
Adaptive SNN for Anthropomorphic Finger Control




Abstract
:1. Introduction
2. Related Works
2.1. SMA Actuators
2.2. Adaptive SNNs
2.3. SNNs in Robotics
2.4. Adaptive SNN for Motion Control
2.5. Proposed Concept
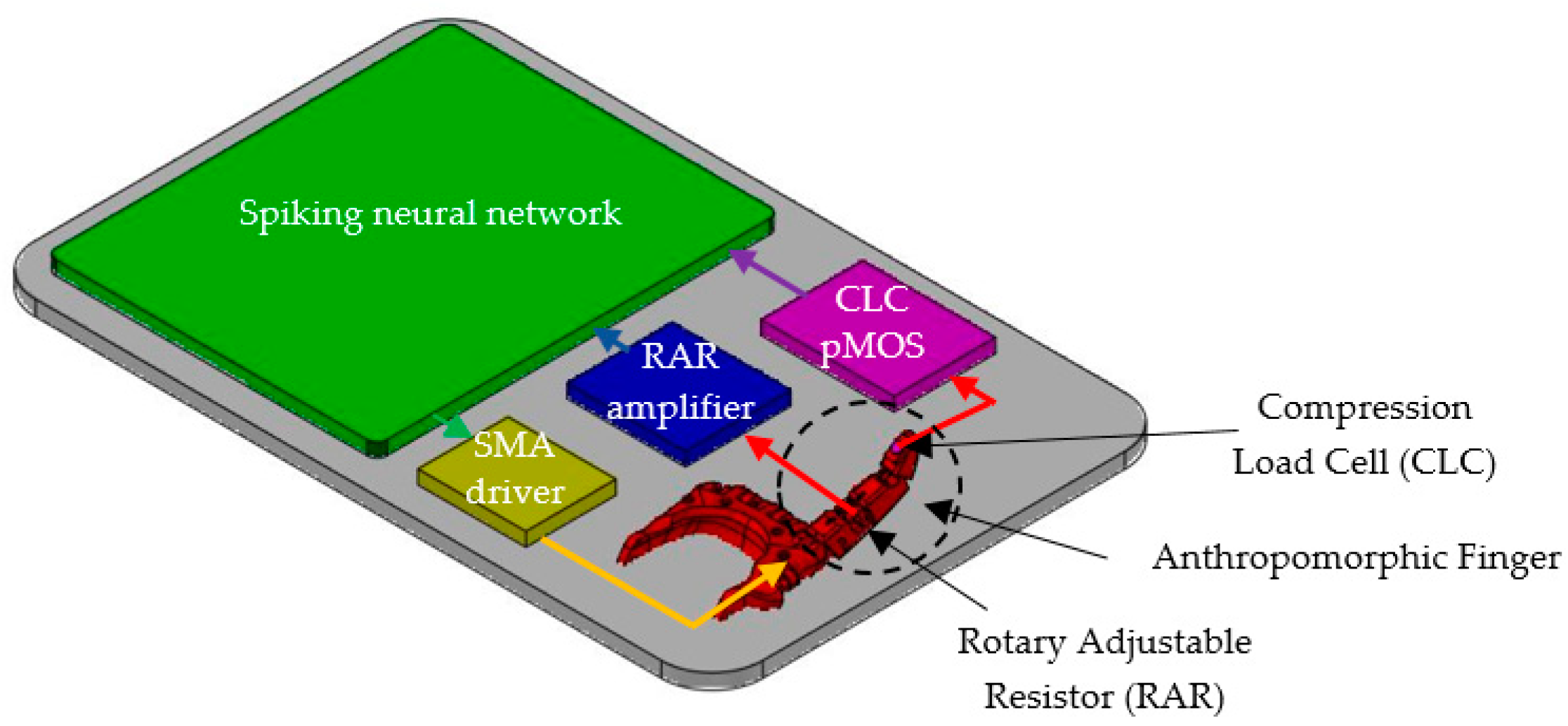
3. Bioinspired System Design
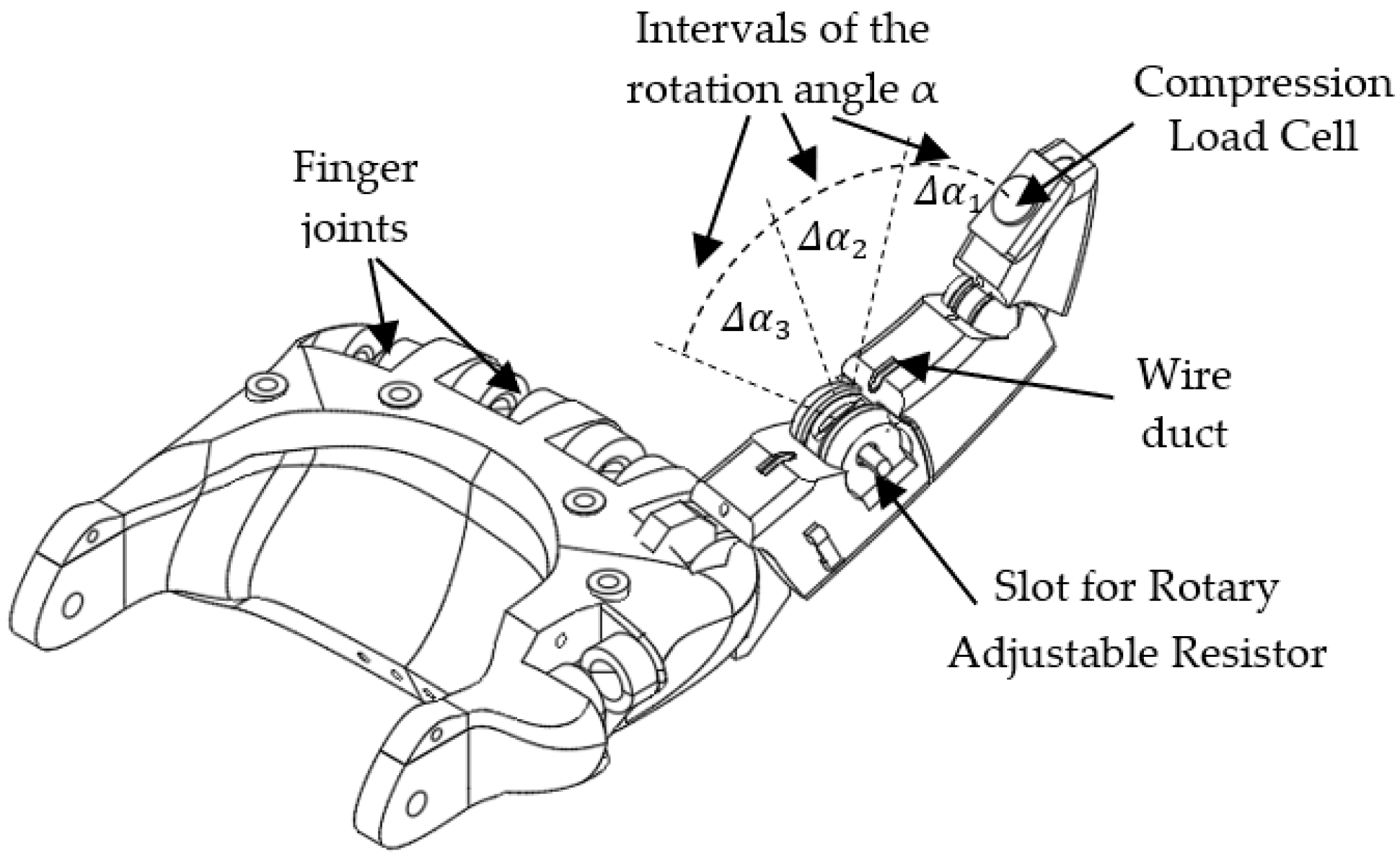
3.1. Artificial Finger
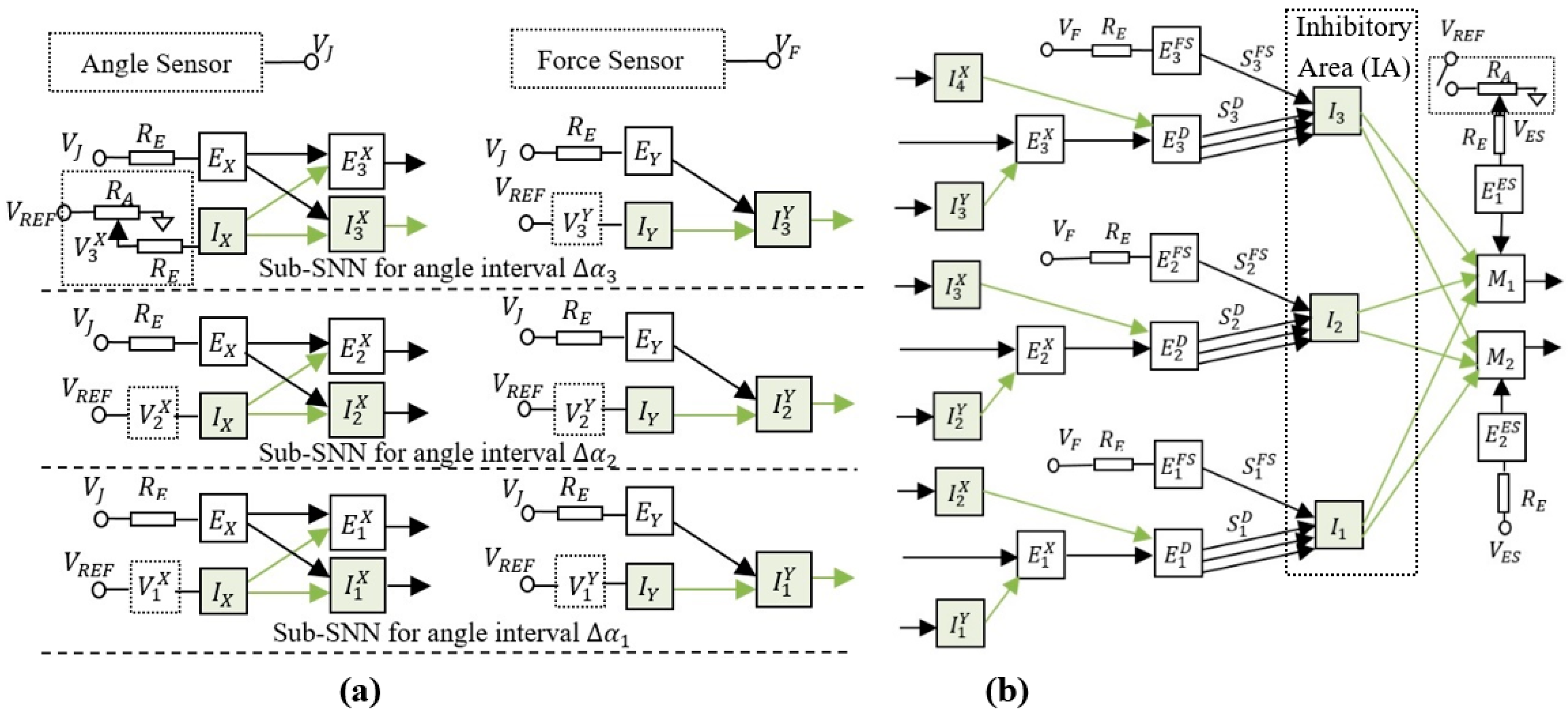
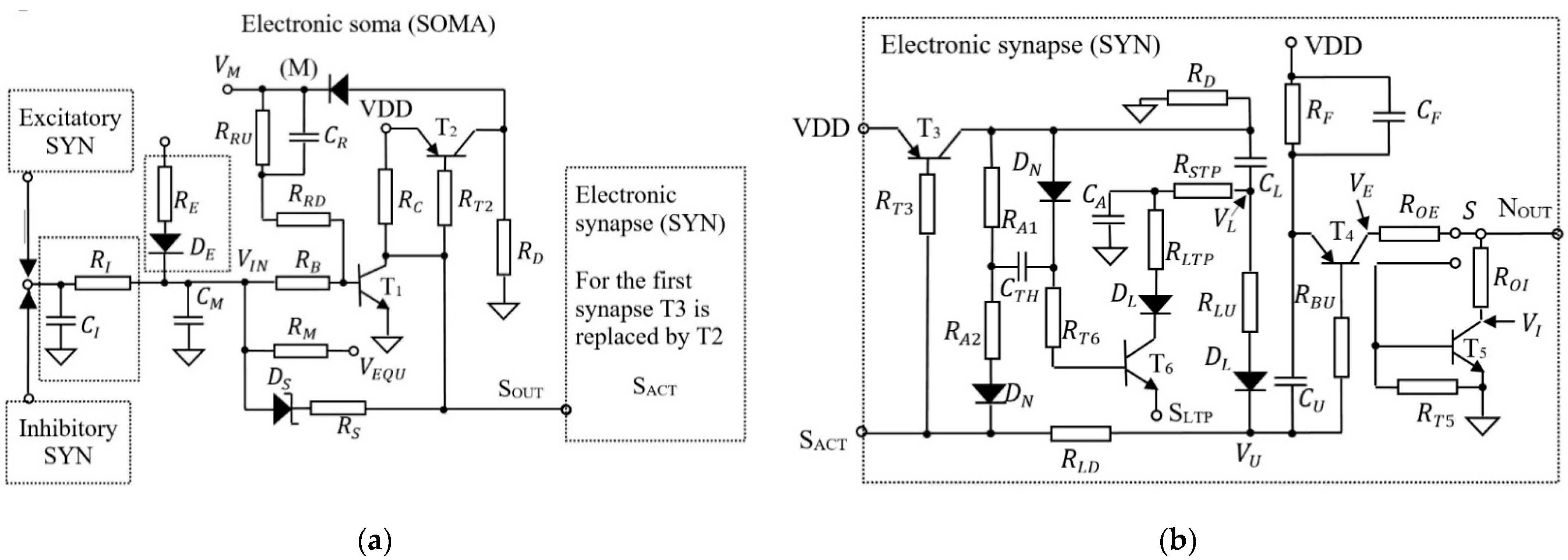
3.2. The Structure of the Adaptive SNN
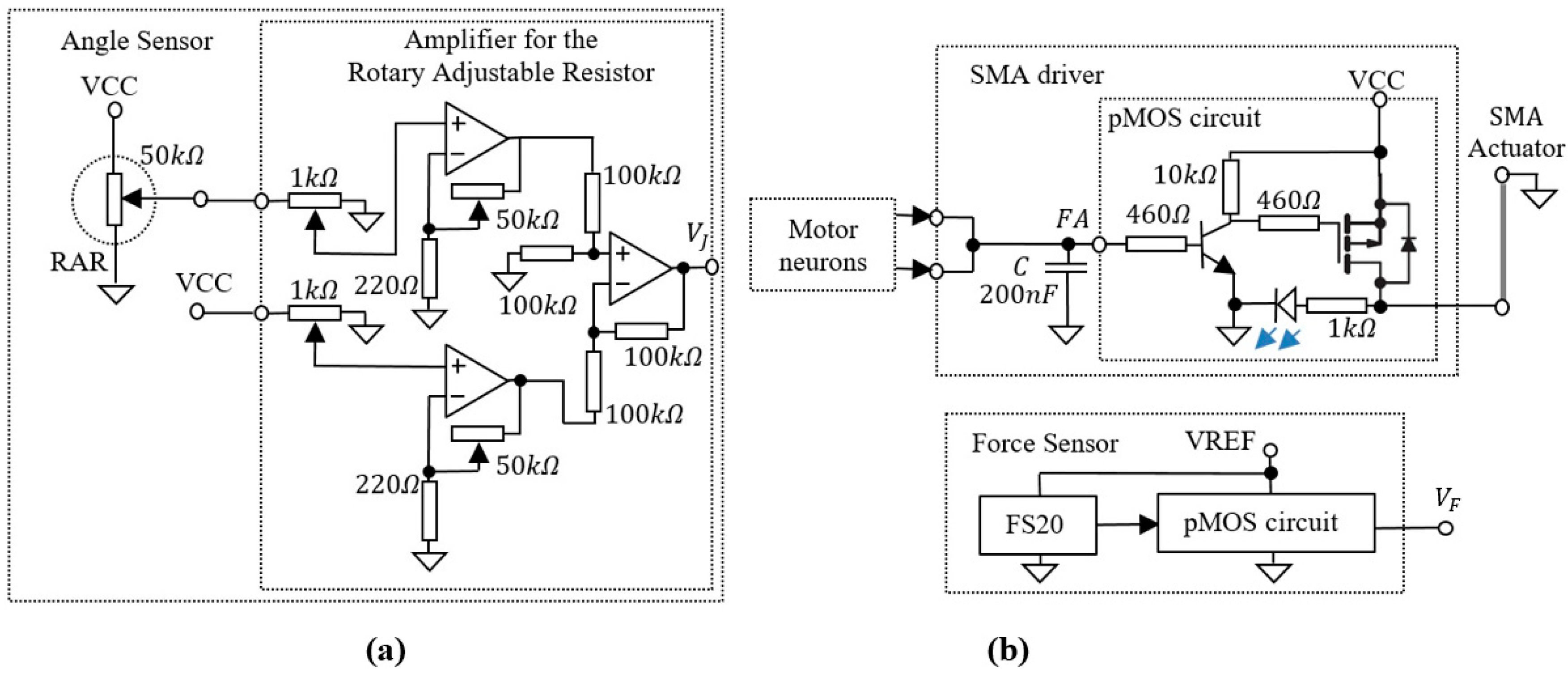
3.3. Auxiliary Electronics
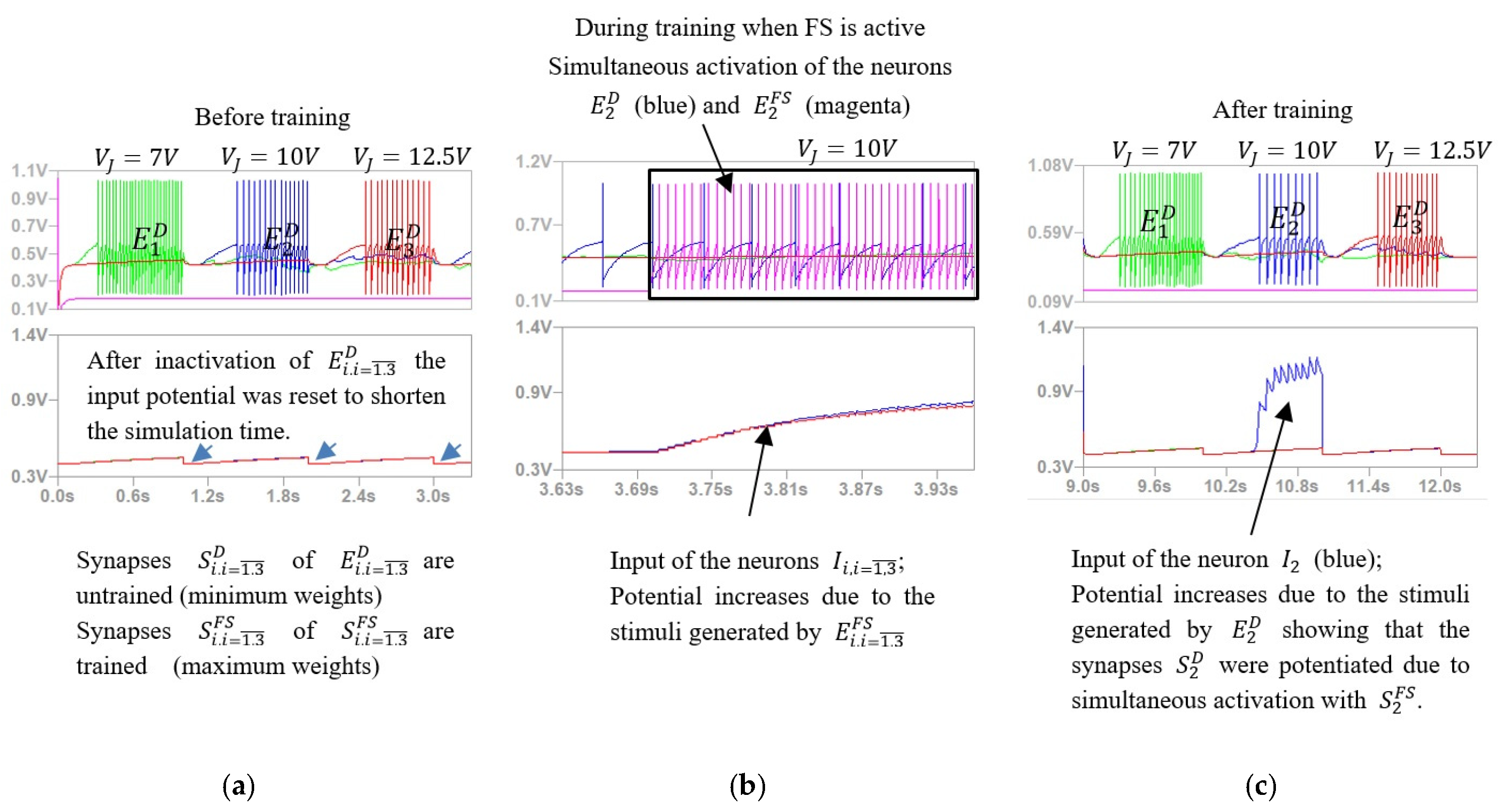
4. Evaluation by Simulation of the SNN Activity
5. Experimental Investigation
5.1. Experimental Setup
5.2. Experiments Overview
5.3. Experimental Results
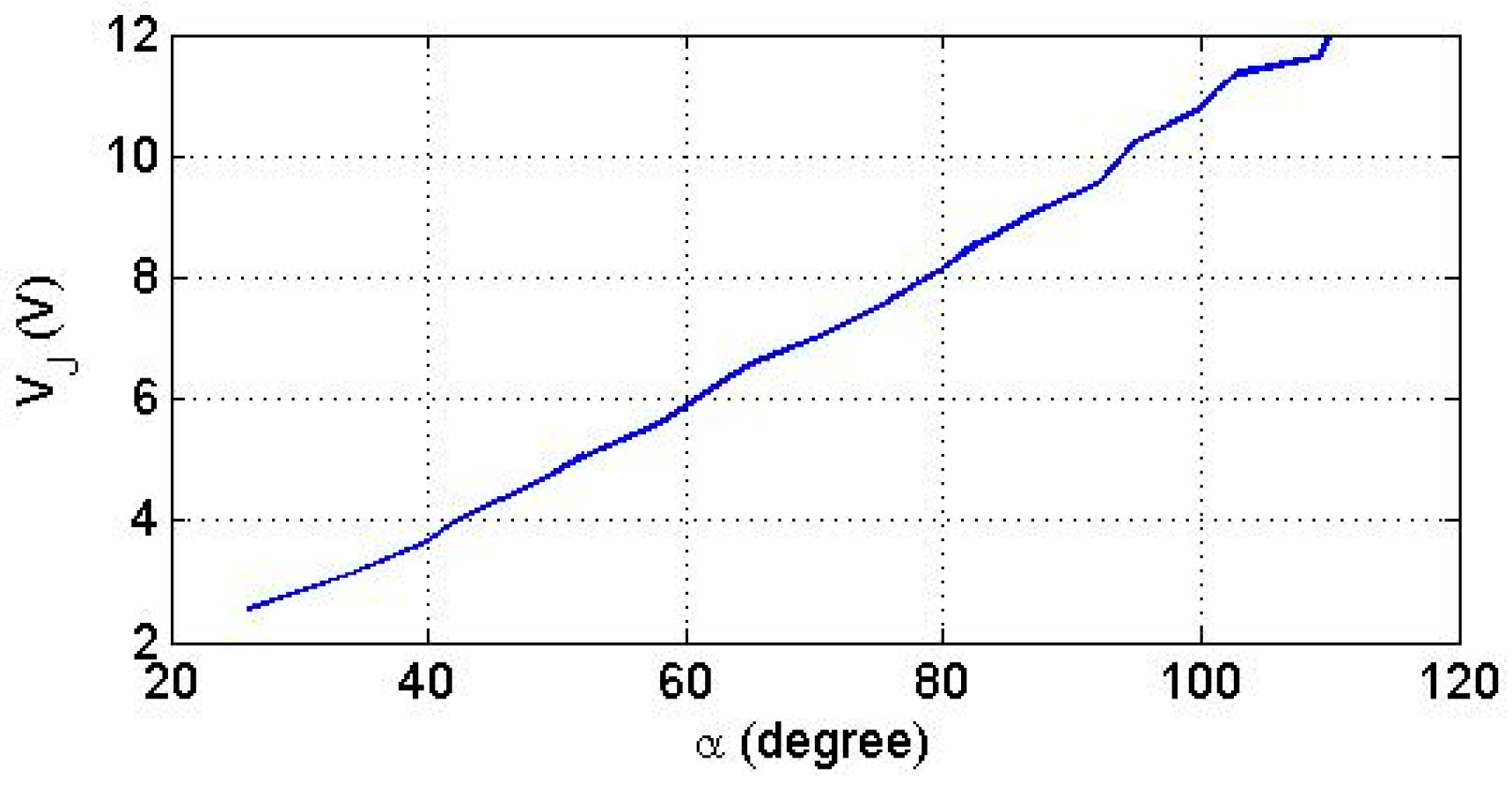
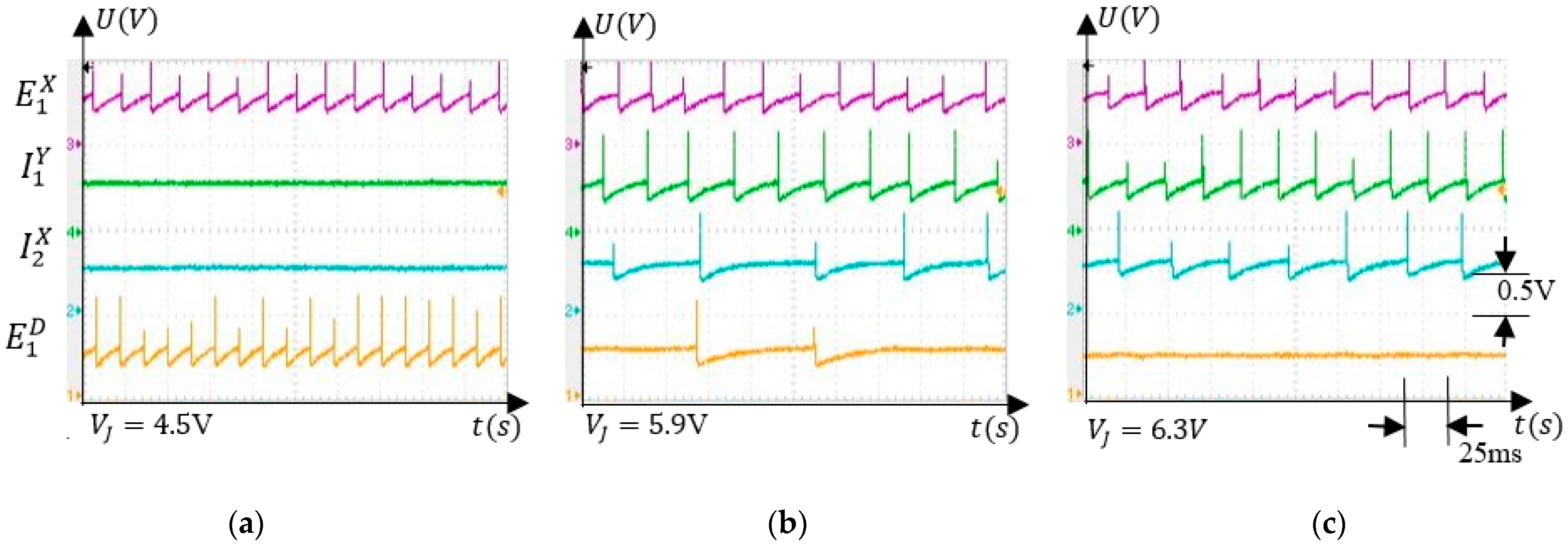
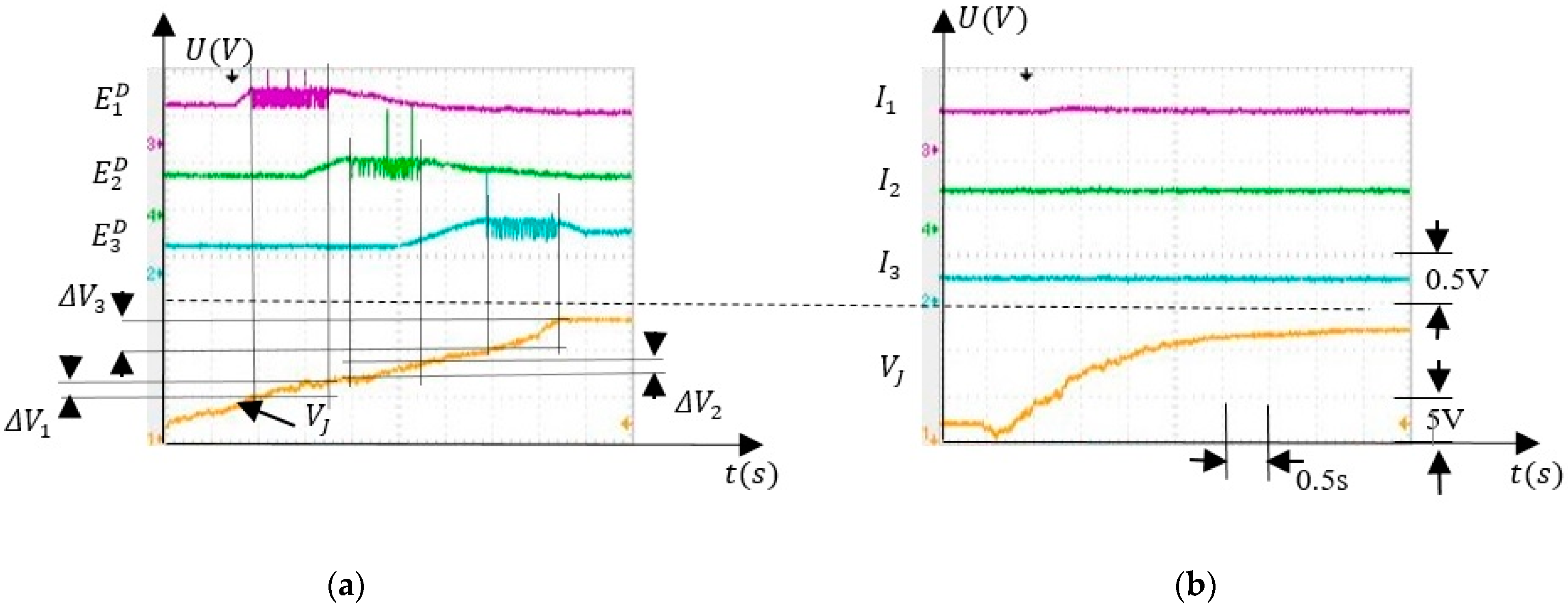
5.3.1. Voltage Interval Selectivity
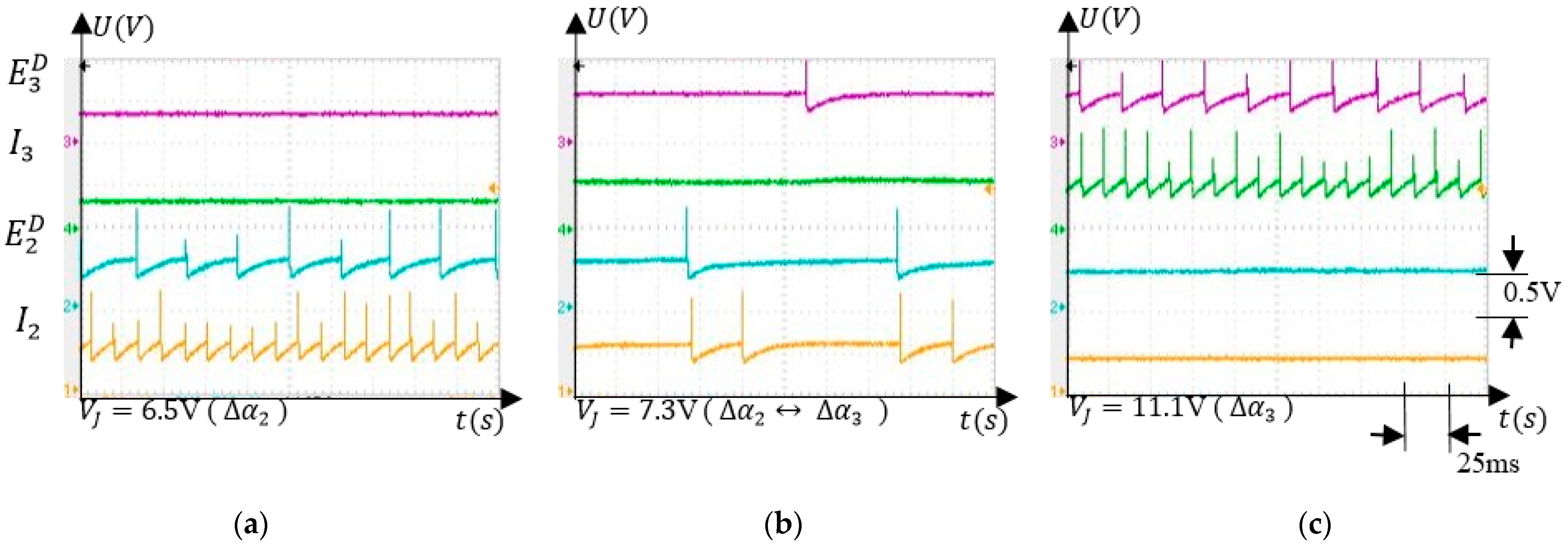
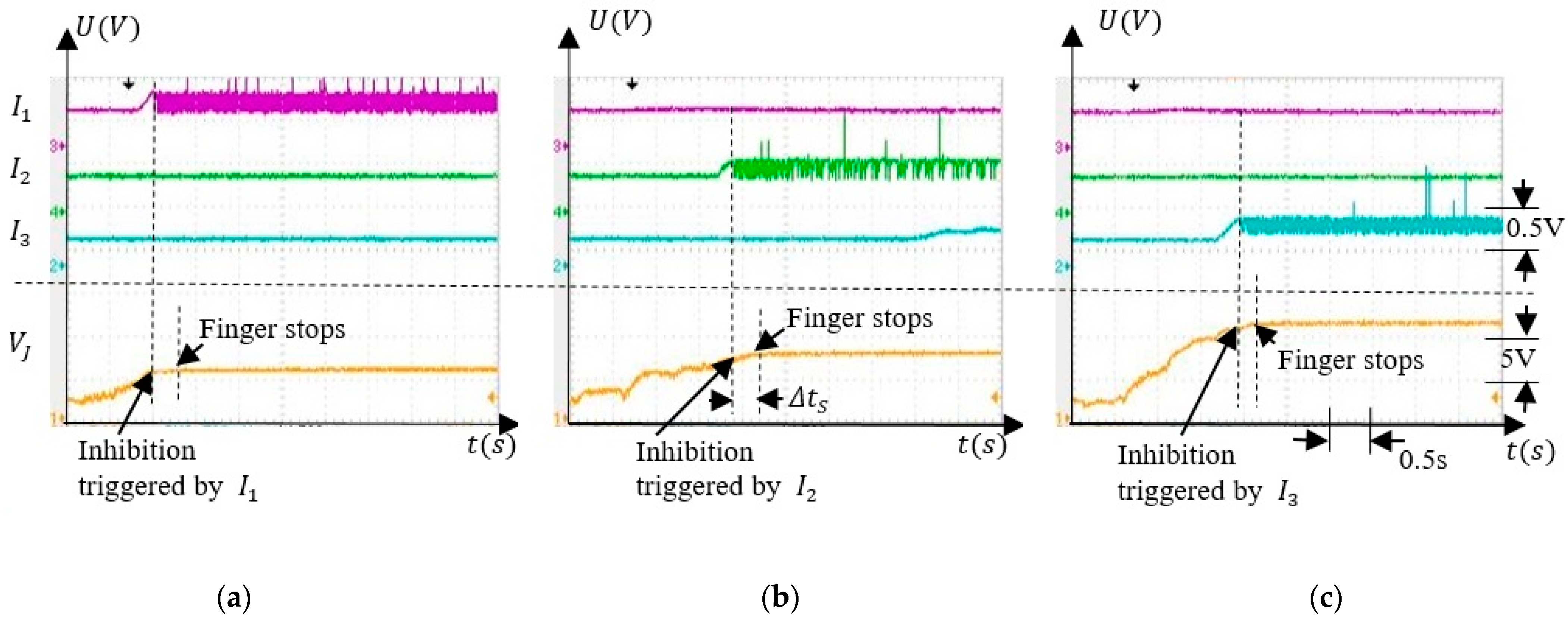
5.3.2. Associative Learning
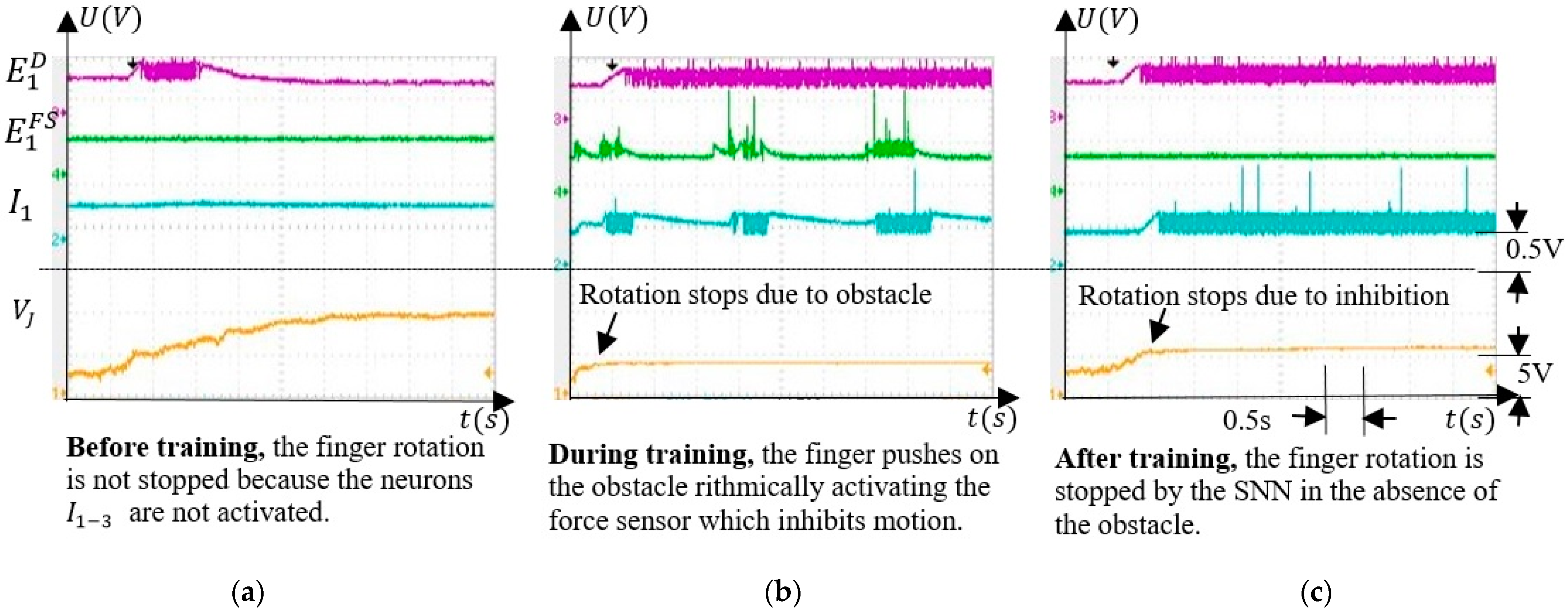
5.3.3. Finger Operation
5.4. Discussions
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Param. | Value | Param. | Value |
|---|---|---|---|
| RRU | 20 kΩ | T1 | BC848C |
| RRD | 1 kΩ | T2 | BC857C |
| RIN | 220 kΩ | DS | BAR43 |
| RB | 6.2 kΩ | DN | 1N4148 |
| RC | 10 kΩ | DL | BAS45A |
| RD | 1 MΩ | CR | 10 nF |
| RM | 1 MΩ | CI | 1 μF |
| RS | 47 Ω | CM | 100 nF |
| RT2 | 10k Ω |
| Param. | Value | Param. | Value |
|---|---|---|---|
| RD | 1 MΩ | CTH | 10 nF |
| RF | 47 kΩ | CA | 47 nF |
| RT3 | 10 kΩ | CL | 2.2 μF |
| RA1 | 10 kΩ | CU | 221 pF |
| RA2 | 1 kΩ | CF | 1 μF |
| RT6 | 470 Ω | T3 | BC857C |
| RSTP | 10 kΩ | T4 | BC857C |
| RLTP | 470 Ω | T5 | BC848C |
| ROE | 1.8 kΩ | DN | 1N4148 |
| RLU | 1 MΩ | DL | BAS45A |
| RLD | 470 kΩ | RE | 560 kΩ |
| RBU | 10 kΩ | RA | 5 kΩ |
| ROI | 470 Ω | ||
| RT5 | 47 kΩ |
References
- Bing, Z.; Meschede, C.; Röhrbein, F.; Huang, K.; Knoll, A.C. A survey of robotics control based on learning-inspired spiking neural networks. Front. Neurorobot. 2018, 12, 35. [Google Scholar] [CrossRef] [Green Version]
