
Assessing Walking Stability Based on Whole-Body Movement Derived from a Depth-Sensing Camera


Abstract
:1. Introduction
2. Materials and Methods
2.1. Secondary Data Analysis
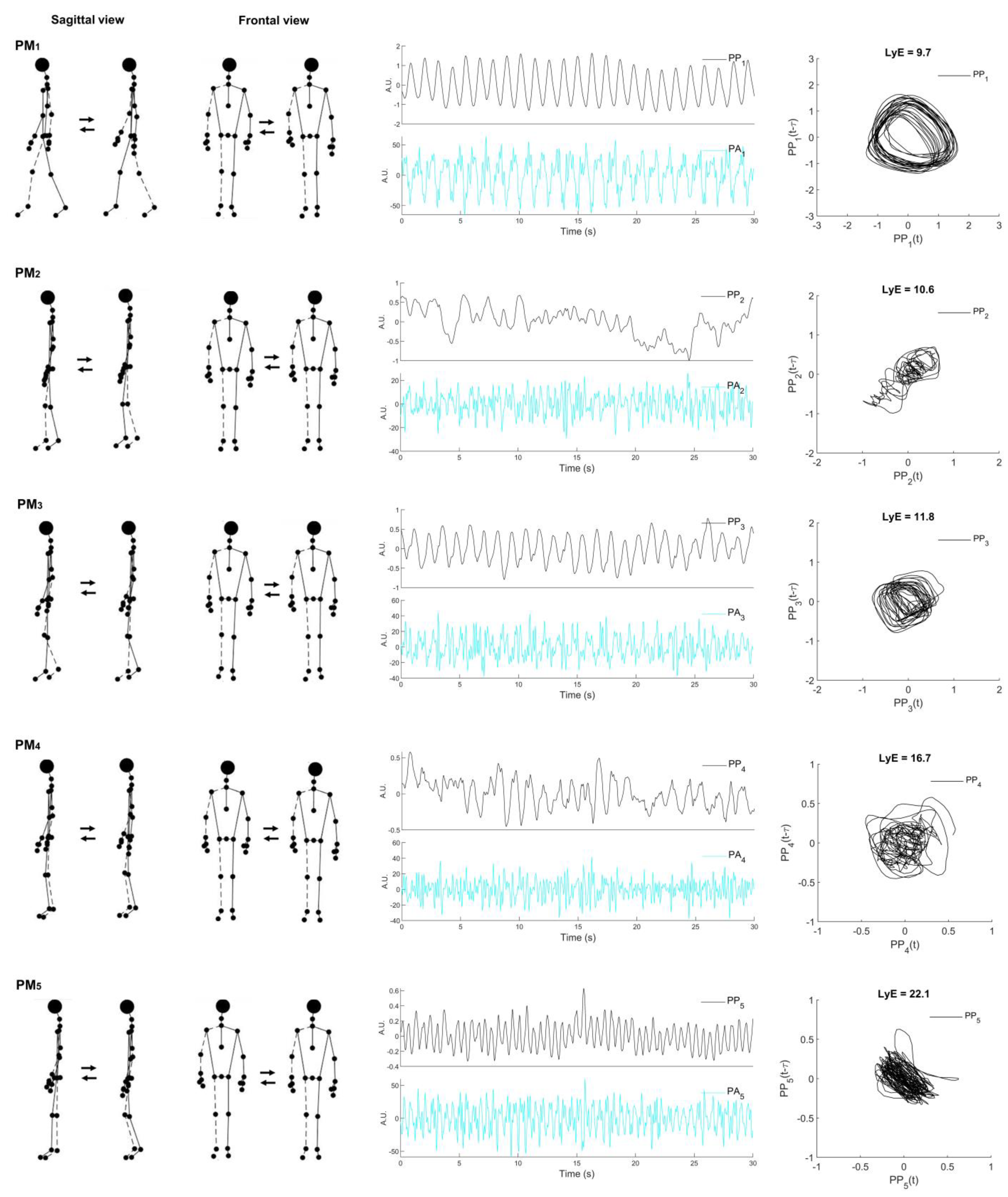
2.2. Movement Synergy Extraction
2.3. Independent Variable Computation
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Callisaya, M.L.; Blizzard, L.; Schmidt, M.D.; McGinley, J.L.; Srikanth, V.K. Ageing and Gait Variability—A Population-Based Study of Older People. Age Ageing 2010, 39, 191–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamacher, D.; Singh, N.B.; Van Dieën, J.H.; Heller, M.O.; Taylor, W.R. Kinematic Measures for Assessing Gait Stability in Elderly Individuals: A Systematic Review. J. R. Soc. Interface 2011, 8, 1682–1698. [Google Scholar] [CrossRef] [PubMed]
- Bruijn, S.M.; Meijer, O.G.; Beek, P.J.; Van Dieen, J.H.; van Dieën, J.H. Assessing the Stability of Human Locomotion: A Review of Current Measures. J. R. Soc. Interface 2013, 10, 20120999. [Google Scholar] [CrossRef] [PubMed]
- Dingwell, J.B.; Cusumano, J.P.; Cavanagh, P.R.; Sternad, D. Local Dynamic Stability Versus Kinematic Variability of Continuous Overground and Treadmill Walking. J. Biomech. Eng. 2001, 123, 27–32. [Google Scholar] [CrossRef] [Green Version]
- Dingwell, J.B.; Marin, L.C. Kinematic Variability and Local Dynamic Stability of Upper Body Motions When Walking at Different Speeds. J. Biomech. 2006, 39, 444–452. [Google Scholar] [CrossRef]
- Bernstein, N.A. The Co-Ordination and Regulation of Movements: Conclusions towards the Study of Motor Co-Ordination. Biodyn. Locomot. 1967, 104–113. [Google Scholar]
- Bizzi, E.; Cheung, V.C.K.K. The Neural Origin of Muscle Synergies. Front. Comput. Neurosci. 2013, 7, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Federolf, P.A. A Novel Approach to Study Human Posture Control: “Principal Movements” Obtained from a Principal Component Analysis of Kinematic Marker Data. J. Biomech. 2016, 49, 364–370. [Google Scholar] [CrossRef] [Green Version]
- Haid, T.H.; Zago, M.; Promsri, A.; Doix, A.-C.M.; Federolf, P.A. PManalyzer: A Software Facilitating the Study of Sensorimotor Control of Whole-Body Movements. Front. Neuroinform. 2019, 13, 24. [Google Scholar] [CrossRef] [Green Version]
- Promsri, A.; Haid, T.; Federolf, P. Complexity, Composition, and Control of Bipedal Balancing Movements as the Postural Control System Adapts to Unstable Support Surfaces or Altered Feet Positions. Neuroscience 2020, 430, 113–124. [Google Scholar] [CrossRef]
