatial Memory in a Spiking Neural Network with Robot Embodiment. Sensors. 2021; 21(8):2678. https://doi.org/10.3390/s21082678
Chicago/Turabian StyleLobov, Sergey A., Alexey I. Zharinov, Valeri A. Makarov, and Victor B. Kazantsev. 2021. "Spatial Memory in a Spiking Neural Network with Robot Embodiment" Sensors 21, no. 8: 2678. https://doi.org/10.3390/s21082678
APA StyleLobov, S. A., Zharinov, A. I., Makarov, V. A., & Kazantsev, V. B. (2021). Spatial Memory in a Spiking Neural Network with Robot Embodiment. Sensors, 21(8), 2678. https://doi.org/10.3390/s21082678