- Al-Falahe, N.A.; Nagaoka, M.; Vallbo, A.B. Response profiles of human muscle afferents during active finger movements. Brain 1990, 113, 325–346. [Google Scholar] [CrossRef] [PubMed]
- Hulliger, M. The mammalian muscle spindle and its central control. Rev. Physiol. Biochem. Pharmacol. 1984, 101, 1–110. [Google Scholar] [PubMed]
- Blum, K.P.; Lamotte D’Incamps, B.; Zytnicki, D.; Ting, L.H. Force encoding in muscle spindles during stretch of passive muscle. PLoS Comput. Biol. 2017, 13, e1005767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, R.; Dalton, B.; Blouin, J.-S.; Inglis, T. Precise coding of ankle angle and velocity by human calf muscle spindles. Neuroscience 2017, 349, 98–105. [Google Scholar] [CrossRef]
- Vallbo, A.B. Afferent discharge from human muscle spindles in non-contracting muscles. Steady state impulse frequency as a function of joint angle. Acta Psychiatr. Scand. 1974, 90, 303–318. [Google Scholar] [CrossRef] [PubMed]
- Vallbo, A.B.; Al-Falahe, N.A. Human muscle spindle response in a motor learning task. J. Physiol. 1990, 421, 553–568. [Google Scholar] [CrossRef]
- Dimitriou, M. Enhanced muscle afferent signals during motor learning in humans. Curr. Biol. 2016, 26, 1062–1068. [Google Scholar] [CrossRef] [Green Version]
- Grillner, S.; Robertson, B.; Kotaleski, J.H. Basal ganglia—A motion perspective. Compr. Physiol. 2020, 10, 1241–1275. [Google Scholar]
- Hulea, M. Analogue Electronic Circuit for Implementation of an Artificial Neuron. Patent RO126249 (A2), 29 June 2018. [Google Scholar]
- Hulea, M.; Ghassemlooy, Z.; Rajbhandari, S.; Younus, O.I.; Barleanu, A. Optical axons for electro-optical neural networks. Sensors 2020, 20, 6119. [Google Scholar] [CrossRef]
- Lester, B.; Baxevanis, T.; Chemisky, Y.; Lagoudas, D. Review and perspectives: Shape memory alloy composite systems. Acta Mech. 2015, 226, 3907–3960. [Google Scholar] [CrossRef] [Green Version]
- Mohd, J.; Leary, J.M.; Subic, A.; Gibson, M. A review of shape memory alloy research, applications and opportunities. Mater. Des. 2014, 56, 1078–1113. [Google Scholar] [CrossRef]