- Troje, N.F. Decomposing Biological Motion: A Framework for Analysis and Synthesis of Human Gait Patterns. J. Vis. 2002, 2, 371–387. [Google Scholar] [CrossRef] [Green Version]
- Ó’ Reilly, D. Slow Walking Synergies Reveal a Functional Role for Arm Swing Asymmetry in Healthy Adults: A Principal Component Analysis with Relation to Mechanical Work. Gait Posture 2021, 85, 126–130. [Google Scholar] [CrossRef]
- Federolf, P.; Tecante, K.; Nigg, B. A Holistic Approach to Study the Temporal Variability in Gait. J. Biomech. 2012, 45, 1127–1132. [Google Scholar] [CrossRef]
- Promsri, A.; Mohr, M.; Federolf, P. Principal Postural Acceleration and Myoelectric Activity: Interrelationship and Relevance for Characterizing Neuromuscular Function in Postural Control. Hum. Mov. Sci. 2021, 77, 102792. [Google Scholar] [CrossRef]
- Promsri, A. Sex Difference in Running Stability Analyzed Based on a Whole-Body Movement: A Pilot Study. Sport 2022, 10, 138. [Google Scholar] [CrossRef]
- Cho, S.H.; Park, J.M.; Kwon, O.Y. Gender Differences in Three Dimensional Gait Analysis Data from 98 Healthy Korean Adults. Clin. Biomech. 2004, 19, 145–152. [Google Scholar] [CrossRef]
- Bruening, D.A.; Frimenko, R.E.; Goodyear, C.D.; Bowden, D.R.; Fullenkamp, A.M. Sex Differences in Whole Body Gait Kinematics at Preferred Speeds. Gait Posture 2015, 41, 540–545. [Google Scholar] [CrossRef] [Green Version]
- McKean, K.A.; Landry, S.C.; Hubley-Kozey, C.L.; Dunbar, M.J.; Stanish, W.D.; Deluzio, K.J. Gender Differences Exist in Osteoarthritic Gait. Clin. Biomech. 2007, 22, 400–409. [Google Scholar] [CrossRef]
- Phinyomark, A.; Osis, S.T.; Hettinga, B.A.; Kobsar, D.; Ferber, R. Gender Differences in Gait Kinematics for Patients with Knee Osteoarthritis. BMC Musculoskelet. Disord. 2016, 17, 157. [Google Scholar] [CrossRef] [Green Version]
- Phinyomark, A.; Osis, S.; Hettinga, B.A.; Leigh, R.; Ferber, R. Gender Differences in Gait Kinematics in Runners with Iliotibial Band Syndrome. Scand. J. Med. Sci. Sport. 2015, 25, 744–753. [Google Scholar] [CrossRef]
- Zago, M.; Condoluci, C.; Pau, M.; Galli, M. Sex Differences in the Gait Kinematics of Patients with Down Syndrome: A Preliminary Report. J. Rehabil. Med. 2019, 51, 144–146. [Google Scholar] [CrossRef]
- Federolf, P.; Angulo-Barroso, R.M.; Busquets, A.; Ferrer-Uris, B.; Gløersen, Ø.; Mohr, M.; Ó’ Reilly, D.; Promsri, A.; van Andel, S.; Wachholz, F.; et al. Letter to the Editor Regarding “The Assessment of Center of Mass and Center of Pressure during Quiet Stance: Current Applications and Future Directions”. J. Biomech. 2021, 128, 110729. [Google Scholar] [CrossRef]