- Coral, W.; Rossi, C.; Colorado, J.; Barrientos, A. SMA-Based Muscle-Like Actuation in Biologically Inspired Robots: A State of the Art Review. In Smart Actuation and Sensing Systems–Recent Advances and Future Challenges; IntechOpen: London, UK, 2012; pp. 52–82. [Google Scholar]
- Hulea, M.; Caruntu, C.F. Spiking Neural Network for Controlling the Artificial Muscles of a Humanoid Robotic Arm. In Proceedings of the 18th Conference on System Theory, Control and Computing, Sinaia, Romania, 17–19 October 2014; pp. 163–168. [Google Scholar]
- Hulea, M. Bio-Inspired Control Method Based on Spiking Neural Networks and SMA Actuator Wires for LASER Spot Tracking. In Nature-Inspired Computing for Control Systems; Hiram, P., Ed.; Springer International Publishing: Cham, Switzerland, 2015; pp. 13–38. [Google Scholar]
- Hulea, M.; Burlacu, A.; Caruntu, C.F. Intelligent motion planning and control for robotic joints using bio-inspired spiking neural networks. Int. J. Hum. Robot. 2019, 16, 1950012. [Google Scholar] [CrossRef]
- Quintanar-Guzmán, S.; Kannan, S.; Olivares-Mendez, M.A.; Voos, H. Lightweight Robotic Arm Actuated by Shape Memory Alloy (SMA) Wires. In Proceedings of the 2016 8th International Conference on Electronics, Computers and Artificial Intelligence, Ploiesti, Romania, 30 June–2 July 2016. [Google Scholar]
- Grillner, S. The motor infrastructure from ion channels to neuronal networks. Nat. Rev. Neurosci. 2003, 4, 573–586. [Google Scholar] [CrossRef] [PubMed]
- Hulea, M.; Uleru, G.; Burlacu, A.; Caruntu, C. Bioinspired SNN for Robotic Joint Control. In Proceedings of the IEEE International Conference on Automation, Quality and Testing, Robotics, Cluj-Napoca, Romania, 21–23 May 2020; pp. 1–5. [Google Scholar]
- Golgouneh, A.; Holschuh, B.; Dunne, L. A Controllable Biomimetic SMA-Actuated Robotic Arm. In Proceedings of the 2020 8th IEEE RAS/EMBS International Conference for Biomedical Robotics and Biomechatronics, New York, NY, USA, 29 November–1 December 2020. [Google Scholar]
- Vasquez Tieck, J.C.; Donat, H.; Kaiser, J.; Peric, I.; Ulbrich, S.; Roennau, A.; Zöllner, M.; Dillmann, R. Towards Grasping with Spiking Neural Networks for Anthropomorphic Robot Hands. In Lecture Notes in Computer Science; Springer: Cham, Switzerland, 2017; Volume 10613, pp. 43–51. [Google Scholar]
- Arena, P.; De Fiore, S.; Patané, L.; Pollino, M.; Ventura, C. STDP-based behavior learning on the TriBot robot. In Proceedings of the Bioengineered and Bioinspired Systems IV, Dresden, Germany, 20 May 2009. [Google Scholar]
- Andrianesis, K.; Tzes, A. Development and control of a multifunctional prosthetic hand with shape memory alloy actuators. J. Intell. Robot. Syst. 2014, 78, 257–289. [Google Scholar] [CrossRef]
- Ulloa, C.C.; Terrile, S.; Barrientos, A. Soft underwater robot actuated by shape-memory alloys jellyrobcib for path tracking through fuzzy visual control. Appl. Sci. 2020, 10, 7160. [Google Scholar] [CrossRef]
- Garcia-Cordova, F.; Lopez-Coronado, J.; Guerrero-Gonzalez, A. Design of an Anthropomorphic Finger Using Shape Memory Alloy Springs. In Proceedings of the IEEE International Conference on Systems, Man, and Cybernetics (Cat. No.99CH37028), Tokyo, Japan, 12–15 October 1999; pp. 794–799. [Google Scholar]
- Sugita, K.; Tanaka, D.; Ono, S.; Chiba, S.; Iwata, K.; Han, Y.; Takato, M.; Uchikoba, F.; Saito, K. SMA Actuator and Pulse-Type Hardware Neural Networks IC for Fast Walking Motion of Insect-Type MEMS Microrobot. In Proceedings of the IEEE International Conference on Advanced Intelligent Mechatronics, Banff, AB, Canada, 12–15 July 2016; pp. 431–435. [Google Scholar]
- Brailovski, V.; Terriault, P.; Georges, T.; Coutu, D. SMA actuators for morphing wings. Phys. Procedia 2010, 10, 197–203. [Google Scholar] [CrossRef] [Green Version]
- Koh, J.-S.; Kim, S.-W.; Noh, M.; Cho, K.-J. Biologically inspired robots using Smart Composite Microstructures. In Proceedings of the 8th International Conference on Ubiquitous Robots and Ambient Intelligence, Incheon, Korea, 23–26 November 2011; p. 871. [Google Scholar]