- Buckley, C.; Alcock, L.; McArdle, R.; Ur Rehman, R.Z.; Del Din, S.; Mazzà, C.; Yarnall, A.J.; Rochester, L. The Role of Movement Analysis in Diagnosing and Monitoring Neurodegenerative Conditions: Insights from Gait and Postural Control. Brain Sci. 2019, 9, 34. [Google Scholar] [CrossRef] [Green Version]
- Springer, S.; Seligmann, G.Y. Validity of the Kinect for Gait Assessment: A Focused Review. Sensors 2016, 16, 194. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.E.; Kon, S.S.C.; Canavan, J.L.; Patel, M.S.; Clark, A.L.; Nolan, C.M.; Polkey, M.I.; Man, W.D.-C. The Five-Repetition Sit-to-Stand Test as a Functional Outcome Measure in COPD. Thorax 2013, 68, 1015–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, R.A.; Pua, Y.H.; Bryant, A.L.; Hunt, M.A. Validity of the Microsoft Kinect for Providing Lateral Trunk Lean Feedback during Gait Retraining. Gait Posture 2013, 38, 1064–1066. [Google Scholar] [CrossRef]
- Cerfoglio, S.; Ferraris, C.; Vismara, L.; Amprimo, G.; Priano, L.; Pettiti, G.; Galli, M.; Mauro, A.; Cimolin, V. Kinect-Based Assessment of Lower Limbs during Gait in Post-Stroke Hemiplegic Patients: A Narrative Review. Sensors 2022, 22, 4910. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Mithraratne, K.; Wilson, N.; Zhang, Y.; Wang, X. Kinect V2-Based Gait Analysis for Children with Cerebral Palsy: Validity and Reliability of Spatial Margin of Stability and Spatiotemporal Variables. Sensors 2021, 21, 2104. [Google Scholar] [CrossRef]
- Guffanti, D.A.; Brunete, A.; Hernando, M. Kinematic Gait Data Using a Microsoft Kinect V2 Sensor during Gait Sequences over a Treadmill. IEEE Dataport 2020. [Google Scholar] [CrossRef]
- Albert, J.A.; Owolabi, V.; Gebel, A.; Brahms, C.M.; Granacher, U.; Arnrich, B. Evaluation of the Pose Tracking Performance of the Azure Kinect and Kinect v2 for Gait Analysis in Comparison with a Gold Standard: A Pilot Study. Sensors 2020, 20, 5104. [Google Scholar] [CrossRef]
- Scano, A.; Caimmi, M.; Chiavenna, A.; Malosio, M.; Tosatti, L.M. A Kinect-Based Biomechanical Assessment of Neurological Patients’ Motor Performances for Domestic Rehabilitation; IGI Global: Hershey, PA, USA, 2016; pp. 252–279. Available online: https://services.igi-global.com/resolvedoi/resolve.aspx?doi=10.4018/978-1-4666-9740-9.ch013 (accessed on 26 February 2022).