- Yu, Q.; Ma, C.; Song, S.; Zhang, G.; Dang, J.; Tan, K. Constructing Accurate and Efficient Deep Spiking Neural Networks with Double-threshold and Augmented Schemes. arXiv 2020, arXiv:2005.03231. [Google Scholar]
- Taherkhani, A.; Belatreche, A.; Li, Y.; Cosma, G.; Maguire, L.; McGinnity, T.M. A Review of Learning in Biologically Plausible Spiking Neural Networks. Neural Netw. 2020, 122, 253–272. [Google Scholar] [CrossRef]
- López, L.J.; Del Ser, J.; Bifet, A.; Kasabov, N. Spiking Neural Networks and online learning: An overview and perspectives. Neural Netw. 2020, 121, 88–100. [Google Scholar]
- Wang, X.; Lin, X.; Dang, X. Supervised learning in spiking neural networks: A review of algorithms and evaluations. Neural Netw. 2020, 125, 258–280. [Google Scholar] [CrossRef]
- Kim, J.; Kim, K.; Kim, J. Unifying Activation- and Timing-based Learning Rules for Spiking Neural Networks. arXiv 2020, arXiv:2006.02642. [Google Scholar]
- Hao, Y.; Huang, X.; Dong, M.; Xu, B. A biologically plausible supervised learning method for spiking neural networks using the symmetric STDP rule. Neural Netw. 2019, 121, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Lobov, S.A.; Mikhaylov, A.N.; Shamshin, M.; Makarov, V.A.; Kazantsev, V.B. Spatial Properties of STDP in a Self-Learning Spiking Neural Network Enable Controlling a Mobile Robot. Front. Neurosci. 2020, 14, 88. [Google Scholar] [CrossRef]
- Demin, V.; Nekhaev, D.; Surazhevsky, I.A.; Nikiruy, K.; Emelyanov, A.; Nikolaev, S.; Rylkov, V.; Kovalchuk, M.V. Necessary conditions for STDP-based pattern recognition learning in a memristive spiking neural network. Neural Netw. 2020, 134, 64–75. [Google Scholar] [CrossRef]
- Shi, M.; Zhang, T.; Zeng, Y. Corrigendum: A Curiosity-Based Learning Method for Spiking Neural Networks. Front. Comput. Neurosci. 2020, 14, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shim, Y.; Philippides, A.; Staras, K.; Husbands, P. Unsupervised Learning in an Ensemble of Spiking Neural Networks Mediated by ITDP. PLoS Comput. Biol. 2016, 12, e1005137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, L.; Zhao, Z.; Wang, L.; Wang, Y. Efficient and hardware-friendly methods to implement competitive learning for spiking neural networks. Neural Comput. Appl. 2020, 32, 13479–13490. [Google Scholar] [CrossRef]
- Kumarasinghe, K.; Kasabov, N.; Taylor, D. Deep learning and deep knowledge representation in Spiking Neural Networks for Brain-Computer Interface. Neural Netw. 2020, 121, 169–185. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.; Shrestha, A.; Zhao, Z.; Qiu, Q. Exploiting Neuron and Synapse Filter Dynamics in Spatial Temporal Learning of Deep Spiking Neural Network. In Proceedings of the Twenty-Ninth International Joint Conference on Artificial Intelligence (IJCAI), Yokohama, Japan, 7–15 January 2021. [Google Scholar]
- Tang, G.; Kumar, N.; Michmizos, K.P. Reinforcement Co-Learning of Deep and Spiking Neural Networks for Energy-Efficient Mapless Navigation with Neuromorphic Hardware. In Proceedings of the International Conference on Intelligent Robots and Systems (IROS), Las Vegas, NV, USA, 24 January 2021 pp. 6090–6097. [Google Scholar]
- Payvand, M.; Fouda, M.E.; Kurdahi, F.; Eltawil, A.; Neftci, E. On-Chip Error-triggered Learning of Multi-layer Memristive Spiking Neural Networks. IEEE J. Emerg. Sel. Top. Circuits Syst. 2020, 10, 522–535. [Google Scholar] [CrossRef]
- Skatchkovsky, N.; Jang, H.; Simeone, O. Spiking Neural Networks—Part II: Detecting Spatio-Temporal Patterns. IEEE Commun. Lett. 2021. [Google Scholar] [CrossRef]
- Cheng, X.; Zhang, T.; Jia, S.; Xu, B. Finite Meta-Dynamic Neurons in Spiking Neural Networks for Spatio-temporal Learning. arXiv 2020, arXiv:2010.03140. [Google Scholar]
- Baagyere, E.; Moses, A.; Zhen, Q.; Oyetunji, E.; Qin, Z. A Multi-Spiking Neural Network Learning Model for Data Classification. IEEE Access 2020, 8, 72360–72371. [Google Scholar]
- Maciąg, P.; Kryszkiewicz, M.; Bembenik, R.; López, L.J.; Del Ser, J. Unsupervised Anomaly Detection in Stream Data with Online Evolving Spiking Neural Networks. Neural Netw. 2019, 139, 118–139. [Google Scholar] [CrossRef] [PubMed]