- Promsri, A.; Longo, A.; Haid, T.; Doix, A.-C.M.; Federolf, P. Leg Dominance as a Risk Factor for Lower-Limb Injuries in Downhill Skiers—A Pilot Study into Possible Mechanisms. Int. J. Environ. Res. Public Health 2019, 16, 3399. [Google Scholar] [CrossRef]
- Promsri, A.; Haid, T.; Werner, I.; Federolf, P. Leg Dominance Effects on Postural Control When Performing Challenging Balance Exercises. Brain Sci. 2020, 10, 128. [Google Scholar] [CrossRef] [Green Version]
- Promsri, A.; Federolf, P. Analysis of Postural Control Using Principal Component Analysis: The Relevance of Postural Accelerations and of Their Frequency Dependency for Selecting the Number of Movement Components. Front. Bioeng. Biotechnol. 2020, 8, 480. [Google Scholar] [CrossRef]
- Promsri, A.; Haid, T.; Federolf, P. How Does Lower Limb Dominance Influence Postural Control Movements during Single Leg Stance? Hum. Mov. Sci. 2018, 58, 165–174. [Google Scholar] [CrossRef]
- De Leva, P. Adjustments to Zatsiorsky-Seluyanov’s Segment Inertia Parameters. J. Biomech. 1996, 29, 1223–1230. [Google Scholar] [CrossRef]
- Promsri, A.; Haid, T.; Werner, I.; Federolf, P. Influence of Lower-Limb Dominance on Coordinative Movement Structures Observed during Single-Leg Balancing on a Multiaxial Unstable Surface. Gait Posture 2018, 65, 60–61. [Google Scholar] [CrossRef]
- Wachholz, F.; Kockum, T.; Haid, T.; Federolf, P. Changed Temporal Structure of Neuromuscular Control, Rather than Changed Intersegment Coordination, Explains Altered Stabilographic Regularity after a Moderate Perturbation of the Postural Control System. Entropy 2019, 21, 614. [Google Scholar] [CrossRef] [Green Version]
- Haid, T.H.; Doix, A.-C.M.; Nigg, B.M.; Federolf, P.A. Age Effects in Postural Control Analyzed via a Principal Component Analysis of Kinematic Data and Interpreted in Relation to Predictions of the Optimal Feedback Control Theory. Front. Aging Neurosci. 2018, 10, 22. [Google Scholar] [CrossRef] [Green Version]
- Wachholz, F.; Tiribello, F.; Promsri, A.; Federolf, P. Should the Minimal Intervention Principle Be Considered When Investigating Dual-Tasking Effects on Postural Control? Brain Sci. 2019, 10, 1. [Google Scholar] [CrossRef] [Green Version]
- Ó’Reilly, D.; Federolf, P. Identifying Differences in Gait Adaptability across Various Speeds Using Movement Synergy Analysis. PLoS ONE 2021, 16, e0244582. [Google Scholar] [CrossRef]
- Wolf, A.; Swift, J.B.; Swinney, H.L.; Vastano, J.A. Determining Lyapunov Exponents from a Time Series. Phys. D Nonlinear Phenom. 1985, 16, 285–317. [Google Scholar] [CrossRef]
- Longo, A.; Haid, T.; Meulenbroek, R.; Federolf, P. Biomechanics in Posture Space: Properties and Relevance of Principal Accelerations for Characterizing Movement Control. J. Biomech. 2019, 82, 397–403. [Google Scholar] [CrossRef]
- Fraser, A.M.; Swinney, H.L. Independent Coordinates for Strange Attractors from Mutual Information. Phys. Rev. A 1986, 33, 1134–1140. [Google Scholar] [CrossRef]
- Kantz, H. A Robust Method to Estimate the Maximal Lyapunov Exponent of a Time Series. Phys. Lett. A 1994, 185, 77–87. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences, 2nd ed.; Cox, B.J., Endler, N.S., Swinson, R.P., Eds.; Erlbaum: Hillsdale, NJ, USA, 1998. [Google Scholar]