- Bullock, D.; Grossberg, S.; Guenther, F.H. A self-organizing neural model of motor equivalent reaching and tool use by a multijoint arm. J. Cogn. Neurosci. 1993, 5, 408–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouganis, A.; Shanahan, M. Training a spiking neural network to control a 4-DoF robotic arm based on Spike Timing-Dependent Plasticity. In Proceedings of the 2010 International Joint Conference on Neural Networks, Barcelona, Spain, 18–23 July 2010; pp. 1–8. [Google Scholar]
- Shuzhi, S.G.; Hang, C.C.; Woon, L.C. Adaptive neural network control of robot manipulators in task space. IEEE Trans. Ind. Electron. 1997, 44, 746–752. [Google Scholar] [CrossRef] [Green Version]
- Almusawi, A.R.J.; Dülger, L.; Sadettin, K. A new artificial neural network approach in solving inverse kinematics of robotic arm (Denso VP6242). Comput. Intell. Neurosci. 2016, 2016, 5720163. [Google Scholar] [CrossRef] [Green Version]
- Ligutan, D.D.; Abad, A.C.; Dadios, E.P. Adaptive Robotic Arm Control using Artificial Neural Network. In Proceedings of the 2018 IEEE 10th International Conference on Humanoid, Nanotechnology, Information Technology, Communication and Control, Environment and Management, Baguio City, Philippines, 29 November–2 December 2018. [Google Scholar]
- Tieck, J.C.V.; Steffen, L.; Kaiser, J.; Roennau, A.; Dillmann, R. Controlling a robot arm for target reaching without planning using spiking neurons. In Proceedings of the 2018 IEEE 17th International Conference on Cognitive Informatics & Cognitive Computing, Berkeley, CA, USA, 16–18 July 2018. [Google Scholar]
- Yang, J.; Na, J.; Gao, G.; Zhang, C. Adaptive neural tracking control of robotic manipulators with guaranteed nn weight convergence. Complexity 2018, 2018, 7131562. [Google Scholar] [CrossRef]
- Chadderdon, G.; Neymotin, S.; Kerr, C.; Lytton, W. Reinforcement learning of targeted movement in a spiking neuronal model of motor cortex. PLoS ONE 2012, 7, e47251. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.S.; Bucci, L.D.; Krichmar, J.L. Learning touch preferences with a tactile robot using dopamine modulated STDP in a model of insular cortex. Front. Neurorobot. 2015, 9, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tieck, J.C.V.; Becker, P.; Kaiser, J.; Peric, I.; Akl, M.; Reichard, D.; Roennau, A.; Dillmann, R. Learning Target Reaching Motions with a Robotic Arm Using Brain-Inspired Dopamine Modulated STDP. In Proceedings of the 2019 IEEE 18th International Conference on Cognitive Informatics & Cognitive Computing, Milan, Italy, 23–25 July 2019; pp. 54–61. [Google Scholar]
- Chakravarthy, S.; Joseph, D.; Surampudi, B. What do the basal ganglia do? A modeling perspective. Biol. Cybern. 2010, 103, 237–253. [Google Scholar] [CrossRef] [PubMed]
- Camilo, V.T.J.; Pogančić, M.V.; Kaiser, J.; Roennau, A.; Gewaltig, M.-O.; Dillmann, R. Learning Continuous Muscle Control for a Multi-joint Arm by Extending Proximal Policy Optimization with a Liquid State Machine. In Lecture Notes in Computer Science; Springer: Cham, Switzerland, 2018; Volume 11139. [Google Scholar]
- Nichols, E.; McDaid, L.J.; Siddique, N. Biologically Inspired SNN for Robot Control. IEEE Trans. Cybern. 2013, 43, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Clawson, T.S.; Ferrari, S.; Fuller, S.B.; Wood, R.J. Spiking neural network (SNN) control of a flapping insect-scale robot. In Proceedings of the 2016 IEEE 55th Conference on Decision and Control, Las Vegas, NV, USA, 12–14 December 2016; pp. 3381–3388. [Google Scholar]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hulea, M.; Uleru, G.I.; Caruntu, C.F. Adaptive SNN for Anthropomorphic Finger Control. Sensors 2021, 21, 2730. https://doi.org/10.3390/s21082730
Hulea M, Uleru GI, Caruntu CF. Adaptive SNN for Anthropomorphic Finger Control. Sensors. 2021; 21(8):2730. https://doi.org/10.3390/s21082730
Chicago/Turabian StyleHulea, Mircea, George Iulian Uleru, and Constantin Florin Caruntu. 2021. "Adaptive SNN for Anthropomorphic Finger Control" Sensors 21, no. 8: 2730. https://doi.org/10.3390/s21082730
APA StyleHulea, M., Uleru, G. I., & Caruntu, C. F. (2021). Adaptive SNN for Anthropomorphic Finger Control. Sensors, 21(8), 2730. https://doi.org/10.3390/s21082730