- Rosenthal, R.; Rubin, D.B. R (Equivalent): A Simple Effect Size Indicator. Psychol. Methods 2003, 8, 492–496. [Google Scholar] [CrossRef] [Green Version]
- Perry, J.; Burnfield, J.M.; Cabico, L.M. Gait Analysis: Normal and Pathological Function; SLACK: Thorofare, NJ, USA, 2010; p. 551. [Google Scholar]
- Hewett, T.E. Neuromuscular and Hormonal Factors Associated with Knee Injuries in Female Athletes: Strategies for Intervention. Sport. Med. 2000, 29, 313–327. [Google Scholar] [CrossRef]


{kind=link}
| Male (n = 50) | Female (n = 54) | p-Value | |
|---|---|---|---|
| Age (years) | 22.4 ± 3.4 | 21.3 ± 3.5 | 0.124 |
| Weight (kg) | 71.6 ± 9.2 | 61.6 ± 10.0 | <0.001 * |
| Height (cm) | 177.6 ± 6.3 | 165.3 ± 6.1 | <0.001 * |
| Body mass index (kg/m2) | 22.7 ± 2.6 | 22.5 ± 3.2 | 0.091 |
| PMk | PPk_rVAR (%) | Descriptive Movements |
|---|---|---|
| 1 | 53.3 ± 9.5 | The swing phase: anti-phase arm and leg movements in the sagittal plane |
| 2 | 19.7 ± 9.0 | The single-limb support phase closely related to the terminal stance phase: anti-phase hip flexion and extension movements |
| 3 | 9.9 ± 2.6 | The single-limb support phase closely related to the mid-stance phase: anti-phase knee flexion and extension movements combined with a lateral shift of the upper body onto the stance leg |
| 4 | 6.7 ± 3.1 | Weight acceptance of the stance phase: lateral weight shift with small knee flexion posture |
| 5 | 2.3 ± 1.1 | Weight acceptance of the stance phase: knee flexion/extension movements and vertical whole-body movements combined with the sliding of the treadmill |
| PPk_rVAR | Male | Female | p-Value | Effect Size | Observed Power |
| 1 | 55.0 ± 9.4 | 51.8 ± 9.4 | 0.064 | −0.371 | 0.753 |
| 2 | 18.8 ± 7.9 | 20.6 ± 9.8 | 0.359 | −0.183 | 0.582 |
| 3 | 10.4 ± 2.6 | 9.3 ± 2.3 | 0.020 * | −0.465 | 0.827 |
| 4 | 6.4 ± 3.2 | 7.0 ± 2.9 | 0.079 | −0.351 | 0.736 |
| 5 | 2.1 ± 0.5 | 2.5 ± 1.4 | 0.080 | −0.350 | 0.735 |
| PPk_LyE | Male | Female | p-Value | Effect size | Observed power |
| 1 | 9.1 ± 1.7 | 10.0 ± 2.0 | 0.013 * | 0.484 | 0.847 |
| 2 | 9.8 ± 2.1 | 9.6 ± 2.7 | 0.348 | 0.082 | 0.518 |
| 3 | 10.8 ± 2.2 | 11.5 ± 1.8 | 0.086 | 0.367 | 0.757 |
| 4 | 13.9 ± 2.2 | 14.0 ± 2.3 | 0.393 | 0.133 | 0.547 |
| 5 | 17.5 ± 1.8 | 18.1 ± 1.5 | 0.089 | 0.362 | 0.753 |
| PAk_N | Male | Female | p-Value | Effect size | Observed power |
| 1 | 124.4 ± 34.9 | 140.0 ± 30.9 | 0.017 * | 0.483 | 0.846 |
| 2 | 155.4 ± 22.7 | 156.6 ± 26.4 | 0.810 | 0.079 | 0.517 |
| 3 | 167.6 ± 32.0 | 183.1 ± 25.9 | 0.008 * | 0.548 | 0.884 |
| 4 | 193.7 ± 24.9 | 202.9 ± 27.1 | 0.075 | 0.345 | 0.737 |
| 5 | 182.3 ± 22.5 | 187.6 ± 24.8 | 0.260 | 0.212 | 0.611 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Promsri, A. Assessing Walking Stability Based on Whole-Body Movement Derived from a Depth-Sensing Camera. Sensors 2022, 22, 7542. https://doi.org/10.3390/s22197542
Promsri A. Assessing Walking Stability Based on Whole-Body Movement Derived from a Depth-Sensing Camera. Sensors. 2022; 22(19):7542. https://doi.org/10.3390/s22197542
Chicago/Turabian StylePromsri, Arunee. 2022. "Assessing Walking Stability Based on Whole-Body Movement Derived from a Depth-Sensing Camera" Sensors 22, no. 19: 7542. https://doi.org/10.3390/s22197542
APA StylePromsri, A. (2022). Assessing Walking Stability Based on Whole-Body Movement Derived from a Depth-Sensing Camera. Sensors, 22(19), 7542. https://doi.org/10.3390/s22197542